

Abstract
Myosin binding protein-C (MyBP-C) was first discovered as an impurity during the purification of myosin from skeletal muscle. However, soon after its discovery MyBP-C was also shown to bind actin. While the unique functional implications for a protein that could cross-link thick and thin filaments together were immediately recognized, most early research nonetheless focused on interactions of MyBP-C with the thick filament. This was in part because interactions of MyBP-C with the thick filament could adequately explain most (but not all) effects of MyBP-C on actomyosin interactions and in part because the specificity of actin binding was uncertain. However, numerous recent studies have now established that MyBP-C can indeed bind to actin through multiple binding sites, some of which are highly specific. Many of these interactions involve critical regulatory domains of MyBP-C that are also reported to interact with myosin. Here we review current evidence supporting MyBP-C interactions with actin and discuss these findings in terms of their ability to account for the functional effects of MyBP-C. We conclude that the influence of MyBP-C on muscle contraction can be explained equally well by interactions with actin as by interactions with myosin. However, because data showing that MyBP-C binds to either myosin or actin has come almost exclusively from in vitro biochemical studies, the challenge for future studies is to define which binding partner(s) MyBP-C interacts with in vivo.
Keywords: Myosin, actin, myosin binding protein-C, sarcomere, muscle
Introduction
In the forty years since the discovery of myosin binding protein-C (MyBP-C) [35] it has become clear that MyBP-C is a key regulator of muscle contraction that affects interactions between myosin and actin and that is essential for normal cardiac muscle contraction (for reviews [3, 4, 42]). The importance of MyBP-C to muscle contraction is further underscored by the discovery that mutations in genes encoding MyBP-C cause myopathies in both skeletal [15, 29] and cardiac muscle [7, 17], and that MyBP-C is involved in cardiac stress pathways during both normal physiologic signaling and in pathological states such as heart failure [42].
What is far less certain and what remains a critical unresolved question is the precise mechanism(s) by which MyBP-C affects muscle contraction. Because of its ability to bind to myosin through two discrete binding sites, early hypotheses focused on the idea that interactions of MyBP-C with the thick filament alone were sufficient to account for the regulatory effects of MyBP-C [47]. However, other early data [31, 32, 52] and more recent studies provide a compelling case that MyBP-C can also interact with the thin filament to affect contraction [43, 49]. The distinction is important not only for better understanding effects of MyBP-C on contraction but also for better insight in how sarcomeres function. Linkage of the thick and thin filaments by MyBP-C could represent a previously unrecognized filament system working in concert with thick filaments, thin filaments and titin to impact force development, transmission, sensing, and signaling during striated muscle contraction.
Structure and Position of MyBP-C in Sarcomeres
MyBP-C is expressed in vertebrate striated muscles where it occurs as distinct isoforms originating from three separate genes: 2 skeletal genes that correspond to expression predominantly in slow and fast skeletal muscle [48] and a third cardiac gene expressed in heart [9]. All isoforms consist of a series of immunoglobin (Ig) and fibronectin III (Fn)-like domains numbered C1 through C10 (Fig. 1a), whereas cardiac (c) MyBP-C has an extra N-terminal Ig domain referred to as C0. Importantly, the cardiac isoform also has additional phosphorylation sites within the regulatory motif (also referred to as the M-domain) which is a linker sequence of ~100 amino acids located between C1 and C2 that is unique to MyBP-C proteins.
Fig. 1.

a Schematic representation of myosin binding protein C (MyBP-C). Shown are lg-like domains (ovals) and Fn-III like domains (rectangles). The cardiac isoform of MyBP-C has an extra domain at the N-terminus (C0) and additional phosphorylation sites in the M-domain between C1 and C2 (asterisks) b Electron micrograph image demonstrating the stripes where MyBP-C is located. Pictures from Luther et al. [52]
Within a sarcomere MyBP-C is localized to a characteristic set of 7-9 discrete stripes (with the exact number of stripes depending on the type of muscle [26]) that are evenly spaced in the C-zone of the A-band. Within the C-zone, i.e., the region where the thick and thin filaments overlap, MyBP-C stripes can be visualized in negatively stained electron microscopy sections or using immuno-EM labeling (Fig. 1b). The stripe spacing (~43 nm apart) corresponds roughly to every third crown of myosin heads emerging from the thick filament backbone. Thus, MyBP-C is present at a limited stoichiometry, but in discrete positions relative to myosin.
Myosin, the first binding partner of MyBP-C
Given its co-purification with, and strong binding to myosin, it is not surprising that early studies investigating the influence of MyBP-C on actomyosin interactions initially focused on its connections to myosin. MyBP-C was shown to bind to both the light meromyosin (LMM) and the S2 subfragment of myosin [33, 47]. The binding site for LMM was identified in the C10 domain [30, 36], whereas regions of MyBP-C that interacted with myosin S2 were identified closer to the N-terminus of the molecule within domains C1-M-C2 [13, 14]. Binding to S2 was further found to be eliminated by phosphorylation of the motif of cardiac MyBP-C, suggesting a role in mediating the inotropic effects of cMyBP-C [13, 14]. The corresponding location on S2 where the motif bound was narrowed down to the proximal 126 amino acids of myosin S2 (S2δ) [13]. Ababou et al. [1, 2] demonstrated that C1 and C2 also bound to the S2δ segment of myosin, albeit through much weaker interactions.
Actin, the second binding partner of MyBP-C
Soon after the discovery of MyBP-C it was found that MyBP-C could also bind to actin. MyBP-C was shown to interact with F-actin, isolated thin filaments, and the I-bands of sarcomeres [31, 32, 52]. Importantly, binding appeared specific in that it could be competed off by myosin S1 heads in the absence of ATP (i.e., by rigor S1 heads) or that MyBP-C could displace S1 heads in the presence of ATP [32].
The finding that MyBP-C could bind thin filaments was striking in that it marked the discovery of the only myofilament protein (aside from myosin) that could simultaneously link thick and thin filaments together within the region of active cross-bridge cycling. Functional roles for a protein that could span the two filament systems were immediately suggested, such as stabilization of the sarcomere lattice through weak coupling of filaments in relaxed muscle or that MyBP-C could make transient contacts with actin such that myosin cross-bridge kinetics could be affected in contracting muscle [32]. Other possibilities include direct effects of MyBP-C to regulate thin filament activation and relaxation as well as mechano-sensing and signaling [20, 43].
Renewed interest in the physiological significance of the ability of MyBP-C to bind actin came with the discovery that the same recombinant truncated MyBP-C proteins that were found to bind myosin S2 could also bind to actin and thin filaments [43]. Although initially surprising, results were confirmed in multiple studies by different labs [34, 51] collectively showing that N-terminal domains of MyBP-C could bind actin with an affinity similar to binding myosin S2 [43].
Specific sequences that mediate actin binding were next identified in the C0 [22, 25, 37, 51] and C1 domains [6, 43], within the first 17 amino acids of the motif [50], and within the folded tri-helix structure of the motif [5]. In the tri-helix bundle a highly conserved sequence was found that bears homology to the actin binding sequences of troponin I, twitchin, and other actin binding proteins [8, 11, 23]. Missense variants associated with hypertrophic cardiomyopathy also affected actin binding, further supporting the idea that tri-helix bundle mediates actin binding of the motif [5].
The possibility that multiple interaction sites dispersed throughout multiple N-terminal domains contribute to the actin binding properties of MyBP-C is supported by the ability of C1C2 to cross-link F-actin filaments [34, 43] and by 3-D reconstructions of actin decorated with N-terminal MyBP-C domains showing that the recombinant proteins can span multiple actin monomers [34, 37]. Similar conclusions were reached through modeling of small angle X-ray and neutron scattering data and NMR data [25, 51] .
Additional studies have pointed to other actin binding sites located in the prolinealanine rich repeat [46], or domains in C-terminal half of the molecule [41]. However, a consensus regarding these sites has yet to emerge from multiple labs. Importantly, differences between labs, especially with respect to C0 and the proline-alanine rich region, are likely to reflect the existence of species-specific differences in the behavior of the recombinant proteins (e.g., mouse versus human sequences) because the human C0 and proline-alanine regions are both able to activate motility or force in the absence of Ca2+, whereas the corresponding mouse domains are not effective [45]. Differences in experimental conditions are also likely to contribute to differences between labs because even modest changes in pH or phosphorylation state had large effects on the amount of C1-M-C2 bound to actin [43]. The latter is especially relevant considering the use of different protein expression systems (e.g., bacterial versus eukaryotic systems) that result in proteins with varying degrees of phosphorylation or other post-translational modifications [12].
Can interactions with myosin or actin adequately explain effects of MyBP-C on contraction?
Interactions with myosin
Starr and Offer [47] first suggested an elegant mechanism by which MyBP-C could influence cross bridge function solely through MyBP-C interactions with the thick filament (Fig 2). According to these authors, interactions of MyBP-C with myosin S2 could restrict the outward movement of cross-bridges toward the thin filament. They further suggested that regulation of this mechanism could provide a means to regulate interaction of myosin heads with actin. Indeed, because phosphorylation of cardiac MyBP-C abolishes interactions with myosin S2 [14] and increases the proximity of myosin heads to actin [10] their model provides a straightforward mechanism to account for the ability of MyBP-C to limit cross-bridge interactions with actin. Alternatively, by connecting S2 to the LMM region of myosin MyBP-C could act as a drag on slowly cycling cross bridges as proposed by Hofmann et al. [19] (Fig. 2). In addition, by binding to the critical S1/S2 junction MyBP-C is in an ideal location to influence interactions with other thick filament proteins such as RLC [28, 39] or to otherwise affect myosin S1 head position or head-head interactions.
Fig. 2.
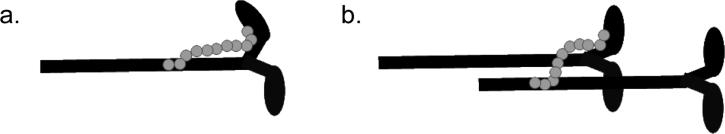
Thick filament model of regulation by MyBP-C. MyBP-C (grey) can crossink myosin heads to the rod portion of the same (a) or adjacent (b) myosin moleucles (black). By doing so, MyBP-C can prevent myosin heads from moving towards the actin filament, as proposed by Starr and Offer [9], or MyBP-C could form a physical drag on long-lived cross bridges, as suggested by Hofman et al. [42].
However, a limitation of the thick filament model is that it cannot account for effects of MyBP-C in ATPase assays where the thick filament is entirely absent or in assays that use only the S1 subfragment of myosin and the possibility of intra-myosin interactions is eliminated [44]. Perhaps even more difficult to reconcile with a thick filament mechanism are observations that MyBP-C can activate actin-S1 interactions in the presence of thin filaments (i.e., actin plus troponin and tropomyosin) even when Ca2+ is absent [18, 40, 44]. Therefore, other mechanisms must exist by which MyBP-C can directly influence cross bridge interactions, either in addition to or in place of, mechanisms that rely solely on inhibitory interactions with S2.
Interactions with actin
On the other hand, the ability of MyBP-C to both activate and inhibit actomyosin interactions can be readily explained if MyBP-C binds directly to actin or the thin filament to affect its activation state (Fig. 3a). Structural evidence in support of this idea comes from reconstructions of F-actin or thin filaments decorated with recombinant truncated MyBP-C proteins showing that they bind in positions that overlap with binding sites of myosin S1 heads and that they can sterically clash with the position of tropomyosin in the closed, low Ca2+ state, but not in the high Ca2+ open position [34, 51]. Thus, by binding to the thin filament MyBP-C is in a position to both inhibit actomyosin interactions by directly competing with myosin S1 heads or to activate the thin filament by interfering with the closed state of tropomyosin (Fig. 3). The ability of MyBP-C to directly activate the thin filament may be important in shortening muscle where MyBP-C could offset the tendency of the thin filament to deactivate during shortening [16]. If so, then loss of thin filament activating effects could explain the abbreviated systolic ejection phase in mice lacking cMyBP-C [38]. Binding of MyBP-C to thin filaments can also readily explain the ability of MyBP-C to slow muscle shortening velocity and limit power output in cMyBP-C knockout myocytes [21]. In this scenario MyBP-C would act as a physical drag by crosslinking the thin and thick filaments. Lastly, the ability of MyBP-C to compete with myosin S1 heads and inhibit actomyosin interactions [32, 44] could provide an explanation for the long-standing puzzle that MyBP-C is present at a limited stoichiometry with respect to myosin: by being present at a low concentration relative to myosin heads, the activating effects of MyBP-C on the thin filament may be optimized while inhibitory competitive effects with myosin cross bridges are minimized.
Fig. 3.

Thin filament regulation of contraction by MuBP-C. a Direct activation of the thin filament by displacement of tropomyosin by MyBP-C, enabling across bridge formation during low Ca2+ conditions. b Blocking of S1 binding sited on acting by MyBP-C, inhibing cross bridge formation despite the open state of tropomyosin. Optimization of the activating effects of MyBP-C (a) while limiting competition with myosin (b) may explain the limited localization of MyBP-C in sarcomeres.
MyBP-C binding interactions in vivo
While there is considerable evidence to conclude that MyBP-C can bind to both myosin S2 and the thin filament, at present there is no direct evidence that MyBP-C interacts with either myosin S2 or actin in sarcomeres. It is possible that the densities attributed to MyBP-C in reconstructions of the cardiac thick filament [53] are due to binding of MyBP-C N-terminal domains to S2 (rather than the C-terminal domains thought to interact with the thick filament backbone), but this possibility has yet to be critically tested. Perhaps the most detailed visualization of whole MyBP-C in sarcomeres to date has come from Luther and colleagues [27] who used electron microscope tomography to reveal that MyBP-C extends out away from the thick filament at sufficient distance to interact with the thin filament. These results are in excellent agreement with previous conclusions from X-ray diffraction data that MyBP-C extends radially outward from the thick filament supporting the idea that MyBP-C interacts with the thin filament in sarcomeres [46].
Resolution of the central question of whether MyBP-C binds to actin or myosin S2 to affect contraction will thus ultimately require innovations in structural methods that allow more precise visualization of the position(s) of MyBP-C in the sarcomere. Of course, it is possible that MyBP-C binds to both myosin S2 and to the actin filament to affect contraction. The essential questions then become whether binding interactions are simultaneous, competitive, or sequential. For example, it is an intriguing possibility that MyBP-C regulates both the relaxed state of the thick filament by binding to myosin S2 and that phosphorylation or other changes in environment then release MyBP-C so that it can influence the activation state of the thin filament by binding to actin or to other thin filament regulatory proteins. Dynamic changes in binding to the thick versus thin filament could be signaled through additional regulatory partners such as Ca2+/calmodulin binding to the regulatory M-domain [24] or by forced extension of the M-domain to reveal cryptic ligand binding sites [5, 20]. Time-resolved methods such as FRET and X-ray diffraction that can differentiate dynamic changes in binding to the thick and thin filaments will thus be essential in further defining the mechanism(s) by which MyBP-C affects contraction. These approaches combined with genetic methods to enhance the actin binding affinity of MyBP-C in vivo should prove insightful in probing the significance of MyBP-C interactions with actin in sarcomeres. These experiments are underway.
Footnotes
This article is published as part of the Special Issue on Cardiac Myosin Binding Protein-C as a Central Target of Cardiac Sarcomere Signaling.
References
- 1.Ababou A, Gautel M, Pfuhl M. Dissecting the N-terminal myosin binding site of human cardiac myosin-binding protein C. Structure and myosin binding of domain C2. J Biol Chem. 2007;282:9204–9215. doi: 10.1074/jbc.M610899200. [DOI] [PubMed] [Google Scholar]
- 2.Ababou A, Rostkova E, Mistry S, et al. Myosin binding protein C positioned to play a key role in regulation of muscle contraction: structure and interactions of domain C1. J Mol Biol. 2008;384:615–630. doi: 10.1016/j.jmb.2008.09.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ackermann MA, Kontrogianni-Konstantopoulos A. Myosin binding protein-C: a regulator of actomyosin interaction in striated muscle. J Biomed Biotechnol. 2011;2011:636403. doi: 10.1155/2011/636403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bennett PM, Furst DO, Gautel M. The C-protein (myosin binding protein C) family: regulators of contraction and sarcomere formation? Rev Physiol Biochem Pharmacol. 1999;138:203–234. doi: 10.1007/BFb0119628. [DOI] [PubMed] [Google Scholar]
- 5.Bezold KL, Shaffer JF, Khosa JK, et al. A gain-of-function mutation in the M-domain of cardiac myosin-binding protein-C increases binding to actin. J Biol Chem. 2013;288:21496–21505. doi: 10.1074/jbc.M113.474346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bhuiyan MS, Gulick J, Osinska H, et al. Determination of the critical residues responsible for cardiac myosin binding protein C's interactions. J Mol Cell Cardiol. 2012;53:838–847. doi: 10.1016/j.yjmcc.2012.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bonne G, Carrier L, Richard P, et al. Familial hypertrophic cardiomyopathy: from mutations to functional defects. Circ Res. 1998;83:580–593. doi: 10.1161/01.res.83.6.580. [DOI] [PubMed] [Google Scholar]
- 8.Butler TM, Mooers SU, Narayan SR, Siegman MJ. The N-terminal region of twitchin binds thick and thin contractile filaments: redundant mechanisms of catch force maintenance. J Biol Chem. 2010;285:40654–40665. doi: 10.1074/jbc.M110.166041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Carrier L, Bonne G, Bahrend E, et al. Organization and sequence of human cardiac myosin binding protein C gene (MYBPC3) and identification of mutations predicted to produce truncated proteins in familial hypertrophic cardiomyopathy. Circ Res. 1997;80:427–434. [PubMed] [Google Scholar]
- 10.Colson BA, Bekyarova T, Locher MR, et al. Protein kinase A-mediated phosphorylation of cMyBP-C increases proximity of myosin heads to actin in resting myocardium. Circ Res. 2008;103:244–251. doi: 10.1161/CIRCRESAHA.108.178996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Friederich E, Vancompernolle K, Huet C, et al. An actin-binding site containing a conserved motif of charged amino acid residues is essential for the morphogenic effect of villin. Cell. 1992;70:81–92. doi: 10.1016/0092-8674(92)90535-k. [DOI] [PubMed] [Google Scholar]
- 12.Ge Y, Rybakova IN, Xu Q, Moss RL. Top-down high-resolution mass spectrometry of cardiac myosin binding protein C revealed that truncation alters protein phosphorylation state. Proc Natl Acad Sci USA. 2009;106:12658–12663. doi: 10.1073/pnas.0813369106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gruen M, Gautel M. Mutations in β-Myosin S2 taht cause Familial Hypertrophic Cardiomyopathy (FHC) Abolish the Interaction with the Regulatory Domain of Myosin-binding Protein-C. J Mol Biol. 1999;286:933–949. doi: 10.1006/jmbi.1998.2522. [DOI] [PubMed] [Google Scholar]
- 14.Gruen M, Prinz H, Gautel M. cAPK-phosphorylation controls the interaction of the regulatory domain of cardiac myosin binding protein C with myosin-S2 in an on-off fashion. FEBS Lett. 1999;453:254–259. doi: 10.1016/s0014-5793(99)00727-9. [DOI] [PubMed] [Google Scholar]
- 15.Gurnett CA, Desruisseau DM, McCall K, et al. Myosin binding protein C1: a novel gene for autosomal dominant distal arthrogryposis type. 1. Hum Mol Genet. 2010;19:1165–1173. doi: 10.1093/hmg/ddp587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hanft LM, Korte FS, McDonald KS. Cardiac function and modulation of sarcomeric function by length. Cardiovasc Res. 2008;77:627–636. doi: 10.1093/cvr/cvm099. [DOI] [PubMed] [Google Scholar]
- 17.Harris SP, Lyons RG, Bezold KL. In the thick of it: HCM-causing mutations in myosin binding proteins of the thick filament. Circ Res. 2011;108:751–764. doi: 10.1161/CIRCRESAHA.110.231670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Herron TJ, Rostkova E, Kunst G, et al. Activation of myocardial contraction by the N-terminal domains of myosin binding protein-C. Circ Res. 2006;98:1290–1298. doi: 10.1161/01.RES.0000222059.54917.ef. [DOI] [PubMed] [Google Scholar]
- 19.Hofmann PA, Greaser ML, Moss RL. C-protein limits shortening velocity of rabbit skeletal muscle fibres at low levels of Ca2+ activation. J Physiol. 1991;439:701–715. doi: 10.1113/jphysiol.1991.sp018689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Karsai A, Kellermayer MS, Harris SP. Mechanical unfolding of cardiac myosin binding protein-C by atomic force microscopy. Biophysj. 2011;101:1968–1977. doi: 10.1016/j.bpj.2011.08.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Korte FS, McDonald KS, Harris SP, Moss RL. Loaded shortening, power output, and rate of force redevelopment are increased with knockout of cardiac myosin binding protein-C. Circ Res. 2003;93:752–758. doi: 10.1161/01.RES.0000096363.85588.9A. [DOI] [PubMed] [Google Scholar]
- 22.Kulikovskaya I, McClellan G, Flavigny J, et al. Effect of MyBP-C binding to actin on contractility in heart muscle. J Gen Physiol. 2003;122:761–774. doi: 10.1085/jgp.200308941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Levine BA, Moir AJ, Perry SV. The interaction of troponin-I with the N-terminal region of actin. Eur J Biochem. 1988;172:389–397. doi: 10.1111/j.1432-1033.1988.tb13899.x. [DOI] [PubMed] [Google Scholar]
- 24.Lu Y, Kwan AH, Jeffries CM, et al. The Motif of Human Cardiac Myosin Binding Protein C is Required for its Ca2+-Dependent Interaction with Calmodulin. J Biol Chem. 2012 doi: 10.1074/jbc.M112.383299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lu Y, Kwan AH, Trewhella J, Jeffries CM. The C0C1 fragment of human cardiac myosin binding protein C has common binding determinants for both actin and myosin. J Mol Biol. 2011;413:908–913. doi: 10.1016/j.jmb.2011.09.026. [DOI] [PubMed] [Google Scholar]
- 26.Luther PK, Craig R. Modulation of striated muscle contraction by binding of myosin binding protein C to actin. BioArchitecture. 2011;1:277–283. doi: 10.4161/bioa.1.6.19341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Luther PK, Winkler H, K. T, et al. Direct visualization of myosin-binding protein C bridging myosin and actin filaments in intact muscle. Proc Natl Acad Sci. 2011;108:11423–11428. doi: 10.1073/pnas.1103216108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Margossian SS. Reversible dissociation of dog cardiac myosin regulatory light chain 2 and its influence on ATP hydrolysis. J Biol Chem. 1985;260:13747–13754. [PubMed] [Google Scholar]
- 29.Markus B, Narkis G, Landau D, et al. Autosomal recessive lethal congenital contractural syndrome type 4 (LCCS4) caused by a mutation in MYBPC1. Hum Mutat. 2012;33:1435–1438. doi: 10.1002/humu.22122. [DOI] [PubMed] [Google Scholar]
- 30.Miyamoto CA, Fischman DA, Reinach FC. The interface between MyBP-C and myosin: site-directed mutagenesis of the CX myosin-binding domain of MyBP-C. J Muscle Res Cell Motil. 1999;20:703–715. doi: 10.1023/a:1005513312939. [DOI] [PubMed] [Google Scholar]
- 31.Moos C. Fluorescence microscope study of the binding of added C protein to skeletal muscle myofibrils. J Cell Biol. 1981;90:25–31. doi: 10.1083/jcb.90.1.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Moos C, Mason CM, Besterman JM, et al. The Binding of Skeletal Muscle C-protein to F-actin, and its Relation to the Interaction of Actin with Myosin Subfragment-1. J Mol Biol. 1978;124:571–586. doi: 10.1016/0022-2836(78)90172-9. [DOI] [PubMed] [Google Scholar]
- 33.Moos C, Offer G, Starr R, Bennett P. Interaction of C-protein with myosin, myosin rod and light meromyosin. J Mol Biol. 1975;97:1–9. doi: 10.1016/s0022-2836(75)80017-9. [DOI] [PubMed] [Google Scholar]
- 34.Mun JY, Gulick J, Robbins J, et al. Electron microscopy and 3D reconstruction of F-actin decorated with cardiac myosin-binding protein C (cMyBP-C). J Mol Biol. 2011;410:214–225. doi: 10.1016/j.jmb.2011.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Offer G, Moos C, Starr R. A New Protein of the Thick filaments of Vertebrate Skeletal Myofybrils. J Mol Biol. 1973;74:653–376. doi: 10.1016/0022-2836(73)90055-7. [DOI] [PubMed] [Google Scholar]
- 36.Okagaki T, Weber FE, Fischman DA, et al. The major myosin-binding domain of skeletal muscle MyBP-C (C protein) resides in the COOH-terminal, immunoglobulin C2 motif. J Cell Biol. 1993;123:619–626. doi: 10.1083/jcb.123.3.619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Orlova A, Galkin VE, Jeffries CM, et al. The N-terminal domains of myosin binding protein C can bind polymorphically to F-actin. J Mol Biol. 2011;412:379–386. doi: 10.1016/j.jmb.2011.07.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Palmer BM, Georgakopoulos D, Janssen PM, et al. Role of cardiac myosin binding protein C in sustaining left ventricular systolic stiffening. Circ Res. 2004;94:1249–1255. doi: 10.1161/01.RES.0000126898.95550.31. [DOI] [PubMed] [Google Scholar]
- 39.Ratti J, Rostkova E, Gautel M, Pfuhl M. Structure and interactions of myosin-binding protein C domain C0: cardiac-specific regulation of myosin at its neck? J Biol Chem. 2011;286:12650–12658. doi: 10.1074/jbc.M110.156646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Razumova MV, Shaffer JF, Tu AY, et al. Effects of the N-terminal domains of myosin binding protein-C in an in vitro motility assay: Evidence for long-lived cross-bridges. J Biol Chem. 2006;281:35846–35854. doi: 10.1074/jbc.M606949200. [DOI] [PubMed] [Google Scholar]
- 41.Rybakova IN, Greaser ML, Moss RL. Myosin binding protein C interaction with actin: characterization and mapping of the binding site. J Biol Chem. 2011;286:2008–2016. doi: 10.1074/jbc.M110.170605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sadayappan S, de Tombe PP. Cardiac myosin binding protein-C: redefining its structure and function. Biophys Rev. 2012;4:93–106. doi: 10.1007/s12551-012-0067-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Shaffer JF, Kensler RW, Harris SP. The myosin-binding protein C motif binds to F-actin in a phosphorylation-sensitive manner. J Biol Chem. 2009;284:12318–12327. doi: 10.1074/jbc.M808850200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Shaffer JF, Razumova MV, Tu AY, et al. Myosin S2 is not required for effects of myosin binding protein-C on motility. FEBS Lett. 2007;581:1501–1504. doi: 10.1016/j.febslet.2007.03.007. [DOI] [PubMed] [Google Scholar]
- 45.Shaffer JF, Wong P, Bezold KL, Harris SP. Functional differences between. the N-terminal domains of mouse and human myosin binding protein protein-C. J Biomed Biotechnol. 2010;2010:789798. doi: 10.1155/2010/789798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Squire JM, Luther PK, Knupp C. Structural Evidence for the Interaction of C-protein (MyBP C) with Actin and Sequence Identification of a Possible Actin Actin-binding Domain. J Mol Biol. 2003;331:713–724. doi: 10.1016/s0022-2836(03)00781-2. [DOI] [PubMed] [Google Scholar]
- 47.Starr R, Offer G. The Interaction of C C-Prot ein with Heavy Meromysin and Subfragment-2. Biochem J. 1978;171:813–816. doi: 10.1042/bj1710813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Weber FE, Vaughan KT, Reinach FC, Fischman DA. Complete sequence of human fast-type and slow slow-type muscle myosin-binding-protein C (MyBP-C). (MyBP Differential expression, conserved domai n structure and chromosome assignment. Eur J Biochem. 1993;216:661–669. doi: 10.1111/j.1432-1033.1993.tb18186.x. [DOI] [PubMed] [Google Scholar]
- 49.Weith A, Sadayappan S, Gulick J, et al. Unique single molecule binding of cardiac myosin binding protein protein-C to actin and phosphorylation-dependent inhibition of actomyosin motility requires 17 amino acids of the motif domain. J Mol Cell Cardiol. 2012;52:219–227. doi: 10.1016/j.yjmcc.2011.09.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Weith AE, Previs MJ, Hoeprich GJ, et al. The extent of cardiac myosin binding protein-C phosphorylation modulates actomyosin function in a graded manner. J Muscle Res Cell Motil. 2012;33:449–459. doi: 10.1007/s10974-012-9312-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Whitten AE, Jeffries CM, Harris SP, Trewhella J. Cardiac myosin myosin-binding protein C decorates F-actin: implications for cardiac function. Proc Natl Acad Sci USA. 2008;105:18360–18365. doi: 10.1073/pnas.0808903105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Yamamoto K. The binding of skeletal muscle C-protein to regulated actin. FEBS Lett. 1986;208:123–127. doi: 10.1016/0014-5793(86)81545-9. [DOI] [PubMed] [Google Scholar]
- 53.Zoghbi ME, Woodhead JL, Moss RL, Craig R. Three-dimensional structure of vertebrate cardiac muscle myosin filaments. Proc Natl Acad Sci USA. 2008;105:2386–2390. doi: 10.1073/pnas.0708912105. [DOI] [PMC free article] [PubMed] [Google Scholar]