

Abstract
Parents know the transformative nature of having and caring for a child. Among many mammals, giving birth leads from an aversion to infant stimuli to irresistible attraction. Here, we review the biological mechanisms governing this shift in parental motivation in mammals. Estrogen and progesterone prepare the uterus for embryo implantation and placental development. Prolactin stimulates milk production, whereas oxytocin initiates labor and triggers milk ejection during nursing. These same molecules, interacting with dopamine, also activate specific neural pathways to motivate parents to nurture, bond with, and protect their offspring. Parenting in turn shapes the neural development of the infant social brain. Recent work suggests that many of the principles governing parental behavior and its effect on infant development are conserved from rodent to humans.
Giving birth is among the most transformative experiences in a parent's lifetime. Furthermore, from the offspring's perspective, the nurturing relationship between parent and infant profoundly affects the development of the brain systems regulating social behavior. Here, we explore the hormonal and neural regulation of mammalian parenting and its consequences for infant social development. The hormones of reproduction (i.e., estrogen, progesterone, oxytocin, and prolactin) create a hospitable intrauterine environment for fetal development, ensure timely birth, and provide sustenance for the infant through lactation, but also orchestrate a set of neural systems to ensure maternal nurturing, bonding, and protection of young. Similar systems along with vasopressin and testosterone influence paternal care in biparental species. Parental nurturing has long-term effects on these same neural systems in infants, resulting in nongenomic transmission of parenting and attachment styles. We review recent studies suggesting that the neural mechanisms regulating parental care and its effect on infant development are notably conserved from rodent to human.
Hormonal synchronization of physiology, brain, and behavior in rodents
Virgin females and males of many species generally avoid infants, finding infant stimuli aversive. Yet parturient mothers typically find infants irresistible and display a suite of maternal nurturing behaviors to ensure survival of their offspring (Fig. 1). For example, virgin female rats avoid or attack pups, but postpartum dams will press a lever more than 100 times per hour to have a pup delivered into their nest box with each press, provided that the number of pups in the nest is maintained below 20. Thus, the onset of maternal care involves a switch in the valence of pup stimuli, resulting from inhibition of avoidance and activation of approach neural systems in response to infant stimuli (1). The power of humoral factors to induce maternal behavior was first illustrated by showing that blood transfusions from a pregnant rat to a virgin female elicited the simultaneous onset of maternal responsiveness in both. Subsequent research revealed that the rise in circulating estrogen and progesterone secreted by the ovaries during pregnancy, followed by the precipitous drop in progesterone at the end of pregnancy, signals that parturition is eminent and maximizes brain sensitivity to oxytocin and prolactin by increasing production of their receptors (Fig. 2).
Fig. 1. Giving birth in mammals leads to a transformation in maternal responsiveness toward infants.
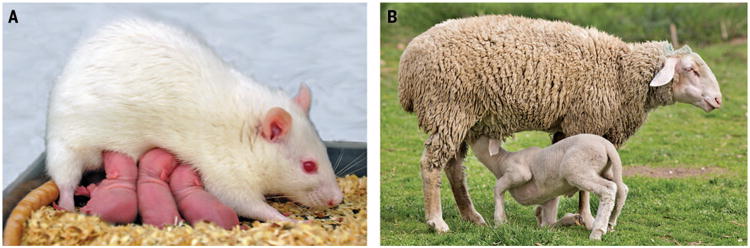
In rats (A), this includes increases in nest building, pup retrieval, nursing, and defense of pups. Although many rodent mothers will care for any pup they encounter, sheep (B) develop selective bonds with their own lambs and reject lambs that are not their own. Experimental research using rodents and sheep have revealed some of the hormonal and neural mechanisms responsible for the onset of maternal behavior. (A) Photo courtesy of Doris Bayerl and Oliver Bosch.
Fig. 2. Schematic illustrating the hormonal and neural synchronization of reproductive physiology and maternal behavior in rodents.
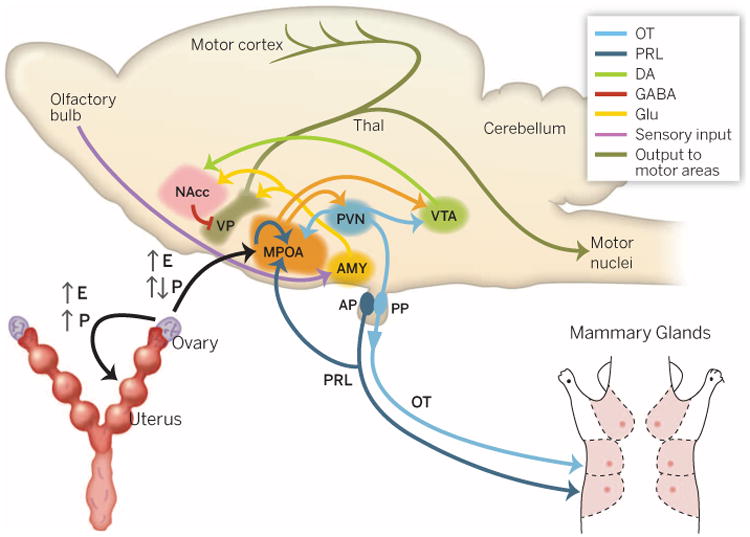
Estrogen (E) and progesterone (P) secreted by the ovaries prepare the uterus for embryonic implantation, support placental development, and sensitize the MPOA to respond to oxytocin (OT) and prolactin (PRL). Prolactin released from anterior pituitary (AP) stimulates milk production in the mammary glands. Oxytocin released from the posterior pituitary (PP) stimulates uterine contractions during labor and milk letdown during nursing. OT, PRL, and dopamine (DA) signaling in the brain modulates communication between several neural pathways to initiate the onset of maternal behavior. GLU, glutamate; NAcc, nucleus accumbens; AMY, amygdala; PVN, paraventricular nucleus of the hypothalamus; VP, ventral pallidum; Thal, thalamus.
The steroid receptor–rich medial preoptic area (MPOA) senses the course of pregnancy by monitoring changes in steroid hormone concentrations, and is likely the region responsible for the transition from pup aversion to attraction at parturition through suppressing amygdala to anterior hypothalamic and enhancing mesolimbic dopaminergic pathways. MPOA neurons are robustly activated by pup stimuli, and destruction of the MPOA abolishes maternal care. Depositing estrogen, oxytocin, prolactin, or dopamine into the MPOA of virgin female rats facilitates maternal responsiveness, demonstrating the pivotal role for this region in synchronizing the onset of maternal behavior with delivery and nursing (2). Elegant molecular genetic studies are beginning to dissect the contributions of specific neural populations within the MPOA for initiating parental care (1).
Dopamine and maternal care in rodents
Infusion of dopamine D1 receptor agonists into the MPOA in the absence of estrogen is sufficient to facilitate maternal behavior, suggesting an intra-MPOA interaction of estrogen and dopamine. However, it is the connection of the MPOA to the mesolimbic dopaminergic ventral tegmental area (VTA) that plays a pivotal role in maternal motivation. Like the MPOA, the VTA is activated by pup stimuli through direct MPOA-VTA connections, leading to increased extracellular dopamine in the nucleus accumbens during maternal care. Dopamine D1 agonists in the nucleus accumbens induce maternal responsiveness without hormonal stimulation. This elevated dopamine in the nucleus accumbens is mediated in part by oxytocin, because females that display higher levels of licking and grooming have higher densities of oxytocin projections to the VTA and elevated dopamine release in the nucleus accumbens in response to pups, which is diminished by oxytocin antagonist in the VTA (2). Thus, the MPOA is the master control region that senses the timing of parturition through the dynamic changes in estrogen, progesterone, oxytocin, and prolactin. At parturition, in response to pup stimuli, the MPOA activates the VTA directly and indirectly through the paraventricular nucleus of the hypothalamus via oxytocin, leading to elevated dopamine in the nucleus accumbens and activating dopamine D1 receptors. This releases the inhibitory control of the ventral pallidum by the nucleus accumbens, allowing excitatory input elicited by pup stimuli from the basolateral amygdala to activate the ventral pallidum. The ventral pallidum is a major output relay of the nucleus accumbens and modulates motor output in response to reinforcing stimuli via projections to the thalamus and cortical and mesencephalic motor nuclei, culminating in the expression of maternal nurturing responses toward pups (Fig. 2).
Neural correlates of human parental care
In recent years, it has been possible to explore the biological correlates of human parenting through a variety of approaches, including brain imaging of the response to infant stimuli, endocrine studies, and gene association studies. Evidence to date suggests that similar mechanisms support animal and human parenting, with a shift to greater involvement of cortical systems in humans.
Neural response to infant and child visual stimuli
Multiple functional magnetic resonance imaging (fMRI) studies report activation in the mesolimbic dopamine system (i.e., VTA, nucleus accumbens, and medial orbitofrontal cortex) as parents view pictures or videos of their children, and these activations are related to positive parenting behaviors (3) (Fig. 3A). For example, fathers who are more involved in instrumental caregiving show stronger VTA activation when viewing pictures of their children. In addition, mothers displaying more coordinated positive engagement and less intrusiveness with their infants more strongly activate the nucleus accumbens when viewing videos of their infants. Finally, mothers who exhibit more praise and positive affect while interacting with their child show stronger medial orbitofrontal cortex activation when viewing pictures of their children. Nonparents can also activate these regions when viewing children, and it may be the appealing facial features of children that drive these activations. In nulliparous women, activation of the nucleus accumbens scales to the degree of “baby schema” (i.e., cuteness) of the child stimuli, as well as with the reported motivation to care for the child. This implies that adult human attraction to infants is less tightly regulated by hormones compared with other species but does not rule out that the hormones of pregnancy intensify the attractiveness of infant stimuli (3–6).
Fig. 3. Paternal brain function in humans revealed by fMRI.
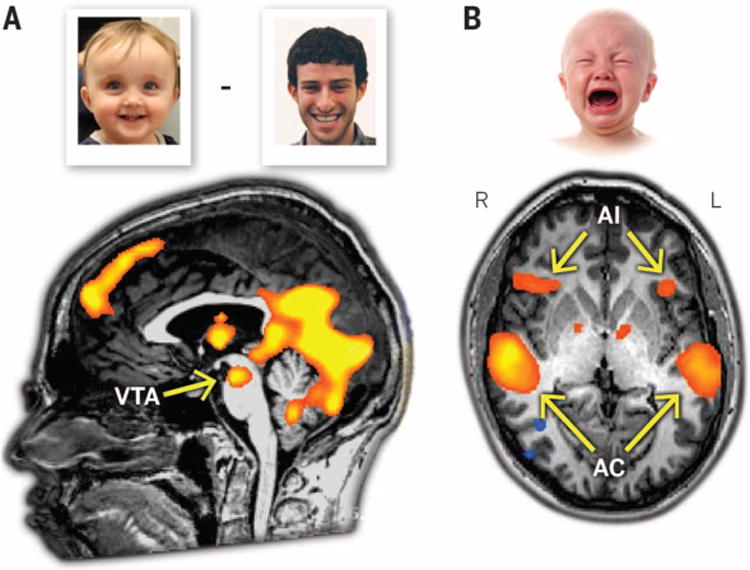
(A) The VTA in fathers is activated to a greater extent when viewing pictures of their own children compared with pictures of unknown adults. (B) The anterior insula in fathers is activated to a greater extent when listening to infant-cry stimuli compared with an auditory control tone. AC, auditory cortex.
Neural response to infant crying
Infants solicit parental caregiving not only by their appearance but also through crying. Infant cries can also activate their parents' mesolimbic dopamine system. However, depressed mothers show attenuated activation to their own infant's cries in the nucleus accumbens, consistent with reduced caregiving motivation. Infant cries also reliably elicit activation in two other brain regions: the anterior insula and the prefrontal cortex. The insula is a visceral somatosensory cortex that represents not only the state of one's own body but also what others are feeling, and the right anterior insula may be where we become aware of those feelings. As such, the anterior insula is critically involved in emotional empathy, and more empathic mothers more strongly activate the anterior insula when viewing pictures of their children. Substance-using mothers show reduced anterior insula activation in response to infant cries, consistent with an attenuated empathic response. Although empathy is essential for parental care, empathic overarousal can lead to distress that interferes with compassionate behavior and effective parenting. For example, in high-risk mothers, stronger anterior insula responses to own-infant cries were related to more intrusive parenting. Fathers also show robust activation of the anterior insula to infant cries (Fig. 3B), and fathers with moderate anterior insula activation are the most involved in instrumental caregiving. Fathers with low and high insula activation may be less involved due to empathic under- and overarousal to cries (3) (7) (8).
The prefrontal cortex is thought to be involved in regulating the initial negative emotional response to infant crying. Frustration induced by inconsolable infant crying is a risk factor for infant abuse (9), highlighting the importance of emotion regulation for sensitive parenting. Recruitment of prefrontal cortex during own-infant crying is associated with increased maternal sensitivity, decreased stress hormone responses to separation, and more secure child attachment behaviors upon reunion (10). Interestingly, depressed mothers and substance-using mothers are less able to engage the prefrontal cortex during infant crying and may therefore be less able to regulate their negative reactions to crying (7,11).
Fewer imaging studies support a role for the MPOA in human parenting, although hypothalamic activation in response to infant stimuli has been reported in a minority of studies (12). It is not clear whether this is due to a diminished role of the MPOA, technical issues with imaging, or because this region is not treated as a separate entity from the hypothalamus in human imaging studies. One possibility is that activation of subpopulations of MPOA neurons generates less fMRI signal as an output node than reflected in downstream targets such as the VTA and nucleus accumbens. In light of its pivotal role in regulating parental behavior in animals, we urge investigators to focus on MPOA/hypothalamus in their analyses.
Oxytocin and parenting in rodents and sheep
Oxytocin has received considerable attention in recent years for modulating many aspects of social relationships, including parenting and social bonding. Disrupting brain oxytocin signaling pharmacologically or genetically disrupts maternal behavior (13). CD38 knockout mice with impaired oxytocin secretion display disrupted maternal care, which is restored by subcutaneous injection of oxytocin (13). Whereas the role of peripheral oxytocin signaling in initiating maternal responsiveness has largely been unexplored, most, but not all, studies suggest that oxytocin signaling in the brain facilitates the onset— and to a lesser degree the maintenance of—maternal responsiveness in rodents. Oxytocin released centrally during parturition and nursing appears to play a role in the transition toward approach behaviors in mothers. Rodent mothers are promiscuously maternal, nurturing any pup they encounter; however, in herding animals like sheep, strong and selective mother-infant bonds are formed. Oxytocin signaling is necessary and sufficient to form a selective bond between a ewe and a lamb (13). Thus oxytocin affects not only maternal motivation but also the formation of mother-infant attachment where it exists. Oxytocin, along with the related peptide vasopressin, also elicits protective maternal aggression toward intruders (14). The essential roles of oxytocin in generating nurturing, bonding, and infant defense in mothers may be the evolutionary antecedent to the more generalized roles of this peptide in other social contexts, including pair-bonding behavior, empathy, trust and in-group favoritism.
Oxytocin and human parenting
There is considerable evidence that oxytocin modulates human parenting from three experimental approaches. The first involves correlating parental behavior and peripheral oxytocin concentrations. Although peripheral oxytocin is less likely to be relevant to behavior than central measures, it is not feasible to measure central oxytocin levels noninvasively in humans. The degree to which peripheral oxytocin is correlated with oxytocin activity in the brain is a point of controversy, as are the methods for sample processing and the assays used. Nevertheless, we discuss studies relating peripheral oxytocin and behavior, with the caveat that little rigorous evidence supports a relationship between circulating plasma, salivary, or urinary oxytocin and brain oxytocin activity, although dynamic changes in peripheral oxytocin could possibly parallel central oxytocin release (13).
In both mothers and fathers, plasma oxytocin is positively correlated with affectionate contact (15) and coordinated positive engagement with infants during dyadic interactions (16). Both of these parental behaviors are important for healthy psychosocial development (17). Depressed mothers, who may be less responsive to their children, have lower salivary oxytocin than non-depressed mothers (18). Baseline plasma oxytocin concentrations in fathers, but not mothers, correlate with stimulatory parent-infant contact such as proprioceptive touch, stimulatory touch, and exploratory play (16). Thus, oxytocin may facilitate sexually differentiated styles of parent-infant interaction that could support different aspects of healthy child development.
Intranasal oxytocin administration has become popular for exploring the role of oxytocin on parenting. These studies should, however, be interpreted with caution with regard to brain mechanisms because nasally administered oxytocin elevates plasma oxytocin significantly, with only modest evidence for elevations within the brain (19–21). Most studies have been conducted in fathers due to concerns with administering oxytocin to lactating mothers. In fathers, intranasal oxytocin increases stimulatory and exploratory play with toddlers and increases the duration of episodes of father-infant touch and social reciprocity. These augmented paternal behaviors in turn increase the duration of episodes of infant gaze to the father and infant object manipulation, as well as infant salivary oxytocin (16). Notably, paternal head speed and acceleration during father-infant interactions are also increased by intranasal oxytocin and positively correlated with infant salivary oxytocin (22). Overall, oxytocin seems to motivate paternal behaviors that facilitate father-infant bonding. Intranasal oxytocin also decreases paternal hostility during interactions with toddlers and decreases frustration in response to infant cries among nulliparous women (3, 12, 16). Finally, in parallel with animal studies where oxytocin supports maternal aggression, intranasal oxytocin increases maternal protective responses in the presence of a socially intrusive stranger among women with postpartum depression (23).
Genetic evidence also supports a role for oxytocin in human parenting. Polymorphisms in genes encoding the oxytocin receptor (OXTR) and CD38 have been reported to predict parenting behaviors. The G/G genotype of OXTR rs53576 is associated with increased maternal sensitivity toward toddlers at risk for externalizing behavior problems and with a more pronounced heart-rate response to infant cries (24). In another study, OXTR rs2254298 and rs1042778 and CD38 rs3796863 risk alleles were each associated with lower plasma oxytocin. Reduced plasma oxytocin and both OXTR and CD38 risk alleles were related to less parental touch (16). Moreover, a recent study showed that children with two A alleles at rs53576 tend to exhibit more negative emotionality, which in turn partially explains some aspects of parental behavior (25). Thus, associations between rs53576 genotype and parental behavior could be partially mediated by genetically influenced child temperament and behavior. These genetic studies, although intriguing, have small samples and await replication.
Oxytocin modulation of the neural response to infants and children
Research in animals suggests that oxytocin acts in the MPOA and VTA to activate the mesolimbic dopamine approach system and inhibit amygdala-based avoidance, rendering infant stimuli reinforcing rather than aversive. A similar mechanism may be operational in humans. Plasma oxytocin is positively correlated with nucleus accumbens response to viewing pictures or videos of one's own children. Intranasal oxytocin also attenuates the amygdala response to unknown infant cries among nulliparous women (26), consistent with inhibition of an avoidance pathway. The same study also found that intranasal oxytocin enhanced the anterior insula response to unknown infant cries, suggesting that oxytocin may also enhance empathic responses to unknown infant cries. Although animal research has emphasized the role of oxytocin in the onset of parental behavior, the human studies discussed above instead generally demonstrate a role for oxytocin in the maintenance of parental behavior. Two recent human studies suggest that peripartum exposure to oxytocin may alter maternal neural responses to infant stimuli. Compared with nonbreastfeeding mothers, breastfeeding mothers more strongly activate the insula and prefrontal cortex in response to own-infant cries, and mothers who deliver their babies vaginally have a stronger insula response to own-infant cries than do mothers who deliver by Caesarean section (3). Collectively, these neuroimaging data are consistent with the above findings, suggesting that oxytocin supports sensitive caregiving.
Regulation of paternal care in rodents
The regulation of paternal care in mammals has received less attention than maternal care, primarily because of its rarity in rodent models, although there appear to be common elements with maternal care (1). In some species, testosterone is necessary for the maintenance of paternal care, perhaps by its conversion to estrogen in the brain (27). However, in several biparental rodent and primate species, the onset of paternal care is associated with a decrease in testosterone and an increase in prolactin. However, there is little evidence of a causal relationship between elevated prolactin concentrations and paternal behavior in any mammalian species (28).
The role of oxytocin in nonhuman mammalian paternal behavior has received little attention, although vasopressin appears to be involved. In monogamous male prairie voles, mating decreases vasopressin fiber content in the septum and increases vasopressin synthesis, consistent with intraseptal vasopressin release. Furthermore, infusion of a vasopressin V1a receptor antagonist into the septum disrupts paternal care in voles (29).
Testosterone, testes size, and paternal caregiving in humans
In many species, testosterone supports mating effort at the expense of parenting effort. This may also be true of human males. Men with higher testosterone report less sympathy toward an unknown newborn infant cry, testosterone decreases when men become involved fathers, and fathers experiencing larger decreases in testosterone report less sexual intercourse (30). Moreover, among fathers, those with higher testosterone are less involved in paternal caregiving (4) and less responsive to infants (31). Low testosterone may facilitate paternal caregiving by allowing more empathy for the child, by increasing frustration tolerance, or by decreasing sexual motivation that could compete with parenting effort. Although married fathers have an attenuated nucleus accumbens response to visual sexual stimuli compared with unmarried nonfathers, this response was not correlated with testosterone levels. However, testosterone was negatively correlated with activation in face-emotion processing regions when viewing pictures of unknown children, consistent with a negative effect on neural systems involved with empathy (32).
Testes size is also correlated with increased investment in mating, both across and within nonhuman species (4). In human fathers, testes size has a weak but significant negative correlation with instrumental caregiving, such that men with smaller testes are more involved. Testes size also has a robust inverse correlation with the VTA activation in response to viewing pictures of one's own child (Fig. 4), implying stronger motivation to approach children in fathers with smaller testes (4).
Fig. 4. Relationship between testes volume and VTA fMRI signal in fathers in response to viewing pictures of their own children.
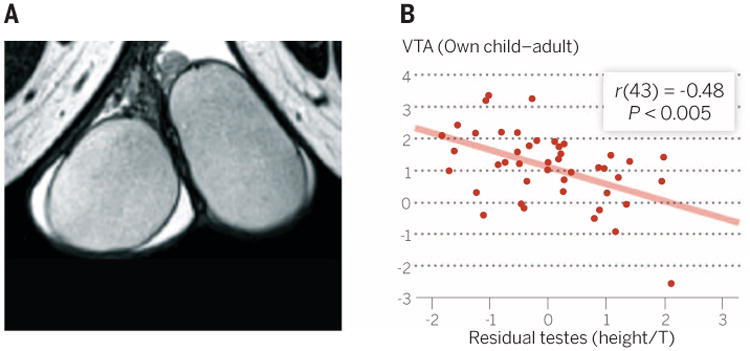
(A) Structural MRI was used to estimate testicular volume in fathers. (B) VTA activation in fathers while viewing pictures of their own children is negatively correlated with testicular volume, controlling for both height and testosterone level. Adapted from (4).
The number of CAG repeats in exon 1 of the androgen receptor gene (AR) is inversely correlated with AR expression. Fathers with more CAG repeats, and presumably fewer AR, have stronger anterior insula response to infant cries (8). This suggests that fathers with less sensitivity to androgens have a more empathic response to infant cries.
Effect of parenting on social development in rodents
Variation in parental nurturing affects the developing offspring's brain, affecting future social behaviors. Rat dams vary in the extent to which they lick and groom their pups. Notably, pups reared by low licking and grooming mothers display low licking and grooming when they become mothers, regardless of the maternal style of their biological mothers. This nongenomic transmission of maternal style is mediated by alterations in estrogen receptor and oxytocin receptor expression (33). High licking and grooming mothers have higher densities of estrogen receptor in the MPOA than low licking and grooming mothers as a result of differential methylation of the estrogen receptor (ERα) promoter. This lower estrogen receptor density leads to decreased sensitivity to estrogen and thus lower oxytocin receptor transcription in the MPOA. Being reared by a low licking and grooming mother also significantly alters several aspects of the mesolimbic dopamine system through adulthood (34). Parenting also has life-long effects on the oxytocin system in primates, as rhesus macaques raised by human caregivers have lower central oxytocin than mother-reared animals (13).
Variation in parental nurturing can affect other social behaviors as well. Repeated neonatal social isolations disrupt later-life pair bonding behaviors in monogamous voles (35). In contrast, pharmacologically stimulating oxytocin neurons with neonatal melanocortin agonists facilitates later-life pair bonding (36). Paternal care can also affect rodent social development and parenting style. Monogamous prairie vole pups of both sexes raised in the absence of the father show impairments in adult pair-bonding behavior and lower levels of licking and grooming compared with biparentally reared animals (37). Likewise, paternal licking and grooming style is nongenomically transmitted from father to son in the monogamous, biparental California mouse (27).
Effect of parenting on neural and social development in humans
A considerable body of work has investigated alterations in brain development in children initially raised without parents in orphanages who were subsequently adopted into stable families. The observed alterations have been focused on the amygdala and prefrontal cortex (38–40). Postinstitutionalized children have larger amygdala volumes than children not raised in orphanages, and amygdala volume is positively correlated with both anxiety symptoms and internalizing problems. Postinstitutionalized children also exhibit increased amygdala responses to fearful faces. Typically developing children have a stronger amygdala response to pictures of their mothers compared with female strangers, which presumably reflects the special affective salience of the mother. In contrast, amygdala response to strangers matches that to the mother among postinstitutionalized children, who have an increased tendency to approach unfamiliar adults. Finally, postinstitutionalized children show altered connectivity between the amygdala and medial prefrontal cortex, a key emotion-regulation pathway. We cannot be certain that parental deprivation is the primary contributor to these altered pathways, but the overlap with findings from experimental studies in rodents supports this possibility (38, 39).
As described in rodents, the effect of variation in parenting on offspring development may be partially mediated by oxytocin in humans. For example, women experiencing childhood neglect or abuse have decreased oxytocin concentrations in cerebrospinal fluid as adults (13). Additionally, children previously raised in orphanages have an attenuated urinary oxytocin response to interactions with their mothers, although the caveats mentioned above regarding peripheral oxytocin and assay techniques must be considered (41). Furthermore, G/G genotype carriers at OXTR rs53576 have a greater risk of displaying depressive symptomology and emotional dysregulation in response to childhood maltreatment but are also more likely to benefit from positive family environments than are A/A genotype carriers (42, 43). Finally, attachment styles are often transmitted across generations, and oxytocin may be involved. Insecurely attached mothers have an attenuated plasma oxytocin response to interactions with their children (12), plasma oxytocin concentrations are positively correlated with parental affection toward infants (18), and infants respond to this affection with parent-directed behaviors that support parent-infant bonding (44). Thus, high levels of oxytocin in securely attached parents may facilitate greater affectionate behavior toward the child, who consequently becomes more securely attached to the parent (Fig. 5).
Fig. 5. Postulated mechanisms supporting the intergenerational transmission of secure attachment in humans.
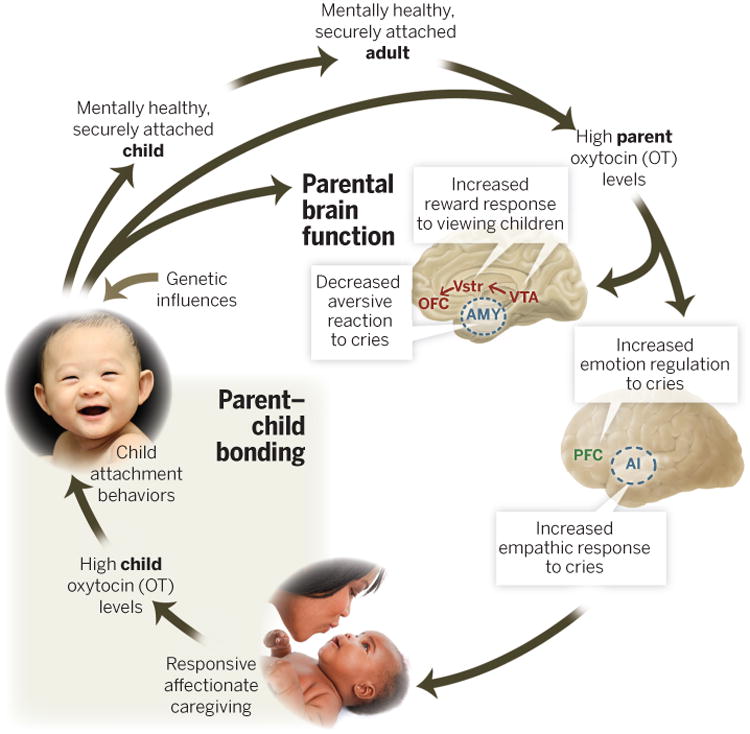
Securely attached parents have higher baseline oxytocin (OT) and a more pronounced OT response to interactions with their children. OT augments the mesolimbic DA system response to visual child stimuli, enhancing their reward value. It also inhibits AMY and augments anterior insula (AI) responses to infant-cry stimuli, facilitating a more empathic reaction. OTmay also modulate prefrontal cortex activity to suppress negative emotional reactions to infant crying. These neurobiological influences promote responsive, affectionate caregiving, which in turn promotes OT activity in children, along with child attachment behaviors that further engage parental brain systems, resulting in a positive feedback cycle that culminates in a mentally healthy, securely attached child. Genetic influences of the child can also either support or interfere with child attachment behaviors and parent-offspring bonding. OFC, orbitofrontal cortex; PFC, prefrontal cortex.
Conclusion
The human parenting experience is likely to have some unique features relative to other mammals by virtue of the increased cortical complexity and control over emotion and behavior in our species. However, there is now strong evidence that human and animal parenting share many subcortical neural and neurochemical mechanisms. In rodents, the MPOA likely plays a primary role in the dramatic transformation of the maternal brain in response to hormonal changes though inhibiting avoidance pathways and activating mesolimbic dopamine-approach pathways. In humans, these mechanisms may modulate parental sensitivity, motivation, and drive, whereas higher-order cortical, hormone-independent mechanisms play a more prominent role in cognitively enriching the parental experience. As technology progresses, we will undoubtedly gain a deeper understanding of how neurochemistry and brain systems influence mammalian and human parenting, as well as how parental nurturing shapes the social neural systems of our offspring. We will likely discover even more parallels in the regulation of parenting and its consequences between rodents and humans. Perhaps this understanding will lead to new efforts to systematically improve parenting in all facets of society to benefit generations to come.
Acknowledgments
The authors acknowledge the support of a Positive Neuroscience Award from the John Templeton Foundation to J.K.R., National Institute of Mental Health (NIMH) R01MH096983 to L.J.Y., and NIMH 1P50MH100023 to J.K.R. and L.J.Y., as well as NIH OD P51OD11132 to the Yerkes National Primate Research Center during the writing of this manuscript.
References and Notes
- 1.Dulac C, O'Connell LA, Wu Z. Science. 2014;345:765–770. doi: 10.1126/science.1253291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Stolzenberg DS, Numan M. Neurosci Biobehav Rev. 2011;35:826–847. doi: 10.1016/j.neubiorev.2010.10.003. [DOI] [PubMed] [Google Scholar]
- 3.Rilling JK. Neuropsychologia. 2013;51:731–747. doi: 10.1016/j.neuropsychologia.2012.12.017. [DOI] [PubMed] [Google Scholar]
- 4.Mascaro JS, Hackett PD, Rilling JK. Proc Natl Acad Sci U S A. 2013;110:15746–15751. doi: 10.1073/pnas.1305579110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Michalska KJ, et al. Front Behav Neurosci. 2014;8:21. doi: 10.3389/fnbeh.2014.00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Glocker ML, et al. Proc Natl Acad Sci U S A. 2009;106:9115–9119. doi: 10.1073/pnas.0811620106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Landi N, et al. Front Psychiatry. 2011;2:32. doi: 10.3389/fpsyt.2011.00032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mascaro JS, Hackett PD, Gouzoules H, Lori A, Rilling JK. Soc Cogn Affect Neurosci. 2013 doi: 10.1093/scan/nst166. 10.1093/scan/nst166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Barr RG. Proc Natl Acad Sci U S A. 2012;109(suppl. 2):17294–17301. doi: 10.1073/pnas.1121267109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Laurent HK, Ablow JC. Infant Behav Dev. 2012;35:761–772. doi: 10.1016/j.infbeh.2012.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Laurent HK, Ablow JC. Soc Cogn Affect Neurosci. 2012;7:125–134. doi: 10.1093/scan/nsq091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Strathearn L, Fonagy P, Amico J, Montague PR. Neuropsychopharmacology. 2009;34:2655–2666. doi: 10.1038/npp.2009.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ross HE, Young LJ. Front Neuroendocrinol. 2009;30:534–547. doi: 10.1016/j.yfrne.2009.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bosch OJ, Neumann ID. Horm Behav. 2012;61:293–303. doi: 10.1016/j.yhbeh.2011.11.002. [DOI] [PubMed] [Google Scholar]
- 15.Apter-Levi Y, Zagoory-Sharon O, Feldman R. Brain Res. 2013 doi: 10.1016/j.brainres.2013.10.052. 10.1016/j.brainres.2013.10.052. [DOI] [PubMed] [Google Scholar]
- 16.Feldman R. Horm Behav. 2012;61:380–391. doi: 10.1016/j.yhbeh.2012.01.008. [DOI] [PubMed] [Google Scholar]
- 17.Feldman R. J Child Psychol Psychiatry. 2007;48:329–354. doi: 10.1111/j.1469-7610.2006.01701.x. [DOI] [PubMed] [Google Scholar]
- 18.Apter-Levy Y, Feldman M, Vakart A, Ebstein RP, Feldman R. Am J Psychiatry. 2013;170:1161–1168. doi: 10.1176/appi.ajp.2013.12121597. [DOI] [PubMed] [Google Scholar]
- 19.Modi ME, Connor-Stroud F, Landgraf R, Young LJ, Parr LA. Psychoneuroendocrinology. 2014;45:49–57. doi: 10.1016/j.psyneuen.2014.02.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Striepens N, et al. Sci Rep. 2013;3:3440. doi: 10.1038/srep03440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Neumann ID, Maloumby R, Beiderbeck DI, Lukas M, Landgraf R. Psychoneuroendocrinology. 2013;38:1985–1993. doi: 10.1016/j.psyneuen.2013.03.003. [DOI] [PubMed] [Google Scholar]
- 22.Weisman O, et al. Biol Lett. 2013;9:20130828. doi: 10.1098/rsbl.2013.0828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mah BL, Bakermans-Kranenburg MJ, Van Ijzendoorn MH, Smith R. Depress Anxiety. 2014 doi: 10.1002/da.22245. 10.1002/da.22245. [DOI] [PubMed] [Google Scholar]
- 24.Riem MM, Pieper S, Out D, Bakermans-Kranenburg MJ, van Ijzendoorn MH. Soc Cogn Affect Neurosci. 2011;6:294–300. doi: 10.1093/scan/nsq035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kryski KR, Smith HJ, Sheikh HI, Singh SM, Hayden EP. Pers Individ Dif. 2014;64:107–110. [Google Scholar]
- 26.Riem MM, et al. Biol Psychiatry. 2011;70:291–297. doi: 10.1016/j.biopsych.2011.02.006. [DOI] [PubMed] [Google Scholar]
- 27.Gleason ED, Marler CA. Proc Biol Sci. 2013;280:20130824. doi: 10.1098/rspb.2013.0824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ziegler TE, Prudom SL, Zahed SR, Parlow AF, Wegner F. Horm Behav. 2009;56:436–443. doi: 10.1016/j.yhbeh.2009.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang Z, Ferris CF, De Vries GJ. Proc Natl Acad Sci U S A. 1994;91:400–404. doi: 10.1073/pnas.91.1.400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gettler LT, McDade TW, Agustin SS, Feranil AB, Kuzawa CW. Horm Behav. 2013;64:755–763. doi: 10.1016/j.yhbeh.2013.08.019. [DOI] [PubMed] [Google Scholar]
- 31.Weisman O, Zagoory-Sharon O, Feldman R. Prog Neuropsychopharmacol Biol Psychiatry. 2014;49:47–52. doi: 10.1016/j.pnpbp.2013.11.006. [DOI] [PubMed] [Google Scholar]
- 32.Mascaro JS, Hackett PD, Rilling JK. Psychoneuroendocrinology. 2014;46:153–163. doi: 10.1016/j.psyneuen.2014.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Champagne FA. Horm Behav. 2011;60:4–11. doi: 10.1016/j.yhbeh.2011.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Peña CJ, Neugut YD, Calarco CA, Champagne FA. Eur J Neurosci. 2014;39:946–956. doi: 10.1111/ejn.12479. [DOI] [PubMed] [Google Scholar]
- 35.Yu P, et al. Psychoneuroendocrinology. 2013;38:3128–3138. doi: 10.1016/j.psyneuen.2013.09.012. [DOI] [PubMed] [Google Scholar]
- 36.Barrett CE, et al. Neuropsychopharmacology. 2013;85:357–366. [Google Scholar]
- 37.Ahern TH, Hammock EA, Young LJ. Dev Psychobiol. 2011;53:118–131. doi: 10.1002/dev.20498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tottenham N, Shapiro M, Telzer EH, Humphreys KL. Dev Sci. 2012;15:307–319. doi: 10.1111/j.1467-7687.2011.01128.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gee DG, et al. Proc Natl Acad Sci U S A. 2013;110:15638–15643. doi: 10.1073/pnas.1307893110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hanson JL, et al. Child Dev. 2013;84:1566–1578. doi: 10.1111/cdev.12069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wismer Fries AB, Ziegler TE, Kurian JR, Jacoris S, Pollak SD. Proc Natl Acad Sci U S A. 2005;102:17237–17240. doi: 10.1073/pnas.0504767102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bradley B, Davis TA, Wingo AP, Mercer KB, Ressler KJ. Eur J Psychotraumatol. 2013;4:21659. doi: 10.3402/ejpt.v4i0.21659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.McQuaid RJ, McInnis OA, Stead JD, Matheson K, Anisman H. Front Neurosci. 2013;7:128. doi: 10.3389/fnins.2013.00128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Weisman O, Zagoory-Sharon O, Feldman R. Biol Psychiatry. 2012;72:982–989. doi: 10.1016/j.biopsych.2012.06.011. [DOI] [PubMed] [Google Scholar]