

Abstract
Natural product Hypericum perforatum L. has been used in folk medicine to improve mental performance. However, the effect of H. perforatum L. on metabolism is still unknown. In order to test whether H. perforatum L. extract (EHP) has an effect on metabolic syndrome, we treated diet induced obese (DIO) C57BL/6J mice with the extract. The chemical characters of EHP were investigated with thin-layer chromatography, ultraviolet, high-performance liquid chromatography (HPLC), and HPLC-mass spectrometry fingerprint analysis. Oral glucose tolerance test (OGTT), insulin tolerance test (ITT), and the glucose infusion rate (GIR) in hyperinsulinemic–euglycemic clamp test were performed to evaluate the glucose metabolism and insulin sensitivity. Skeletal muscle was examined for lipid metabolism. The results suggest that EHP can significantly improve the glucose and lipid metabolism in DIO mice. In vitro, EHP inhibited the catalytic activity of recombinant human protein tyrosine phosphatase 1B (PTP1B) and reduced the protein and mRNA levels of PTP1B in the skeletal muscle. Moreover, expressions of genes related to fatty acid uptake and oxidation were changed by EHP in the skeletal muscle. These results suggest that EHP may improve insulin resistance and lipid metabolism in DIO mice. © 2014 The Authors. Phytotherapy Research published by John Wiley & Sons Ltd.
Keywords: Hypericum perforatum L., PTP1B inhibitor, insulin resistance, dyslipidemia
Introduction
Obesity and type 2 diabetes increase the numbers of death worldwide (Williams, 2008). Insulin resistance is a key factor in the etiology of type 2 diabetes and other obesity-related chronic diseases (Grundy, 2006; Ye, 2013). Lipid metabolic disorder plays an important role in insulin resistance (Novgorodtseva et al., 2011; Takasu et al., 2012). The intramyocellular lipid accumulation is associated with insulin resistance and recognized as a hallmark of type 2 diabetes in obese humans and animals (Haus et al., 2011; Korach-André et al., 2005; Kuhlmann et al., 2005; Raz et al., 2005). Treatment of insulin resistance is limited by the availability of effective and safe medicines (Ye, 2011).
Protein tyrosine phosphatase 1B (PTP1B), the negative regulator of insulin and leptin signaling pathways, has been considered as a promising molecular target in the treatment of insulin resistance and lipid metabolic disorder (Basavarajappa et al., 2012; Johnson et al., 2002; Sun et al., 2007). PTP1B inhibition leads to improvement in insulin signaling and fatty acid oxidation in muscles (Zabolotny et al., 2004; Zhang, 2002). So, numerous efforts directly towards the discovery of specific PTP1B inhibitors have been made.
Natural product Hypericum perforatum L. has been used in folk medicine to improve mental performance. It contains several kinds of chemical compounds, such as flavonoids, proanthocyanidins, naphthodianthrones (including hypericin and pseudohypericin), and acylphloroglucinols (including hyperforin and adhyperforin) (Nahrstedt and Butterweck, 1997). Recently, evidence suggests that H. perforatum L has multiple activities with potential pharmacological interest, including antibacterial, antitumoral, and antiangiogenic effects (Medina et al., 2006). However, it is still unknown whether H. perforatum L. has an effect on metabolic syndrome. Here, we found that the extract from H. perforatum L. (EHP) inhibited PTP1B and improved insulin resistance and lipid metabolism.
Materials and Methods
Plant material and extractions
The aerial parts of H. perforatum L. were collected in Huocheng County, Xinjiang, China, and identified by Prof. Fa Liu of Xinjiang Institute of Materia Medica. The voucher specimen (ID-S-2250) was deposited at the herbarium of Institute of Materia Medica, Chinese Academy of Medical Sciences, China. The dried aerial parts of H. perforatum L. were extracted with 95% ethanol at room temperature. The solvent was removed under reduced pressure for isolation of the extract. To this extract, H2O was added and the solution was filtered with a Diaion HP-20 column. The retained compounds were eluted from the column with H2O-95% ethanol (1:0–0:1) to yield four fractions. The fraction 4 was named as EHP (the extraction rate was about 0.75% of dry weight of H. perforatum L.), and the chemical character was determined with thin-layer chromatography, ultraviolet (UV) and high-performance liquid chromatography (HPLC), HPLC-mass spectrometry (MS) fingerprint analysis, respectively.
Data analysis: In order to identify the chemical components in the extract, the fingerprints of EHP were analyzed using the on-line diode array detection-electrospray ionization (DAD-ESI)-MS techniques. It was found that the negative mode contained more ions than the positive mode; therefore, ESI in the negative mode was selected for the following analysis.
HPLC analysis: HPLC analysis was performed with an Agilent 1100 series system-(Palo Alto, CA, USA). An Agilent Zorbax Eclipse XDB-C18 column (150 mm × 2.1 mm, i.d., 5 µm) at 35 °C eluented 30% 20 mM NH4AC water solution (A) and 70% CH3CN(B) in 30 min at a flow rate of 0.3 mL/min, which was monitored at 590 nm.
LC-MS analysis: The MS analysis was performed using an Agilent 1100 series LC/MSD Trap system, the conditions of HPLC as An Agilent Zorbax Eclipse XDB-C18 column (150 mm × 2.1 mm, i.d., 5 µm) maintained at 35 °C. The mobile phase was 30% 20 mM NH4AC water solution (A) and 70% CH3CN (B) in 30 min at a flow rate of 0.3 mL/min, monitored at 590 nm. The conditions of the ESI source were as follows: source voltage, 3500 V; drying gas (N2) flow rate 9.00 L/min; drying gas temperature, 35 °C; pressure of nebulizer, 40.00 psi. Scan range of MS spectra was set between 100 U and 1000 U in negative mode.
Animals and treatments
Four-week-old, male C57BL/6J mice were obtained from the Animal Center of the Institute of Laboratory Animal Sciences, Chinese Academy of Medical Sciences. The animals were housed at 21–23 °C and at a humidity level of 40–60%. They were exposed to a 12-h lighting cycle (06:00–18:00 light, 18:00–06:00 dark) and had free access to water and diets. After a 14-week feeding with the high-fat-diet (50% fat, 36% carbohydrate, and 14% protein in calorie), the DIO mice were randomly divided into four groups: untreated (Model), Rosi (positive control), EHP-L (low dose), and EHP-H (high dose). These groups were treated with water, rosiglitazone (15 mg/kg/day), EHP 50 mg/kg/day and 200 mg/kg/day by gavage (once a day) for 3 weeks. In addition, age-matched male C57BL/6J mice, fed with chow (12% fat, 62% carbohydrate, and 26% protein in calorie), were used as normal control (Con). All experimental procedures were approved by the Animal Care and Use Committee in our institute.
The oral glucose tolerance test (OGTT), the insulin tolerance test (ITT), and the hyperinsulinemic–euglycemic clamp test were performed as described previously (Ye et al., 2008) with some modification. The PTP1B activity was determined according to the method reported (Ma et al., 2011).
Materials
Commercial kits for the measurements of serum triglyceride (TG) and total cholesterol (TC) were obtained from BIOSINO Bio-Technology and Science INC (Beijing, China). Insulin was purchased from Novo Nordisk (Denmark); sodium pentobarbital, glucose, and other reagents were purchased from the Beijing Chemical Reagents Company (Beijing, China). The reagents para-NitroPhenyl Phosphate (pNPP) and para-nitrophenol (pNP) were produced by Sigma-Aldrich Trading Co., Ltd., USA. Anti-PTP1B antibody was obtained from Upstate (CA, USA), and anti-Actin antibody were purchased from Santa Cruz Biotechnology (CA, USA).
Biochemical assays
Blood glucose level was determined with a glucose analyzer (Biosen 5030, EKF Diagnostic, Germany). Protein concentration was determined with the Bio-Rad protein assay reagent (Bio-Rad, Hercules, CA, USA) with a microplate spectrophotometer (μQuant, BioTek Instruments, Inc, USA). The TG and TC levels were determined according to the instructions in the assay kit. Muscle triglyceride content was determined as glycerol residues after extraction and separation of the muscle samples according to the methods as reported (Ragheb et al., 2009). Serum insulin concentration was estimated with a radioimmunoassay kit obtained from the Northern Bioengineering Institute (Beijing, China) using a liquid scintillation and luminescence counter (1450 MicroBeta, PerkinElmer, USA).
The mRNA levels were analyzed by reverse transcription polymerase chain reaction (RT-PCR). Total RNA was isolated from skeletal muscle with Trizol reagent and reverse transcribed by a two-step method with the Super Script First-Strand Synthesis System. The samples were chosen randomly from each group, and the assay of every gene (Table1) in each sample was replicated three times. The β-actin gene was amplified as a loading control. The PCR products were separated by electrophoresis on a 1% agarose gel.
Table 1.
Sequences of the primers used in the PCR measurements
| Gene | Sense | Sequence (5′ to 3′) | Gen bank no./ref. |
|---|---|---|---|
| PTP1B | Forward | TTCTCCTACCTGGCTGTCATCG | NM 011201 |
| Reverse | CCCACCATCCGTCTCCTAACT | ||
| ACCβ | Forward | CTGCTCCAGGCTAAGCGATT | NM 133904.1 |
| Reverse | AGCCGATTTGGAAGGTGATG | ||
| CD36 | Forward | TGTACCTGGGAGTTGGCGAG | NM 007643 |
| Reverse | CTGCTGTTCTTTGCCACGTC | ||
| FATP1 | Forward | AGCCATCTGGGAGGAGTTCA | NM 011977 |
| Reverse | AGAGGGTCCTGCTGGTTGAT | ||
| CPT-1 | Forward | GTTCAACACTACACGCATCCC | NM 009948 |
| Reverse | GGTCACAAAGAAAGCAGCAC | ||
| Actin | Forward | CCCATCTACGAGG GCTAT | NM 007393 |
| Reverse | TGGCC AGTAA TGTCC AGG |
The protein expression was estimated by western blot analysis with the method reported (Ma et al., 2011). Immunoblotting analysis was performed with enhanced chemiluminescence (ECL) reagent and detected by a gel analysis system (Flurochem 5500, Alpha Innotech, USA).
Statistical analysis
Data were analyzed by SPSS version 11.0. All results are expressed as mean ± SD (standard deviation.). P values less than 0.05 were considered statistically significant.
Results
EHP extraction and isolation procedures
In the chromatogram recorded of EHP extracts at 590 nm (Fig.1A), two peaks were found, Rt = 2.28 min and Rt = 6.48 min; and 590 nm in their UV–vis spectra (Fig.1B) was the characteristic signal of napthodianthrones.
Figure 1.
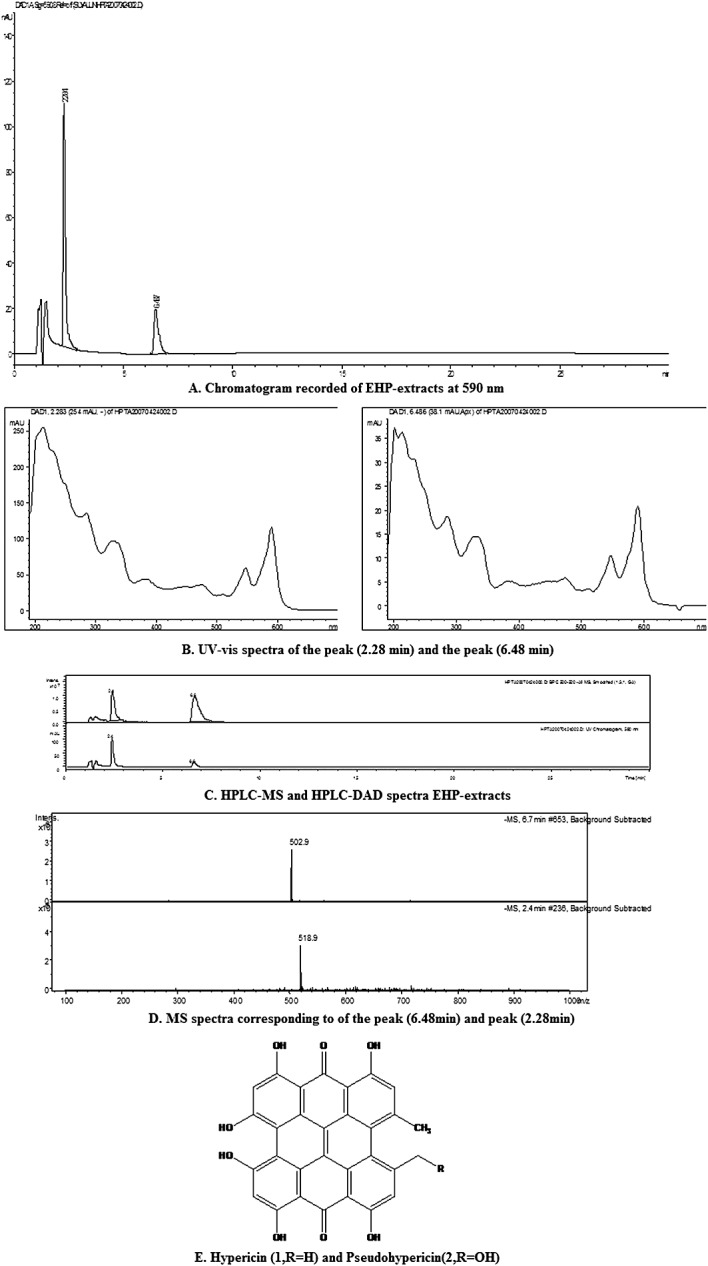
EHP-HPLC fingerprint analysis A, Chromatogram recorded of EHP-extracts at 590 nm; B, UV–vis spectra of peak (2.28 min) and peak (6.48 min); C, HPLC-MS and HPLC-DAD spectra EHP-extracts; D, MS spectra corresponding to the peak (2.28 min and 6.48 min); E, Hypericin (1, R = H) and Pseudohypericin (2, R = OH).
In the negative ESI-MS spectra of EHP-extracts (Fig.1C, D) the quasi-molecular ion peaks of Rt = 2.28 min and Rt = 6.48 min appear m/z 518.9 and 502.9 as [M − H]− ions, we finally tentatively elucidated compounds to be hypericin (1, Rt = 6.48 min) and pseudohypericin (2, Rt = 2.28 min) (Fig.1E) combined with UV–vis spectra. EHP extract was separated and identified by means of HPLC-MS analysis that it contained mainly hypericin analogues.
Effects of EHP on insulin resistance
After feeding with the high-fat-diet for 14 weeks, DIO mice developed significant insulin resistance. Fasting insulin and the homeostasis model assessment of insulin resistance (HOMA-IR) index were increased by 12.11 and 7.97 folds compared with those in the chow group. After 1-week treatment, the levels of serum insulin and HOMA-IR were markedly decreased by 64.6% and 74.6% in EHP-L group, and by 69.7% and 74.6% in EHP-H group. Similar results were observed in Rosi group (Table2). These data suggest that EHP is able to improve insulin sensitivity in DIO mice.
Insulin sensitivity related plasma parameters in mice
| FBG (mg/dL) | FINS (μIU/L) | HOMA-IR (mg/dl × μIU/L) | |
|---|---|---|---|
| Con | 74.1 ± 15.4 | 6.8 ± 2.8 | 23.5 ± 13.3 |
| Model | 105.7 ± 14.4### | 54.2 ± 34.7### | 284.5 ± 151.7### |
| Rosi | 76.4 ± 11.2*** | 9.3 ± 2.5** | 30.8 ± 4.6*** |
| EHP-L | 81.5 ± 32.6* | 19.2 ± 7.3** | 72.2 ± 46.8** |
| EHP-H | 95.7 ± 19.3 | 16.4 ± 4.4** | 72.4 ± 32.1*** |
FBG, the level of fasting blood glucose; FINS, the level of fasting serum insulin. HOMA-IR index was calculated by the formula (HOMA-IR = FINS × FBG/22.5). n = 9.
, P < 0.001 vs Con; *,**,***, P < 0.05, 0.01, 0.001 vs Model.
After EHP treatment for 1 week, OGTT was performed. The blood glucose and the area under the curve (AUC) in Model group were significantly higher than those in the chow control group. Compared with those in Model group, the levels of blood glucose at 30, 60, and 120 min were decreased by 28.6% in EHP-L group and by 36.0% in EHP-H group; and the AUC values were also decreased by 19.6% and 22.4% in EHP-L and EHP-H groups (Fig.2A, B). Similar results were obtained in Rosi group. These data suggest that EHP is able to improve glucose metabolism in DIO mice.
Figure 2.
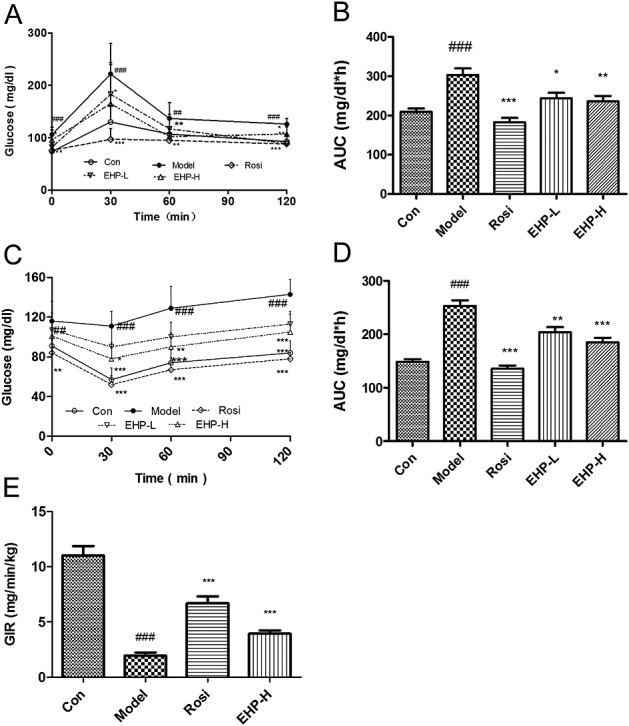
Effects of EHP on insulin resistance in mice A, Blood glucose levels in OGTT; B, Areas under the blood glucose-time curves in OGTT; C, Blood glucose levels in ITT; D, Areas under the blood glucose–time curves in ITT; E, GIR value in hyperinsulinemiceuglycemic clamp test. n = 9. #P < 0.05, ##P < 0.01, ###P < 0.001 vs Con; *P < 0.05, **P < 0.01, ***P < 0.001 vs Model.
After EHP treatment for 1 week, ITT was performed. The blood glucose level was decreased by 36.9% in chow group and by 4.3% in Model group at 30 min, compared with their own baseline. The AUC value in Model group was much higher than that in chow group. As expected, the mice treated with EHP exhibited an enhanced ability of glucose clearance in ITT. There was a greater decline in blood glucose levels by 16.6% and 30.0% in EHP-L and EHP-H groups. The reduction was as associated with 19.7% and 29.9% reduction in AUC compared with those in Model group (Fig.2C, D). The effect of EHP was similar to that of rosiglitazone in ITT. The data suggest that EHP is able to improve insulin tolerance in DIO mice and the activity is comparable to rosiglitazone.
In order to confirm insulin sensitizing effect of EHP, the hyperinsulinemic–euglycemic clamp test was performed in mice after EHP treatment for 2 weeks. The value of the glucose infusion rate (GIR) was 5.5-fold lower in Model group than that in chow group. After rosiglitazone and EHP treatments, the GIR values were increased by 240% and 101% in Rosi and EHP-H groups, respectively, compared with that in Model group (Fig.2E).
Inhibition of PTP1B by EHP
In order to understand the mechanism by which EHP improves insulin sensitivity, we tested EHP for inhibition of PTP1B in vitro. EHP exhibited a strong inhibitory effect on the catalytic activity of recombinant hPTP1B protein with an EC50 of 1.08 µg/mL (Fig.3A).
Figure 3.
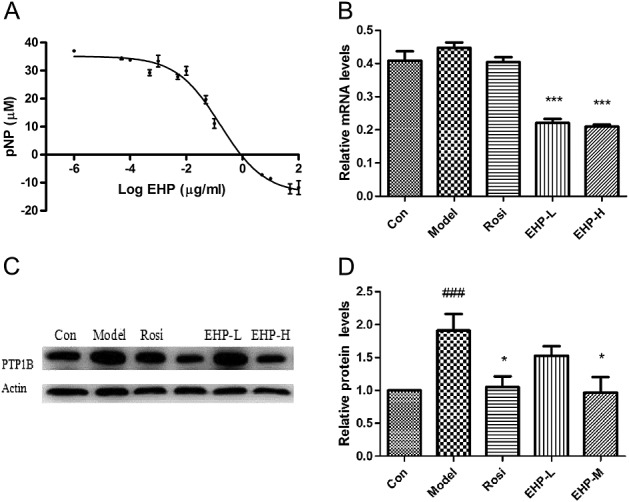
Effects of EHP on target PTP1B A, Inhibition on recombinant hPTP1B protein activity, n = 3; B, The mRNA levels of PTP1B in skeletal muscle, n = 5; C, The expression of PTP1B in skeletal muscle. #P < 0.05, ###P < 0.001 vs Con; *P < 0.05, ***P < 0.001 vs Model.
After treatment with EHP for 3 weeks, the mRNA levels of PTP1B were decreased by 32.8% and 50.7% in a dose dependent manner in the skeletal muscle of the EHP-L and EHP-H groups (Fig.3B). Meanwhile, the protein levels of PTP1B were markedly up-regulated in the skeletal muscle of DIO mice, and the increase was reversed by 21.8% and 48.8% in EHP-L and EHP-H groups (Fig.3C).
Effects of EHP on lipid metabolic profiles
DIO mice developed hypercholesterolemia, but not hypertriglyceridemia. Compared with the levels in DIO mice, the serum TC was significantly decreased by 16.2% and 22.2%, and the levels of serum TG were decreased by 13.1% and 22.6% in the EHP-L and EHP-H groups in a dose-dependent manner (Table 3).
Effects of EHP on lipid profiles in high-fat-fed mice
| Serum TC (mg/dL) | Serum TG (mg/dL) | Muscle TG (mg/g pro) | |
|---|---|---|---|
| Con | 83 ± 7 | 114 ± 38 | 3.0 ± 0.6 |
| Model | 216 ± 28### | 84 ± 14# | 4.5 ± 0.9## |
| Rosi | 199 ± 18 | 47 ± 11*** | 4.4 ± 1.3 |
| EHP-L | 181 ± 26* | 73 ± 16 | 4.1 ± 1.2 |
| EHP-H | 168 ± 30** | 65 ± 19* | 3.4 ± 0.8* |
n = 9.
,
,
, P < 0.05, 0.01, 0.001 vs Con;
,
,
, P < 0.05, 0.01, 0.001 vs Model.
The intromyocellular lipid accumulation in skeletal muscle has been associated with insulin resistance and dyslipidemia. The results showed that triglyceride content in the skeletal muscles was significantly increased in DIO mice, and the change was reversed by EHP treatment. Compared with those in DIO mice, the triglyceride contents in the skeletal were decreased by 8.9% and 25.3% in EHP-L and EHP-H groups in a dose-dependent manner (Table3).
Effects of EHP on the lipid metabolic pathways in the skeletal muscle
In order to understand the mechanisms of EHP action on dyslipidemia, gene expression was examined for those involved in lipid metabolism. As a major transporter of fatty acid, FATP1, was significantly increased in the skeletal muscle of DIO mice. Also, EHP decreased the gene expression of FATP1 in a dose-dependent manner. However, there were no significant changes in mRNA levels of CD36 from EHP treatment (Fig.4).
Figure 4.
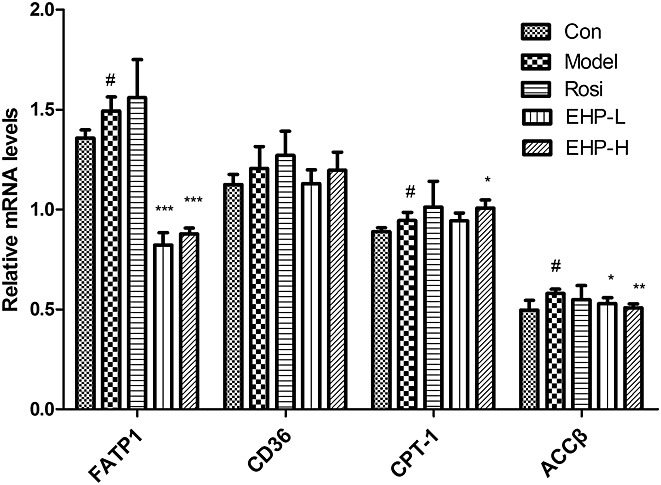
Effects of EHP on the factors involved fatty acid metabolism in skeletal muscle. Each sample was from 3 independent experiments of 9 mice. #P < 0.05 vs Con; *P < 0.05, **P < 0.01, ***P < 0.001 vs Model.
As an important regulator of fatty acid oxidation, CPT-1 was mildly increased in the skeletal muscle of DIO mice. EHP increased CPT-1 expression at the high dosage. The mRNA level of ACCβ was higher in Model group than that in chow group, and the change was reversed by EHP treatment (Fig.4).
Discussion
Insulin resistance, characterized by reduced responsiveness to circulating insulin, is a common feature in type 2 diabetes (Ye, 2013; Choudhary et al., 2012). DIO mice are characterized with obesity and insulin resistance and area valuable tool in the study of insulin sensitizer (Almind and Kahn, 2004; Biddinger and Ludwig, 2005). Consistently, our results showed the mice challenged with high-fat-diet developed serious insulin resistance and dyslipidemia.
As the largest organ in the body, the skeletal muscle plays a very important role in the glucose and lipid metabolism (Pedersen and Febbraio, 2012). Many proteins produced by the skeletal muscle may be involved in the association between sedentary behavior and chronic diseases such as cardiovascular diseases, type 2 diabetes. Therefore, our study of EHP was conducted in the skeletal muscle.
Defects in the insulin signaling cascade play a key role in the pathogenesis of insulin resistance (Choi and Kim, 2010; Ye, 2013). PTP1B is a negative regulator of both insulin and leptin signaling through antagonizing the activities of insulin receptor and Janus kinase 2 (JAK2) (Koren and Fantus, 2007). Whole-body Ptp1b−/− mice show enhanced insulin sensitivity and increased phosphorylation of tyrosine residues on insulin receptor (Klaman et al., 2000). PTP1B inhibitor can augment insulin sensitivity through enhancing insulin receptor and leptin receptor activities (Picardi et al., 2008). Here, both the catalytic activity and gene expression of PTP1B were decreased by EHP treatment, suggesting that EHP is an inhibitor of PTP1B. HEP led to improvement of hyperinsulinmia, hyperglycemia, insulin tolerance, and GIR in the hyperinsulinemic–euglycemic clamp test.
PTP1B is a molecular target for lipid disorder in addition to insulin sensitization (Picardi et al., 2008). EHP can decrease the serum triglyceride, cholesterol, and ectopic lipid accumulation in skeletal muscle. The putative mechanism is increasing fatty acid oxidation in skeletal muscle of DIO mice.
Insulin resistance in skeletal muscle is associated with intramuscular lipid accumulation, which is a result of insulin signaling inhibition by lipids (Holland et al., 2007). FATP1 that mediates fatty acid uptake into cells is implicated in the development of insulin resistance. FATP1 deletion in KO mice protected mice from fat-induced insulin resistance and intramuscular accumulation of fatty acyl-CoA (Kim et al., 2004). FATP1 inactivation led to reduction in skeletal muscle lipid accumulation and improvement in insulin sensitivity (Wu et al., 2006). In this study, the mRNA levels of FATP1 in DIO mice were increased by HFD and reduced by EHP. FATP1 reduction may explain the decreased TG content in skeletal muscle of EHP groups. ACCβ catalyzes malonyl CoA formation and high expression of ACCβ increases malonyl CoA level, which lead to inhibition of CPT-1 activity. CPT-1 inhibition results in suppression of the fatty acid β-oxidation and increase in intracellular lipid accumulation (Cheng et al., 2002). Here, EHP significantly increased mRNA levels of genes related to fatty acid oxidation, suggesting that EHP regulates fatty acid oxidation. However, EHP did not markedly decrease the liver TG content (date not shown). These results suggested that the skeletal muscles might be the major target tissue of EHP.
In conclusion, the present results demonstrated that EHP, an extraction from H. perforatum L., serves as a new PTP1B inhibitor. It improves insulin resistance and dyslipidemia in DIO C57BL/6J mice. EHP is a potential drug candidate in the treatment of metabolic syndrome.
Acknowledgments
This work was supported by the National Major Special Project on New Drug Innovation of China (no. 2012ZX09103-101-063; 2012ZX09301002-004). We thank Prof. Fa Liu of Xinjiang Institute of Materia Medica (China) for the plant identification and Servier Company (France) for the kind contribution of PTP1B plasmid.
Conflict of interest
The authors declare no conflict of interest.
References
- Almind K, Kahn CR. Genetic determinants of energy expenditure and insulin resistance in diet-induced obesity in mice. Diabetes. 2004;53:3274–3285. doi: 10.2337/diabetes.53.12.3274. [DOI] [PubMed] [Google Scholar]
- Basavarajappa DK, Gupta VK, Rajala RV. Protein Tyrosine Phosphatase 1B: A Novel Molecular Target for Retinal Degenerative Diseases. Adv Exp Med Biol. 2012;723:829–834. doi: 10.1007/978-1-4614-0631-0_106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biddinger SB, Ludwig DS. The insulin-like growth factor axis: a potential link between glycemic index and cancer. Am J Clin Nutr. 2005;82:277–278. doi: 10.1093/ajcn.82.2.277. [DOI] [PubMed] [Google Scholar]
- Cheng A, Uetani N, Simoncic PD, et al. Attenuation of leptin action and regulation of obesity by protein tyrosine phosphatase 1B. Dev Cell. 2002;2:497–503. doi: 10.1016/s1534-5807(02)00149-1. [DOI] [PubMed] [Google Scholar]
- Choi K, Kim YB. Molecular mechanism of insulin resistance in obesity and type 2 diabetes. Korean J Intern Med. 2010;25:119–129. doi: 10.3904/kjim.2010.25.2.119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choudhary N, Kalra S, Unnikrishnan AG, et al. Preventive pharmacotherapy in type 2 diabetes mellitus. Indian J Endocrinol Metab. 2012;16:33–43. doi: 10.4103/2230-8210.91183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grundy SM. Drug therapy of the metabolic syndrome: minimizing the emerging crisis in polypharmacy. Nat Rev Drug Discov. 2006;5:295–309. doi: 10.1038/nrd2005. [DOI] [PubMed] [Google Scholar]
- Haus JM, Solomon TP, Lu L, et al. Intramyocellular lipid content and insulin sensitivity are increased following short-term low-glycemic index diet and exercise intervention. Am J Physiol Endocrinol Metab. 2011;301:E511–E516. doi: 10.1152/ajpendo.00221.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holland WL, Knotts TA, Chavez JA, et al. Lipid mediators of insulin resistance. Nutr Rev. 2007;65(6 Pt 2):S39–S46. doi: 10.1111/j.1753-4887.2007.tb00327.x. [DOI] [PubMed] [Google Scholar]
- Johnson TO, Ermolieff J, Jirousek MR. Protein tyrosine phosphatase 1B inhibitors for diabetes. Nat Rev Drug Discov. 2002;1:696–709. doi: 10.1038/nrd895. [DOI] [PubMed] [Google Scholar]
- Kim JK, Gimeno RE, Higashimori T, et al. Inactivation of fatty acid transport protein 1prevents fat-induced insulin resistance in skeletal muscle. J Clin Invest. 2004;113:756–763. doi: 10.1172/JCI18917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klaman LD, Boss O, Peroni OD, et al. Increased energy expenditure, decreased adiposity, and tissue-specific insulin sensitivity in protein-tyrosine phosphatase 1B-deficient mice. Mol Cell Biol. 2000;15:5479–5489. doi: 10.1128/mcb.20.15.5479-5489.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korach-André M, Gounarides J, Deacon R, et al. Age and muscle-type modulated role of intramyocellular lipids in the progression of insulin resistance in nondiabetic Zucker rats. Metabolism. 2005;54:522–528. doi: 10.1016/j.metabol.2004.11.006. [DOI] [PubMed] [Google Scholar]
- Koren S, Fantus IG. Inhibition of the protein tyrosine phosphatase PTP1B: potential therapy for obesity, insulin resistance and type-2 diabetes mellitus. Best Pract Res Clin Endocrinol Metab. 2007;21:621–640. doi: 10.1016/j.beem.2007.08.004. [DOI] [PubMed] [Google Scholar]
- Kuhlmann J, Neumann-Haefelin C, Belz U, et al. Correlation between insulin resistance and intramyocellular lipid levels in rats. Magn Reson Med. 2005;53:1275–1282. doi: 10.1002/mrm.20501. [DOI] [PubMed] [Google Scholar]
- Ma YM, Tao RY, Liu Q, et al. PTP1B inhibitor improves both insulin resistance and lipid abnormalities in vivo and in vitro. Mol Cell Biochem. 2011;357:65–72. doi: 10.1007/s11010-011-0876-4. [DOI] [PubMed] [Google Scholar]
- Medina MA, Martínez-Poveda B, Amores-Sánchez MI, et al. Hyperforin: more than an antidepressant bioactive compound? Life Sci. 2006;79:105–111. doi: 10.1016/j.lfs.2005.12.027. [DOI] [PubMed] [Google Scholar]
- Nahrstedt A, Butterweck V. Biologically active and other chemical constituents of the herb of Hypericumperforatum L. Pharmacopsychiatry. 1997;2:129–134. doi: 10.1055/s-2007-979533. [DOI] [PubMed] [Google Scholar]
- Novgorodtseva TP, Karaman YK, Zhukova NV, et al. Composition of fatty acids in plasma and erythrocytes and eicosanoids level in patients with metabolic syndrome. Lipids Health Dis. 2011;10:82. doi: 10.1186/1476-511X-10-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pedersen BK, Febbraio MA. Muscles, exercise and obesity: skeletal muscle as a secretory organ. Nat Rev Endocrinol. 2012;8:457–465. doi: 10.1038/nrendo.2012.49. [DOI] [PubMed] [Google Scholar]
- Picardi PK, Calegari VC, Prada PO, et al. Reduction of hypothalamic protein tyrosine phosphatase improves insulin and leptin resistance in diet-induced obese Rats. Endocrinology. 2008;149:3870–3880. doi: 10.1210/en.2007-1506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ragheb R, Shanab GM, Medhat AM, et al. Free fatty acid-induced muscle insulin resistance and glucose uptake dysfunction: Evidence for PKC activation and oxidative stress-activated signaling pathways. Biochem Biophys Res Commun. 2009;389:211–216. doi: 10.1016/j.bbrc.2009.08.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz I, Eldor R, Cernea S, Shafrir E. Diabetes: insulin resistance and derangements in lipid metabolism. Cure through intervention in fat transport and storage. Diabetes Metab Res Rev. 2005;21:3–14. doi: 10.1002/dmrr.493. [DOI] [PubMed] [Google Scholar]
- Sun T, Wang Q, Yu Z, et al. Hyrtiosal, a PTP1B inhibitor from the marine sponge Hyrtios erectus, shows extensive cellular effects on PI3K/AKT activation, glucose transport, and TGFbeta/Smad2 signaling. ChemBioChem. 2007;8:187–193. doi: 10.1002/cbic.200600349. [DOI] [PubMed] [Google Scholar]
- Takasu S, Mutoh M, Takahashi M, et al. Lipoprotein lipase as a candidate target for cancer prevention/therapy. Biochem Res Int. 2012;2012:398697. doi: 10.1155/2012/398697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams KJ. Molecular processes that handle-and mishandle-dietary Lipids. J Clin Invest. 2008;118:3247–3259. doi: 10.1172/JCI35206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu Q, Ortegon AM, Tsang B, et al. FATP1 is an insulin-sensitive fatty acid transporter involved in diet-induced obesity. Mol Cell Biol. 2006;26:3455–3467. doi: 10.1128/MCB.26.9.3455-3467.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye J. Challenges in drug discovery for thiazolidinedione substitute. Acta Pharma Sinica B. 2011;1:137–142. doi: 10.1016/j.apsb.2011.06.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye J. Mechanisms of insulin resistance in obesity. Front Med. 2013;7:14–24. doi: 10.1007/s11684-013-0262-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye F, Tao R, Cong W, et al. Utilization of fluorescence tracer in hyperinsulinemic-euglycemic clamp test in mice. J Biochem Biophys Methods. 2008;70:978–984. doi: 10.1016/j.jprot.2008.01.003. [DOI] [PubMed] [Google Scholar]
- Zabolotny JM, Haj FG, Kim YB, et al. Transgenic overexpression of protein-tyrosine phosphatase 1B in muscle causes insulin resistance, but overexpression with leukocyte antigen-related phosphatase does not additively impair insulin action. J Biol Chem. 2004;279:24844–24851. doi: 10.1074/jbc.M310688200. [DOI] [PubMed] [Google Scholar]
- Zhang YZ. Protein tyrosine phosphatases: structure and function, substrate specificity, and inhibitor development. Annu Rev Pharmacol Toxicol. 2002;42:209–234. doi: 10.1146/annurev.pharmtox.42.083001.144616. [DOI] [PubMed] [Google Scholar]