

Abstract
Introduction
The ‘diurnal slope’ of salivary cortisol has been used as a measure of stress and circadian function in a variety of reports with several detailing its association with cancer progression. The relationship of this slope, typically a negative value from high morning concentrations to low evening concentrations, to the underlying daily variation in total plasma cortisol throughout the 24-hour cycle, however, has never been reported.
Methods
To examine the relationship between diurnal salivary cortisol slope and the underlying pattern of plasma cortisol in individuals with cancer, we examined a cohort of women with advanced breast cancer (n=97) who had saliva and plasma collected during a modified 24-hour, constant posture protocol.
Results
We found that steepness of the diurnal slope of salivary cortisol was correlated with the amplitude of plasma cortisol rhythm when the slope was calculated from samples taken at wake+30 minutes and 9PM (r=−0.29, p>0.05). Other variants of salivary slope calculations were not significantly correlated with the amplitude of the plasma cortisol rhythm. Diurnal salivary cortisol slope steepness was not correlated with the time between habitual waking and the computed circadian peak of cortisol, but there was a correlation between diurnal slope steepness and the time between habitual waking and the time of the awakening spike of morning cortisol (r’s<−0.23, p’s<0.05).
Conclusion
It therefore appears that in women with advanced breast cancer, diurnal salivary cortisol slope primarily represents aspects of the cortisol awakening response in relation to evening levels more than the circadian rhythm of total plasma cortisol.
Keywords: cortisol, diurnal rhythm, saliva, plasma, sleep, breast cancer, women, circadian rhythm
Introduction
Many studies have reported on a ‘diurnal slope’ of salivary cortisol concentrations during waking hours, with the slope representing the change in salivary cortisol concentrations between the morning (typically at or near wake time) and evening (typically at or near bed time). The normal pattern is described as a negative slope (higher in the morning, lower in the evening), though not all healthy individuals exhibit this pattern [1]. An altered diurnal slope of cortisol has been reported in association with a variety of pathologies, including, but certainly not limited to breast cancer [2;3], ovarian cancer [4], lung cancer [5], coronary artery disease [6], fibromyalgia [7], depression [8], early-life abuse [9], and disrupted sleep [10]. Flattening of the diurnal slope has been found to predict more rapid disease progression and shorter survival in breast [2], ovarian [4], and lung [5] cancers. Many of these reports use the diurnal slope of cortisol to impute the strength or normality of circadian rhythms. There is a robust circadian variation in the release of cortisol from the adrenal cortex such that levels begin to rise prior to habitual wake time and reach a nadir in the evening [11]. There is a small, suppressive effect of sleep on plasma cortisol patterns [12], but otherwise, the underlying 24-hour pattern of cortisol is believed to be primarily driven by the suprachiasmatic nuclei, the central circadian pacemaker in mammals [11]. Salivary concentrations of cortisol are well correlated with free (unbound) plasma concentrations of cortisol as this fraction of plasma cortisol moves freely between the blood and salivary compartments [13]. There is, therefore, logical reason to connect the pattern of salivary cortisol with underlying circadian timing.
There are, however, several reasons why the diurnal slope of daytime salivary cortisol might not accurately represent the underlying 24-hour circadian pattern of plasma cortisol. First, as salivary cortisol is often collected in circumstances in which the environment is not controlled (i.e., ambulatory), concentrations may be subject to acute stressors more so than in the laboratory environment in which plasma cortisol is collected. For example, an acute stress in the evening that evoked a spike of cortisol release could result in a positive diurnal slope even though the underlying circadian rhythm was normal. A second issue is that the diurnal slope is dependent on the circadian phase angle of entrainment – the relative position of the circadian rhythm of cortisol to the timing of sleep. For example, if the rhythm of cortisol were phase delayed (later than normal), the morning saliva sample would be obtained prior to the peak of cortisol. Likewise, a phase advanced rhythm (earlier than normal) would have the morning sample being collected after the peak and the evening sample being collected during the rising phase. Under both of these circumstances, the underlying circadian rhythm of cortisol would be of normal shape and amplitude, but the timing vis-à-vis sleep would be abnormal and the diurnal slope would be flat or positive due to a lower-than-peak morning sample. A third possible issue relates to the pulsatile release of cortisol. Cortisol is released in a highly pulsatile manner. Infrequent sampling, such as occurs with saliva samples, makes determination of a diurnal slope more prone to the vagaries of this pulsatility. For example, if a morning saliva sample is obtained just prior to pulse and an evening sample is obtained at the peak of a pulse, the slope would appear flat despite a normal underlying diurnal rhythm.
To determine whether differences in salivary diurnal slope are related to real or perceived differences in the underlying circadian rhythm, we compared the complete, diurnal pattern of plasma cortisol with that of contemporaneously collected salivary cortisol and the calculated diurnal slope of salivary cortisol concentrations. We did this comparison in a group of women with advanced breast cancer, a population in whom diurnal cortisol has been frequently reported [2;3;14;15].
Materials and Methods
Participants
Ninety seven women diagnosed with Advanced Breast Cancer (ABC) were recruited to participate. Participants had to meet the following inclusion criteria: documented metastatic or recurrent breast cancer, aged between 45 and 75 years, proficient in English, willing to travel to Stanford, California for three nights, postmenopausal, non-smoking, Karnofsky ratings of 70% or higher [16], and at least a high school diploma or equivalent. Exclusion criteria included: bilateral lymph node removal, low hematocrit, active cancers (other than breast cancer, basal cell or squamous cell carcinomas of the skin, or in situ cancer of the cervix) within the past 10 years, any concurrent medical condition likely to influence short-term survival, use of corticosteroids, glucocorticoids, benzodiazepines, or melatonin within the week preceding and during the in-laboratory study, history of major psychiatric illness that required hospitalization in the preceding year, current substance/alcohol abuse/dependence, or engagement in shift work during three months prior to the study. We did allow participants to be on chronic (>1 month), stable doses of antidepressant medications and medications to ameliorate pain; post hoc analyses found that the use of these medications did not affect the results presented below. We examined individual cortisol profiles of women who had been on long term glucocorticoids or corticosteroids to ensure that the use of such medications did not explain any of the findings described below. Participants were required to abstain from travelling two or more time zones away from their local time zone for the two weeks prior to and during their participation in the study. If not living within the Pacific time zone, participants agreed to maintain their local time zone schedule during a three-night visit to California.
Participants were recruited through the Stanford Cancer Center, the Army of Women website, and posting of study flyers on the internet and at various bay area clinics. All procedures were approved by the Stanford University Institutional Review Board functioning according to the third edition of the Guidelines on the Practice of Ethical Committees in Medical Research issued by the Royal College of Physicians of London. Consent was obtained from each participant after full explanation of the purpose and nature of all procedures used.
Protocol
Using a combination of sleep logs and wrist-worn actigraphs (Actiwatch 2, Philips-Respironics, Bend OR), ambulatory sleep/wake patterns were recorded in all participants for two weeks. Actigraphs record three-dimensional arm movement, which is useful for determining gross patterns of sleep and wake [17]. Actigraph data were interpreted using commercially available software (Actiware 5, Respironics, Bend OR) in combination with the sleep logs. From these data, habitual bed and wake times were calculated from the average bed and wake times on the 14 recorded days. Divergences greater than 60 minutes from these averages were excluded from the calculations.
Following the two-week at-home protocol, individuals participated in a 27-hour stay at either the Clinical Translational Research Unit (CTRU) or the Stanford Sleep Disorders Clinic at Stanford Hospital. Rooms were modified to control ambient light exposure and allow for simultaneous recording of sleep and sampling of blood without sleep disruption. Participants arrived at Stanford Hospital approximately seven hours after habitual wake time (e.g., 3 PM for an individual who habitually woke at 8am). After approximately an hour of orientation and medical screening (hematocrit, vital signs), participants entered their room and stayed in this room for the duration of their study, which was scheduled to end approximately ten hours after habitual wake time the next day (e.g., 6 PM for an individual who habitually woke at 8am). During hours of scheduled wakefulness, participants typically read, did light stretching (strenuous exercise was not allowed), or talked to staff members. An eight hour period of darkness was scheduled to occur starting four hours before the midpoint of habitual sleep and wake times. Lighting during periods of scheduled wake was dim (<15 lux), and lights were off (<0.05 lux) during the period of scheduled sleep. To avoid the metabolic consequences of large meals, participants were provided equicaloric hourly snacks, the sum total of which satisfied their nutritional requirements for the duration of the study [18].
Blood samples were collected via an indwelling venous catheter kept patent with a 0.9% sodium chloride/1000 U heparin solution delivered at a 10 mL/hr drip rate. The catheter was placed in the arm contralateral to any axillary lymph node resection. The catheter was attached to the infusion pump with 1.5 m or 3.0 m extension tubing, the former being used during the participant’s waking hours and the latter being used during the participant’s scheduled sleep time. During scheduled sleep, blood samples were collected in an adjacent room, so as not to disturb the participant’s sleep, with the intravenous tubing passing through a shielded porthole in the wall. Blood samples for determination of cortisol concentrations were collected at 20 – 60 minute intervals, with the elevated collection rate occurring from three hours prior to habitual bed time until an hour after habitual wake time.
Plasma cortisol assays and data fitting
At least 1 cc of whole blood was collected via the catheter into a tube coated with ethylenediaminetetraacetic acid (EDTA). Samples were immediately spun for 15 minutes at 1300 × g in a 4 °C centrifuge; 40 μL of the resultant plasma was decanted and stored at −80 °C until assay. All samples were assayed in duplicate using a commercially-available solid phase enzyme-linked immunosorbent assay (ELISA) (Immuno-Biological Laboratories, Minneapolis MN). Briefly, plasma samples for an individual participant were brought to room temperature and 20 μL of plasma was added into a single well on a 96-well plate. To each sample, 200 μL of enzyme conjugate (cortisol conjugated to horseradish peroxidase) was added, mixed, and incubated at room temperature for 60 minutes. The plate was washed (Wellwash AC, Thermo, Waltham MA) and dried before 100 μL of a substrate solution (tetramethylbenzidine) was added to each well, followed by a 15 minute incubation at room temperature. The reaction was stopped with 100 μL of stop solution (1N acid) and the plate was read by a microplate photometer (Multiskan, Fisher Scientific, Pittsburgh PA) at 450 nm. Standards (0, 20, 50, 100, 200, 400, 800 ng/mL) were processed in a manner identical to the samples. After adjusting for a blank well, results from the standards assay were fit with a four-parameter logistic model, from which concentrations of the unknowns were calculated. Data quality was ensured with an internal control and reviews of each curve fit and individual sample results. Intra-assay coefficients of variation for 43.5 ng/mL, 226.5 ng/mL, and 403.6 ng/mL were 8.1%, 3.2%, and 5.6% respectively. Interassay coefficients of variation for 55 ng/mL, 209 ng/mL, and 361 ng/mL were 6.6%, 7.7%, and 6.5% respectively. Assay sensitivity was 2.5 ng/mL.
Plasma cortisol has a characteristic diurnal rhythm, with a nadir occurring around the time of habitual bedtime and a peak occurring just after wake time. This pattern is only slightly altered by the presence of sleep. There are many different curve fitting techniques that have been used to describe the diurnal cortisol rhythm, including cosinor [19], two-hour smoothing [20], L2 norm approximation [21], and harmonic regression [22]. Based on preliminary analyses from an already published data set (data not shown) [23], we found that harmonic regression analysis using a three harmonic regression analysis was robust, and for the phase of the first harmonic to be most representative of circadian phase, as indicated by the timing of the rhythm of plasma melatonin [22]. As such, we fit all plasma cortisol data with a three-harmonic regression analysis (OriginPro 8.0, Microcal, Northampton MA), with fixed harmonics (waveforms) of 24−, 12−, and 8-hours. Using this technique, we were able to characterize the time of the peak and trough of the composite curve, the peak of the first harmonic (a marker of circadian phase), the amplitude of the curve (one-half peak-to-trough), the mesor (Midline Estimating Statistic Of Rhythm), and the goodness of fit (adjusted r2). We then calculated phase angles between various markers (i.e., the time between two definable portions of rhythmic behavior), including habitual wake time and the peak of the composite curve and habitual wake time and the peak of the first harmonic. The timing of the peak of the first harmonic relative to habitual wake time is a useful marker of the phase angle of entrainment – that is the position of the circadian system relative to the normal timing of sleep.
Salivary cortisol assays and data fitting
Saliva samples were collected at three time points – 9 PM on Day 1, awakening on Day 2, and 30 minutes after awakening on Day 2. These times were selected as they are commonly used in this area of research [24], although, there is considerable variation in the timing of samples used to determine the diurnal slope of salivary cortisol [25]. Three days of saliva sampling is posited as ideal to determine slope [24], so our use of a single day limits some of the statistical reliability of our analyses. As the 9PM sample was taken on Day 1, we carried forward the value of this sample as 9PM on Day 2. Given relatively little drift of the human circadian timing system during this type of protocol [26], we believe this is a valid imputation. Saliva was obtained by having the participant chew on an untreated cotton swab, which was then stored in a plastic container (Salivette, Sarstedt, Newton NC). The Salivette was stored at −80 °C until assay. For assessment of salivary concentration of cortisol, the Salivette was brought to room temperature and then spun for 15 minutes at 1500 × g in a room temperature centrifuge, with the Salivette filtering out the particulate matter in the saliva. Salivary cortisol concentrations were determined in duplicate with a commercially available enzyme immunoassay kit (Salimetrics, State College PA). In brief, 24 μL of sample was added to an individual well on a 96-well plate. After adding 200 μL of conjugate (cortisol conjugated to horseradish peroxidase) to each well, the plate was rotated for 5 minutes at 500 rpm and then left at room temperature for 55 minutes. The plate was washed and dried before 200 μL of a tetramethylbenzidine solution was added to each well. The plate was again rotated and left in the dark at room temperature for 25 minutes. The reaction was stopped with 50 μL of a stop solution, the plate was rotated for 3 minutes, and then the plate was read by the microplate photometer at 450 nm. Standards (0, 0.012, 0.037, 0.111, 0.333, 1.000, 3.000 μg/dL) were processed in a manner identical to the subject samples. After adjusting for a blank well, results from the standards assay were fit with a four-parameter logistic model, from which concentrations of the unknowns were calculated. Data quality was ensured with commercially-available controls and reviews of each curve fit and individual sample results. Intra-assay coefficients of variation for 0.097 μg/dL and 0.999 μg/dL were 3.65% and 3.35% respectively. Interassay coefficients of variation for 0.101 μg/dL and 1.020 μg/dL were 6.41% and 3.75% respectively. Assay sensitivity was 0.003 μg/dL.
Statistics
Summary data were generated using Excel (v.11.8332.8333, Microsoft, Redmond WA). Regression analyses were completed using OriginPro (Origin Labs, Northampton MA). Functional principal component analyses (fPCA) were completed using R [27;28]. fPCA is similar to PCA except that it was applied to semicontinuous data (plasma cortisol) rather than discrete data. In this fPCA analysis, participants who had cortisol concentrations available from at least 38 of 51 plasma samples and no greater than a gap of 1 hour of missing data were used (n=61). Included data gaps (3.3% of sample) were filled by linear interpolation such that all participants had a continuous string of 24 hours of data with a data point every 20 minutes. Data from individual participants were fit with a nine-Fourier-based function. These equations were then subjected to functional data analysis that computed the equations underlying the variance in the Fourier equations [29]. The relative weight of each of these underlying equations (fPCA component) was calculated for each participant and represented as an eigenvalue that could be subjected to parametric statistics. The first fPCA component explains the greatest amount of variance and each subsequent component explains less. We calculated the first four components of the fPCA as these typically explain most of the variance and remaining components often have low probative value (<1% variance explained). Each eigenvalue represents the relative amount that a cortisol pattern is represented by a specific equation or shape. For example, the first fPCA component represented the amplitude differences between subjects without changes to the overall shape of the cortisol pattern. The first four components of the fPCA were compared with diurnal slope of salivary cortisol concentrations.
Data are presented as mean ± SD.
Results
General results
Of the 97 women with advanced breast cancer (ABC) who took part in the protocol, saliva samples were available from 91 (age: 57.4 ± 7.58 years, BMI = 28.1 ± 5.78). Salivary concentrations of cortisol were 0.33 ± 0.19 μg/dL at wake time (n = 91), 0.38 ± 0.20 μg/dL at wake + 30 minutes (n = 86), and 0.077 ± 0.12 μg/dL at 9 PM (n = 91). In 83 of the women (age: 57.7 ± 7.80 years, BMI = 28.0 ± 5.76), there were contemporaneous blood samples obtained at these three times. Plasma concentrations of cortisol were 143.0 ± 49.90 ng/mL at wake time (n = 77), 154.7 ± 46.30 ng/mL at wake + 30 minutes (n = 72), and 30.46 ± 23.98 ng/mL at 9 PM (n = 82). For both the saliva and plasma, due to single missing samples in individual participants, the totals at each time point vary. Each of the pairs of blood and saliva samples was obtained within 30 minutes of one another (6.41 ± 4.78 minutes). Plasma and salivary concentrations were well correlated with one another (n=229, adjusted r2=0.47, p<0.001, linear regression), with salivary concentrations of cortisol being 2.6 ± 2.3 % of that which are found in plasma. Thus, as anticipated, the cortisol concentration within each individual salivary sample was reliably indicative of the plasma concentration of cortisol.
Salivary slope vs. plasma curve fitting
Given the analytic variability in the literature [25], we calculated the diurnal slope of salivary cortisol in a variety of ways, each using regression analysis: (1) wake sample to 9PM sample, (2) wake sample to 9PM sample with both samples natural log transformed prior to calculation of slope [24], (3) wake time + 30 minutes sample to 9PM sample, and (4) wake + 30 minutes sample to 9PM sample with both samples natural log transformed prior to calculation of slope. The diurnal slope was −0.018 ± 0.016 (Method 1, n=91), −0.12 ± 0.084 (Method 2, n=91), −0.023 ± 0.015 (Method 3, n=90), and −0.13 ± 0.095 (Method 4, n=90). Methods 1 and 3 (untransformed data) were moderately correlated (r=0.39, p<0.001) while Methods 2 and 4 (natural log transformed data) were quite strongly correlated (r=0.86, p<0.001). This is expected as the natural log transformation decreases the impact of elevated values, such as occurs at the wake + 30 minute sample, yielding relatively little difference in the slope calculated from such transformed data when the data are anchored to either the wake sample or the wake + 30 sample.
To examine our a priori hypothesis that the diurnal slope of salivary cortisol was related to the position (i.e., phase angle) of the circadian system relative to the timing of sleep, we ran regression analyses between circadian phase angle (time between habitual wake time and the peak of the first harmonic of the three-harmonic fit to the plasma cortisol data) and the diurnal slope (as calculated by four methods) (available from 78 participants due to missing plasma samples). We could, however, find no such association between diurnal slope and the phase angle of the circadian timing of cortisol (Table 1).
Table 1.
Simple linear regression (r) of four different methods of diurnal slope calculation against four measures derived from three-harmonic regression analysis of 24-hours of plasma cortisol data. The four salivary slopes (all two-point linear regression): wake time and 9PM concentrations, natural log of wake time and 9PM concentrations, wake time + 30 min and 9PM concentrations, and natural log of wake time + 30 min and 9PM concentrations. The four variables derived from plasma cortisol: the phase angle (ψ) between habitual wake time (HWT) and the peak of the composite three harmonic model (P3), the phase angle between HWT and the peak of the first harmonic (P1), the amplitude (one-half peak-to-trough) of the fit, and the mesor of the fit.
| Method of Calculation Diurnal Slope of Salivary Cortisol | ψ (HWT-P3) [hrs] | ψ (HWT-P1) [hrs] | Amplitude [μg/dL] | Mesor [μg/dL] | |
|---|---|---|---|---|---|
| 1 | Wake → 9PM | −0.25* | 0.07 | −0.10 | 0.08 |
| 2 | ln(Wake) → ln(9PM) | −0.23* | 0.09 | −0.04 | 0.14 |
| 3 | Wake+30 → 9PM | −0.19 | −0.07 | −0.29* | −0.06 |
| 4 | ln(Wake+30) → ln(9PM) | −0.23* | −0.05 | −0.11 | 0.08 |
p<0.05, linear regression.
To examine our other a priori hypothesis that the diurnal slope of salivary cortisol was related to the amplitude of the plasma cortisol rhythm, we ran simple regression analyses between the amplitude of the three-harmonic fit to the plasma cortisol data and the salivary diurnal slope (as calculated by four methods). Diurnal amplitude of the three-harmonic fit to the plasma cortisol was significantly correlated with the salivary diurnal slope only when the slope was calculated using the untransformed wake+30 as the morning value (Table 1), which would be the slope calculated from the presumptive highest to lowest points in the diurnal cortisol curve. Given that the log transformation is used to remove variability associated with peak cortisol values, the lack of association with log-transformed slopes is not altogether unsurprising. We also examined whether the mesor of the three-harmonic fit to the plasma cortisol data was associated with the salivary diurnal slope, but could find no such relationship (Table 1).
The wake+30 time point is likely to represent the cortisol awakening response – an extra elevation in the already high early morning concentrations that usually follows a morning awakening. As it was the slope of the untransformed data using the wake+30 time point of saliva collection that was associated with the amplitude of plasma cortisol, we also examined whether the phase angle of habitual wake time with the peak of the composite curve of the three-harmonic fit (presumably influenced more by the awakening response than the peak of the first harmonic of the three-harmonic fit) had a relationship to the diurnal slope of salivary cortisol. The diurnal slopes as calculated using either the log-transformed data (both techniques) and the non-transformed data (wake to 9PM only) were indeed associated with the phase angle between habitual wake time and the peak of the three-harmonic fit (Table 1). Thus the greater the temporal distance between the peak of the cortisol rhythm and habitual wake time, the less steep the slope. Thus, as a group, a “flattened” diurnal rhythm in salivary cortisol is representative of a diminution of both the amplitude of the overall plasma cortisol rhythm and a cortisol peak that is abnormally timed vis-à-vis the timing of sleep. Note, the timing of the circadian system as indicated by the time of the peak of the first harmonic, is not associated with this flattening – rather it is a delay in the morning awakening response of cortisol that is associated with the flattening of the diurnal slope.
Salivary slope vs. plasma fPCA
Of the 91 women in whom we were able to calculate the diurnal slope of salivary cortisol, 10 (11%) had a positive slope (Figure 1). As can be seen in the figure (note, due to missing blood, there are only 8 of the 10 individuals in Figure 1), there appear to be a variety of “reasons” underlying the positive value of the salivary slope. In order to analyze these patterns and determine if a specific pattern of plasma cortisol was associated with a positive or flatter diurnal slope of salivary cortisol, the plasma cortisol data were subjected to functional principal component analysis (fPCA) and then compared with salivary diurnal slope (described in Methods). Complete plasma data were available from 61 women, 6 of whom (10%) had a positive diurnal slope of salivary cortisol. The first four components of the fPCA were able to explain 81.8% of the variance of the plasma cortisol data. In comparing the first four components to the diurnal slope, the first (~r2=−0.02, p=0.90; linear regression), second (~r2=0.02, p=0.17; linear regression), and fourth (~r2=0.01, p=0.19; linear regression) components of the fPCA did not appear related to the diurnal slope. The third component of the fPCA, however, was linearly related to the diurnal slope of salivary cortisol (~r2=0.08, p<0.05; linear regression). Examination of both the individual plots (Figure 1) and the results of the fPCA (Figure 2) reveal that in some participants, the flattening of the slope is associated with a lower morning peak and an elevated evening nadir (cf., red and blue lines in Figure 2).
Figure 1.
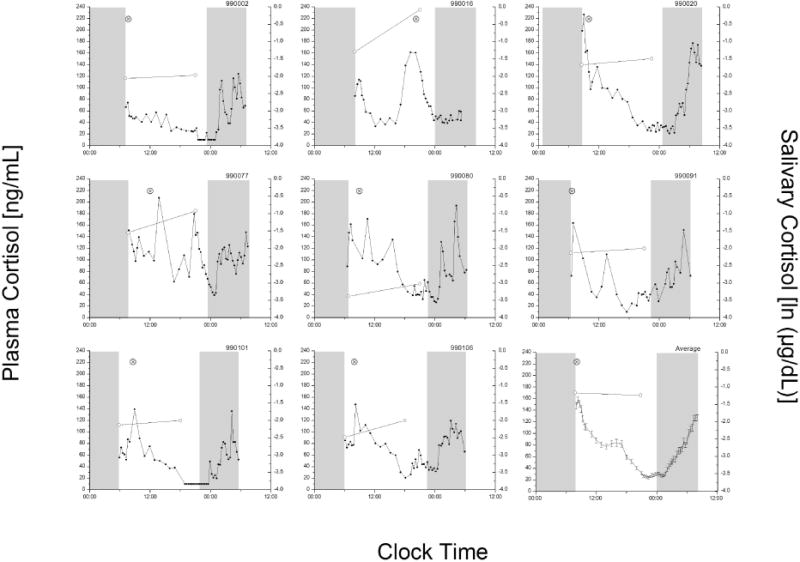
Individual plots of participants with positive diurnal salivary cortisol slopes. Each panel (except the lower right) has data collected during the protocol for the eight participants with positive slopes. The average ± SEM curves for the remaining available participants (n=75) is shown in the lower right panel. The cortisol concentration for each plasma sample (left y-axis, solid circles) and saliva sample (right y-axis, open circles) is plotted at the precise time of collection (wake and 9PM). The grey boxes represent the timing of lights-off when sleep was permitted. Circadian phase (⊗, peak of the first harmonic of the three-harmonic fit) is plotted for each participant.
Figure 2.
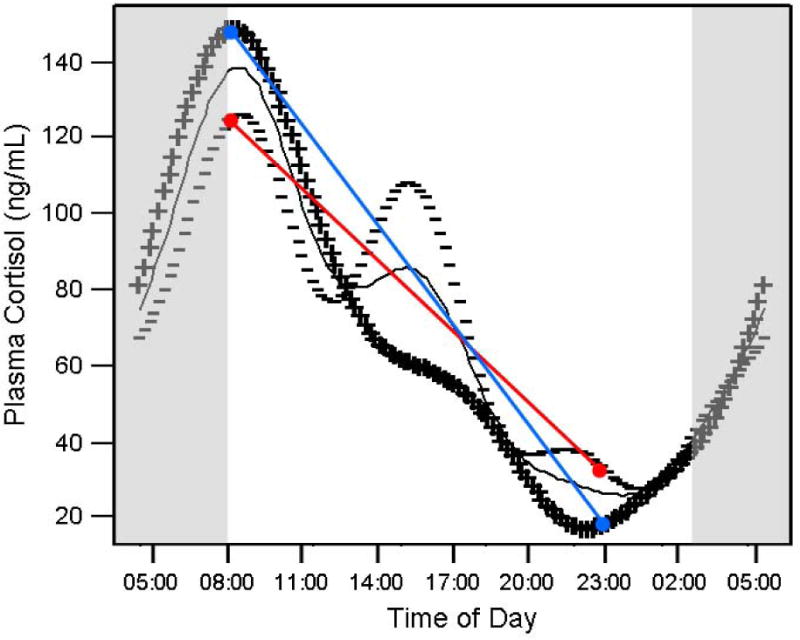
fPCA component 3 and its relationship to diurnal salivary cortisol slope. Data from high (+++) and low (−−−) eigenvalues of fPCA component 3 are plotted along with the average (solid grey line) cortisol curve of all participants. The grey boxes represent the relative time of sleep. The red and blue lines represent the possible salivary slopes as might be derived from a sample taken at wake time and 9pm, from individuals with higher (blue) or lower (red) fPCA component 3 scores.
Discussion
Our data indicate that there is no single, underlying plasma pattern that is represented by the observation of a flat or positive diurnal slope of salivary cortisol. Rather, from a statistical standpoint, there appear to be two main associations between diurnal slope and the plasma pattern of cortisol. First, in some individuals, a delay in the fitted peak of the three harmonic curve, but not the circadian peak of plasma cortisol is associated with a flattening of the salivary diurnal slope. This may be a slightly later than normal peak (e.g., participant 990016) or a peak with highly aberrant timing (e.g., participant 990101). The flat slope would, therefore, be primarily due to the morning saliva sample being obtained before the actual peak in morning cortisol. As there was no association between the steepness of the slope and the relative timing of the circadian clock (phase angle between the time of the first harmonic and the timing of sleep), this finding is unlikely to represent an abnormality of circadian timing. Rather, it is more likely to represent a delay in the so-called cortisol awakening response [30–32]. Confirmation of circadian timing with an independent marker such as melatonin, however, would be necessary to confirm the normality of circadian timing.
A second association is with a pattern that has a slightly reduced morning peak and an elevated nadir in plasma cortisol in the evening (e.g., participant 990080). The flat slope would be, therefore, due to the morning sample being slightly lower than normal and the evening sample being slightly higher than normal. This pattern has been observed before in individuals with insomnia [33] and in older individuals [34]. It is not likely due to chronic stress or depression, both of which are likely to alter the constitutive expression of cortisol, as these are sometimes characterized by constantly low (chronic stress) or elevated (depression) levels of plasma cortisol [35;36]. As the slope obtained between untransformed salivary cortisol data obtained near the cortisol awakening and the evening was the only diurnal salivary cortisol slope method that was correlated with the diurnal amplitude of plasma cortisol (Table 1), use of this slope derivation would appear to be important to accurately capture the underlying amplitude in plasma cortisol concentrations. Use of slope derived from natural log transformed data would, however, appear to capture some of the variance associated with the timing of the morning peak of cortisol vis-à-vis its timing relative to habitual wake time.
There are still participants (e.g., participant 990020) in whom there is no apparent explanation for a flattened slope. It is possible that there may be an alteration in corticosteroid binding globulins (transcortin) such that the relationship between total cortisol (plasma) and fractional free cortisol (saliva) could be inconsistent during a single day, possibly due to acute stress [37] or the breast cancer or its treatment [38], including the use of specific drugs that might change the properties of steroid binding proteins (e.g., see [39]). Further study of free plasma cortisol concentrations versus salivary cortisol concentrations would be necessary to explore this hypothesis. In one study of 109 relatively healthy individuals, 51% had normal diurnal slopes of salivary cortisol on two consecutive days and 17% had flattened slopes on two consecutive days, with the remaining 31% having a normal slope one day and a flattened slope the other day, indicating either a relative lack of reproducibility of a flat slope or a very high interday variability in salivary cortisol slope [40]. To our knowledge, this type of day-to-day variability in diurnal rhythmicity has never been reported for plasma cortisol concentrations in any normal or pathological state, raising the question as to the physiologic significance of this “flat” pattern.
Our data indicate that it is likely that individuals with a flattened diurnal slope of salivary cortisol represent a heterogeneous group. In several, but not all [3], previous studies, including those studying women with breast cancer, diurnal slope was not different between controls and patients. The presence of flatter diurnal slopes of salivary cortisol among patients, however, is associated with reduced survival or other negative health outcomes, implying an important physiological meaning for the construct [2;4;5;41;42]. In previous work, flattening of diurnal slopes of salivary cortisol among women with metastatic breast cancer has been shown to be related to higher waking rise of cortisol and reduced adrenal suppression the day after administration of 0.5 mg of dexamethasone, suggesting reduced feedback inhibition once cortisol levels are elevated, rather than hypersensitivity to activation [43].
Our current data indicate that flatter diurnal slopes of salivary cortisol are unlikely to represent an abnormality of the circadian system, but are more likely associated with a change in the morning cortisol awakening response and the timing of salivary sampling in relation to that response. As such, it beckons the question as to the physiological upstream cause of the flattened diurnal slopes that might ultimately be associated with reduced survival and negative health outcomes and why such “flattening” does not appear to be associated with acute negative outcomes in healthy individuals. Future research on the causal mechanisms of flattened diurnal slopes will be necessary to clarify this physiology.
Acknowledgments
We thank our research participants for their time, wisdom, and willingness to participate, and the Dr. Susan Love Research Foundation’s Love/Avon Army of Women Program for their assistance in recruitment. We thank Dr. Dirk Hellhammer for helpful comments on this manuscript. We also wish to thank the nursing staff of the Clinical Translational Research Unit for staffing the studies and Ms. Chung-Ping Liao and Mr. Ryan Fisicaro for conducting the cortisol assays. This work was supported by the National Cancer Institute (R01CA118567) and the National Center for Research Resources (UL1 RR025744).
References
- 1.Stone AA, Schwartz JE, Smyth J, Kirschbaum C, Cohen S, Hellhammer D, Grossman S. Individual differences in the diurnal cycle of salivary free cortisol: a replication of flattened cycles for some individuals. Psychoneuroendocrinol. 2001;26:295–306. doi: 10.1016/s0306-4530(00)00057-3. [DOI] [PubMed] [Google Scholar]
- 2.Sephton SE, Sapolsky RM, Kraemer HC, Spiegel D. Diurnal cortisol rhythm as a predictor of breast cancer survival. Journal of the National Cancer Institute. 2000;92:994–1000. doi: 10.1093/jnci/92.12.994. [DOI] [PubMed] [Google Scholar]
- 3.Abercrombie HC, Giese-Davis J, Sephton S, Epel ES, Turner-Cobb JM, Spiegel D. Flattened cortisol rhythms in metastatic breast cancer patients. Psychoneuroendocrinol. 2004;29:1082–1092. doi: 10.1016/j.psyneuen.2003.11.003. [DOI] [PubMed] [Google Scholar]
- 4.Schrepf A, Clevenger L, Christensen D, Degeest K, Bender D, Ahmed A, Goodheart MJ, Dahmoush L, Penedo F, Lucci JA, III, Ganjei-Azar P, Mendez L, Markon K, Lubaroff DM, Thaker PH, Slavich GM, Sood AK, Lutgendorf SK. Cortisol and inflammatory processes in ovarian cancer patients following primary treatment: relationships with depression, fatigue, and disability. Brain Behavior and Immunity. 2013;30(Suppl):S126–S134. doi: 10.1016/j.bbi.2012.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sephton SE, Lush E, Dedert EA, Floyd AR, Rebholz WN, Dhabhar FS, Spiegel D, Salmon P. Diurnal cortisol rhythm as a predictor of lung cancer survival. Brain Behavior and Immunity. 2013;30(Suppl):S163–S170. doi: 10.1016/j.bbi.2012.07.019. [DOI] [PubMed] [Google Scholar]
- 6.Cohen S, Schwartz JE, Epel E, Kirschbaum C, Sidney S, Seeman T. Socioeconomic status, race, and diurnal cortisol decline in the Coronary Artery Risk Development in Young Adults (CARDIA) Study. Psychosomatic Med. 2006;68:41–50. doi: 10.1097/01.psy.0000195967.51768.ea. [DOI] [PubMed] [Google Scholar]
- 7.Weissbecker I, Floyd A, Dedert E, Salmon P, Sephton S. Childhood trauma and diurnal cortisol disruption in fibromyalgia syndrome. Psychoneuroendocrinol. 2006;31:312–324. doi: 10.1016/j.psyneuen.2005.08.009. [DOI] [PubMed] [Google Scholar]
- 8.Stetler C, Dickerson SS, Miller GE. Uncoupling of social zeitgebers and diurnal cortisol secretion in clinical depression. Psychoneuroendocrinol. 2004;29:1250–1259. doi: 10.1016/j.psyneuen.2004.03.003. [DOI] [PubMed] [Google Scholar]
- 9.van der Vegt EJ, van der EJ, Kirschbaum C, Verhulst FC, Tiemeier H. Early neglect and abuse predict diurnal cortisol patterns in adults A study of international adoptees. Psychoneuroendocrinol. 2009;34:660–669. doi: 10.1016/j.psyneuen.2008.11.004. [DOI] [PubMed] [Google Scholar]
- 10.Lasikiewicz N, Hendrickx H, Talbot D, Dye L. Exploration of basal diurnal salivary cortisol profiles in middle-aged adults: associations with sleep quality and metabolic parameters. Psychoneuroendocrinol. 2008;33:143–151. doi: 10.1016/j.psyneuen.2007.10.013. [DOI] [PubMed] [Google Scholar]
- 11.Czeisler CA, Klerman EB. Circadian and sleep-dependent regulation of hormone release in humans. Rec Prog Hormone Res. 1999;54:97–132. [PubMed] [Google Scholar]
- 12.Weitzman ED, Zimmerman JC, Czeisler CA, Ronda J. Cortisol secretion is inhibited during sleep in normal man. J Clin Endocrinol Metab. 1983;56:352–358. doi: 10.1210/jcem-56-2-352. [DOI] [PubMed] [Google Scholar]
- 13.Kirschbaum C, Hellhammer DH. Salivary cortisol. In: Fink G, editor. Encyclopedia of Stress v3. San Diego: Academic Press; 2000. pp. 379–383. [Google Scholar]
- 14.Bower JE, Ganz PA, Dickerson SS, Petersen L, Aziz N, Fahey JL. Diurnal cortisol rhythm and fatigue in breast cancer survivors. Psychoneuroendocrinol. 2005;30:92–100. doi: 10.1016/j.psyneuen.2004.06.003. [DOI] [PubMed] [Google Scholar]
- 15.Vedhara K, Tuinstra J, Miles JN, Sanderman R, Ranchor AV. Psychosocial factors associated with indices of cortisol production in women with breast cancer and controls. Psychoneuroendocrinol. 2006;31:299–311. doi: 10.1016/j.psyneuen.2005.08.006. [DOI] [PubMed] [Google Scholar]
- 16.Karnofsky DA, Burchenal JH. The clinical evaluation of chemotherapeutic agents in cancer. In: MacLeod CM, editor. Evaluation of Chemotherapeutic Agents. New York: Columbia University Press; 1949. pp. 191–205. [Google Scholar]
- 17.Ancoli-Israel S, Cole R, Alessi C, Chambers M, Moorcroft W, Pollack CP. The role of actigraphy in the study of sleep and circadian rhythms. Sleep. 2003;26:342–392. doi: 10.1093/sleep/26.3.342. [DOI] [PubMed] [Google Scholar]
- 18.Mifflin MD, St Jeor ST, Hill LA, Scott BJ, Daugherty SA, Koh YO. A new predictive equation for resting energy expenditure in healthy individuals. Amer J Clin Nutrition. 1990;51:241–247. doi: 10.1093/ajcn/51.2.241. [DOI] [PubMed] [Google Scholar]
- 19.Halberg F, Johnson EA, Nelson W, Runge W, Sothern RB. Autorhythmometry – procedures forphysiologic self-measurements and their analysis. Physiology Teacher. 1972;7:1–11. [Google Scholar]
- 20.Dijk DJ, Duffy JF, Silva EJ, Shanahan TL, Boivin DB, Czeisler CA. Amplitude reduction and phase shifts of melatonin, cortisol and other circadian rhythms after a gradual advance of sleep and light exposure in humans. PLoS ONE. 2012;7:e30037. doi: 10.1371/journal.pone.0030037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chakraborty A, Krzyzanski W, Jusko WJ. Mathematical modeling of circadian cortisol concentrations using indirect response models: comparison of several methods. J Pharmacokinetics Biopharm. 1999;27:23–43. doi: 10.1023/a:1020678628317. [DOI] [PubMed] [Google Scholar]
- 22.Klerman EB, Gershengorn HB, Duffy JF, Kronauer RE. Comparisons of the variability of three markers of the human circadian pacemaker. J Biol Rhythms. 2002;17:181–193. doi: 10.1177/074873002129002474. [DOI] [PubMed] [Google Scholar]
- 23.Zeitzer JM, Dijk D-J, Kronauer RE, Brown EN, Czeisler CA. Sensitivity of the human circadian pacemaker to nocturnal light: melatonin phase resetting and suppression. J Physiol. 2000;526:695–702. doi: 10.1111/j.1469-7793.2000.00695.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kraemer HC, Giese-Davis J, Yutsis M, O’Hara R, Neri E, Gallagher-Thompson D, Taylor CB, Spiegel D. Design decisions to optimize reliability of daytime cortisol slopes in an older population. American Journal of Geriatric Psychiatry. 2006;14:325–333. doi: 10.1097/01.JGP.0000201816.26786.5b. [DOI] [PubMed] [Google Scholar]
- 25.Adam EK, Kumari M. Assessing salivary cortisol in large-scale, epidemiological research. Psychoneuroendocrinol. 2009;34:1423–1436. doi: 10.1016/j.psyneuen.2009.06.011. [DOI] [PubMed] [Google Scholar]
- 26.Duffy JF, Dijk D-J. Getting through to circadian oscillators: why use constant routines? J Biol Rhythms. 2002;17:4–13. doi: 10.1177/074873002129002294. [DOI] [PubMed] [Google Scholar]
- 27.R Development Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing; 2010. [Google Scholar]
- 28.Zeitzer JM, David R, Friedman L, Mulin E, Garcia R, Wang J, Yesavage JA, Robert PH, Shannon W. Phenotyping apathy in individuals with Alzheimer disease using functional principal component analysis. American Journal of Geriatric Psychiatry. 2013;21:391–397. doi: 10.1016/j.jagp.2012.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ramsay JO, Silverman BW. Functional Data Analysis. 2. New York: Springer; 2005. [Google Scholar]
- 30.Pruessner JC, Wolf OT, Hellhammer DH, Buske-Kirschbaum A, von AK, Jobst S, Kaspers F, Kirschbaum C. Free cortisol levels after awakening: a reliable biological marker for the assessment of adrenocortical activity. Life Sci. 1997;61:2539–2549. doi: 10.1016/s0024-3205(97)01008-4. [DOI] [PubMed] [Google Scholar]
- 31.Pruessner JC, Hellhammer DH, Kirschbaum C. Burnout, perceived stress, and cortisol responses to awakening. Psychosomatic Med. 1999;61:197–204. doi: 10.1097/00006842-199903000-00012. [DOI] [PubMed] [Google Scholar]
- 32.Wust S, Federenko I, Hellhammer DH, Kirschbaum C. Genetic factors, perceived chronic stress, and the free cortisol response to awakening. Psychoneuroendocrinol. 2000;25:707–720. doi: 10.1016/s0306-4530(00)00021-4. [DOI] [PubMed] [Google Scholar]
- 33.Vgontzas AN, Bixler EO, Lin HM, Prolo P, Mastorakos G, Vela-Bueno A, Kales A, Chrousos GP. Chronic insomnia is associated with nyctohemeral activation of the hypothalamic-pituitary-adrenal axis: clinical implications. J Clin Endocrinol Metab. 2001;86:3787–3794. doi: 10.1210/jcem.86.8.7778. [DOI] [PubMed] [Google Scholar]
- 34.Vgontzas AN, Zoumakis M, Bixler EO, Lin HM, Prolo P, Vela-Bueno A, Kales A, Chrousos GP. Impaired nighttime sleep in healthy old versus young adults is associated with elevated plasma interleukin-6 and cortisol levels: physiologic and therapeutic implications. J Clin Endocrinol Metab. 2003;88:2087–2095. doi: 10.1210/jc.2002-021176. [DOI] [PubMed] [Google Scholar]
- 35.Yehuda R, Teicher MH, Trestman RL, Levengood RA, Siever LJ. Cortisol regulation in posttraumatic stress disorder and major depression: a chronobiological analysis. Biol Psychiatry. 1996;40:79–88. doi: 10.1016/0006-3223(95)00451-3. [DOI] [PubMed] [Google Scholar]
- 36.Linkowski P, Mendlewicz J, Leclercq R, Brasseur M, Hubain P, Golstein J, Copinschi G, Van CE. The 24-hour profile of adrenocorticotropin and cortisol in major depressive illness. J Clin Endocrinol Metab. 1985;61:429–438. doi: 10.1210/jcem-61-3-429. [DOI] [PubMed] [Google Scholar]
- 37.Murphy BEP. Corticosteroid-binding globulin (transcortin) In: Fink G, editor. Stress Science: Neuroendocrinology. San Francisco: Elsevier; 2010. pp. 245–252. [Google Scholar]
- 38.Bell E, Bulbrook RD, Deshpande N. Transcortin in the plasma of patients with breast cancer. Lancet. 1967;290:395–397. doi: 10.1016/s0140-6736(67)92012-0. [DOI] [PubMed] [Google Scholar]
- 39.Löfgren L, Wallberg B, Wilking N, Fornander T, Rutqvist LE, Carlstrom K, von SB, von SE. Tamoxifen and megestrol acetate for postmenopausal breast cancer: diverging effects on liver proteins, androgens, and glucocorticoids. Medical Oncology. 2004;21:309–318. doi: 10.1385/MO:21:4:309. [DOI] [PubMed] [Google Scholar]
- 40.Smyth JM, Ockenfels MC, Gorin AA, Catley D, Porter LS, Kirschbaum C, Hellhammer DH, Stone AA. Individual differences in the diurnal cycle of cortisol. Psychoneuroendocrinol. 1997;22:89–105. doi: 10.1016/s0306-4530(96)00039-x. [DOI] [PubMed] [Google Scholar]
- 41.Weinrib AZ, Sephton S, Degeest K, Penedo F, Bender D, Zimmerman B, Kirschbaum C, Sood AK, Lubaroff DM, Lutgendorf SK. Diurnal cortisol dysregulation, functional disability, and depression in women with ovarian cancer. Cancer. 2010;116:4410–4419. doi: 10.1002/cncr.25299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Cohen L, Cole SW, Sood AK, Prinsloo S, Kirschbaum C, Arevalo JMG, Jennings NB, Scott S, Vence L, Wei Q, Kentor D, Radvanyi L, Tannir N, Jonasch E, Tamboli P, Pisters L. Depressive symptoms and cortisol rhythmicity predict survival in patients with renal cell carcinoma: role of inflammatory signaling. PLoS ONE. 2012;7 doi: 10.1371/journal.pone.0042324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Spiegel D, Giese-Davis J, Taylor CB, Kraemer H. Stress sensitivity in metastatic breast cancer: analysis of hypothalamic-pituitary-adrenal axis function. Psychoneuroendocrinol. 2006;31:1231–1244. doi: 10.1016/j.psyneuen.2006.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]