

Abstract
Δ-6-fatty acid desaturase (FADS2) is the key enzyme in the biosynthesis of polyunsaturated fatty acids (PUFAs), the essential structural determinants of mammalian membrane lipid-bilayers. We developed the auxotrophic fads2−/− mouse mutant to assess the enigmatic role of ω3- and ω6-PUFAs in lipid homeostasis, membrane structure and function. Obesity resistance is another major phenotype of the fads2−/− mutant, the molecular basis of which is unknown. Phospholipidomic profiling of membrane systems of fads2−/−mice revealed diacylglycerol-structures, deprived of PUFAs but substituted with surrogate eicosa-5,11,14-trienoic acid. ω6-Arachidonic (AA) and ω3-docosahexaenoic acid (DHA) supplemented diets transformed fads2−/− into AA-fads2−/− and DHA-fads2−/− mutants. Severely altered phospholipid-bilayer structures of subcellular membranes of fads2−/− liver specifically interfered with maturation of transcription factor sterol-regulatory-element-binding protein, the key regulator of lipogenesis and lipid homeostasis. This study strengthens the concept that specific PUFA-substituted membrane phospholipid species are critical constituents of the structural platform operative in lipid homeostasis in normal and disease conditions.
Keywords: AA- and DHA-fads2−/− mouse mutants, abnormal DAG-structures of PL, deregulated lipogenesis, FADS2-deficiency, hepatic steatosis
Introduction
The role of ω3- and ω6- PUFAs in lipid homeostasis and related imbalances in several pathologies have been the subject of intense research for several decades 1. Evidence accumulating during the last decades suggests that the genetic basis of lipid homeostasis is critically regulated by epigenetic factors, particularly nutritional imbalances. The ω3/ω6-PUFA ratio in the Western diet is regarded as the driving force in the development of chronic metabolic, vascular 2 3 4 5, psychiatric and neurodegenerative disorders 6.
Dietary studies have been the approach to better define the systemic role of ω3- and ω6-PUFAs in lipid homeostasis and the molecular pathogenesis of these diseases. Subsequently, a broad range of therapeutic implementations have been proposed. However, until the precise roles of ω3- and ω6-PUFAs are better understood, treatment of PUFA-related diseases may not be adequate.
Mammalian cells transform essential fatty acids (EFAs) ω3-α-linolenic (α-18:3; ALA) and ω6-linoleic (18:2; LA) acid at fatty acid desaturase- and elongase- complexes, localized in the endoplasmic reticulum (ER), into long-chain ω3- and ω6-PUFA families, that include ω3-eicosapentaenoic (20:55,8,11,14,17, EPA), ω3-docosahexaenoic acid(22:64,7,10,13,16,19, DHA), ω6-di-homo-γ-linolenic (20:38,11,14), and ω6-arachidonic acid (20:45,8,11,14, AA). PUFAs are incorporated into the diacylglycerol (DAG)-backbone of respective phospholipids (PLs) by specific acyl-transferases during de novo synthesis 7 or by the deacylation-reacylation cycle 8. PLs are amphipathic molecules with ‘dove tail’ functions, asymmetrically distributed in the outer leaflet [phosphatidyl-choline (PC) and sphingomyelin (SM)] and inner leaflet [phosphatidyl -ethanolamine, PE, -inositol, PI, -serine, PS and bis-phosphatidyl-glycerol, CL (cardiolipin)]. PUFAs are structural determinants within the hydrophobic DAG-core, where they contribute to the hydrophobic scaffold of functionally divergent integral membrane proteins. Their polar head groups at the membrane surface function as docking sites of protein domains in cellular transport and cell signaling.
We applied the loss of structure-function strategy to studies in vivo on the molecular mechanism(s) underlying the structural and metabolic role of individual PUFAs. This approach has become feasible in the fads2−/−mouse mutant 9. Systemic absence of PUFAs and auxotrophy of the fads2−/−mouse mutant are essential prerequisites for conclusive dietary studies in vivo.
Infertility of male and female fads2−/− mice was the first dominant phenotype analyzed 9. The rescue of male and female fertility by a 22:6- (DHA) supplemented diet gave the first hint at the auxotrophy of the fads2−/− mutant. Incorporation of specifically DHA into PLs of Sertoli and follicular granulosa cells rescued cell polarity, germ cell maturation and fertility. These results have been confirmed in another fads2−/− mouse model 10 11.
We report here on another major phenotype of the fads2−/− mutant, perturbed lipogenesis and obesity resistance, and the molecular link to PUFA deficiency.
Profiling of the membrane phospholipidomes of the metabolically most active tissues, liver, muscle, brown adipose tissue (BAT) and white adipose tissue (WAT) generated the structural platform for the phenotypic characterization.
The first striking observation was the activation of a novel abnormal pathway, by which LA acid was transformed into the eicosatrienoic acid ω6-20:35,11,14 with a non canonical double-bond system. Second, 20:35,11,14 was systemically incorporated as a surrogate of PUFAs, specifically of AA (ω6-20:45,8,11,14), into DAGs of all membrane PLs. Third, the severely perturbed core structures of subcellular membranes (ER-, Golgi- and nuclear) critically altered the hydrophobic environment for functions of integral membrane proteins, specifically of ER/Golgi membrane-bound posttranslational processing of SREBP1c, a key transcription factor in the regulation of lipid metabolism. Fourth, this altered the expression of key enzymes of lipogenesis in the fads2−/− mouse.
Finally, we observed that stringent dietary supply of AA systemically transformed the auxotrophic nd-fads2−/− mutant into the ‘ω6-AA−/−’ and DHA into the ‘ω3-DHA−/−’ mouse line with solely AA and DHA, respectively. These three well defined mouse lines of fads2−/− expand the scope of studies, which target the numerous proposed but experimentally unproven functions of PUFAs in normal and disease conditions.
Results and Discussion
Fads2−/− mice show obesity resistance
FADS2-deficient mice are obesity-resistant. Adult male and female nd-fads2−/− mice are lean and 10–15% smaller in body size than nd+/+ mice (Fig1A and B). Weight gain of fads2−/− littermates slowed down after weaning and differed from fads2+/+ by 20 and 25% at age 4–5 months. AA+/+ and −/− mice gained weight at a similar but enhanced rate, but less than nd-fads2+/+ and −/− mice on DHA supplemented diet (Fig1C). Non-invasive NMR revealed a reduction of body fat mass by 10% and correspondingly an increase in lean mass of body weight of fads2−/− mice (Fig1D and E). Weight differences of liver, BAT, and WAT of nd-, AA- and DHA+/+ and −/− mice are depicted in Fig1F. Mass of subcutaneous, abdominal and epididymal adipose tissues of nd- and AA−/− mice remained markedly reduced during life span, but mass of WAT of adult (4 months) DHA −/− mice was twice that of DHA+/+ mice. Size of adipocytes were estimated from area measurements in sections of epididymal WAT.
Figure 1. Fads2−/− mice develop obesity resistance.
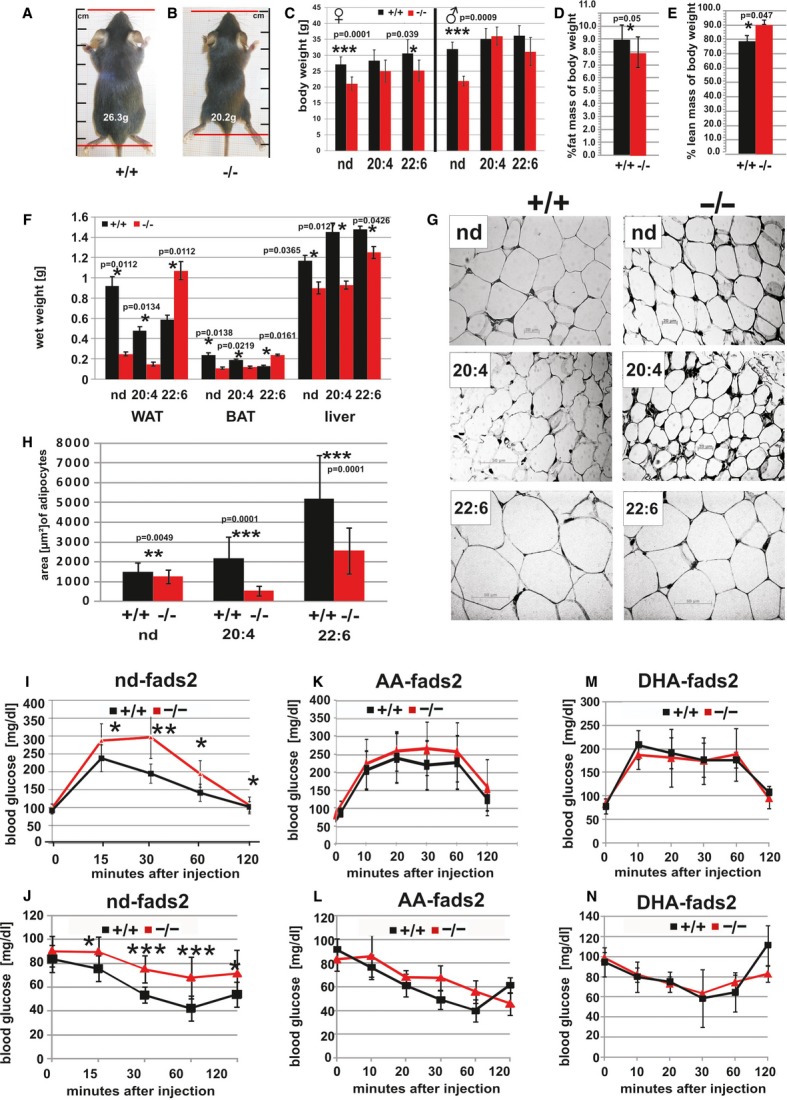
- A–C Body size [cm] (nose-tail root) and weight of male and female fads2−/− mice (age 4 months) differed by about 20 and 25% from control +/+ mice (A, B), similarly of female DHA−/−, unlike those of AA−/− (n = 20) (C).
- D, E NMR-spectroscopy revealed significantly reduced% fat mass (D) and correspondently elevated% lean mass of body weight in fads2−/− (n = 10) (E).
- F Wet weight [g] of WAT of fads2+/+ and AA+/+ (black bars) was four and three times and of BAT about twice higher than that of corresponding fads2−/− mice. Weight of WAT and BAT of DHA−/− exceeded twice that of DHA+/+ mice (n = 10). Liver weight of nd-, AA- and DHA+/+ mice was 1.3, 1.5 and 1.2 times higher than that of corresponding fads2−/− mice.
- G, H In HE-stained sections (5 μm) epididymal adipocytes of nd+/+ were significantly larger than epididymal adipocytes of nd−/− mice, about three times in AA- and twice in DHA+/+ mice.
- I Glucose tolerance test (GTT) (15 min P = 0.0342, 30 min P = 0.0019, 60 min P = 0.0130, 120 min P = 0.0271) of nd-fads2+/+ and −/− mice (red).
- J Insulin tolerance test (ITT) (15 min P = 0.030, 30 min P = 0.0001, 60 min P = 0.0005, 120 min P = 0.030) of nd-fads2+/+ and −/− mice (red).
- K Normal GTT of AA+/+ and AA−/− mice (n = 10).
- L Normal ITT of AA+/+ and AA−/− mice (n = 10).
- M GTT of DHA+/+ and DHA−/− mice (n = 10).
- N ITT of DHA+/+ and DHA−/− mice (n = 10).
Adipocytes of nd−/− mice were about two-third in size of nd +/+ mice, about one third in AA−/− and half in DHA−/− mice (FigG and H). Situs of age- and gender-matched nd-, AA- and DHA+/+ and −/− littermates underlined these parameters (supplementary Fig S1 A–F).
The loss of adipose tissue of nd−/− mice was associated with reduced levels of serum leptin, 1.8 ± 0.43 ng/ml in nd+/+ to 0.8 ± 0.4 ng/ml in nd−/− mice. Leptin levels were similar in AA+/+ and AA−/− mice, 1.25 ± 0.4 and 1.2 ± 0.3 ng/ml, respectively, and those of DHA +/+ mice were elevated (6 ± 2 ng/ml), but normal in DHA−/− mice (1.2 ± 0.4 ng/ml) (n = 10). nd fads2−/− mice showed significantly reduced serum triglyceride and cholesterol concentrations (supplementary Fig S1G and H).
Leanness of nd-fads2−/− was not caused by reduced food uptake, malabsorption or maldigestion. Phenomaster recordings documented a similar food uptake of nd- and AA+/+ and −/− mice (supplementary Fig S1I). DHA+/+ and DHA−/− mice, however, were hyperphagic (supplementary Fig S1J). Metabolic parameters (VO2 uptake, VCO2 production, and heat dissipation) of nd−/− mice were reduced by approximately one third compared to control mice (supplementary Fig S2C–E), but comparable and unchanged in AA- or DHA+/+ and −/− mice. RER reflected a balanced carbohydrate and fat utilization by the three cohorts (supplementary Fig S2).
Enhanced energy production due to perturbed epidermal lipid barrier with trans-epidermal water loss (TEWL) and associated metabolic waste syndrome were excluded by measurement of TEWL of the nd-fads2−/−mutant (supplementary Fig S2A).
Locomotor activity of adult (3–5 months) nd-, AA- and DHA+/+ and −/− mice was compared in the Phenomaster (supplementary Fig S2B). Locomotor activity of nd- and DHA−/− mice was reduced to one third due to the absence of the two main anandamides (N-arachidonoyl-ethanolamide and 2-arachidonoyl-glycerol) in brain extracts of respective cohorts of nd−/− and DHA−/− mice, but AA−/− mice showed normal motility (data not shown). We next searched for biochemical and cellular links between altered lipid metabolism and strongly reduced adipogenesis in the fads2−/− mouse.
Insulin response in the fads2−/− mouse is altered
Fasting serum insulin concentration of fads2−/− mice is about half that of +/+ littermates, whereas in AA- and DHA−/− mice have normal serum insulin concentrations. The glucose tolerance test (GTT) of nd+/+ and −/− mice (Fig1I) showed a significantly retarded return to normal serum glucose concentration in the nd−/− mouse during the 2 h period post intraperitoneal insulin application, suggesting reduced insulin sensitivity, which is also reflected in the insulin tolerance test (ITT) (Fig1J). GTTs and ITTs of AA- and DHA+/+ and −/− mice showed a similar responses (Fig1K–N).
Severe perturbationof the nd-fads2−/−membrane phospholipidome
PUFAs are ubiquitous substituents of the DAG moiety of all PL classes of mammalian tissues. To unravel the mechanisms that link PUFA deficiency and deregulated lipogenesis in the nd−/− mouse, we defined the structural platform by a comprehensive analysis of the phospolipidome of the four metabolically most active tissues liver, muscle, BAT and WAT. PL classes were separated by high-performance thin-layer chromatography (HPTLC) for MS/MS using precursor scanning characteristic for each PL class, depicted for fads2+/+ and −/− liver. The flow diagram in supplementary Fig S3A paradigmatically outlines the separation, isolation, and characterization of individual species of PI of total lipid extract of liver of fads2+/+ and −/− mice by MS/MS. Further detailed structural insight into DAG species of each PL class was obtained by GC/MS analysis of their FA substituents as methylesters.
The first surprising discovery was the induction of a futile biosynthetic pathway, by which LA was transformed into a novel non-mammalian all-cis-eicosa-5,11,14-trienoic acid (20:35,11,14) for substitution of PL-associated DAGs in the fads2−/−mouse (Fig2A and B). 20:35,11,14 was systemically incorporated as a surrogate of AA and PUFAs in the main PL classes PC, PI, PS and PE, not in cardiolipin (Fig2C and D).
Figure 2. Linoleic acid is transformed in a non-canonical reaction sequence to the non-mammalian ‘FADS2-deficiency’ eicosa-5z,11z,14z-trienoic acid (20:35,11,14).
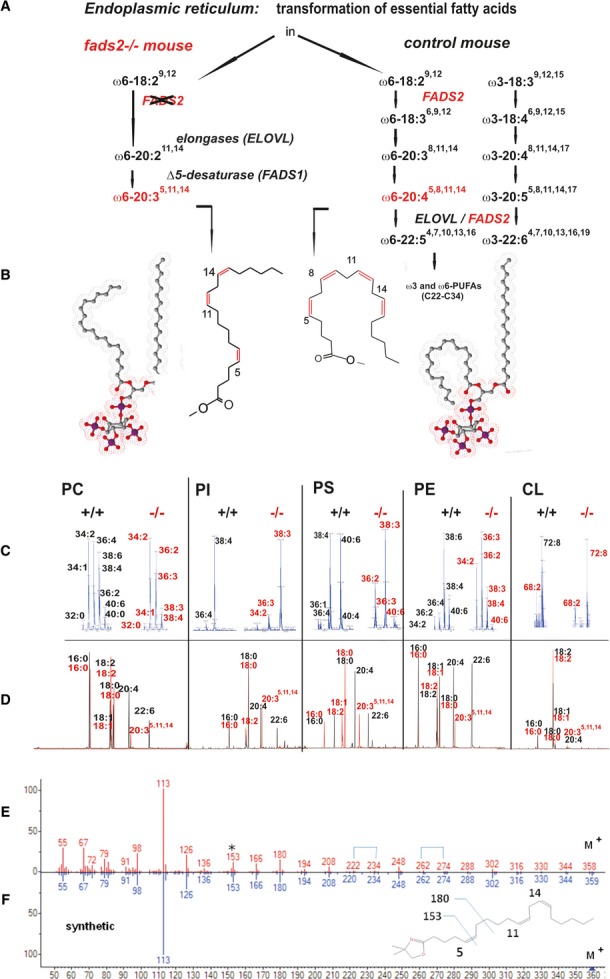
- A Pathway of the transformation of linoleic acid, canonical (right) and aberrant to 20:35,11,14 (left) in the fads2−/− mouse.
- B Space filling models of 18:0/20:45,8,11,14-PIP3 and 18:0/20:35,11,14-PIP3.
- C MS/MS of PC, PI, PS, PE, and cardiolipin species of total lipid extract. Numbers at peaks represent the sum of C-atoms and number of double bonds in acyl-chains of DAGs in respective PL species.
- D GC/MS of FAME of substituents of DAGs of individual PL species. Black bars, +/+; red bars, −/−.
- E, F Structural characterization (head-to-tail presentation) by GC/MS of 2,2-dimethyloxazoline (DMOX) derivatives of 20:35,11,14, of unknown C20-PUFA of liver PI as 20:35,11,14 (E) and of synthetic 20:35,11,14(F).
Figure 3. Phospholipidome of total liver unveils the absence of all PUFAs and their substitution by surrogate 20:35,11,14.
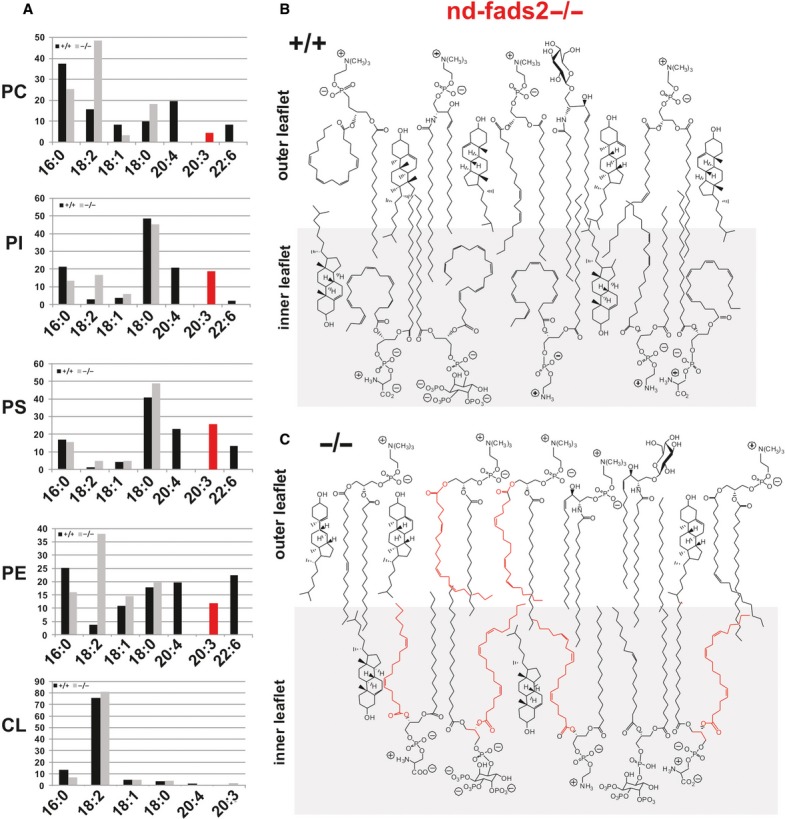
- A Profiling of the phospholipidome of total liver of fads2+/+ (black) and −/− (red) mice (age 4 months). GC/MS of FAME of substituents of DAGs of individual PC, PI, PS, PE, and cardiolipin species. Black bars, +/+; grey bars, −/−. Aberrant 20:35,11,14 (red bar) is the only PUFA in PLs of liver. y-axis,% of total FAME.
- B, C Schematic presentation of the assembly of PL species in a domain of the asymmetric bilayer of membranes of control +/+ (B) and fads2−/−mice (C), and highlights the severely distorted inner leaflet of the bilayer.
Figure 4. Dietary arachidonic (AA) and docosahexaenoic (DHA) acid transform the auxotrophic nd-fads2−/− mouse into the ‘AA-’ and ‘DHA-fads2−/−’ mouse mutants.
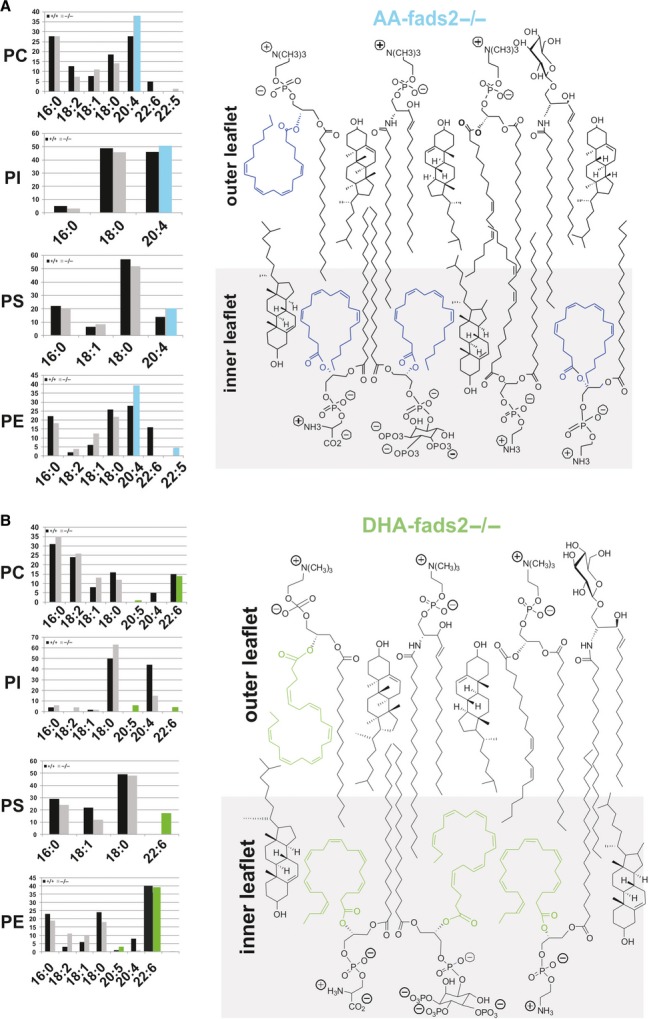
- A AA and DHA supplemented diets systemically suppress the synthesis of 20:35,11,14. The bar diagrams present the quantitative analysis of the fatty acid substituents of the main PL classes of liver of AA−/− mice. AA (blue) are the only surrogates for PUFAs in all PL species. y-axis:% of total FAME. Right panel: schematic structural view of the putative assembly of AA-substituted PL classes enriched in the inner leaflet of the PL bilayer in a domain of the asymmetric membrane bilayer.
- B Same as in (A) for DHA−/− mice. DHA (green) are the only surrogates for PUFAs in all PL species. y-axis:% of total FAME. Right panel: schematic structural view of the putative assembly of DHA-substituted PL classes enriched in the inner leaflet of the PL bilayer in a domain of the asymmetric membrane bilayer.
The structure of this unusual PL constituent was established unambiguously by GC/MS analysis of its DMOX (2,2-dimethyl oxazoline) derivative. DMOX derivatives of PUFAs release diagnostic fragments in MS, characteristic of the methylen-interrupted double-bond systems 12 13 14 15.The analytical data were identical with those of synthetic DMOX-20:35,11,14 16 and spectra from published data 13 (FigE and F). The double-bond positions of 20:35,11,14 were confirmed finally by oxidative ozonolysis, which released the expected adipic and glutaric acids, identified as dimethylesters by GC/MS, (supplementary Fig S3).
The analytical data suggested the activation of a novel futile pathway from LA via chain elongation to 20:211,14 and Δ5-desaturation to 20:35,11,14 in the nd−/− mouse.
We next studied the distribution of 20:35,11,14 in DAGs of PC, PI, PS, and PE in the phospholipidome of liver total lipid extract of nd-fads2−/− mice and their microsomal and nuclear membrane fractions, muscle, BAT, and WAT (supplementary Fig S4), the metabolically most active tissues in the nd−/− mouse. Stoichiometry and distribution of 20:35,11,14 in different PL classes in nd−/− mice were close to that of respective 20:45,8,11,14 in fads2+/+mice (Fig S3). No Δ5-desaturation of 18:29,12 to 18:35,9,12 or α-18:39,12,15 to 18:45,9,12,15 was observed. These findings underline the high chain-length specificity of Δ5-desaturase (FADS1).
Transforming the nd-fads2−/− into the ‘arachidonic acid (AA)-’ and ‘docosahexaenoic acid (DHA)-fads2−/−’ mouse mutants
We asked whether 20:35,11,14 substituted PLs in mammalian membrane systems can match the structural and functional properties of AA and DHA. Unexpectedly, nd−/− mice, when raised on 20:4- (AA) and 22:6- (DHA) supplemented diets, completely suppressed the synthesis of 20:35,11,14. 20:35,11,14 was systemically replaced by 20:4 and 22:6 in respective PL classes of AA- and DHA−/− mice, as depicted for total liver and its purified nuclear and ER- membranes, and of muscle, BAT, and WAT (supplementary Fig S4).
These in vivo studies were confirmed by experiments in primary hepatocyte cultures from nd+/+ and auxotrophic nd−/− mice. Supplementary Fig S5 depicts the utilization of 20:45,8,11,14 (AA) and ω6-20:35,11,14 in the synthesis of PL classes in +/+ and −/− primary hepatocytes. sn2-O-Acyltransferases of the family of membrane-bound O-acyltransferases (MBOAT) 17 showed no preference for either substrate in de novo synthesis or PL remodelling.
In conclusion, these dietary studies generated three well-defined fads2−/− mouse models: (i) the nd−/− mouse, all PL classes of which lack the canonical ω3- and ω6-PUFAs but instead are acylated exclusively with 20:35,11,14 as PUFA surrogate, (ii) the ‘ω6-AA-fads2−/−’ mouse with ω6-arachidonic acid, and (iii) the ‘ω3-DHA-fads2−/−’ mouse with ω3-docosahexaenoic acid as the only PUFA as substituent in the sn2-position of DAGs of membrane PLs in the membrane lipid bilayer. The AA−/− mouse resembles a mimicry of mammalian solely on the ‘land food web’ and the DHA−/− of the ‘sea food chain’ during evolution 1.
Gene expression of enzymes of lipid metabolism in fads2−/− mice
We next investigated the expression profiles of genes coding for transcription factors of enzymes involved in lipid metabolism pparα, ß, γ, srebp1c, hmgcoas2, hnf1α, hnf4α and enzymes of fatty acid metabolism cpt1 and 2, scd1-3, Δ5 (fads1) and elongases 2–6 and of gluconeogenesis fruct-1,6-bp, glc-6p and pepck of (i) the nd-fads2 (Fig5A and B), (ii) the AA- (Fig5C and D) and (iii) the DHA+/+ and −/− mice (Fig 5E and F) by real time qRT-PCR. (i) Gene expression of enzymes involved in gluconeogenesis did not change significantly in nd +/+ and −/− liver, except for the upregulation of glc-6p in DHA−/− mouse liver (Fig5A and B). Also expression of transcription factors of lipogenesis, including pparα, ß, γ, hnf1α and hnf4α, and srebp1c, and enzymes of cholesterol synthesis (HMGCoA synthase) were not significantly changed in nd−/− liver. However, steady-state concentrations of the RNA of key enzymes of PUFA synthesis, the SCD1-3 and desaturase Δ5 (FADS1) subunit of the trimeric microsomal desaturase complexes, and the condensing subunits of the tetrameric elongase complexes ELOVL2, 5 and 6 were remarkably elevated. ELOVL2 in nd-, AA- and DHA−/− liver is essential for the synthesis of ω6-very long chain PUFAs (VLCFA) 18. 18:2 and γ-18:3 are the preferred substrates of ELOVL5 19. Elovl3 expression was downregulated and thereby the synthesis of saturated VLCFA and recruitment of saturated triglycerides to BAT 20, but elovl6, which encodes the elongase of saturated C12–C16 fatty acids and also correlates positively with the severity of hepatosteatosis was overexpressed. These enzymes are major targets of SREBP1c, critically involved in the development of obesity and insulin resistance. (ii) In AA−/− liver only the expression of pparγ, the transcription factor of key enzymes of lipid metabolism, and of elovl2 was significantly upregulated (Fig5C and D). (iii) DHA−/− mouse liver showed elevated expression of glc-6p indicating enhanced gluconeogenesis, and of transcription factor srebp1c causing the activation of Δ5 desaturases (fads1) and elovl2 and elovl5 and of elovl1, the elongase yielding 24:0 to 26:0, required for ceramide synthesis. Expression of scd3, a mouse-specific isoform of stearoyl-CoA desaturase, was down regulated in DHA −/− mice (Fig5E and F).
Figure 5. Lipogenesis in the fads2−/− mutant is regulated by the membrane PL environment containing specific PUFA subtituents.

- A – F Expression profiles of genes of transcription factors of enzymes of lipid metabolism, of gluconeogenesis and FA metabolism in liver of nd- (A, B), AA- (C, D) and DHA+/+ and −/− mice (E, F). Elevated gene expression of scd1, Δ5 (fads1), elovl2, elovl5 and elovl6 in liver of nd−/− mice, of pparγ and elovl2 in AA−/−, and of sreb1c, glc-6-P, Δ5 (fads1), elovl1, elovl2 and elovl5 in liver of DHA−/− mice.
- G SREBP1c processing and nuclear import in liver of nd+/+ and nd−/− mice. Western blot analysis of ER-proteins of liver of nd-+/+ and −/− mice shows a reduced concentration of p-SREBP1c, P = 0.042 (C), but enhanced concentration of nuclear m-SREBP1c, P = 0.024. Calnexin was used as ER- and lamin A/C as nuclear protein loading marker.
- H Immuno-histochemistry of nd+/+ and −/− cryo-sections of liver were stained with anti SREBP1c antibodies, nuclei with DAPI. Inserts show magnification of nuclei with anti SREBP1c reactive antigen.
- I Oil red-stained cryo-sections (7 μm) of liver from nd+/+ showed scarce and from fads2−/− extended lipid droplets (I).
- J – O AA+/+ and −/− mice (n = 4) revealed similar concentrations of p-Srebp1c in microsomal (ER) and m-Srebp1c in nuclear protein extract (J), but significantly higher in DHA−/− ER and nuclear extracts (M). In cryosections, nuclei of AA-fads2+/+ and −/− hepatocytes were essential-free of SREBP1c antigen as demonstrated by inserts of magnified nuclei (K). Oil red staining indicated that AA+/+ and −/− hepatocytes were essentially free of lipid deposits (L), but nuclei of DHA+/+ and −/− hepatocytes loaded with SREBP1c antigen (N), massive hepatic steatosis in DHA+/+ liver, ameliorated in DHA−/−mice (O).
Deregulation of lipogenesis and hepatic steatosis in the fads2−/− mouse
Lipid homeostasis and lipid bilayer structures of the ER and Golgi membranes are stringently controlled by SREBP1c, the key transcription factor of enzymes of lipogenesis. Cholesterol concentration in the lipid bilayer of the endoplasmic reticulum and Golgi membranes regulates the membrane-bound steps of processing precursor (p)-SREBP1c to mature (m)-SREBP1c for nuclear import 21.
We therefore studied the impact of the perturbed PL bilayer of microsomal (mic) and of nuclear (nuc) membranes of nd+/+ and −/− mice on the posttranslational modification of p-SREBP1c to m-SREBP1c. Western blot analysis of ER- and nuclear membrane lysates using anti-SREBP1c antibodies indicated elevated levels of active m-SREBP1c in the nuclear fraction of nd−/− liver (Fig5G), which was confirmed by immunohistochemistry (IHC) (Fig5H). Bright field microscopy of Oil red-stained cryo sections of liver of nd−/− mice revealed hepatocytes filled with tiny lipid droplets (steatosis), compared to few larger diameter droplets in nd+/+ hepatocytes (Fig5I).
Liver of AA-fads2+/+ and AA−/− revealed a remarkably similar expression of genes of transcription factors and enzymes of lipogenesis, of except increased expression of pparγ and elovl2 (Fig5C). Western blot analysis showed a comparable processing of microsomal p-SREBP1c to nuclear m-SREBP1c (Fig5J), which is supported by IHC indicating a similar nuclear SREBP1c antigen import (Fig5K). AA−/− hepatocytes were almost free of lipid droplets (Fig5L).
Only DHA−/− liver RNA showed a threefold higher steady-state concentration of screbp1c, of glc-6p, a marker of gluconeogenesis, and of Δ5-desaturase (fads1), elongases elovl 1, 2 and 5, which are marker enzymes of PUFA metabolism (Fig5E and F). Western blot analysis indicated that microsomal p-SREBP1c- and nuclear m-SREBP1c exceeded SREBP1c levels in liver of the DHA+/+ mouse (Fig5M). IHC on liver cryo sections revealed elevated nuclear import of m-SREBP1c and enhanced fluorescent SREBP1c antigen in nuclei of DHA−/− hepatocytes. Nuclei of DHA−/− hepatocytes were heavily loaded with SREBP1c reactive peptides (Fig5N). Oil red-stained sections of DHA+/+ and −/− hepatocytes unveiled hepatic steatosis, characterized by heavy peri-nuclear macro-lipid droplet accumulation. Hepatocytes of DHA+/+ and −/− mice differed in number and size of lipid droplets. DHA+/+ liver showed heavy steatosis, which was ameliorated in hepatocytes of DHA−/− mice (Fig5O).
Conclusion
The genetically and biochemically well-defined auxotrophic fads2−/− and the derived AA- and DHA fads2−/− mutant mouse lines preclude many of the ambiguities of numerous feeding experiments in rodents in the past, which addressed the important role of PUFAs. This study indicates the wide experimental scope of the auxotrophic fads2−/− mouse. The nd-, AA- and DHA−/−mouse lines might become useful models for unveiling the complex mechanisms underlying the impact of dietary ω3- and ω6-PUFAs in membrane biology, human nutrition and the development and prevention of dyslipidemia, vascular and neurodegenerative diseases due to imbalanced dietary PUFA supply.
Materials and Methods
Reagents
The following antibodies were used for IHC and Western blot analysis: SREBP1c, α-tubulin, lamin A/C, calnexin and affinity-purified polyclonal anti-polypeptide I192-K444 specific rabbit antibodies 9. DHASCO and ARASCO (Martek Biosciences, Columbia, MD, USA) were used as sources of docosahexaenoic acid (DHA) and arachidonic acid (AA), respectively. [1-14C] Radioactive and inactive 20:35,11,14 were synthesized in this laboratory 9 16 22. Primers for qRT-PCR are listed under supplementary Table S2.
Animals
Generation and genotyping of fads2+/− and −/− mice have been described before 9. Fads2+/− mice were back crossed 10 times into the C57BL/6N genetic background. Animals were housed in the SPF-barrier mouse facility of the Center of Molecular Medicine (CMMC) with a 12 h light/dark cycle and free access to water and a regular (nd), 20:4-, and 22:6-supplemented (1% of daily caloric uptake) diet (Altromin, Dinslage Germany).The nd-diet was free of PUFAs and optimized with 18:2 and 18:3 to prohibit EFA-deficiency. Colonies of fads2+/+, +/−, and −/− mice were maintained on the respective diets throughout life time, starting with fads2+/− breeding pairs. FA composition of diets is listed under supplementary Table S1.
Laboratory measurements
Serum concentrations of glucose, insulin, leptin, triglycerides and total cholesterol were determined by standard colorimetric assays 9.
Glucose- and insulin tolerance tests
For GTT and ITT mice were fasted overnight (16 h). Glucose (2 g/kg body weight) was injected intra-peritoneally (ip). For ITT, mice were fasted for 16 h, anesthesized, and insulin (0.5 IU/kg) or saline were injected ip as control. Blood glucose and serum insulin concentrations were determined before and at different times after injection, indicated in the figure legends.
Metabolic measurements
Metabolic parameters (VO2, VCO2, H, and RER) were determined in the Phenomaster TSL-system, and fat and lean mass by the Bruker Optics minispec TD-NMRAnalyser. TEWL was measured with the Tewameter, model 210 Courage-Khazaka, Cologne, Germany,
Pulse chase experiments
Primary hepatocyte cultures for pulse chase experiments were prepared from fads2+/+ and −/− mice as previously described 23. Details are provided in Supplementary information.
Gene expression
RNA preparation and real time quantitative RT-PCR (qRT) are described in Supplementary information.
Western blot analysis
Western blot analysis of cell lysates of fads2+/+ and −/− liver, muscle, BAT and WAT and nuclear and microsomal proteins of liver is described in Supplementary information.
Immunohistochemistry
Mice were perfused from the left ventricle with PBS and PBS-buffered 4% paraformaldehyde. Organs were fixed and processed for light- and immuno fluorescence microscopy as described in Supplementary information.
Lipid analysis
Isolation, separation, identification and quantification of lipids from organs and subcellular membranes and of fatty acid constituents are described in Supplementary information.
Mass spectroscopy of phospholipids
PL classes were analyzed by MS/MS using an Applied Biosystems (Darmstadt, Germany) QTrap analyzer. Conditions are described in Supplementary information.
Oxidative ozonolysis
FA mixtures of individual PL classes were dissolved in glacial acetic acid/methylacetate 1:2 (v/v) for oxidative ozonolysis. Methylated oxidation products were analyzed by GC/MS as described in Supplementary information.
Statistical analyses
Data are expressed as means ± s.e.m. Differences between two groups were assessed using the unpaired Student's t-test. Sizes of animal cohorts are listed under respective figures.
Acknowledgments
We gratefully acknowledge the support of this work by the Center of Molecular Medicine, University of Cologne, CECAD (Cluster of Excellence, Cellular Stress Response in Aging Related Diseases), University of Cologne and the Fritz-Thyssen-Stiftung.
Author contributions
WS designed and performed experiments, BJ, EB IH and ISS performed experiments, SB, MO and MT performed data analysis, WS and IH wrote the manuscript.
Conflict of interest
The authors declare that they have no conflict of interest.
Supporting Information
Supplementary information for this article is available online: http://embor.embopress.org
Supporting Information
Review Process File
References
- Crawford MA, Broadhurst CL. The role of docosahexaenoic and the marine food web as determinants of evolution and hominid brain development: the challenge for human sustainability. Nutr Health. 2012;21:17–39. doi: 10.1177/0260106012437550. [DOI] [PubMed] [Google Scholar]
- Calder PC. Polyunsaturated fatty acids and inflammation. Prostaglandins Leukot Essent Fatty Acids. 2006;75:197–202. doi: 10.1016/j.plefa.2006.05.012. [DOI] [PubMed] [Google Scholar]
- Hwang JB, Hernandez J, Leduc R, Frost SC. Alternative glycosylation of the insulin receptor prevents oligomerization and acquisition of insulin-dependent tyrosine kinase activity. Biochim Biophys Acta. 2000;1499:74–84. doi: 10.1016/s0167-4889(00)00109-9. [DOI] [PubMed] [Google Scholar]
- Kim KY, Lee JW, Park MS, Jung MH, Jeon GA, Nam MJ. Expression of a thioredoxin-related protein-1 is induced by prostaglandin E(2) Int J Cancer. 2006;118:1670–1679. doi: 10.1002/ijc.21572. [DOI] [PubMed] [Google Scholar]
- Yaqoob P, Calder PC. N-3 polyunsaturated fatty acids and inflammation in the arterial wall. Eur J Med Res. 2003;8:337–354. [PubMed] [Google Scholar]
- Cole GM, Ma QL, Frautschy SA. Omega-3 fatty acids and dementia. Prostaglandins Leukot Essent Fatty Acids. 2009;81:213–221. doi: 10.1016/j.plefa.2009.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennedy EP. Biosynthesis of complex lipids. Fed Proc. 1961;20:934–940. [PubMed] [Google Scholar]
- Lands WE, Merkl I. Metabolism of glycerolipids. III. Reactivity of various acyl esters of coenzyme A with alpha’-acylglycerophosphorylcholine, and positional specificities in lecithin synthesis. J Biol Chem. 1963;238:898–904. [PubMed] [Google Scholar]
- Holz B, Jenke B, Binczek E, Günter RH, Kiss C, Karakesisoglou I, Thevis M, Weber AA, Arnhold S, Addicks K. Delta6-desaturase (FADS2) deficiency unveils the role of omega3- and omega6-polyunsaturated fatty acids. EMBO J. 2008;27:2281–2292. doi: 10.1038/emboj.2008.156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stroud CK, Nara TY, Roqueta-Rivera M, Radlowski CE, Lawrence P, Zhang Y, Cho BH, Segre M, Brenna JT, Haschek WM, Nakamura MT. Disruption of FADS2 gene in mice impairs male reproduction and causes dermal and intestinal ulceration. J Lipid Res. 2009;50:1870–1880. doi: 10.1194/jlr.M900039-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roqueta-Rivera M, Stroud CK, Haschek WM, Akare SJ, Segre M, Brush RS, Agbaga MP, Anderson RE, Hess RA, Nakamura MT. Docosahexaenoic acid supplementation fully restores fertility and spermatogenesis in male delta-6 desaturase-null mice. J Lipid Res. 2010;51:360–367. doi: 10.1194/jlr.M001180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christie WW. Gas chromatography-mass spectrometry methods for structural analysis of fatty acids. Lipids. 1998;33:343–353. doi: 10.1007/s11745-998-0214-x. [DOI] [PubMed] [Google Scholar]
- Christie WW, Han X. Lipid Analysis - Isolation, Separation, Identification and Lipidomic Analysis. 4th edn. Bridgwater, UK: Oily Press; 2010. p. 446. [Google Scholar]
- Kangani CO, Kelley DE, Evans RW. Synthesis and mass spectrometry of benzoxazoline, dimethyloxazoline and 4-phenyloxazoline derivatives of polyunsaturated fatty acids. Rapid Commun Mass Spectrom. 2007;21:2129–2136. doi: 10.1002/rcm.3071. [DOI] [PubMed] [Google Scholar]
- Svetashev VI. Mild method for preparation of 4,4-dimethyloxazoline derivatives of polyunsaturated fatty acids for GC-MS. Lipids. 2011;46:463–467. doi: 10.1007/s11745-011-3550-4. [DOI] [PubMed] [Google Scholar]
- Nadidai I. 1976. Pathway of arachidonic acid. Synthesis of [1-14C] 8,11,14-20:3 and [1-14C]5,11,14 20:3.
- Matsuda S, Inoue T, Lee HC, Kono N, Tanaka F, Gengyo-Ando K, Mitani S, Arai H. Member of the membrane-bound O-acyltransferase (MBOAT) family encodes a lysophospholipid acyltransferase with broad substrate specificity. Genes Cells. 2008;13:879–888. doi: 10.1111/j.1365-2443.2008.01212.x. [DOI] [PubMed] [Google Scholar]
- Zadravec D, Tvrdik P, Guillou H, Haslam R, Kobayashi T, Napier JA, Capecchi MR, Jacobsson A. ELOVL2 controls the level of n-6 28:5 and 30:5 fatty acids in testis, a prerequisite for male fertility and sperm maturation in mice. J Lipid Res. 2011;52:245–255. doi: 10.1194/jlr.M011346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leonard AE, Kelder B, Bobik EG, Chuang LT, Lewis CJ, Kopchick JJ, Mukerji P, Huang YS. Identification and expression of mammalian long-chain PUFA elongation enzymes. Lipids. 2002;37:733–740. doi: 10.1007/s11745-002-0955-6. [DOI] [PubMed] [Google Scholar]
- Westerberg R, Mansson JE, Golozoubova V, Shabalina IG, Backlund EC, Tvrdik P, Retterstol K, Capecchi MR, Jacobsson A. ELOVL3 is an important component for early onset of lipid recruitment in brown adipose tissue. J Biol Chem. 2006;281:4958–4968. doi: 10.1074/jbc.M511588200. [DOI] [PubMed] [Google Scholar]
- Wang X, Sato R, Brown MS, Hua X, Goldstein JL. SREBP-1, a membrane-bound transcription factor released by sterol-regulated proteolysis. Cell. 1994;77:53–62. doi: 10.1016/0092-8674(94)90234-8. [DOI] [PubMed] [Google Scholar]
- Stoffel W. Biosynthesis of polyenoic fatty acids. Biochem Biophys Res Commun. 1961;6:270–273. doi: 10.1016/0006-291x(61)90376-x. [DOI] [PubMed] [Google Scholar]
- Klingmuller U, Bauer A, Bohl S, Nickel PJ, Breitkopf K, Dooley S, Zellmer S, Kern C, Merfort I, Sparna T, Donaue J, Walz G, Geyer M, Kreutz C, Hermes M, Gotschel F, Hecht A, Walter D, Egger L, Neubert K. Primary mouse hepatocytes for systems biology approaches: a standardized in vitro system for modelling of signal transduction pathways. Syst Biol. 2006;153:433–447. doi: 10.1049/ip-syb:20050067. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supporting Information
Review Process File