

Abstract
Animal groups in nature often display an enhanced collective information-processing capacity. It has been speculated that natural selection will tune this response to be optimal, ensuring that the group is reactive while also being robust to noise. Here, we show that this is unlikely to be the case. By using a simple model of decision-making in a dynamic environment, we find that when individuals behave rationally and are subject to selection based on their accuracy, optimality of collective decision-making is not attained. Instead, individuals overly rely on social information and evolve to be too readily influenced by their neighbours. This is due to a classic evolutionary conflict between individual and collective interest. The result is a sub-optimal system that is poised on the cusp of total unresponsiveness. Individuals in the evolved group exhibit delayed reactions to changes in the environment, before responding with rapid, socially reinforced transitions, reminiscent of familiar human and animal social systems (markets, stampedes, fashions, etc.). Our results demonstrate that behaviour of this type may not be pathological, but instead could represent an evolutionary attractor for such collective systems.
Keywords: collective behaviour, evolution, social information
1. Introduction
Social influence is a powerful force in nature and society. In many contexts, individuals gain an advantage by observing and then copying the actions of others [1–3]. The result of this behaviour can be beneficial for all group members; studies of collective behaviour in humans and animals have shown that the use of social information can dampen individual errors and lead to greater decision accuracy [4–7], and may also result in an emergent collective intelligence [8,9]. While there are benefits to social information use, there can also be downsides [10–13]. Although interaction can lead to enhanced information processing [14] and the spreading of novel technologies [15], it may also lead to a lack of responsiveness to changing environments [16] and an over-reliance on the behaviour of others. When individuals devalue their own personal information in favour of imitating the actions, or opinions, of others, this is termed an information cascade [10]. For example, a lack of individual autonomy has been blamed for disasters such as the Challenger shuttle accident [17] and the 2008 financial collapse [18], while in a more commonplace setting Faria et al. [19] showed that the use of social information led to increased risk taking in road-crossing pedestrians. In the natural world, experiments have shown animals are also susceptible to information cascades [20,21], causing individuals to undervalue their personal information [22]. Further, simulations suggest that social behaviour may lead to hysteresis, which means that collective movements, such as migration, are hard to recover once they are lost [23].
Given the substantial costs and benefits associated with the use of social information, an important question is whether natural selection will tune individual behaviour to optimize information processing at the collective level. To investigate this question, we employ an individual-based model of information use in the presence of an external, dynamic information source. We assume that the fitness of individuals is determined by the accuracy of their response to this environmental cue.
This deliberately abstract model could represent animals selecting a heading based on environmental cues as in [23], or humans responding to an alarm signal as in [24]. Historically, models of this type have been termed ‘binary choice with externalities' [25] and have been applied to socially influenced decisions to wear safety equipment in sports [25] or to adopt a particular currency [26]. For animal groups, this form of model may be applied to decisions, for example, relating to movement in response to predators, or larger scale movements associated with migrations. In the latter case, there is substantial evidence to suggest that both the decision of whether to migrate or not [27] and the accuracy of migration [28,29] are affected by social interaction.
Individuals are able to detect the external (global) information source, which we denote G(t); however, detection is not perfect, thus the actions of others may be used as an additional source of information. Individuals respond to the environment with a binary response variable 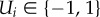 and we define individual i to be correct if
and we define individual i to be correct if
| 1.1 |
Decisions are made based on a personal estimate gi(t) and the observed opinions of individuals in a local neighbourhood, 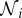 . To model the imperfect environmental detection, the evolution of the personal estimate, gi(t), follows an Ornstein–Uhlenbeck process of the form
. To model the imperfect environmental detection, the evolution of the personal estimate, gi(t), follows an Ornstein–Uhlenbeck process of the form
| 1.2 |
so that individuals make an estimate of the true signal and the quality of this estimate is improved by increasing ωg and deteriorates as the level of noise, σ, increases. We note that this reduces all environmental information to a single dimension, and individuals have no scope for specialization. Functional diversity within groups, as in [30], is therefore precluded.
Social information is contained in a binary vector of the states of individuals within the interaction neighbourhood, 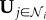 . It should be noted that this vector consists of the observed responses of neighbours, who themselves may be using social information. This is in contrast to the classic ‘wisdom of crowds' model in which independent individual estimates are aggregated [31,32].
. It should be noted that this vector consists of the observed responses of neighbours, who themselves may be using social information. This is in contrast to the classic ‘wisdom of crowds' model in which independent individual estimates are aggregated [31,32].
By employing optimal decision theory [33,34], we next determine the appropriate response of individual i to the total information received, 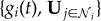 , subject to a single evolvable parameter ωs, which may be interpreted as the level of confidence an individual has that its neighbours are correct.
, subject to a single evolvable parameter ωs, which may be interpreted as the level of confidence an individual has that its neighbours are correct.
2. Optimal individual decision-making
Following the approach of Nitzan & Paroush [33], Pérez-Escudero & de Polavieja [34] and Perreault et al. [35], we determine a weighting of social and personal information by noting that, for each individual i, an optimal strategy is one for which,
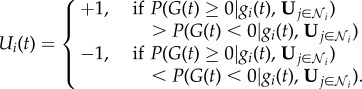 |
2.1 |
In words this means that individual i should set Ui = +1 if, given all available information, it is most likely that the true state of the environment is greater than zero. (As the state G(t) = 0 has Lebesgue measure zero, we arbitrarily assign the optimal response to this state as Ui = 1.)
By applying Bayes' theorem (and for clarity omitting the explicit time dependence), we find
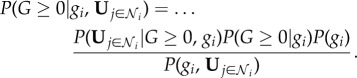 |
2.2 |
Therefore, by using equation (2.2) and the equivalent expression for 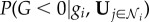 , equation (2.1) may be rearranged so that the state G ≥ 0 is the most probable state if
, equation (2.1) may be rearranged so that the state G ≥ 0 is the most probable state if
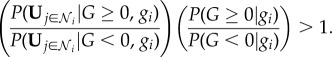 |
2.3 |
This expression is essentially a rewriting of equation (2.1), with the role of social and personal information appearing separately. The likelihood that the environment is in the state G ≥ 0, given the personal information gi may be calculated from the properties of the Ornstein–Uhlenbeck process, as
| 2.4 |
This represents the stationary solution of the process, hence is valid when G(t) varies slowly with respect to the response time defined by ωg. (While this assumption does not hold when the environment is rapidly alternating, simulations shown in the electronic supplementary material, figure S3, demonstrate qualitatively equivalent results.) Again, employing Bayes' theorem, we attain
| 2.5 |
As 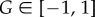 , we have
, we have
| 2.6 |
and, similarly,
| 2.7 |
Combining equations (2.4), (2.6) and (2.7) and cancelling constants provides
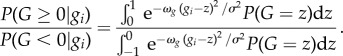 |
2.8 |
We next consider the problem of weighting the social information. As the accuracy of neighbours depends on their own strategies, the question of how to weight this information is analytically intractable. However, we are able to reduce the problem to a single parameter, denoted ωs, that represents the assumed probability an individual has that a randomly selected neighbour is correct. If ωs = 0.5, neighbours are believed to have an even chance of being correct, and thus provide no additional information, while if ωs = 1, neighbours will always be followed and the social information is assumed to be infallible.
We stress that ωs does not represent the true probability an individual is correct but rather a belief about that probability that can be translated into a decision rule. Further, while this analysis represents a formal interpretation of the social confidence level, in terms of a probability, this parameter is free to evolve and may give too much, or too little, weight to observed opinions. The only restriction placed on individuals is that the motivations of neighbours are unknown, meaning a scenario where two neighbours share an opinion based on their independent personal information is indistinguishable from a scenario where one neighbour has copied the other. Effectively, this assumption restricts the domain of our model to systems in which honest communication of personal information, or confidence levels, does not occur.
Formally, individual i assumes
| 2.9 |
and
| 2.10 |
The estimated probability (assuming independence) of observing the vector of responses of neighbours, 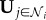 , if the global information G ≥ 0 is then
, if the global information G ≥ 0 is then
| 2.11 |
while if G < 0 then this is
| 2.12 |
Combining these two equations, we get
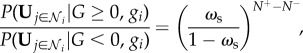 |
2.13 |
where N + and N− are the number of neighbours for which Uj = 1 and Uj =−1, respectively. Finally, we may substitute equations (2.8) and (2.13) into equation (2.3) to attain the optimal individual decision-making strategy, given a social confidence level ωs, as
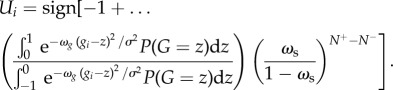 |
2.14 |
3. Numerical simulations
We next simulate an individual-based model that incorporates the decision rule previously described. For these simulations, we first need to define the functional form of G(t) and prescribe an interaction network over which social observations are made. As a first approximation, we use a mean-field model for the interaction network. This assumes that the population is well mixed and observations are drawn at random at each time step. The advantage of this approach is that the social network is simplified to a single parameter k, that defines the size of the interaction neighbourhood, such that 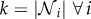 . However, we note that our results are not dependent on this assumption, and simulations incorporating various structured interaction networks are shown in the electronic supplementary material, figures S5–S7.
. However, we note that our results are not dependent on this assumption, and simulations incorporating various structured interaction networks are shown in the electronic supplementary material, figures S5–S7.
To model the environmental information, we employ an alternating, periodic function for G(t). By using a deterministic function, we are able to control both the nature of transitions between environmental states and the length of the time interval between transitions. While this simplifies our analytical calculations, similar results are attained in the case of more realistic, stochastic environments (see the electronic supplementary material, figure S4, for details). For G(t), we use both a periodic triangle wave and a square wave input. Mathematically, these are defined as
| 3.1 |
for the triangle wave, and
| 3.2 |
for the square wave, where Θ is the Heaviside step function, and TE determines the time scale of the environmental variation in both cases. The shape of these functions may be viewed in the time series shown in figure 1a,b (dashed lines).
Figure 1.

Numerical simulations. (a) Time series of average response to the environment 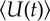 (solid lines) and G(t) (dashed line) for triangle wave. Parameter values are k = 4, N = 200, ωg = 0.2, σ = 1 and TE = 100. Values used for ωs are shown as triangles with corresponding colour in (c). Panel (b) as (a) for square wave and parameter values k = 8, N = 100, ωg = 0.1, σ = 1 and TE = 500. (c) Accuracy as a function of social information weighting for parameters as in (a) (triangle points, grey line) and (b) (square points, black line). Accuracy is defined as the time averaged fraction of individuals for which U = sign(G(t)). The dashed lines illustrate the performance when k independent observations are made directly by individuals (i.e. the performance according to the ‘wisdom of crowds' hypothesis).
(solid lines) and G(t) (dashed line) for triangle wave. Parameter values are k = 4, N = 200, ωg = 0.2, σ = 1 and TE = 100. Values used for ωs are shown as triangles with corresponding colour in (c). Panel (b) as (a) for square wave and parameter values k = 8, N = 100, ωg = 0.1, σ = 1 and TE = 500. (c) Accuracy as a function of social information weighting for parameters as in (a) (triangle points, grey line) and (b) (square points, black line). Accuracy is defined as the time averaged fraction of individuals for which U = sign(G(t)). The dashed lines illustrate the performance when k independent observations are made directly by individuals (i.e. the performance according to the ‘wisdom of crowds' hypothesis).
These two expressions for G(t) provide two optimal rules for equation (2.14) by determining the form of P(G = z). If the triangle wave is used then
| 3.3 |
whereas for the square wave,
| 3.4 |
where δ is the Dirac delta function. By substituting these functions into equation (2.14), an optimal decision rule can be obtained that is a function only of gi(t) (personal information) and 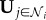 (social information). In figure 1, results from simulations of the model are shown. The beneficial aspects of social information can be clearly observed as ωs increases. However, over-reliance on this information, represented by larger values of ωs, results in steep declines in performance as the group becomes rapidly less responsive.
(social information). In figure 1, results from simulations of the model are shown. The beneficial aspects of social information can be clearly observed as ωs increases. However, over-reliance on this information, represented by larger values of ωs, results in steep declines in performance as the group becomes rapidly less responsive.
The results of the simulations, shown in figure 1, demonstrate the advantages, and disadvantages, of using social information. By following the behaviour of others, individuals increase their accuracy. This increase in accuracy goes beyond the classic ‘wisdom of crowds' concept, whereby the variance in the fraction of correct individuals observed over independent trials converges to zero, an effect most clearly expounded by Condorcet's jury theorem [31]. To enable a comparison to this effect, accuracy is calculated as if independent estimates were aggregated and shown in figure 1c. For these results, neighbours convey their own personal estimate of the cue (gi(t)). Equivalently, this may be considered as the scenario where a single individual makes k independent observations of the cue and then takes the average of those observations.
In the full simulations (solid lines of figure 1), observations of neighbours do not equate to independent estimates, as neighbours are also making use of social information. This can result in a far superior performance, as individuals are effectively accessing a greater number of estimates via the information flow through the social network. However, as the weighting given to social information increases, the benefit is rapidly lost due to high levels of correlation [36,37]. The lack of independence in individual behaviour leads to a sharp decrease in performance, until the system becomes locked into a single response and accuracy is 0.5 (equal to a random chance of being correct).
4. Evolved strategies and collective unresponsiveness
In order to determine where, in parameter space, we should expect to find natural collective systems, we introduce an evolutionary component into our simulations by allowing the parameter ωs to evolve. A selection algorithm [38] is used that ensures an individual's expected number of offspring is proportional to the accuracy of its response to the environment. Offspring inherit the characteristics of the parent individual (ωs) with a small Gaussian mutation. As social interactions occur at random within the population, our simulations preclude the evolution of any altruistic traits that may benefit neighbouring individuals while incurring a personal fitness cost. Hence, our model is focused purely on selection at the individual level.
In figure 2, results from the evolutionary simulations are shown. We clearly observe a substantial distinction between the evolved weighting of social information and the value that gives optimal collective performance. We find natural selection drives the trait beyond the optimal level and moves the collective system towards an unresponsive state. In these simulations we employ homogeneous populations as the initial condition with ωs = 0.5 for all individuals (no social information is used). Results are robust to these assumptions as shown in the electronic supplementary material, figures S8 and S9.
Figure 2.

Evolutionary simulations. (a) Evolution of accuracy for triangle wave (red) and square wave (blue). Parameter values are, triangle wave: k = 8, N = 50, ωg = 0.2, σ = 1, TE = 1000; square wave: k = 4, N = 200, ωg = 0.1, σ = 1, TE = 500. Generations consist of simulations lasting for 100 × TE time units. (b,c) Average response to a change in the environment as evolution proceeds. Colours match to the parameter values and stage of evolution shown in (a). Units are rescaled by TE, with t = 0 corresponding to the point at which G(t) switches from negative to positive. (d) Performance and evolved state. Lines represent the performance for a homogeneous population, while points show the actual ESS value to which the system evolves.
In order to understand the mechanisms underlying the simulation results, we investigate the dynamics of our model within an evolutionary invasion framework [39]. To do so, we must first make some simplifying assumptions regarding model properties. Namely, we assume that the population is large, the number of social observations each individual makes is also large, and that the environmental cue is defined by the step function of equation (3.2). Given these restrictions, we attain an equation that governs the dynamics of the population when the external cue is in the state G(t) = 1 as
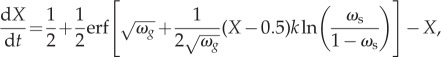 |
4.1 |
where X is the fraction of individuals that are responding correctly to the environment and, for simplicity, we have rescaled the personal information parameter, ωg, by a factor of σ2 to reduce the number of parameters. (For a detailed derivation of this equation, see the electronic supplementary material.)
As the environment alternates between different states on a characteristic time scale, the expected accuracy may be calculated from the performance over a single, representative, time interval. Further, if we expect there to be low accuracy immediately following a transition, we may approximate this initial state as X = 0, i.e. there is zero accuracy within the group at time t = 0. The time-averaged accuracy may then be written as
| 4.2 |
where X(t) is defined as the solution to equation (4.1) with initial condition X(0) = 0. In the limiting case of large populations, and many social observations of the population, a single individual with social weighting of 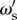 has a probability of being correct that is dependent on the fraction of individuals in the population that are correct, according to
has a probability of being correct that is dependent on the fraction of individuals in the population that are correct, according to
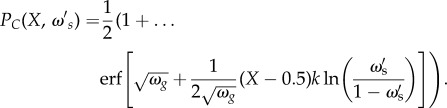 |
4.3 |
The expected accuracy, 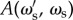 , of this mutant strategy over time is then
, of this mutant strategy over time is then
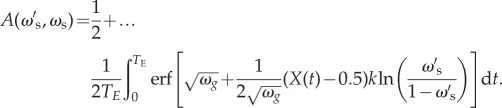 |
4.4 |
The evolutionarily stable strategy is found by locating the value of the resident population ωs around which small mutations will have a lower accuracy [40,41],
| 4.5 |
Solutions to equations (4.2) and (4.5) may be found numerically and these are shown in figure 3 alongside the corresponding full simulation. To verify that the singular strategy is both evolutionarily and convergence stable, second derivatives are taken with respect to the mutant strategy (evolutionary stability) and the resident strategy (convergence stability). These derivatives are numerically evaluated at the location of the singular strategy and it is found that the value of ωs that satisfies equation (4.5) is both evolutionarily stable (no branching occurs) and convergence stable. (See the electronic supplementary material for further details.)
Figure 3.

Evolutionary analysis. (a) Average accuracy of an individual-based simulation of evolution. The square wave is employed with parameter values: k = 80, N = 500, ωg = 0.1, σ = 1 and TE = 200. A Gaussian mutation rate of 1 × 10−6 was used; this level of mutation leads to oscillations between high-accuracy generations and total unresponsiveness. (b) Average accuracy of an individual-based simulation with a mutation rate of 1 × 10−7. Evolution converges to a stable value (average over final 100 generations is shown in red). (c) Comparison of simulation and analytical solutions. The grey line shows accuracy for an individual-based simulation with a fixed weighting of social information (ωs). The red line indicates evolved accuracy from simulation. The black line shows the performance calculated from equation (4.2), and the blue point represents the analytical ESS calculated from equation (4.5).
While this analysis confirms the results of the individual-based simulations, it provides little insight into the underlying mechanisms. To gain a more heuristic understanding of the evolutionary process, we coarse-grain the full dynamics of equation (4.1) and consider a two-stage process that describes the population response to a change in the environment.
The first stage consists of the period of time immediately following an environmental transition, before the population has responded. This corresponds to the situation when most individuals are incorrect as X(t) < 0.5 and G(t) = 1. The length of this phase is the response time, TR, of the collective system and may be calculated as
| 4.6 |
The second stage consists of the rapid transition to the steady-state solution of equation (4.1) and lasts for a period of time TE−TR, i.e. until the next environmental switch. We may then approximate the full ordinary differential equation as a process in which the population switches from a low proportion of individuals in the correct state 0 < X(t) < 0.5, which we denote XL and approximate as XL = 0.25, to the high-accuracy state X∞, which is the steady-state solution of equation (4.1). The switch occurs after time TR, and we use the full dynamics to calculate the length of this period.
As shown in equation (4.3), an individual with social weighting 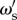 has a probability of being correct that is dependent on the accuracy of other individuals in the population. In this reduced framework, the expected long-term accuracy of an individual with social weighting
has a probability of being correct that is dependent on the accuracy of other individuals in the population. In this reduced framework, the expected long-term accuracy of an individual with social weighting 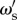 within a population of individuals with social weighting ωs is
within a population of individuals with social weighting ωs is
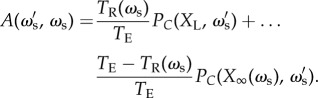 |
4.7 |
The usefulness of this expression lies in the division of the impact of individual and collective properties on accuracy. The steady-state group accuracy, X∞, and the length of the response time are controlled by the resident population parameter ωs, whereas the rare strategy 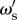 governs the individual-level accuracy in each phase.
governs the individual-level accuracy in each phase.
Again the evolutionarily stable strategy is found by solving the equation
| 4.8 |
Introducing the notation,
| 4.9 |
it can be shown that the evolutionarily stable strategy (ESS) is reached when
| 4.10 |
This expression may be interpreted as a balancing of the gain in accuracy attained by increasing sociality when X is high, with the loss in accuracy when X is low, weighted according to the respective length of time of each phase. The equation is written in this form so that the time spent prior to a collective response to the environment (LHS) may be related to the ratio of accuracy changes at the individual level (RHS).
Next, we follow a similar approach to find the optimal value of the resident population. To find this collectively optimal accuracy, it is necessary to solve
| 4.11 |
i.e. find the value of ωs that maximizes accuracy considering its impact on both individual-level decisions and the aggregate properties of the system. Neglecting the effect of the resident population strategy on the value of X∞, and only including the dominant effect on the lack of responsiveness, equation (4.11) may be rearranged to give
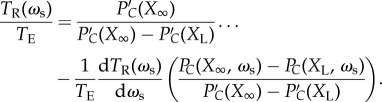 |
4.12 |
By comparing equations (4.10) and (4.12), we observe the key difference between the optimal and evolved solutions; to achieve the optimal level of information processing, the increase in the collective inertia of the system must be considered. This increase in response time with respect to the social weighting manifests itself in the second term on the RHS of equation (4.12). As this term is absent from the ESS, the population will always evolve towards unresponsiveness, moving beyond the optimal value of ωs and stabilizing only when the social information is sufficiently degraded.
To visualize the role of each term in equations (4.10) and (4.12), their values have been plotted in figure 4. This geometric view of the equations illustrates the influence of each term and how they combine to create a sub-optimal collective response. It is worth noting that the rapid increase in the delay time as sociality increases leads to a precipitous drop in responsiveness. This translates into high variation in accuracy across generations caused by fluctuations around the ESS value of ωs. As accuracy declines so steeply in the vicinity of the ESS value, when mutations are high, the system frequently becomes completely unresponsive, as demonstrated by the repeated low accuracy states shown in figure 3a.
Figure 4.
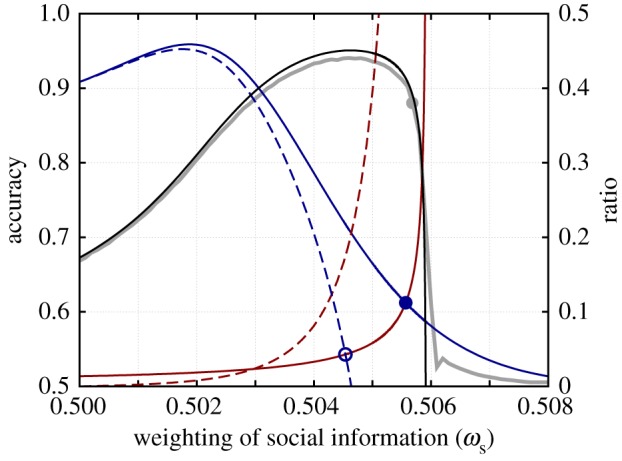
Interpreting evolutionary stability and optimal performance. The grey line shows the accuracy as a function of ωs from the individual-based simulations (k = 80, N = 500, ωg = 0.1, σ = 1, TE = 200), the grey point shows the evolved average accuracy. The black line shows the accuracy calculated from the reduced model of equation (4.7). The solid red line shows the LHS of equation (4.10) 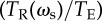 and the solid blue line shows the RHS of equation (4.10), where they intersect corresponds to the ESS value of the social weighting. The red dashed line shows the value of
and the solid blue line shows the RHS of equation (4.10), where they intersect corresponds to the ESS value of the social weighting. The red dashed line shows the value of 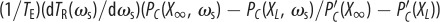 . Subtracting this from the solid blue line as in equation (4.12) and finding the intersection with
. Subtracting this from the solid blue line as in equation (4.12) and finding the intersection with 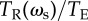 (solid red line) gives the optimal weighting of social information.
(solid red line) gives the optimal weighting of social information.
5. Discussion
Over-reliance on social information is a common and frequently observed behaviour in many species. In humans, this is manifested in many well-studied phenomena such as rapid technology adoption and lock-in [42], or the boom-and-bust cycles of financial markets [11,43]. Non-human animals may also devalue their personal information in favour of copying others [44], resulting in sub-optimal collective behaviour even when individuals are aware of more suitable alternatives [45].
Several previous studies have investigated this effect in the context of human decision-making, notably [10,46] (but also see [47–49], and [50] for a review). These models assume that decision-making is sequential, previous decisions are observed and agents behave rationally given the information that is available. Under these assumptions, information cascades will occur as the number of individuals increases. Fundamentally, this effect arises because individuals are unable to recognize the decisions of others that are based on copying.
A similar mechanism underlies the results of our model, however in our framework individuals are allowed to employ any level of confidence in social information. As strategies evolve based on their performance, there is the potential for individuals to avoid over-dependence on social information. Our results show that while high levels of accuracy are initially attained, natural selection continually drives the population beyond the optimal performance levels, due to the conflict between individual and collective interest [51].
By employing optimal decision theory and evolutionary invasion analysis, we have shown that over-reliance on social information evolves in dynamic environments. As individuals in a well-functioning group provide an excellent source of information, natural selection leads to over-reliance on social cues, even to the point of unresponsiveness. When mutations are large but infrequent, highly conformist individuals take over the population and lead to an almost complete failure in information processing. Once this has occurred, less social individuals are selected for, and higher accuracy levels are re-established. However, in the limit of vanishing mutation rates, analytical conditions show that information processing will evolve to be sub-optimal, as the disadvantages of over-confidence in social information are felt at the collective level (groups become unable to escape from dominant, widely held opinions).
The sharp deterioration in accuracy observed as social weightings are increased is in agreement with other studies of collective movement. For example, in a recent work, Codling & Bode [52] showed that when groups use social information, accuracy is improved, and surprisingly a large amount of social weighting was observed to be optimal. However, in agreement with the results presented here, steep declines in performance occurred as social influence increased. In this context, our findings predict that the high-accuracy navigating groups of Codling & Bode [52] will be unstable, and animal groups in nature are more likely to be found with values of social weighting that place them close to the collapse of accurate motion.
Investigating the ultimate drivers of social behaviour is not feasible in most species; however quorum-sensing bacteria [53] present a promising avenue for future tests of our theory. Bacteria respond to both social and environmental information and may be evolved in a laboratory setting. For bacteria responding to a dynamic environment, we predict that over-reliance on social information and an increased unresponsiveness will evolve over time.
The results we present illustrate that evolution may not lead to effective information-processing groups, even in the absence of any cost to acquiring personal information. This has implications in a variety of contexts. Notably, our findings challenge the notion that we should expect animal groups to be ‘tuned’ to respond optimally to environmental information. Instead, collective inertia should be considered the default, and we predict that social species will display less behavioural plasticity and respond more slowly to changes in their environment. In the context of human interaction, our results suggest that when social information is available, individuals will over use it [54] and this should be considered when attempting to engineer effective groups or organizations.
Supplementary Material
Supplementary Material
Supplementary Material
Funding statement
This research was supported by Army Research Office grant no. W911NG-11–1–0385, National Science Foundation grant no. PHY-0848755, Office of Naval Research Award N00014–09–1–1074, Human Frontier Science Project grant no. RGP0065/2012 and NSF EAGER grant no. IOS-1251585 to I.D.C. T.L. was supported by the FIRB project (RBID08PP3J), the Fondation Sciences Mathématiques de Paris (FSMP) and by a public grant overseen by the French National Research Agency (ANR-10-LABX-0098).
References
- 1.Dall SR, Giraldeau L-A, Olsson O, McNamara JM, Stephens DW. 2005. Information and its use by animals in evolutionary ecology. Trends Ecol. Evol. 20, 187–193. ( 10.1016/j.tree.2005.01.010) [DOI] [PubMed] [Google Scholar]
- 2.Bazazi S, Ioannou CC, Simpson SJ, Sword GA, Torney CJ, Lorch PD, Couzin ID. 2010. The social context of cannibalism in migratory bands of the Mormon cricket. PLoS ONE 5, e15118 ( 10.1371/journal.pone.0015118) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Clark CW, Mangel M. 1984. Foraging and flocking strategies: information in an uncertain environment. Am. Nat. 123, 626–641. ( 10.1086/284228) [DOI] [Google Scholar]
- 4.Simons A. 2004. Many wrongs: the advantage of group navigation. Trends Ecol. Evol. 19, 453–455. ( 10.1016/j.tree.2004.07.001) [DOI] [PubMed] [Google Scholar]
- 5.Codling EA, Pitchford JW, Simpson SD. 2007. Group navigation and the ‘many-wrongs principle’ in models of animal movement. Ecology 88, 1864–1870. ( 10.1890/06-0854.1) [DOI] [PubMed] [Google Scholar]
- 6.Faria JJ, Codling EA, Dyer JR, Trillmich F, Krause J. 2009. Navigation in human crowds; testing the many-wrongs principle. Anim. Behav. 78, 587–591. ( 10.1016/j.anbehav.2009.05.019) [DOI] [Google Scholar]
- 7.Torney CJ, Levin SA, Couzin ID. 2013. Decision accuracy and the role of spatial interaction in opinion dynamics. J. Stat. Phys. 151, 203–217. ( 10.1007/s10955-013-0700-5) [DOI] [Google Scholar]
- 8.Woolley AW, Chabris CF, Pentland A, Hashmi N, Malone TW. 2010. Evidence for a collective intelligence factor in the performance of human groups. Science 330, 686–688. ( 10.1126/science.1193147) [DOI] [PubMed] [Google Scholar]
- 9.Berdahl A, Torney CJ, Ioannou CC, Faria JJ, Couzin ID. 2013. Emergent sensing of complex environments by mobile animal groups. Science 339, 574–576. ( 10.1126/science.1225883) [DOI] [PubMed] [Google Scholar]
- 10.Banerjee AV. 1992. A simple model of herd behavior. Q. J. Econ. 107, 797–817. ( 10.2307/2118364) [DOI] [Google Scholar]
- 11.Lux T. 1995. Herd behaviour, bubbles and crashes. Econ. J. 105, 881–896. ( 10.2307/2235156) [DOI] [Google Scholar]
- 12.Giraldeau L-A, Valone TJ, Templeton JJ. 2002. Potential disadvantages of using socially acquired information. Phil. Trans. R. Soc. Lond. B 357, 1559–1566. ( 10.1098/rstb.2002.1065) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Rieucau G, Giraldeau L-A. 2011. Exploring the costs and benefits of social information use: an appraisal of current experimental evidence. Phil. Trans. R. Soc. B 366, 949–957. ( 10.1098/rstb.2010.0325) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Couzin ID. 2009. Collective cognition in animal groups. Trends Cogn. Sci. 13, 36–43. ( 10.1016/j.tics.2008.10.002) [DOI] [PubMed] [Google Scholar]
- 15.Walden EA, Browne GJ. 2009. Sequential adoption theory: A theory for understanding herding behavior in early adoption of novel technologies. J. Assoc. Inform. Syst. 10, 31–62. [Google Scholar]
- 16.Feldman MW, Aoki K, Kumm J. 1996. Individual versus social learning: evolutionary analysis in a fluctuating environment. Anthropol. Sci. 104, 209–231. ( 10.1537/ase.104.209) [DOI] [Google Scholar]
- 17.Esser JK, Lindoerfer JS. 1989. Groupthink and the space shuttle challenger accident: toward a quantitative case analysis. J. Behav. Decis. Mak. 2, 167–177. ( 10.1002/bdm.3960020304) [DOI] [Google Scholar]
- 18.Tuckett D. 2009. Addressing the psychology of financial markets. Economics 3, 1–21. ( 10.5018/economics-ejournal.ja.2009-40) [DOI] [Google Scholar]
- 19.Faria JJ, Krause S, Krause J. 2010. Collective behavior in road crossing pedestrians: the role of social information. Behav. Ecol. 21, 1236–1242. ( 10.1093/beheco/arq141) [DOI] [Google Scholar]
- 20.Laland KN, Williams K. 1998. Social transmission of maladaptive information in the guppy. Behav. Ecol. 9, 493–499. ( 10.1093/beheco/9.5.493) [DOI] [Google Scholar]
- 21.Sasaki T, Granovskiy B, Mann RP, Sumpter DJ, Pratt SC. 2013. Ant colonies outperform individuals when a sensory discrimination task is difficult but not when it is easy. Proc. Natl Acad. Sci. USA 110, 13 769–13 773. ( 10.1073/pnas.1304917110) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rieucau G, Giraldeau L-A. 2009. Persuasive companions can be wrong: the use of misleading social information in nutmeg mannikins. Behav. Ecol. 20, 1217–1222. ( 10.1093/beheco/arp121) [DOI] [Google Scholar]
- 23.Guttal V, Couzin ID. 2010. Social interactions, information use, and the evolution of collective migration. Proc. Natl Acad. Sci. USA 107, 16 172–16 177. ( 10.1073/pnas.1006874107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bassett DS, Alderson DL, Carlson JM. 2012. Collective decision dynamics in the presence of external drivers. Phys. Rev. E 86, 036105 ( 10.1103/PhysRevE.86.036105) [DOI] [PubMed] [Google Scholar]
- 25.Schelling TC. 1973. Hockey helmets, concealed weapons, and daylight saving: a study of binary choices with externalities. J. Confl. Resolution 17, 381–428. ( 10.1177/002200277301700302) [DOI] [Google Scholar]
- 26.Young HP. 2001. Individual strategy and social structure: an evolutionary theory of institutions. Princeton, NJ: Princeton University Press. [Google Scholar]
- 27.Dumond M, Lee DS. 2013. Dolphin and union caribou herd status and trend. Arctic 66, 329 ( 10.14430/arctic4311) [DOI] [Google Scholar]
- 28.Berdahl A, Westley PA, Levin SA, Couzin ID, Quinn TP. In press A collective navigation hypothesis for homeward migration in anadromous salmonids. Fish Fish. ( 10.1111/faf.12084) [DOI] [Google Scholar]
- 29.Dittman A, Quinn T. 1996. Homing in Pacific salmon: mechanisms and ecological basis. J. Exp. Biol. 199, 83. [DOI] [PubMed] [Google Scholar]
- 30.Hong L, Page SE. 2004. Groups of diverse problem solvers can outperform groups of high-ability problem solvers. Proc. Natl Acad. Sci. 101, 16 385–16 389. ( 10.1073/pnas.0403723101) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Condorcet M. 1976. Reprinted in Condorcet: selected writings, Keith Michael Baker, 33rd edn Indianapolis, IN: Bobbs-Merrill. [Google Scholar]
- 32.Galton F. 1907. Vox Populi. Nature 75, 450–451. ( 10.1038/075450a0) [DOI] [Google Scholar]
- 33.Nitzan S, Paroush J. 1982. Optimal decision rules in uncertain dichotomous choice situations. Int. Econ. Rev. 23, 289 ( 10.2307/2526438) [DOI] [Google Scholar]
- 34.Pérez-Escudero A, de Polavieja GG. 2011. Collective animal behavior from Bayesian estimation and probability matching. PLoS Comp. Biol. 7, e1002282 ( 10.1371/journal.pcbi.1002282) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Perreault C, Moya C, Boyd R. 2012. A Bayesian approach to the evolution of social learning. Evol. Hum. Behav. 33, 449–459. ( 10.1016/j.evolhumbehav.2011.12.007) [DOI] [Google Scholar]
- 36.Ladha KK. 1992. The Condorcet Jury theorem, free speech, and correlated votes. Am. J. Political Sci. 36, 617–634. ( 10.2307/2111584) [DOI] [Google Scholar]
- 37.Kao AB, Couzin ID. 2014. Decision accuracy in complex environments is often maximized by small group sizes. Proc. R. Soc. B 281, 20133305 ( 10.1098/rspb.2013.3305) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bäck T. 1996. Evolutionary algorithms in theory and practice. Oxford, UK: Oxford university press. [Google Scholar]
- 39.Dercole F, Rinaldi S. 2008. Analysis of evolutionary processes: the adaptive dynamics approach and its applications. Princeton, NJ: Princeton University Press. [Google Scholar]
- 40.Eshel I. 1983. Evolutionary and continuous stability. J. Theor. Biol. 103, 99–111. ( 10.1016/0022-5193(83)90201-1) [DOI] [PubMed] [Google Scholar]
- 41.Hofbauer J, Sigmund K. 1990. Adaptive dynamics and evolutionary stability. Appl. Math. Lett. 3, 75–79. ( 10.1016/0893-9659(90)90051-C) [DOI] [Google Scholar]
- 42.Arthur WB. 1989. Competing technologies, increasing returns, and lock-in by historical events. Econ. J. 99, 116 ( 10.2307/2234208) [DOI] [Google Scholar]
- 43.Pan W, Altshuler Y, Pentland AS. 2012. Decoding social influence and the wisdom of the crowd in financial trading network. In Privacy, security, risk and trust (PASSAT), 2012 International Conference on Social Computing (SocialCom), pp. 203–209. Piscataway, NJ: IEEE. [Google Scholar]
- 44.Howell D. 1979. Flock foraging in nectar-feeding bats: advantages to the bats and to the host plants. Am. Nat. 114, 23–49. ( 10.1086/283452) [DOI] [Google Scholar]
- 45.Bates L, Chappell J. 2002. Inhibition of optimal behavior by social transmission in the guppy depends on shoaling. Behav. Ecol. 13, 827–831. ( 10.1093/beheco/13.6.827) [DOI] [Google Scholar]
- 46.Bikhchandani S, Hirshleifer D, Welch I. 1992. A theory of fads, fashion, custom, and cultural change as informational cascades. J. Political Econ. 100, 992–1026. ( 10.1086/261849) [DOI] [Google Scholar]
- 47.Smith L, Sørensen P. 2000. Pathological outcomes of observational learning. Econometrica 68, 371–398. ( 10.1111/1468-0262.00113) [DOI] [Google Scholar]
- 48.Çelen B, Kariv S. 2004. Observational learning under imperfect information. Games Econ. Behav. 47, 72–86. ( 10.1016/S0899-8256(03)00179-9) [DOI] [Google Scholar]
- 49.Ellison G, Fudenberg D. 1993. Rules of thumb for social learning. J. Political Econ. 101, 612–643. ( 10.1086/261890) [DOI] [Google Scholar]
- 50.Easley D, Kleinberg J. 2010. Networks, crowds, and markets. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 51.Hardin G. 1968. The tragedy of the commons. Science 162, 1243–1248. ( 10.1126/science.162.3859.1243) [DOI] [PubMed] [Google Scholar]
- 52.Codling EA, Bode NW. 2014. Copycat dynamics in leaderless animal group navigation. Mov. Ecol. 2, 11 ( 10.1186/2051-3933-2-11) [DOI] [Google Scholar]
- 53.Miller MB, Bassler BL. 2001. Annu. Rev. Microbiol. 55, 165–199. ( 10.1146/annurev.micro.55.1.165) [DOI] [PubMed] [Google Scholar]
- 54.Saavedra S, Hagerty K, Uzzi B. 2011. Synchronicity, instant messaging, and performance among financial traders. Proc. Natl Acad. Sci. USA 108, 5296–5301. ( 10.1073/pnas.1018462108) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.