

Abstract
AIM: To investigate the role of NF-κB in the pathogenesis of TNBS-induced colitis in rats.
METHODS: Thirty-two healthy adult Sprague-Dawley (SD) rats were randomly divided into four groups of eight each: normal, NS, model I, model II groups in our study. Rat colitis model was established through 2-,4-,6-trinitrobenzene sulfonic acid (TNBS) enema. At the end of four weeks, the macroscopical and histological changes of the colon were examined and mucosa myeloperoxidase (MPO) activities assayed. NF-κB p65 expression was determined by Western blot assessment in cytoplasmic and nuclear extracts of colon tissue, and the expressions of TNF-α and ICAM-1 protein in colon tissue were examined by immunohistochemistry. The relativities between expression of NF-κB p65 and other parameters were analyzed.
RESULTS: TNBS enema resulted in pronounced pathological changes of colonic mucosa in model II group (macroscopic and histological injury indices 6.25±1.39 and 6.24±1.04, respectively), which were in accordance with the significantly elevated MPO activity (1.69±0.11). And the nuclear level of NF-κB and expression of TNF-α, ICAM-1 in rats of model II group were higher than that of normal control (9.7±1.96 vs 1.7±0.15, 84.09±14.52 vs 16.03±6.21, 77.69±8.09 vs 13.41±4.91 P<0.01), Linear correlation analysis revealed that there were strong correlations between the nuclear level of NF-κB and the tissue positive expression of TNF-α and ICAM-1, MPO activities, macroscopical and histological indices in TNBS-induced colitis, respectively (r = 0.8235, 0.8780, 0.8572, 0.9152, 0.8247; P<0.05).
CONCLUSION: NF-κB plays a pivotal role in the pathogenesis of ulcerative colitis, which might account for the up-regulation the expression of TNF-α and ICAM-1.
Keywords: NF-κB, Ulcerative colitis, TNBS
INTRODUCTION
Ulcerative colitis (UC), a major inflammatory bowel disease (IBD), is chronic inflammatory disorder of the gastrointestinal tract. Although the exact etiology and pathogenetic mechanisms remain obscure, dysregulated immune and inflammatory responses have been recognized as involved in the pathogenesis of IBD[1-4]. There is substantial evidence that both proinflammatory cytokines-adhesion molecules and other inflammatory mediators including tumor necrosis factor (TNF-α) and intercellular adhesion molecule-1(ICAM-1) exhibit a key role in the inflammatory process[5-7]. Their increased production in the intestinal mucosa is thought to be an important factor in the pathophysiology of intestinal inflammation in UC. However, the pivotal elements in the regulation of the increased inflammatory activity remains unclear.
Thus, the focus of recent researches have been on the identification of signaling pathways and transcription factors that bind to gene promoter regions and govern proinflammatory cytokines and mediators gene transcription in UC. Recent studies have demonstrated that NF-κB is a key transcription factor of lymphocytes and macrophages with important regulatory functions in the immune system and inflammatory processes[8-10]. These functions are at least partially based on its ability to regulate the promoters of a variety of genes whose products, such as cytokines, adhesion molecules and acute phase proteins, are critical for inflammatory processes. And this factor is formed by several different dimers of members of the Rel family, which include NF-κB1 (p50), NF-κB2 (p52), RelA (p65), RelB, and C-Rel[11,12]. The heterodimers composed of the subunits NF-κB1 and RelA are the most frequent and active forms of NF-κB[12]. Characteristically, NF-κB proteins are sequestered in the cytoplasm as a result of retention by a class of inhibitory proteins, referred to as the IκB family[13]. On stimulation, IκBα is phosphorylated and proteolytic degradation takes place rapidly. The free NF-κB is then translocated to the nucleus and bound to decameric DNA sequences and activated transcription of target genes[13,14-17].
In the present study, we investigate whether increased activation of NF-κB is important in TNBS-induced colitis in rats and attempt to demonstrate the regulating role of NF-κB in inflammatory response.
MATERIALS AND METHODS
Animals and experimental protocol
Thirty-two healthy adult Sprague-Dawley (SD) rats of both sexes, weighing 150-200 g, obtained from the Experimental Animal Center of Wuhan University, were housed under specific pathogen-free conditions, and allowed access to standard rat chow and water. Before the experiments, they were acclimatized to the surroundings for one week. The study protocol was in accordance with the guideline for animal research and was approved by the Ethical and Research Committee of the hospital. Rat model of colitis induced with 2-,4-,6-trinitrobenzene sulfonic acid (TNBS, Sigma Corporation) enema is described in the literature[18]. Thirty-two experimental animals were randomly divided into four groups of eight each: normal control group, NS group treated with saline enema, and model I and II groups inflicted with TNBS/40% ethanol enema at doses of 50 and 150 mg/kg, respectively. At the end of four weeks, the animals were killed and the colon samples were collected.
Colon tissues were fixed in 4% paraformaldehyde, dehydrated and paraffin-embedded. Four-micrometer sections were cut transversely and stained with hematoxylin and eosin. Colon macroscopic and histological damage indices were evaluated by an independent pathologist at random as reported previously[19]. At the same time, colon samples from the same sites were also obtained and frozen immediately in liquid nitrogen for subsequent Western blot analysis for NF-κB p65 protein and MPO activity determination.
Extracts and Western blot analysis
Colon samples were snap frozen in liquid nitrogen at the time of removal and later mechanically homogenized in liquid nitrogen. Nuclear and cytoplasmic extracts were prepared by adaptation of previously described techniques through freeze/thaw cycles between crushed ice and liquid nitrogen[20,21]. Cytoplasmic extracts were collected in an aqueous buffer containing 10 mmol/L Hepes (pH7.9), 1.5 mmol/L MgCl2, and 10 mmol/L KCl. Nuclear extracts were prepared by solubilizing the remaining nuclei in a buffer containing 20 mmol/L Hepes (pH7.9), 420 mmol/L NaCl, 1.5 mmol/L MgCl2, 0.2 mmol/L EDTA and 25% glycerol. Both buffers were supplemented with 1 mmol/L dithiothreitol, 0.5 mmol/L phenylmethanesulfonyl fluoride, 1 μg/mL aprotinin, 1 μg/mL pepstatin, 1 mmol/L benzamidine, 1 mmol/L sodium vanadate, and 1 mmol/L NaF. And colon tissues from the same sites were homogenized for the assessment of total tissue concentrations of NF-κB p65 protein. Protein concentration of samples were assessed using the method of Bradford protein assay, and all samples were adjusted to an equal protein content before analysis. Samples (20 μg of total protein) were separated on a 12% denaturing polyacrylamide gel. Separated proteins were transferred to a nitrocellulose membrane (20 V, 90 min; transfer buffer 25 mmol/L Tris, 190 mmol/L glycine, 20% methanol, 0.5% sodium dodecyl sulfate) by electroblotter (Bio-rad Corporation). The membrane was placed into blocking buffer (5% non-fat milk in 20 mmol/L Tris/HCl, pH7.6, 140 mmol/L NaCl, 0.5% Tween 20) for 1 h at room temperature. Blocking buffer was decanted and the membrane was incubated with the primary antibody (1:1000 diluted in blocking buffer, Sigma Corporation) on a shaker at 4 °C overnight. After being washed (20 mmol/L Tris/HCl, pH7.6, 140 mmol/L NaCl, 0.1% Tween 20), the membrane was incubated with a peroxidase conjugated secondary antibody, which was diluted in 5% non-fat milk in wash buffer (1 h; room temperature; gentle shaking). Being washed, the membrane was exposed to sensitive film several minutes after incubating in Western blotting luminol reagent (Cell Signal Corporation). The bands were quantified by densitometry.
Immunohistochemistry detection
The expression of TNF-α and ICAM-1 in colon tissue was exhibited immunohistochemically, in which the employed primary polyclonal antibody was 1:100 rabbit-anti-rat-TNF-α and 1:50 goat-anti-rat-ICAM-1 (Santa Cruz Biotechnology Corporation), and the second antibody was biotinylation goat-anti-rabbit IgG and rabbit-anti-goat IgG, respectively. The slide was developed in 0.05% freshly prepared diaminobenzidine solution for several minutes, and then counterstained with hematoxylin. The expression of target protein was further semiquantitated according to the percentage of positively-stained cells and positive cell density. In each slide, the number of positive cells was determined in 16 D ocular micrometer (area of 0.1024 mm2, magnification×400) in 10 different areas, the average was positive cell density.
Myeloperoxidase assay
Colon samples were weighed and homogenized in a solution prepared from the assay kit (Nanjing Jiancheng Bioengineering Co. Ltd, China), and homogenates of 5% were obtained and used for MPO assay. MPO can catalyze the redox reaction of H2O2 and 3, 3, 5, 5-tetramethylbenzidine, and produces yellow colored compounds through whose absorbance at 460 nm MPO activity was calculated and expressed as U/g wet tissue. One unit of MPO activity was defined as the quantity of enzyme that degraded 1 μmoL H2O2 at 37 °C per g wet tissue.
Statistical analysis
Experimental results were analyzed by one-way ANOVA and t-tests for multiple comparisons between groups. Linear correlation analysis was performed to investigate the relationship between activity of NF-κB and other parameters. Data were finally expressed as mean±SE of the mean. P value less than 0.05 was considered statistically significant.
RESULTS
Macroscopic and histological evidence of inflammation
Pronounced pathological changes of colonic mucosa similar to that in human IBD were observed in rats with colitis induced by TNBS enema. Most animals inflicted with TNBS enema had disease limited to the rectum and rectosigmoid, some had disease extending beyond the sigmoid or had total colitis. The changes were most severe in the rectum and extended for a variable extent around the colon. The mucosa was hyperemic, edematous, and granular, and some small punctate ulcers were visible. In some samples, the ulceration was linear along the line of the teniae coli, exuberant regeneration of the epithelium was found in the colon. The inflammation was predominantly confined to the mucosa, and the lamina propria became edematous and the capillaries are dilated and congested, the ulcers were deep and undermine the surrounding epithelium. There was an inflammatory infiltrate of neutrophils, lymphocytes, and macrophages. Eosinophils and mast cells were also present in increased numbers. The colon mucosal macroscopic and histological injury indices were significantly increased in these experimental animals compared with normal controls (P<0.01). Colonic MPO activities of rats in model group were significantly increased compared with normal group and NS group (P<0.01, Figure 1 and Table 1).
Figure 1.
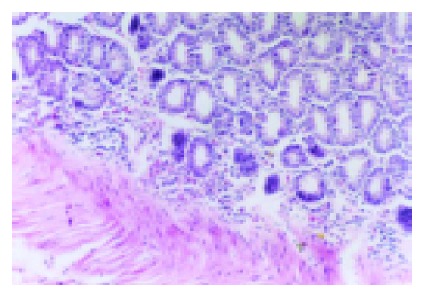
HE staining showed inflammatory infiltration of colon tissue in model group×200. A lot of inflammatory cells infiltrated into the colon tissue.
Table 1.
Macroscopical and histological damage indices and MPO activity in rats (n = 10).
| Group | Macroscopical | Histological | MPO activity (U/g) |
| Normal control | 0.75±0.71 | 1.13±0.83 | 0.31±0.08 |
| NS group | 1.63±0.74 | 1.38±0.52 | 0.39±0.07 |
| Model group I | 2.00±0.76bf | 3.88±0.99bf | 0.71±0.06bf |
| Model group II | 6.25±1.39bdf | 6.24±1.04bdf | 1.69±0.11bdf |
P<0.01 vs normal control;
P<0.01 vs NS group;
P<0.01 vs model group I.
Western blot of NF-κB P65
Nuclear level of NF-κB p65 protein were evaluated by Western blot in colonic samples. Rats of model group II (150 mg/kg TNBS enema) exhibited higher levels of nuclear NF-κB p65 than those of model group I (50 mg/kg TNBS enema), NS group, and normal group (P<0.01). In rats of model group I, more nuclear NF-κB p65 was found than that of normal controls (P<0.01), but no statistical difference was seen in comparison with NS group. But total levels of NF-κB p65 in colon tissues from the same sites were not different in rats of model group, NS group and normal controls (Figures 2, 3).
Figure 2.
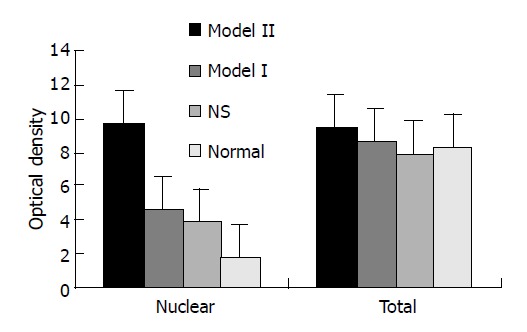
levels of NF-κB p65 in colon tissue of rats with TNBS-indued colitis.
Figure 3.
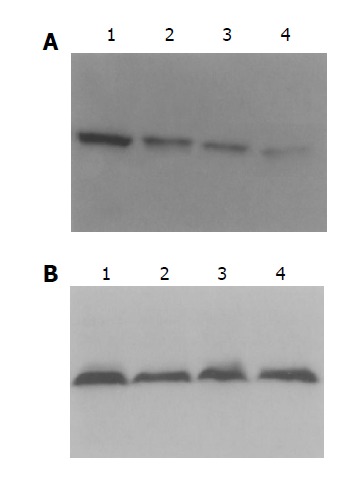
Western blotting showed levels of NF-κB P65 in colon tissue of rats. A: Rats of model group II exhibited higher levels of nuclear NF-κB P65 than others; B: total levels of NF-κB P65 were not different in rats. Lanes 1–4: model II, model I, NS, normal groups.
Expression of TNF-α and ICAM-1 in colon tissue
The TNF-α and ICAM-1 negatively expressed cells were manifested as blue-stained nuclei and the positive cell was with brown-yellow cytoplasma or nuclear membrane. Colonic TNF-α and ICAM-1 positive expressions were mainly observed on monocytes, neutrophils and there was also a sparse distribution in epithelial cells and vascular endothelial cells. The positive cells were predominantly located within the mucosa and mucosa lamina propria (Figures 4, 5).The expression of TNF-α and ICAM-1 in colon tissues were significantly increased in rats inflicted with TNBS enema compared with that of the NS group and normal controls (Table 2, P<0.01).
Figure 4.
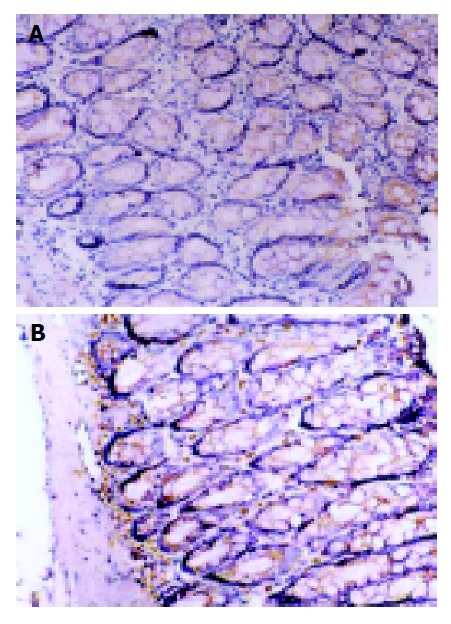
Immunohistochemical staining for TNF-α protein expression. SP×200. A: TNF-α protein expression in normal group; B: TNF-α expression in model II group. The number of TNF-α immunoreactive positive cells in model group was significantly more than that of normal controls.
Figure 5.
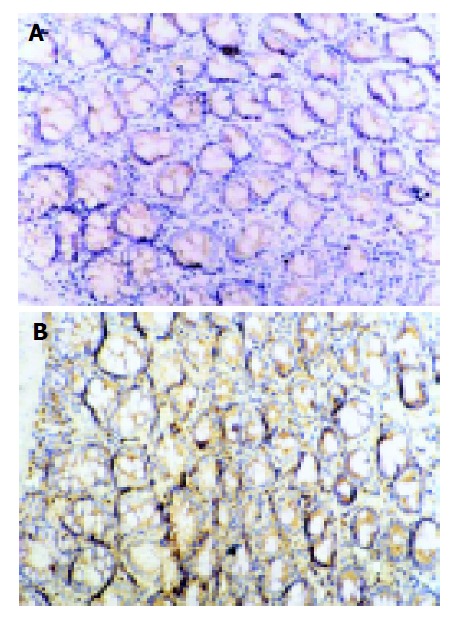
Immunohistochemical staining for ICAM-1 protein expression. SP×200. A: Expression of ICAM-1 in normal group; B: Expression of ICAM-1 in model II group. The ICAM-1 expression was upregulated markedly in model group.
Table 2.
Expression of TNF-α and ICAM-1 in colon tissue (n = 10).
| Group | TNF-α (positive density) | ICAM-1(positive density) |
| Normal control | 16.03±6.21 | 13.41±4.91 |
| NS group | 20.04±6.76 | 17.50±6.95 |
| Model group I | 53.40±8.79bf | 47.01±8.82bf |
| Model group II | 84.09±14.52bdf | 77.69±8.09bdf |
P<0.01 vs normal control;
P<0.01 vs NS group;
P<0.01 vs model group I.
Relativity analysis between TNF-α–ICAM-1, MPO, mucosal damage indices and NF-κB P65
Linear correlation analysis showed that a strong relativity existed between the levels of nuclear NF-κB p65 and the positive expression of TNF-α and ICAM-1, MPO activity, mucosal damage indices in TNBS-induced colitis. (r = 0.8235, 0.8780, 0.8572, 0.9152, 0.8247; P<0.05).
DISCUSSION
High levels of circulating and mucosal proinflammatory molecule are a characteristic feature of IBD[22-26]. Both TNF-α and ICAM-1 have been suggested to be important mediators involved in the initiation and perpetuation of intestinal inflammation in UC[27,28]. Infiltration and persistence of inflammatory cells within tissues are hallmarks of UC. Recruitment and activation of these cells to the inflammatory focus are multifactorial events involving adhesion of circulating cells to the vascular endothelium followed by tissue migration, and ICAM-1 is an active participant in mediating leukocyte adhesion to endothelial and epithelial cells. We confirmed that colon tissue obtained from rats with TNBS-induced colitis exhibited significantly more TNF-α and ICAM-1 expressions than normal and negative controls. This finding supported the idea that proinflammatory molecules participate in the occurrence and development of UC.
However, the pivotal factors in modulation of proinflammatory molecule synthesis remain unknown. Important candidates are transcription factors that bind to gene promoter regions and are involved in the regulation of inflammation gene transcription. Activation of NF-κB may be a pivotal event in proinflammatory signal transduction. In most resting cells, transcription factor members of the Rel-NF-κB are bound to cytoplasmic inhibitory factors IκBs, and are released on activation induced degradation of their cytoplasmic inhibitors by the proteasome/ubiquitin complex[14-17]. NF-κB p65 hetero- or homodimers migrate to the nucleus and regulate transcriptional activity by binding to specific DNA sequences in promoter/enhancer regions of inflammation genes[8,12,15,29]. We therefore investigated the levels of nuclear NF-κB P65 protein and found that increased amounts of NF-κB P65 protein in nuclear extracts of mucosal colon samples of rats with TNBS-induced colitis in comparison with both those treated with saline enema and normal controls. The highest levels were seen in rats inflicted with TNBS at larger dose. Moreover, the protein concentrations of NF-κB p65 were not different in intestinal mucosa from rats with TNBS-induced colitis, normal controls and those treated with saline enema if total tissue extracts were not separated into cytoplasmic and nuclear extracts. This finding is expected because activation of NF-κB results in a shift from the cytoplasm to the nucleus.
Translocation into the nucleus and binding to target DNA sequences are important events for NF-κB to control transcription process. Genes regulated by NF-κB include those encoding IL-2-IL-6–IL-8-the IL-2 receptor-the IL-12 p40 subunit- VCAM-1-ICAM-1-TNF-α, etc. And studies have also shown that the ICAM-1 and TNF-α gene promoter have binding sites for NF-κB. In our study, we observed that a strong relativity existed between the levels of nuclear NF-κB p65 and the positive expression of TNF-α and ICAM-1. These findings suggested that the heightened activation of NF-κB could be a major regulator of proinflammatory molecule synthesis and secretion in UC.
Our studies also observed that there was a positive relativity between the levels of nuclear NF-κB p65 and mucosal damage indices and MPO activity, which was considered as a marker of inflammation in colitis[30,31]. The results suggested that levels of nuclear NF-κB could reflect severity of diseases and be potentially helpful for us to evaluate therapeutic efficacy.
In summary, our studies suggested that activation of NF-κB may serve a center function as a key factor in the pathophysiology of chronic intestinal inflammation. Activation of NF-κB up-regulated the expression of proinflammatory molecule including TNF-α and ICAM-1 and participated in the occurrence and development of TNBS-induced colitis. Although further studies are needed to explore the specific mechanisms of activation and regulation, NF-κB could be a good target for therapy in UC, undoubtedly.
Footnotes
Science Editor Li WZ Language Editor Elsevier HK
References
- 1.MacDonald TT, Monteleone G, Pender SL. Recent developments in the immunology of inflammatory bowel disease. Scand J Immunol. 2000;51:2–9. doi: 10.1046/j.1365-3083.2000.00658.x. [DOI] [PubMed] [Google Scholar]
- 2.Lakatos L. Immunology of inflammatory bowel diseases. Acta Physiol Hung. 2000;87:355–372. [PubMed] [Google Scholar]
- 3.Ogata H, Hibi T. Cytokine and anti-cytokine therapies for inflammatory bowel disease. Curr Pharm Des. 2003;9:1107–1113. doi: 10.2174/1381612033455035. [DOI] [PubMed] [Google Scholar]
- 4.Inoue S, Matsumoto T, Iida M, Mizuno M, Kuroki F, Hoshika K, Shimizu M. Characterization of cytokine expression in the rectal mucosa of ulcerative colitis: correlation with disease activity. Am J Gastroenterol. 1999;94:2441–2446. doi: 10.1111/j.1572-0241.1999.01372.x. [DOI] [PubMed] [Google Scholar]
- 5.Ishiguro Y. Mucosal proinflammatory cytokine production correlates with endoscopic activity of ulcerative colitis. J Gastroenterol. 1999;34:66–74. doi: 10.1007/s005350050218. [DOI] [PubMed] [Google Scholar]
- 6.Xia B, Guo HJ, Crusius J, Deng CS, Meuwissen S, Pena A. In vitro production of TNF-alpha,IL-6 and sIL-2R in Chinese patients with ulcerative colitis. World J Gastroenterol. 1998;4:252–255. doi: 10.3748/wjg.v4.i3.252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sans M, Panés J, Ardite E, Elizalde JI, Arce Y, Elena M, Palacín A, Fernández-Checa JC, Anderson DC, Lobb R, et al. VCAM-1 and ICAM-1 mediate leukocyte-endothelial cell adhesion in rat experimental colitis. Gastroenterology. 1999;116:874–883. doi: 10.1016/s0016-5085(99)70070-3. [DOI] [PubMed] [Google Scholar]
- 8.Baldwin AS. Series introduction: the transcription factor NF-kappaB and human disease. J Clin Invest. 2001;107:3–6. doi: 10.1172/JCI11891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zaninoni A, Imperiali FG, Pasquini C, Zanella A, Barcellini W. Cytokine modulation of nuclear factor-kappaB activity in B-chronic lymphocytic leukemia. Exp Hematol. 2003;31:185–190. doi: 10.1016/s0301-472x(02)01046-9. [DOI] [PubMed] [Google Scholar]
- 10.Kis A, Yellon DM, Baxter GF. Role of nuclear factor-kappa B activation in acute ischaemia-reperfusion injury in myocardium. Br J Pharmacol. 2003;138:894–900. doi: 10.1038/sj.bjp.0705108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Glasgow JN, Wood T, Perez-Polo JR. Identification and characterization of nuclear factor kappaB binding sites in the murine bcl-x promoter. J Neurochem. 2000;75:1377–1389. doi: 10.1046/j.1471-4159.2000.0751377.x. [DOI] [PubMed] [Google Scholar]
- 12.Tak PP, Firestein GS. NF-kappaB: a key role in inflammatory diseases. J Clin Invest. 2001;107:7–11. doi: 10.1172/JCI11830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ghosh S, May MJ, Kopp EB. NF-kappa B and Rel proteins: evolutionarily conserved mediators of immune responses. Annu Rev Immunol. 1998;16:225–260. doi: 10.1146/annurev.immunol.16.1.225. [DOI] [PubMed] [Google Scholar]
- 14.Karin M, Ben-Neriah Y. Phosphorylation meets ubiquitination: the control of NF-[kappa]B activity. Annu Rev Immunol. 2000;18:621–663. doi: 10.1146/annurev.immunol.18.1.621. [DOI] [PubMed] [Google Scholar]
- 15.May MJ, Ghosh S. Signal transduction through NF-kappa B. Immunol Today. 1998;19:80–88. doi: 10.1016/s0167-5699(97)01197-3. [DOI] [PubMed] [Google Scholar]
- 16.Rothwarf DM, Zandi E, Natoli G, Karin M. IKK-gamma is an essential regulatory subunit of the IkappaB kinase complex. Nature. 1998;395:297–300. doi: 10.1038/26261. [DOI] [PubMed] [Google Scholar]
- 17.Nakano H, Shindo M, Sakon S, Nishinaka S, Mihara M, Yagita H, Okumura K. Differential regulation of IkappaB kinase alpha and beta by two upstream kinases, NF-kappaB-inducing kinase and mitogen-activated protein kinase/ERK kinase kinase-1. Proc Natl Acad Sci USA. 1998;95:3537–3542. doi: 10.1073/pnas.95.7.3537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Morris GP, Beck PL, Herridge MS, Depew WT, Szewczuk MR, Wallace JL. Hapten-induced model of chronic inflammation and ulceration in the rat colon. Gastroenterology. 1989;96:795–803. [PubMed] [Google Scholar]
- 19.Dieleman LA, Palmen MJ, Akol H, Bloemena E, Peña AS, Meuwissen SG, Van Rees EP. Chronic experimental colitis induced by dextran sulphate sodium (DSS) is characterized by Th1 and Th2 cytokines. Clin Exp Immunol. 1998;114:385–391. doi: 10.1046/j.1365-2249.1998.00728.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hou J, Schindler U, Henzel WJ, Ho TC, Brasseur M, McKnight SL. An interleukin-4-induced transcription factor: IL-4 Stat. Science. 1994;265:1701–1706. doi: 10.1126/science.8085155. [DOI] [PubMed] [Google Scholar]
- 21.Osborn L, Kunkel S, Nabel GJ. Tumor necrosis factor alpha and interleukin 1 stimulate the human immunodeficiency virus enhancer by activation of the nuclear factor kappa B. Proc Natl Acad Sci USA. 1989;86:2336–2340. doi: 10.1073/pnas.86.7.2336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Monteleone G, MacDonald TT. Manipulation of cytokines in the management of patients with inflammatory bowel disease. Ann Med. 2000;32:552–560. doi: 10.3109/07853890008998835. [DOI] [PubMed] [Google Scholar]
- 23.Brown KA, Back SJ, Ruchelli ED, Markowitz J, Mascarenhas M, Verma R, Piccoli DA, Baldassano RN. Lamina propria and circulating interleukin-6 in newly diagnosed pediatric inflammatory bowel disease patients. Am J Gastroenterol. 2002;97:2603–2608. doi: 10.1111/j.1572-0241.2002.06030.x. [DOI] [PubMed] [Google Scholar]
- 24.Sasaki M, Jordan P, Houghton J, Meng X, Itoh M, Joh T, Alexander JS. Transfection of IL-10 expression vectors into endothelial cultures attenuates alpha4beta7-dependent lymphocyte adhesion mediated by MAdCAM-1. BMC Gastroenterol. 2003;3:3. doi: 10.1186/1471-230X-3-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tamura K, Fukuda Y, Sashio H, Takeda N, Bamba H, Kosaka T, Fukui S, Sawada K, Tamura K, Satomi M, et al. IL18 polymorphism is associated with an increased risk of Crohn's disease. J Gastroenterol. 2002;37 Suppl 14:111–116. doi: 10.1007/BF03326428. [DOI] [PubMed] [Google Scholar]
- 26.Ito H, Hirotani T, Yamamoto M, Ogawa H, Kishimoto T. Anti-IL-6 receptor monoclonal antibody inhibits leukocyte recruitment and promotes T-cell apoptosis in a murine model of Crohn's disease. J Gastroenterol. 2002;37 Suppl 14:56–61. doi: 10.1007/BF03326415. [DOI] [PubMed] [Google Scholar]
- 27.Louis E. The immuno-inflammatory reaction in Crohn's disease and ulcerative colitis: characterisation, genetics and clinical application. Focus on TNF alpha. Acta Gastroenterol Belg. 2001;64:1–5. [PubMed] [Google Scholar]
- 28.Woywodt A, Ludwig D, Neustock P, Kruse A, Schwarting K, Jantschek G, Kirchner H, Stange EF. Mucosal cytokine expression, cellular markers and adhesion molecules in inflammatory bowel disease. Eur J Gastroenterol Hepatol. 1999;11:267–276. doi: 10.1097/00042737-199903000-00010. [DOI] [PubMed] [Google Scholar]
- 29.Lee JI, Burckart GJ. Nuclear factor kappa B: important transcription factor and therapeutic target. J Clin Pharmacol. 1998;38:981–993. doi: 10.1177/009127009803801101. [DOI] [PubMed] [Google Scholar]
- 30.Shusterman T, Sela S, Cohen H, Kristal B, Sbeit W, Reshef R. Effect of the antioxidant Mesna (2-mercaptoethane sulfonate) on experimental colitis. Dig Dis Sci. 2003;48:1177–1185. doi: 10.1023/a:1023741518385. [DOI] [PubMed] [Google Scholar]
- 31.Vowinkel T, Kalogeris TJ, Mori M, Krieglstein CF, Granger DN. Impact of dextran sulfate sodium load on the severity of inflammation in experimental colitis. Dig Dis Sci. 2004;49:556–564. doi: 10.1023/b:ddas.0000026298.72088.f7. [DOI] [PubMed] [Google Scholar]