

Abstract
Recombinant adenovirus (Ad) vectors are widely used for both in vitro and in vivo gene transfer. However, intravenous administration of Ad vectors results mainly in hepatocyte transduction and subsequent hepatotoxicity. Coxsackie-adenovirus receptor (CAR) and αvβ integrins, which are functional receptors for the fiber and penton proteins, respectively, are the tropism determinants of Ad type 5 (Ad5). We previously developed a system for rapid construction of fiber-modified Ad5 vectors. We also constructed a fiber-modified Ad5 containing an Arg-Gly-Asp (RGD) motif in the HI-loop and showed that it could enhance anti-tumor effects in vitro and in vivo. Here, we constructed a novel Ad5 vector containing two amino acid mutations in the AB loop of the fiber-modified Ad5 fiber knob and showed that it could significantly reduce liver tropism and increase gene transfer in low-CAR or CAR-deficient cancer cells following intravascular delivery. However, anti-tumor effects of the fiber-mutated Ad5 expressing HSV-TK under control of the hTERT promoter was not found when compared with an unmodified Ad5 vector in cancer lines expressing different levels of CAR, likely due to the activity of the hTERT promoter being lower than that of the CMV promoter. Nevertheless, this study describes an enhanced Ad5 vector for intravascular gene delivery, and further modifications such as changes in the promoter may facilitate the development of this vector for cancer treatment.
Keywords: Adenovirus, gene therapy, coxsackie-adenovirus receptor, liver tropism
Introduction
Human adenoviruses (Ad) are currently the most widely used viral vectors in gene therapy clinical trials due to their well-characterized biology, large transgene capacity, ability to transfer genes to a wide variety of cell types, amplification to high titers and relatively safe profile for use in humans [1]. Currently, 57 human Ad serotypes are defined and distributed into seven species, A to G, of which the most commonly used for cancer gene therapy is species C adenovirus, HAdV-C5 (Ad5) [2]. Systemic or intravenous (IV) administration is the ideal mode of delivery for Ad5-based cancer therapy. However, effective IV administration of Ad5-based vectors has been hindered by the innate liver tropism of Ad5 and concerns of hepatotoxicity. Liver tropism causes the Ad5 vector to be diverted away from the desired tumor targets and to be rapidly cleared from the body, thereby necessitating higher doses of administered vector and resulting in poor antitumor efficacy [3].
The mechanisms of systemic Ad administration that leads to efficient liver tropism and transduction remain controversial. Infectivity of Ad5, at least in vitro, is known to be mediated via a primary cell tethering interaction between the Ad5 fiber knob domain and the coxsackie and adenovirus receptor (CAR) [4], followed by a secondary, endocytosis-stimulating interaction between Arg-Gly-Asp (RGD) tri-peptide motifs in the Ad penton-base protein and αvβ3 and αvβ5 cellular integrins [5]. In adult humans, CAR is highly expressed in different tissues, including the liver. Its distribution is comparable to that in adult mice, except that higher levels are expressed in the mouse liver. However, the introduction of point mutations in the Ad5 fiber knob domain to abrogate CAR binding was shown to have a profound impact on transduction efficiency in vitro, but it did not necessarily have an apparent effect on levels of liver transduction when introduced systemically in vivo [6,7].
Ablating Ad5 liver tropism only partially contributes to improving the tumor targeting of vectors. In this context, Ad5-based vectors selectively transduce target cells via binding of the Ad fiber protein to CAR. However, CAR is poorly expressed on many tumor targets, thereby limiting transduction [8]. Prior studies showed that RGD-integrin interactions could be used as an alternative infection pathway and could dramatically improve the ability of the Ad vector to transduce several types of cells [9]. In a previous study, we constructed a fiber-modified Ad5 containing an RGD motif in the HI-loop and demonstrated that it could enhance anti-tumor effects in vitro and in vivo [10].
In the current study, a modified vector was made using this developed system for rapid construction of recombinant fiber-modified Ad5 vectors, which can be generated by in vitro ligation and easily monitored by a reporter gene in vitro and in vivo. This novel Ad5 vector containing two amino acid mutations in the AB loop of the fiber-modified Ad5 fiber knob with the aim of reducing liver tropism and increasing the anti-tumor efficacy of the vector in CAR-deficient cancers.
Materials and methods
Cell culture
Human embryonic kidney HEK293 cells were purchased from the American Type Culture Collection (Manassas, VA, USA). HL7702 (human normal liver), A549 (human lung adenocarcinoma), Caco-2 (human colorectal adenocarcinoma) and Skov3 (human ovarian cancer) cells were obtained from the Cell Bank of the Chinese Academy of Science (Shanghai, China). All cell lines were cultured in compatible media supplemented with 10% fetal bovine serum (FBS) (Invitrogen, Carlsbad, CA, USA), penicillin (50 U/ml) and streptomycin (50 U/ml) in the presence of 5% CO2.
Flow cytometry analysis
Expression levels of CAR, αvβ3 and αvβ5 in cells were determined by flow cytometry analysis with a MoFlo XDP cell sorter (Beckman Coulter, Brea, CA, USA). Cultured cells were washed and harvested with PBS, resuspended in PBS containing 1% bovine serum albumin (BSA) for 30 min at room temperature and then incubated with primary antibodies for 1 h on ice. Subsequently, the cells were washed and incubated with FITC-conjugated IgG for an additional hour. After washing with 1% BSA/PBS, the cells were analyzed by flow cytometry. The same secondary antibodies alone served as the negative control for each cell line. Primary antibodies specific for CAR (Rmcb, Millipore, Bedford, MA, USA), anti-αvβ3 mAb LM609 and anti-αvβ5 mAb P1F6, were purchased from Millipore. The secondary antibody was FITC-conjugated rabbit anti-mouse IgG (Dingguo, Beijing, China).
Construction and production of recombinant Ad5 vectors
Ad5-Luc, Ad5-RGD-Luc, Ad5-hT-TK and Ad5-RGD-hT-TK were previously constructed in our lab [10,11]. Ad5-MU-RGD-Luc and Ad5-MU-RGD-hT-TK also were constructed as previously described [10]. First, site-directed mutagenesis was performed to change Ser (408) and Pro (409) to Glu and Ala in the fiber knob domain of Ad5. Primers, 5’-CCACACCAGCTCCAGAGGCCAACTGTAGACTAAATGCA-3’ (forward) and 5’-TGCATTTAGTCTACAGTTGGCCTCTGGAGCTGGTGTGGT-3’ (reverse) were used to introduce the mutations based on the pT-SM-RGD plasmid, resulting in the pT-SM-RGD-MU plasmid. The knob region in PT-SX was subsequently replaced with the knob-RGD-MU region in pT-SM-RGD-MU by using the Stu I and Mfe I sites to obtain pT-SX-RGD-MU. Finally, the knob region in pAd-Luc was replaced with the knob-RGD-MU region in pT-SX-RGD-MU with the use of the Spe I and Xba I sites to generate pAd-MU-RGD-Luc.
The recombinant Ad plasmid pAd-hT-TK previously constructed in our lab contains the HSV-TK gene under the control of the hTERT promoter in the E1/E3 deletion region. The plasmid pAd-hT-TK was digested with I-Ceu I and PI-Sce I, and then ligated with the I-Ceu I and PI-Sce I-digested pAd-MU-RGD-Luc to generate the pAd-MU-RGD-hT-TK plasmid. The recombinant Ad5 vectors (Ad5-MU-RGD-Luc and Ad5-MU-RGD-hT-TK) were amplified in HEK293 cells and purified by CsCl gradient centrifugation. The titers were determined by the 50% tissue culture infectious dose (TCID50) method. The negative control Ad5-Blank contained the Ad5 genome with no insert in the E1/E3 deletion region (Newgene, Shanghai, China).
Identification of recombinant Ad5 vectors
Cells were seeded at 5 × 105 cells per well in six-well plates. Twenty-four hours later, the cells were infected with recombinant Ad at the multiplicity of infection (MOI) of 50 and then harvested at 48 h post-infection. Viral DNA was extracted from crude lysates using the QIAamp DNA Mini Kit (Qiagen, Valencia, CA, USA) following the manufacturer’s instructions and then analyzed by sequencing (Sangon Biotech, Shanghai, China), restriction enzyme digestion and PCR. To analyze the expression of foreign genes, cells lysates were suspended in a low salt extraction buffer and centrifuged (3,000 rpm for 5 min). The supernatants were collected as samples and mixed with an equal volume of 2 × SDS buffer (125 mM Tris-HCl, pH 6.8; 4% SDS; 20% glycerol; 0.01% bromophenol blue; and 10% β-mercaptoethanol) and boiled for 30 min. The samples were loaded onto a 12% SDS-polyacrylamide gel for electrophoresis and then transferred to a nitrocellulose membrane. The primary TK antibodies were rabbit anti-sera produced by our laboratory raised against full-length HSV-TK protein, and the luciferase antibodies were purchased from Abcam (Cambridge, MA, USA). The secondary antibody was an AP-labeled anti-rabbit antibody (Jackson ImmunoRresearch Laboratories, West Grove, PA, USA).
Ad5 infection assay and cytotoxicity assay in vitro
To assess the infectivity of the fiber-modified Ad5, experimental cells were seeded into 24-well plates (1 × 105 cells/well) and incubated overnight to allow adherence. Viruses (Ad5-Blank, Ad5-Luc, Ad5-RGD-Luc or Ad5-MU-RGD-Luc) were used to infect the cells at different MOIs. The infected cells were incubated at 37°C in 5% CO2 for 3 h, and the medium was then replenished with medium containing 2% FBS. Forty-eight hours later, the cells were rinsed with PBS and assayed for luciferase activity used the Dual-Glo Luciferase Assay System (Promega, Madison, WI, USA) with a Fluoroskan Ascent FL luminometer (Thermo Scientific, Waltham, MA, USA).
To assess the cytotoxicity of the suicide genes in Ad5, experimental cells (5 × 103 cells) were seeded into a 96-well plate overnight before infection with Ad5-Blank, Ad5-hT-TK, Ad5-RGD-hT-TK or Ad5-MU-RGD-hT-TK at MOI = 5 for 3 h, followed by culture with the prodrug ganciclovir (GCV) at various concentrations (0.1, 1, 10 or 100 μM). On the fourth day, cell viability was determined by the MTT assay. MTT (5 mg/ml) was added to each well, and the cells were incubated for another 4 h. The medium containing MTT was then discarded, and 150 μL DMSO was added to each well and shaken to dissolve the crystals. Absorbance (A) values were measured at OD490 nm. Cell survival rates were calculated according to the following formula: survival = [(average A value of experimental group - average A value of control group)/(average A value of positive group - average A value of control group)] × 100%, where the control group includes blank wells, and the positive group is comprised of cells cultured with medium containing serum only.
Liver tropism analysis
For liver tropism studies, five-week-old BALB/c (-/-) nude mice were injected with 1 × 109 plaque forming units (PFU)/mouse of Ad5-luc, Ad5-RGD-luc or Ad5-MU-RGD-luc via the tail vein. After 48 h, mice were anesthetized and injected with D-luciferin approximately 10 min before imaging with a Kodak In-Vivo Multispectral Imaging System. Thereafter, mice were sacrificed and livers harvested. DNA was isolated using the Qiagen DNeasy Tissue Kit. Quantitative PCR was performed on DNA samples using primers specific for the luciferase transgene. All animal procedures were approved by the Jilin University Institutional Animal Care and Use Committee (approval no. 2012011203).
Statistical analysis
One-way ANOVA followed by Fisher’s least significant difference (LSD) test was employed using SPSS 13.0 software. All analyses were performed using GraphPad Prism 5.0, and all results are presented as means ± SEM. A threshold of P < 0.05 was defined as statistically significant.
Results
Construction and identification of fiber-mutated Ad5 vectors
In our recent studies, we built a simplified Ad5 vector platform for fiber modification and quick detection of gene transfer, and a fiber-modified Ad vector containing an RGD motif in the HI loop of the fiber knob was also constructed. Therefore, we could expediently construct a novel fiber-mutated Ad5 vector, Ad5-MU-RGD, based on Ad5-RGD vectors as described in Materials and Methods (Figure 1A). In order to identify the fiber-mutated Ad5 vectors, viral DNA was extracted from Ad5-RGD and Ad5-MU-RGD infected cells and analyzed by sequencing, PCR and restriction enzyme digestion with Hind III. These results showed that the fiber-mutated Ad5 with the complete genome was obtained (Figure 1B and 1C). Western blot analysis also was performed to analyze the protein expression from fiber-mutated Ad5 vectors. Compared with Ad5-RGD, Ad5-MU-RGD showed a lower protein expression level under the control of either the CMV or hTERT promoter (Figure 1D).
Figure 1.
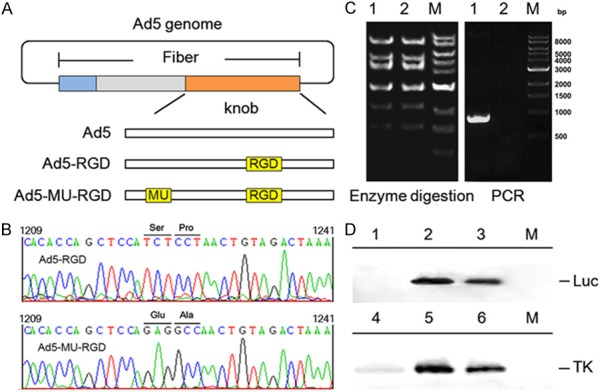
Construction and identification of fiber-mutated Ad5 vectors. A. schematic representation of Ad5 vectors carrying mutations and RGD peptide in the loops of fiber knob. B. sequencing chomatograms of the mutations (S408E, P409A) in the fiber knob. C. agarose gel electrophoresis of viral genome after restriction enzyme digestion with Hind III and PCR using specific primers. Lane 1: Ad5-MU-RGD; lane 2: Ad5-RGD; M: 1 Kb DNA ladder. D. western blot analysis of luciferase expression in 293 cells (upper) and HSV-TK expression in A549 cells (lower) after infection with Ad5 vectors. Lane 1: 293 cells; lane 2: Ad5-RGD-Luc infected 293 cells; lane 3: Ad5-MU-RGD-Luc infected 293 cells; lane 4: A549 cells; lane 5: Ad5-RGD-Luc infected A549 cells; lane 6: Ad5-MU-RGD-Luc infected A549 cells; M: pre-stained protein marker.
CAR and integrin expression in various cell models
To establish the ideal cell model for evaluating the fiber-mutated Ad5 vectors, we investigated the expression of CAR and integrins in a number of human cell lines using indirect flow cytometry analysis with antibodies specific for CAR or the indicated integrins. As shown in Figure 2F, A549 cells expressed the highest level of CAR with a ratio of 88.13%. By contrast, levels of CAR in HL7702, Caco-2 and Skov3 cells were reduced with ratios of 25.66%, 24.48% and 1.09%, respectively (Figure 2B, 2J, 2N). These cells also expressed variable levels of integrins. As shown in Figure 2O and 2L, Skov3 cells expressed higher level of αvβ3 than A549, while Caco-2 cells expressed the highest level of αvβ5 integrin (23.19%). These results indicated that the fiber-mutated Ad5 vectors would have higher transduction efficiency than unmodified Ad5 in Skov3 and Caco-2 cells. Therefore, these two lines may be ideal models for evaluating cancer cell growth inhibition mediated by the proteins expressed from the suicide genes on fiber-mutated Ad5 vectors.
Figure 2.
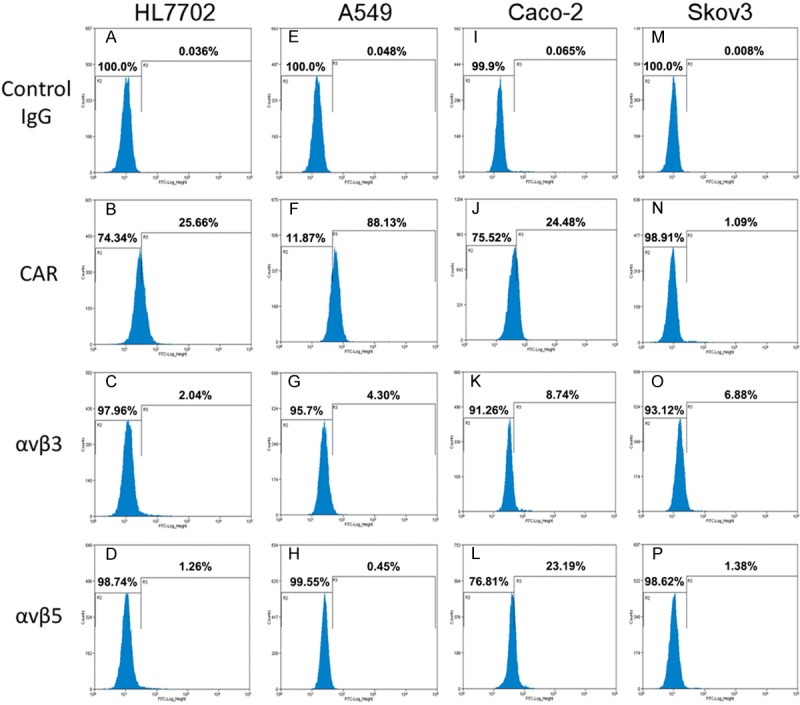
Flow cytometry analysis of levels of CAR and integrins αvβ3 and αvβ5 in experimental cell lines. Cells were labeled with mouse anti-CAR monoclonal antibody, mouse anti-human integrin αvβ3 (LM609) antibody or mouse anti-human integrin αvβ5 (P1F6) antibody. The cells were then incubated with FITC-conjugated rabbit anti-mouse IgG secondary antibody and analyzed by flow cytometry.
Variation in transduction abilities of fiber-mutated Ad5 in vitro
To investigate the infectivity of the fiber-mutated adenoviral vector, cells with different CAR and integrin expression levels, as described above, were infected with Ad5-Blank, Ad5-Luc, Ad5-RGD-Luc or Ad5-MU-RGD-Luc. The log10-transformed values of luciferase activities of these infected cells are presented in Figure 3. The infectivity of Ad5-RGD-Luc was apparently higher than that of Ad5-Luc and Ad5-MU-RGD-Luc in all cell lines at both MOI 1 and 10. Notably, the infectivity of the fiber-mutated Ad5-MU-RGD-Luc was significantly increased in Caco-2 and Skov3 cells, but it was significantly reduced in HL7702 and A549 cells when compared with Ad5-Luc. These findings are consistent with the higher levels of integrins and lower levels of CAR in Caco-2 and Skov3 cells than those of HL7702 and A549 cells shown in Figure 2. These results indicated that the fiber-mutated Ad5-MU-RGD would more effectively transduce some CAR-deficient cancer cell models would be more effective in some CAR-deficient cancer models than the Ad5 vector.
Figure 3.
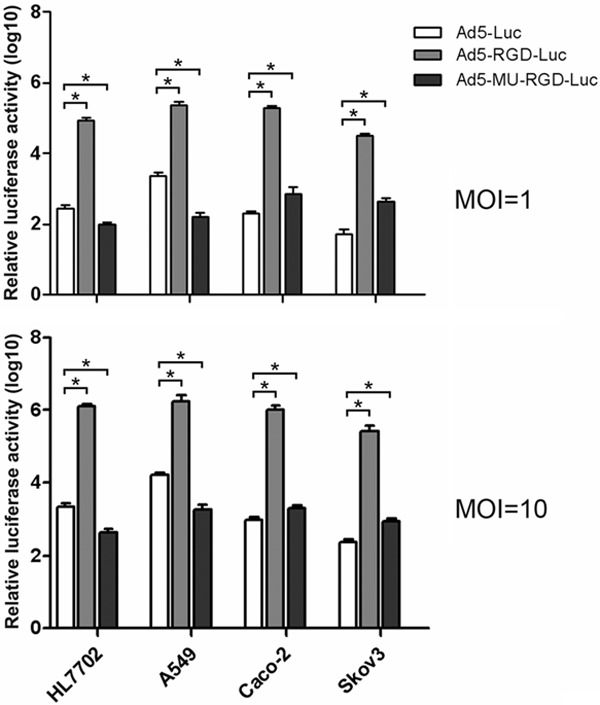
Gene transfer efficiency of fiber-mutated Ad5. Ad vectors were used to infect A549 (high CAR expression), HepG2 (moderate CAR expression), Skov3 (low CAR expression) and HL7702 (normal liver) cells at 10 MOI. The luciferase activity was determined 48 h later. The relative luciferase activity is shown as the fold change in luciferase units relative to the Ad5-blank activity. All data are presented as the mean ± SEM of three experiments. *P < 0.05, compared with Ad5-Luc group.
Mutation in Ad5 fiber decreases liver transduction in mice
To further evaluate whether the fiber-mutated Ad5 vector will decrease liver tropism and transduction, we performed in vivo experiments quantifying gene expression and viral genomes in liver tissues. The mice were injected with Ad5-Luc, Ad5-RGD-Luc or Ad5-MU-RGD-Luc via the tail vein, and luciferase transgene expression was visualized by whole-body bioluminescence imaging 48 h after administration. Results showed that all three Ad5 vectors selectively targeted to the liver as expected, and the fluorescence intensity in liver tissues followed the order of Ad5-RGD-Luc > Ad5-Luc > Ad5-MU-RGD-Luc (Figure 4A). Vector genomes were analyzed by quantitative PCR in liver and other tissues 48 h post-administration of the vectors. Virion levels of Ad5-MU-RGD-Luc were significantly lower than that of Ad5-Luc in the liver (Figure 4B). The results suggest that these mutations in the fiber knob of the Ad5 vector contribute to the attenuation of liver tropism and intrahepatic transduction after intravascular administration.
Figure 4.
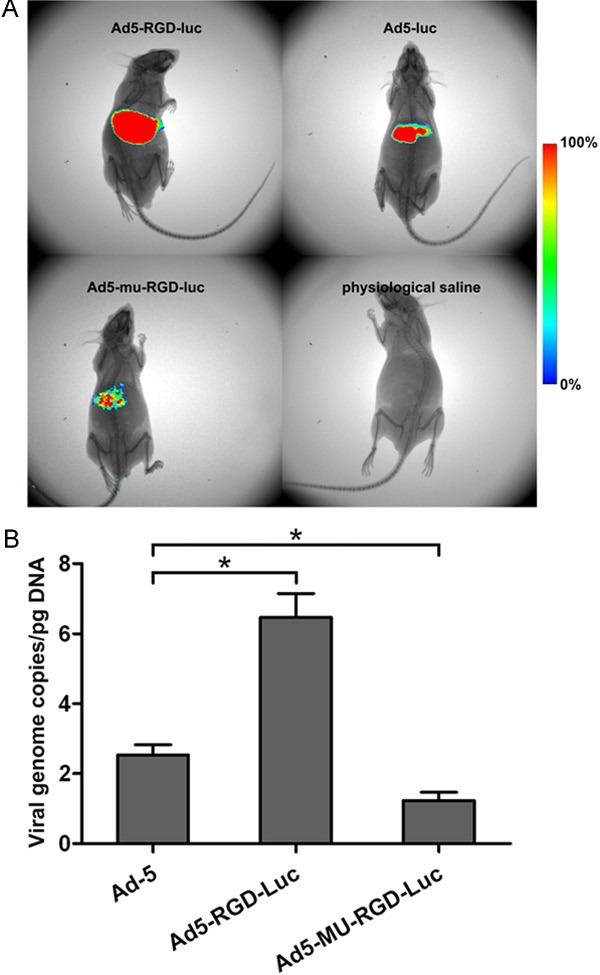
Liver transduction of fiber-mutated Ad5 vectors. A. luciferase expression visualized by whole-body bioluminescence imaging 48 h after systemic administration of Ad5 vectors with 1 × 109 PFU/mouse. B. viral genome content in the liver of mice 48 h after injection with Ad5 vectors. Ad genomes were detected in the liver after DNA extraction and analysis by quantitative PCR (*P < 0.05 vs. Ad5-Luc, n = 5).
Analysis of cytotoxic effects in vitro by fiber-mutated Ad5
In order to investigate whether the fiber-mutated Ad5 vector can be used to more effectively deliver suicide genes to CAR-deficient cancer cells and improve the efficacy of gene therapy, we performed MTT assays on experimental cells infected with either Ad5-hT-TK, Ad5-RGD-hT-TK or Ad5-MU-RGD-hT-TK, followed by treatment with the prodrug GCV. As shown in Figure 5, none of the three Ad5 vectors nor the prodrug showed significant cytotoxicity against the human normal liver HL7702 cells, in which minimal hTERT promoter activity was detected (data not shown). However, in all three cancer cell lines, Ad5-hT-TK/GCV and Ad5-MU-RGD-hT-TK/GCV displayed comparable cytotoxic levels, which were lower than that of Ad5-RGD-hT-TK.
Figure 5.
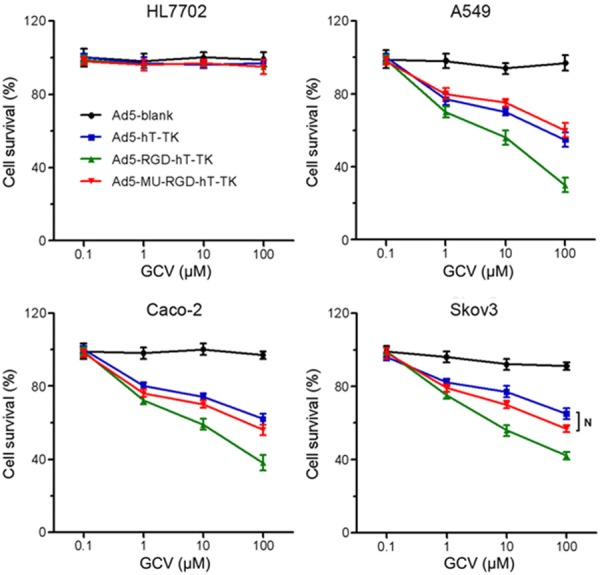
in vitro anti-tumor effects of fiber-mutated Ad5 vectors. Cells were infected with Ad5-Blank, Ad5-hT-TK, Ad5-RGD-hT-TK or Ad5-MU-RGD-hT-TK (MOI = 5) for 3 h and then cultured with medium containing various concentrations of GCV (0.1, 1, 10, 100 μM). Cell survival rates were measured by MTT assays. Each experiment was performed in quintuplicate (N represents P > 0.05).
Discussion
The development of advanced Ad vectors suitable for targeted intravascular gene delivery is required to both improve efficacy and minimize toxicity. However, the use of Ad-based vectors is limited by the inherent hepatic tropism of IV-administered Ad, which precludes targeted delivery to alternative organs or disease sites, and by the associated host inflammatory responses to the vector [12]. Therefore, targeted IV gene delivery with Ad vectors is a long-standing goal in the field of clinical gene therapy. To achieve this aim, platform vectors must combine efficient retargeting strategies with detargeting modifications to ablate native receptor binding (i.e., CAR, integrins) [13].
For efficient targeting of Ad5, the first task is to ablate the virion’s natural tropism, and this ablation has been achieved by introducing mutations into the knob, hexon and penton base within motifs mediating interactions with cell-surface molecules or blood components, leading to reduced transduction efficiency [13-15]. The second challenge is to retarget the Ad5 specifically to the cancer cells and tissues. Since the CAR receptor usually is expressed at low levels in the desired cancer cells, we recently constructed a fiber-modified Ad5 vector containing an RGD peptide in the HI-loop of the fiber knob domain. The incorporation of the RGD peptide has been shown to significantly increase gene transfer to various types of tumors lacking sufficient CAR expression. However, we hypothesized that this vector would have higher levels of accumulation and transduction in liver tissues, which highly express CAR, than unmodified Ad5 vectors following intravascular delivery. Results in the present study confirmed our assumption both in vitro and in vivo (Figures 3, 4).
Because in vitro Ad cell entry was shown to be mediated through interactions of the fiber knob and penton base with CAR and cellular integrins, respectively, roles of both interactions in Ad liver tropism were assessed. Substitutions in fiber knob loops such as the AB loop known to interact with CAR [7,16,17] dramatically reduced the transduction of various CAR-positive cell lines. However, prior studies described conflicting results on whether eliminating CAR binding through discrete knob mutations could alter in vivo distribution following systemic delivery in mice. Leissner P [18] demonstrated that Ad vectors containing the S408E mutation in the fiber AB loop yielded efficient liver transduction in mice, despite having greatly reduced transduction efficiencies on cells in vitro. Similarly, a mutation in the FG loop which ablated CAR-binding did not sufficiently reduce liver transduction [19], and an Ad vector containing a Y477A mutation in the fiber DE loop did not show altered biodistribution or hepatotoxicity [6]. By contrast, Einfeld DA [17] demonstrated that an Ad vector containing a more extensive fiber AB loop mutation (R412S, A415G, E416G, and K417G) had a 10-fold reduction in liver gene expression. In line with that finding and the report by Yun CO [7], our results demonstrated that the combination of such fiber knob AB loop mutations (S408E, P409A) with the RGD motif in the HI loop of the fiber knob resulted in an approximately 5-fold decrease in liver transduction (Figure 4). In addition, our studies showed that both CAR and integrins play important roles in hepatic transduction. However, other mechanisms such as the calcium-dependent interaction between Ad5 and the vitamin K-dependent blood coagulation factor X are also required in governing liver transduction by adenoviral vectors.
As the novel fiber-mutated Ad5 vector could reduce liver tropism and increase gene transfer in CAR-deficient cancer cells Skov3, we expected that it would further enhance anti-tumor effects. However, no significant increase or reduction of in vitro anti-tumor effects was found between Ad5-hT-TK and Ad5-MU-RGD-hT-TK in all three cancer lines expressing different levels of CAR (Figure 5). Therefore, analysis of inhibition of tumor growth by these Ad5 vectors in vivo was not performed. At least two possible explanations account for the above results, one of which could be attributed to the promoter, since we previously showed that the Ad-hT-TK/GCV system was significantly less effective against tumor cells compared with the Ad-CMV-TK/GCV system in vitro [20]. The other explanation is that CAR-deficient cancer cells expressing higher levels of αvβ3 and αvβ5 integrin than that in Skov3 cells may show better inhibition effects. Therefore, changes in the promoter or screening more suitable cell lines to enhance the anti-tumor effects of the fiber-mutated Ad5 vector are likely to yield a safer and more effective gene therapy strategy for the treatment of certain CAR-deficient cancers.
Acknowledgements
This study was supported by the National Nature Science Foundation of China (No. 30872396, 31300756, 81472816) and the Key Projects in the National Science & Technology Pillar Program in the Twelfth Five-year Plan Period (Grant 2012ZX10001-009).
Disclosure of conflict of interest
None.
References
- 1.Kasala D, Choi JW, Kim SW, Yun CO. Utilizing adenovirus vectors for gene delivery in cancer. Expert Opin Drug Deliv. 2014;11:379–392. doi: 10.1517/17425247.2014.874414. [DOI] [PubMed] [Google Scholar]
- 2.Yu B, Zhou Y, Wu H, Wang Z, Zhan Y, Feng X, Geng R, Wu Y, Kong W, Yu X. Seroprevalence of neutralizing antibodies to human adenovirus type 5 in healthy adults in China. J Med Virol. 2012;84:1408–1414. doi: 10.1002/jmv.23325. [DOI] [PubMed] [Google Scholar]
- 3.Imperiale MJ. Keeping adenovirus away from the liver. Cell Host Microbe. 2008;3:119–120. doi: 10.1016/j.chom.2008.02.007. [DOI] [PubMed] [Google Scholar]
- 4.Bergelson JM, Cunningham JA, Droguett G, Kurt-Jones EA, Krithivas A, Hong JS, Horwitz MS, Crowell RL, Finberg RW. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science. 1997;275:1320–1323. doi: 10.1126/science.275.5304.1320. [DOI] [PubMed] [Google Scholar]
- 5.Wickham TJ, Mathias P, Cheresh DA, Nemerow GR. Integrins alpha v beta 3 and alpha v beta 5 promote adenovirus internalization but not virus attachment. Cell. 1993;73:309–319. doi: 10.1016/0092-8674(93)90231-e. [DOI] [PubMed] [Google Scholar]
- 6.Alemany R, Curiel DT. CAR-binding ablation does not change biodistribution and toxicity of adenoviral vectors. Gene Ther. 2001;8:1347–1353. doi: 10.1038/sj.gt.3301515. [DOI] [PubMed] [Google Scholar]
- 7.Yun CO, Yoon AR, Yoo JY, Kim H, Kim M, Ha T, Kim GE, Kim JH. Coxsackie and adenovirus receptor binding ablation reduces adenovirus liver tropism and toxicity. Hum Gene Ther. 2005;16:248–261. doi: 10.1089/hum.2005.16.248. [DOI] [PubMed] [Google Scholar]
- 8.Kanerva A, Mikheeva GV, Krasnykh V, Coolidge CJ, Lam JT, Mahasreshti PJ, Barker SD, Straughn M, Barnes MN, Alvarez RD, Hemminki A, Curiel DT. Targeting adenovirus to the serotype 3 receptor increases gene transfer efficiency to ovarian cancer cells. Clin Cancer Res. 2002;8:275–280. [PubMed] [Google Scholar]
- 9.Dmitriev I, Krasnykh V, Miller CR, Wang M, Kashentseva E, Mikheeva G, Belousova N, Curiel DT. An adenovirus vector with genetically modified fibers demonstrates expanded tropism via utilization of a coxsackievirus and adenovirus receptor-independent cell entry mechanism. J Virol. 1998;72:9706–9713. doi: 10.1128/jvi.72.12.9706-9713.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhan Y, Yu B, Wang Z, Zhang Y, Zhang HH, Wu H, Feng X, Geng RS, Kong W, Yu XH. A fiber-modified adenovirus co-expressing HSV-TK and Coli. NTR enhances antitumor activities in breast cancer cells. Int J Clin Exp Pathol. 2014;7:2850–2860. [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang Y, Yu XH, Zha X, Kong W. Adenovirus-mediated HSV-TK Gene Therapy Using hTERT Promoter in CNE Cells in vitro. Chemical Research in Chinese Universities. 2009;25:60–63. [Google Scholar]
- 12.Parker AL, Nicklin SA, Baker AH. Interactions of adenovirus vectors with blood: implications for intravascular gene therapy applications. Curr Opin Mol Ther. 2008;10:439–448. [PubMed] [Google Scholar]
- 13.Coughlan L, Alba R, Parker AL, Bradshaw AC, McNeish IA, Nicklin SA, Baker AH. Tropism-modification strategies for targeted gene delivery using adenoviral vectors. Viruses. 2010;2:2290–2355. doi: 10.3390/v2102290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Khare R, Chen CY, Weaver EA, Barry MA. Advances and future challenges in adenoviral vector pharmacology and targeting. Curr Gene Ther. 2011;11:241–258. doi: 10.2174/156652311796150363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dreier B, Honegger A, Hess C, Nagy-Davidescu G, Mittl PR, Grutter MG, Belousova N, Mikheeva G, Krasnykh V, Pluckthun A. Development of a generic adenovirus delivery system based on structure-guided design of bispecific trimeric DARPin adapters. Proc Natl Acad Sci U S A. 2013;110:E869–877. doi: 10.1073/pnas.1213653110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kirby I, Davison E, Beavil AJ, Soh CP, Wickham TJ, Roelvink PW, Kovesdi I, Sutton BJ, Santis G. Identification of contact residues and definition of the CAR-binding site of adenovirus type 5 fiber protein. J Virol. 2000;74:2804–2813. doi: 10.1128/jvi.74.6.2804-2813.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Einfeld DA, Schroeder R, Roelvink PW, Lizonova A, King CR, Kovesdi I, Wickham TJ. Reducing the native tropism of adenovirus vectors requires removal of both CAR and integrin interactions. J Virol. 2001;75:11284–11291. doi: 10.1128/JVI.75.23.11284-11291.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Leissner P, Legrand V, Schlesinger Y, Hadji DA, van Raaij M, Cusack S, Pavirani A, Mehtali M. Influence of adenoviral fiber mutations on viral encapsidation, infectivity and in vivo tropism. Gene Ther. 2001;8:49–57. doi: 10.1038/sj.gt.3301343. [DOI] [PubMed] [Google Scholar]
- 19.Mizuguchi H, Koizumi N, Hosono T, Ishii-Watabe A, Uchida E, Utoguchi N, Watanabe Y, Hayakawa T. CAR- or alphav integrin-binding ablated adenovirus vectors, but not fiber-modified vectors containing RGD peptide, do not change the systemic gene transfer properties in mice. Gene Ther. 2002;9:769–776. doi: 10.1038/sj.gt.3301701. [DOI] [PubMed] [Google Scholar]
- 20.Yu B, Zhang Y, Zhan Y, Zha X, Wu Y, Zhang X, Dong Q, Kong W, Yu X. Co-expression of herpes simplex virus thymidine kinase and Escherichia coli nitroreductase by an hTERT-driven adenovirus vector in breast cancer cells results in additive anti-tumor effects. Oncol Rep. 2011;26:255–264. doi: 10.3892/or.2011.1285. [DOI] [PubMed] [Google Scholar]