

Abstract
Gene therapy appears as a promising strategy to treat incurable diseases. In particular, combined gene therapy has shown improved therapeutic efficiency. Internal ribosome entry sites (IRESs), RNA elements naturally present in the 5’ untranslated regions of a few mRNAs, constitute a powerful tool to co-express several genes of interest. IRESs are translational enhancers allowing the translational machinery to start protein synthesis by internal initiation. This feature allowed the design of multi-cistronic vectors expressing several genes from a single mRNA. IRESs exhibit tissue specificity, and drive translation in stress conditions when the global cell translation is blocked, which renders them useful for gene transfer in hypoxic conditions occurring in ischemic diseases and cancer. IRES-based viral and non viral vectors have been used successfully in preclinical and clinical assays of combined gene therapy and resulted in therapeutic benefits for various pathologies including cancers, cardiovascular diseases and degenerative diseases.
Keywords: Vector, Gene transfer, Internal ribosome entry site, Gene therapy
Core tip: Combined gene therapy has emerged for a few years as a promising strategy to improve treatments of many diseases including cancer, cardiovascular diseases and degenerative diseases. In this context, internal ribosome entry site (IRES)-based vectors provide a powerful system to co-express several therapeutic genes from the same transcription unit. IRESs are translational enhancers, exhibiting tissue-specificity, and activated by stress. Different IRES-based vectors including plasmids, adeno-associated virus-derived and lentiviral vectors have been used successfully in many preclinical protocols of gene therapy. Moreover the few clinical assays launched with IRES-based multicistronic vectors resulted in therapeutic benefits.
INTRODUCTION
Combined gene therapy has appeared for a few years as an attractive approach to optimize the therapeutic benefits of gene transfer. In the field of cancer, the first examples of antitumoral cooperative effect have been provided by co-expression of the co-stimulation molecules CD70 and CD80, and of the two anti-angiogenic factors, angiostatin and endostatin, respectively[1-4]. Synergistical effects have also been obtained with co-expression of angiogenic growth factors generating therapeutic angiogenesis in ischemic diseases. This rational has been proven first with co-administration of vascular endothelial growth factor A (VEGFA) and angiopoietin as recombinant proteins as well as by co-administration of two plasmids coding these growth factors[5]. A few years later, combination of recombinant fibroblast growth factor 2 (FGF2) and PDGF-B also improved hindlimb ischemia in rats whereas a bicistronic vector expressing FGF2 and VEGFA efficiently induced vessel formation in a mouse angiogenesis assay[6,7]. These studies launched the concept of combined biotherapy. They also revealed that combined gene therapy is a promising therapeutic approach, allowing long term efficiency of treatments compared to recombinant proteins whose half life is often very short.
Internal ribosome entry sites (IRESs) are translational enhancers naturally present in a series of mRNAs, mediating internal initiation of translation when present between the genes of interest (Figure 1). IRESs thus allow the design of multicistronic expression cassettes resembling bacterial operons, able to drive translation of several genes coded by the same mRNA[8]. We have demonstrated that the use of IRES-based vectors co-expressing two genes of interest allows stable transgene expression with a constant ratio of the proteins of interest, in contrast to the use of two different plasmids expressing each transgene[9]. Actually, a bicistronic IRES-based vector co-expressing FGF2 and Cyr61 has revealed more efficient to generate therapeutic angiogenesis at low doses than the monocistronic vectors expressing large amounts of only one of these angiogenic factors[10]. It must be underlined that the IRES-based vector had no side effects on promotion of tumoral angiogenesis in contrast to the monocistronic ones, a very important feature for increased safety in clinical assays. These observations prompted us to deepen the features of IRESs applicable to vectorology and assess progress made in the field of gene transfer and combined gene therapy clinical assays using IRES-based vectors.
Figure 1.
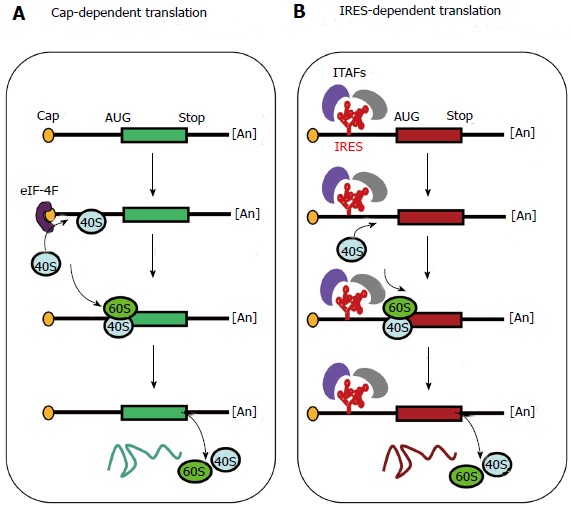
Cap-dependent and internal ribosome entry site-dependent initiation, two alternative mechanisms of translation. A: The so-called cap-dependent ribosome scanning mechanism predicts that ribosome 40S subunit binds to the mRNA 5’ end. Ribosome binding requires the initiation factor 4F (eIF-4F, composed of the three proteins eIF-4E, -4A and -4G). Then the mRNA is unwound under the control of the helicases eIF-4A and -4B, allowing the ribosome to scan the mRNA until recognition of an initiation codon (classically AUG)[11,12]; B: When an Internal ribosome entry site (IRES) is present in the mRNA 5’ untranslated region, IRES trans-acting factors (ITAFs) allow ribosome 40S internal recruitment, independently of the presence of cap and eIF-4F. The IRES-dependent mechanism occurs in the case of picornavirus uncapped mRNAs as well as for cellular capped mRNAs.
IRESS, TRANSLATIONAL ACTIVATORS FOR COMBINED TRANSGENE EXPRESSION
At a time when it was admitted that initiation of translation in eukaryotes required recognition of the capped mRNA 5’ end to recruit ribosomes, translation of the uncapped picornavirus mRNAs from an internal start codon remained a mystery. Indeed, the so-called ribosome scanning mechanism predicted that ribosomes bound to the mRNA 5’ end scanned the mRNA molecules until they recognized an AUG codon[11,12] (Figure 1). The event of internal ribosome binding was thought impossible. This puzzle raised by picornaviruses was solved by the discovery of RNA elements, called IRES, present in the 5’ untranslated regions of their mRNAs, which allow internal recruitment of ribosomes[13,14]. The dogma of the scanning mechanism was thus broken. In addition, it was quickly extended to cellular mRNAs as the first cellular IRES was discovered three years later in the BiP mRNA, coding for the immunoglobulin chaperone also known as GRP78[15]. This discovery was followed by the finding of several other IRESs in cellular mRNAs, in particular in the mRNAs of angiogenic growth factors such as FGF2, proto-oncogenes such as c-myc, pro and anti-apoptotic proteins such as X chromosome-linked inhibitor-of-apoptosis protein and apoptotic peptidase activating factor 1[16-20]. IRESs were also found in retroviruses, whose mRNAs are capped as cellular mRNAs, leading to the design of IRES-containing retroviral vectors[21,22].
The existence of IRESs in capped cellular mRNAs asked the question of their pathophysiological function[23]. Actually, several reports showed that IRESs from cellular mRNAs are regulated in various physiological processes including cell differentiation, spermatogenesis, neurone plasticity[24-27]. Several IRESs are also activated during cell cycle mitosis[28,29]. Recent reports have also shown that IRESs are aberrantly activated in tumor cells, and are thus involved in dysregulation of gene expression in cancer[30]. Furthermore, cellular IRES activity is stimulated in stress conditions such as apoptosis and hypoxia when cap-dependent translation is blocked[31-36].
IRES-dependent internal initiation of translation reminds the prokaryotic initiation mechanism which can translate polycistronic mRNAs[37,38]. This observation gave the idea that such operons could be created in eukaryotes using IRESs to design expression vectors[39]. A large majority of expression vectors allow co-expression of two genes under the control of two promoters. However such an approach has revealed that one of the genes may be silenced despite of the expression of the other one even though it expresses an antibiotic[40]. This can result from competition between the two promoters or counterselection of the gene of interest in case of toxicity or of cell growth inhibition. In such a context, IRESs have been used to generate transgene co-expression under the control of a single promoter (Figure 2).
Figure 2.
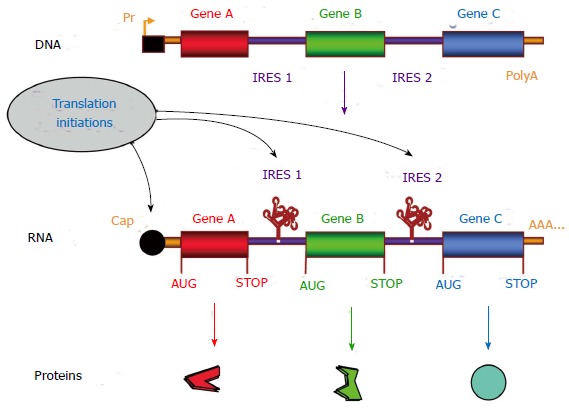
Internal ribosome entry site-based multicistronic vector concept. The internal ribosome entry site (IRES)-based expression cassette contains several genes, separated by IRESs, under the control of the same promoter (Pr). This transcription unit gives rise to a single mRNA coding the different genes. Translation initiation occurs at the 5’ end by the cap-dependent mechanism, resulting in translation of the first open reading frame (ORF, Gene A). Internal initiations of translation occur at each IRES, resulting in translation of the other ORFs (Genes B and C). Thus the multicistronic mRNA generates several proteins from a single transcription unit, allowing more stable long term expression and stable transgene ratio[9,48]. For each ORF, initiation (AUG) and termination (STOP) codons are indicated.
The first retroviral tricistronic IRES-based vector appeared in 1992, providing an exciting potential for gene therapy[41]. This vector successfully co-expressed adenosine desaminase with neomycin (NEO) resistance and chloramphenicol acetyltransferase reporter genes, using the two picornavirus IRESs from poliovirus and encephalomyocarditis virus (EMCV), respectively. Two years later, a therapeutic tricistronic vector expressing the two interleukin-12 subunits with NEO validated the concept of IRES-based vectors to co-express two subunits of a protein with an adequate stoechiometry together with a resistance gene[42]. In the following years, bicistronic vectors were used successfully to select cell clones expressing a protein of interest with a resistance gene, preventing the problems generated by the use of two promoters[40,43].
TISSUE-SPECIFICITY OF CELLULAR IRESS
Most IRES-based vectors developed up to now use picornavirus IRESs, based on the strong efficiency of such IRESs in transient transfection, compared to cellular IRESs. It has been observed that cellular IRESs often exhibit a low efficiency in transiently transfected cells. Such a feature may result from the cell and tissue specificity of the cellular IRES activities. Actually, the FGF2 IRES activity varies with the cell type, the lowest being in fibroblasts, and the highest in neuroblastoma and osteosarcoma cells[44]. Similar variations have been observed for other cellular IRESs[45] (Creancier L and Prats AC, unpublished results). The strongest regulation of cellular IRESs has been shown in vivo, in transgenic mice expressing bicistronic dual luciferase constructs containing different IRESs. Clearly, the EMCV IRES was active in most tissues and organs, while the FGF2 IRES was very low in most organs except for testis and brain where its activity increased 200 to 400 times, at least 10 times higher than the EMCV IRES activity[44]. A similar behavior was observed with other cellular IRESs such as c-myc and VEGFA IRESs[31,45].
The tissue-specific features of cellular IRESs are useful to control transgene expression. Thus they can be considered as translational enhancers, if one makes a parallel with transcriptional enhancers upstream of promoters, governing the tissue-specificity of gene expression. The concept of translational tissue-specificity may be applied to gene transfer by coupling tissue-specific IRESs with tissue-specific promoters to create vectors with increased safety. This concept should also remember us that EMCV is not always the best IRES to be used. A recent study reports the failure of expression of the second cistron of a bicistronic adeno-associated virus (AAV) vector using the EMCV IRES, in murine cerebellar Purkinje neurons[46].
The advantage of using a cellular IRES has also been demonstrated for gene transfer into skeletal muscle. The FGF1 IRES is as efficient as the EMCV IRES in mouse muscle after plasmid DNA electrotransfer[47]. Moreover, when this IRES is used in a bicistronic AAV vector, its activity is significantly superior to that of the EMCV IRES in myoblasts and allows a transgene expression 10 times more efficient when this AAV is injected in mouse muscle[48]. Such a difference may be due to the presence of specific FGF1 IRES trans-acting factors (ITAFs) (Ainaoui et al, in revision). Alternatively, it can result from the lower ability of the EMCV IRES to maintain a stable long term compared to cellular IRESs, shown in a previous report[9].
On the basis of these different data, it can be recommended to choose the adequate IRES to be used according to the cell type or tissue to be targeted, rather than using systematically the EMCV IRES as presently proposed in all commercial IRES-based vectors.
IRES-MEDIATED GENE EXPRESSION IN STRESS CONDITIONS
In many diseases cells are subjected to different stresses such as hypoxia, apoptosis or ER stress. In stress conditions, translation initiation is inhibited by two ways: blockade the mammalian target of rapamycin pathway which affects ribosome recruitment on the cap, and phosphorylation of eIF2-α which prevents charged initiator Met-tRNA formation. Interestingly, IRES-dependent translation is not affected by these two ways of silencing[35,49,50].
As mentioned above, IRESs are naturally present in messenger RNAs coding for proteins involved in the stress response, especially apoptosis and hypoxia. In particular, an IRES is present in the mRNA of the hypoxia-induced factor 1α (HIF1α), the key of the cell response to hypoxia that induces transcription of all the genes containing a hypoxia responsive element (HRE) in their promoters[51]. This IRES allows HIF mRNA translation to be activated during hypoxia despite of the blockade of global translation[32,52]. Such activation occurs under the control of an ITAF, the pyrimidine tract binding protein, also known as a regulator for various IRESs[52,53].
An important consequence of hypoxia is the stimulation of angiogenesis in order to generate new vessels able to restore the cell supply with oxygen. This process occurs in cancers when cells in the tumor core are oxygen deprivated, as well as in ischemic diseases such as heart and lower limb ischemia when tissues are not any more irrigated due to artery occlusion. Strikingly, the major angiogenic factors VEGFA (vascular endothelial growth factor A), FGF1 and FGF2, possess IRESs in their mRNAs[20,47,54-56]. VEGFA expression, transcriptionally induced by HIF1α, is also translationally enhanced via the IRES in hypoxic tumors and in ischemic mouse legs[31,32,36]. In contrast to VEGFA, FGF2 is not induced transcriptionally by hypoxia but its synthesis is translationally induced by the IRES-dependent mechanism in ischemic tissues[31,33]. The same phenomenon has been observed for the major lymphangiogenic factor VEGFC, induced by hypoxia at the translational level via an IRES, but not at the transcriptional level, in tumors and lymph nodes[36,57]. FGF2 and VEGFC induction is exclusively translational and HIF-independent, revealing that IRESs provide an alternative HIF-independent way of response to hypoxia.
On a biotechnological point of view, the sensitivity of IRESs to hypoxia may be an advantage for several applications. Gene transfer vectors can benefit from this feature as the presence of IRESs allows increased transgene expression in ischemic conditions in vivo. Once again, one can see that data from basic research have to be taken into account in the design of optimized expression cassettes. The use of IRES-based vectors seems particularly adequate for gene therapy of ischemic diseases and cancer, as in both cases the transgenes have to be expressed in hypoxic conditions.
BIOMEDICAL APPLICATIONS OF IRESS
IRESs have found biomedical applications for several years. As mentioned above, the first biomedical use of IRESs in an expression vector has been co-expression of subunits of a therapeutic protein with a gene of resistance, as shown for interleukin 12 subunits with a gene of resistance[42]. However this application is limited to therapeutic genes composed of several subunits. In addition, the use of resistance genes is not recommended as it may prevent the use of the vector in a clinical assay.
Another application of IRESs raised during the last decade, resulting form the emerging concept of combined gene therapy. Several studies have validated this concept using a cocktail of two vectors to transfer two genes simultaneously. This has been particularly documented in the field of cardiovascular diseases and cancer, with therapeutic benefits obtained in different animal models using different combinations of angiogenic or anti-angiogenic factors[4,5,58-61] (Table 1). Interestingly the combination of VEGFA and PDGFB successfully induced therapeutic angiogenesis both in ischemic leg and in ischemic heart. In the field of rare diseases, two AAV vectors expressing microdystrophin and IGF1 resulted in increased muscle mass and strength, reduced myofiber degeneration and increased protection against contraction-induced injury in mdx mice[62]. These different studies were performed either with naked DNA or with recombinant adeno-associated virus vectors.
Table 1.
Preclinical studies of combined gene therapy with co-administration of monocistronic vectors
| Pathology | Therapeutic genes | Animal model | Vector type | Ref. |
| Cancers | ||||
| Leukemia, melanoma | Angiostatin + endostatin | Mouse | Retrovirus | Scappaticci et al[4], 2001 |
| Ovarian cancer | Angiostatin + endostatin | Mouse | AAV | Ponnazhagan et al[3], 2004 |
| Glioblastoma | VEGF-R1 + angio-endo (Statin AE) | Mouse | SB transposon | Ohlfest et al[60], 2005 |
| Pancreatic cancer | TSP1 + endostatin | Mouse | AAV | Zhang et al[61], 2007 |
| Cardiovascular diseases | ||||
| Limb ischemia | VEGFA + angiopoietin-1 | Rabbit | Plasmid | Chae et al[5], 2000 |
| Limb ischemia | VEGFA + FGF2 | Mouse | Plasmid | Lee et al[59], 2007 |
| Limb ischemia | VEGFA + PDGFB | Rabbit | AAV | Kupatt et al[58], 2010 |
| Heart ischemia | VEGFA + PDGFB | Pig | AAV | Kupatt et al[58], 2010 |
| Rear diseases | ||||
| DMD | Microdystrophin + IGF1 | Mouse | AAV | Abmayr et al[62], 2005 |
DMD: Duchenne muscular dystrophy; VEGF: Vascular endothelial growth factor; FGF2: Fibroblast growth factor 2; AAV: Adeno-associated virus.
The use of two different vectors for multiple transgene expression exhibits disadvantages: on the one hand, the ratio of the therapeutic molecules cannot be controlled, leading in the loss of the cooperative effect: expression of one of the vectors often decrease or is silenced earlier than the other one[40]. On the other hand, the cost of two therapeutic vectors in a clinical perspective is higher than a single one. These disadvantages are still more important in case of a cocktail of three or more therapeutic genes.
The concept of IRES-based vectors for combined gene therapy has been validated for combined immunotherapy of cancer using a tricistronic retrovirus expressing the two co-stimulation molecules CD70 and CD80[2] (Table 2). In addition to the EMCV IRES, several cellular or retroviral IRESs were successful in this approach[63]. In vivo gene therapy has also been validated for the treatment of ischemic limb in a mouse model, following intramuscular injection and electrotransfer of a plasmid containing the FGF1 IRES for co-expression of FGF2 and Cyr61[10]. This study showed than the two angiogenic factors, although expressed at lower doses from the bicistronic vector than from the monocistronic ones, have a synergistical effect in stimulating therapeutic angiogenesis, rendering the bicistronic construct more efficient. More importantly, due to the lower doses of therapeutic molecules, the bicistronic vector induces no side effects on tumoral angiogenesis, in contrast to one of the monocistronic vectors expressing huge amounts of Cyr61. Thus combined gene therapy using IRES-based vectors is also a safer therapeutic approach.
Table 2.
Preclinical studies of combined gene therapy using multicistronic vectors
| Pathology | Therapeutic genes | Animal model | IRES | Vector type | Ref. |
| Cancers | |||||
| Fibrosarcoma | CD70 + CD80 | Mouse | EMCV | Retrovirus | Couderc et al[2], 1998 |
| Melanoma | Angio-endo fusion | Mouse | None (fusion) | Retrovirus | Scappaticci et al[79], 2001 |
| Multiple myeloma | IL12 subunits + CD80 | Mouse | EMCV + FMDV | Retrovirus | Wen et al[69], 2001; Li et al[67], 2003 |
| Melanoma | CD70 + CD80 | Mouse | EMCV, c-myc, FGF2, HTLV1 | Retrovirus | Douin et al[63], 2004 |
| Ovarian cancer | Angiostatin + endostatin | Mouse | EMCV | AAV | Isayeva et al[66], 2005 |
| Head and neck cancer | Angio-endo fusion | Mouse | None (fusion) | Vaccinia virus | Tysome et al[80], 2011 |
| Pancreas cancer | CXCL4L1 + fibstatin | Mouse | FGF1 | AAV, Lentivector | Prats et al[68], 2013 |
| Cardiovascular diseases | |||||
| Limb ischemia | FGF2 + Cyr 61 | Mouse | FGF1 | Plasmid | Rayssac et al[10], 2009 |
| Limb ischemia | VEGFA + BMP7 | Rabbit | EMCV | AAV | Zhang et al[65], 2010 |
| Limb ischemia | VEGFA + FGF4 | Mouse | EMCV | AAV | Jazwa et al[64], 2013 |
| Heart ischemia | Gata4 + Mef2C + Tbx5 | Rat | None (2A element) | Lentivector | Mathison et al[75], 2014 |
| Neurodegenerative diseases | |||||
| Parkinson | TH + AADC + CH1 | Rat | EMCV | Lentivector | Azzouz et al[73], 2002 |
| Stewart et al[74], 2011 | |||||
| AMD | Angiostatin + endostatin | Mouse | EMCV | Lentivector | Kachi et al[70], 2009 |
BMP7: Bone morphogenetic protein 7; Gata4: GATA binding protein 4; Mef2C: Myocyte-specific enhancer factor 2C; Tbx5: T-box transcription factor 5; TH: Tyrosine hydroxylase; AADC: aromatic L-amino acid decarboxylase; CH1: GTP cyclohydrolase-1; AMD: Age-related macular degeneration.
Additional studies have confirmed the successful use of IRES-based vectors for combined treatment of limb ischemia with VEGFA and FGF4 or bone morphogenetic protein7 (BMP7)[64,65]. Combined gene therapy of cancer was also reported using IRES-based vectors co-expressing IL-12 and CD80, as well as antiangiogenic factors angiostatin and endostatin, or CXCL4I and fibstatin[66-69] (Table 2). Combination of angiostatin and endostatin in an IRES-based vector was also successful to treat age-related macular degeneration in a mouse model[70]. In the field of degenerative diseases, mucopolysaccharidosis type IIIA has been addressed in presymptomatic MPSIIIA mice by intrastriatal administration of an AAV vector co-expressing N-sulfoglycosamine sulfohydrase (SGSH) with the sulfatase-modifying factor (SUMF1) (Winner et al, submitted). This study has resulted in a clinical assay[71] see below). Only one report has obtained better data with two separate AAV vectors to deliver FGF14 and a fluorescent protein into purkinje neurons, than with an IRES[46]. This study used the EMCV IRES previously reported to function in neurons[72]. However it must be underlined that the EMCV IRES is not very active in neurons in vivo, by comparison with the FGF2 IRES that is at least ten times more active[24,44]. In such a case, one can expect that the choice of the FGF2 IRES would provide better data.
Multigene transfer has also been validated for combina-tions of three genes. A tricistronic IRES-based lentivector expressing three catecholaminergic proteins, Prosavin, was administrated by bilateral striatal injection for treatment of Parkinson in rats, resulting in important therapeutic benefits[73,74] (Table 2). Moreover, a tricistronic 2A-based lentivector administrated in situ was also efficient in co-expressing Gata4, Mef2c and Tbx5 for postinfarct ventricular functional improvement in rats[75].
It is often mentioned that the IRES-driven translation of the downstream cistrons is lower than the cap-dependent first cistron translation. This issue can easily be addressed by intelligent vector design: First, one can take into account the tissue specificity of the IRES by choosing the most adequate IRES rather than using systematically the EMCV IRES. Most bi- and- tricistronic vectors use this IRES although it is far to be the best one in many tissues such as muscle or brain[24,48]. Second, the IRES efficiency can be improved. It must be noticed that the EMCV IRES activity is very sensitive to the position of the start codon of the gene of interest. This IRES, in contrast to the FGF1 IRES, exhibits no flexibility: the AUG must be positioned just downstream from the IRES. The insertion of a single restriction site between the IRES and the AUG codon is sufficient to inactivate the IRES[76]. The insertion of a spacer between the first gene and the IRES is also susceptible to enhance the IRES activity by preventing IRES structural alterations by RNA sequences located upstream[77]. In addition, mutations of the upstream AUG codons in the EMCV IRES improve its efficiency[78]. Finally, an important parameter is the IRES regulation by microenvironment. In particular, FGF or VEGF IRES activities are more sensitive to hypoxia than the EMCV IRES and may allow a more efficient transgene expression in ischemic diseases.
ALTERNATIVES TO IRESS FOR MULTICISTRONIC VECTORS
IRES-based vectors are not the only approach to co-express several gene products under the control of a single promoter. The first alternative is gene fusion. It has been successfully used to combine endostatin and angiostatin in a treatment of melanoma and of head and neck cancer[79,80]. A second alternative to IRESs is the use of alternative splicing-based vectors. Such an approach had been proposed many years ago using retroviral vectors, using the natural alternative splicing features of retrovirus genome[81,82]. This concept has been developed more recently in the purpose of co-expressing two immunoglobulin chains[83]. The interest of this system is the ability to adapt the ratio of the two transgenes by mutating the splicing sites. However one limit of this attractive system is that splicing site efficiency and consequently the ratio of the two proteins of interest, is influenced by the presence of exon splicing enhancers or silencers in the transgene sequences, preventing the design of vectors with a stable transgene ratio applicable to co-expression of any pair of therapeutic proteins.
A third exciting system of co-expression is provided by the 2A peptides. Such peptides, occurring in many viral genomes, are peptide sequences of about 19 amino-acid residues, which can produce a discontinuity in the translated polypeptide when encoded in a longer open reading frame (ORF)[84]. In contrast to what is currently admitted, 2A peptides do not catalyze a protein cleavage, but they catalyze termination of translation in the absence of a stop codon, followed by reinitiation. They are currently used as a tool to co-express two or more separate proteins from a single ORF[85]. 2A peptides thus constitute an alternative to IRESs, but do not work in all systems. By example, in the study in purkinje neurons mentioned above, a 2A peptide was used but did not function, resulting in detection of the longer ORF rather than the two expected proteins[46]. In another report comparing bicistronic constructs expressing Sox9 and EGFP separated by the EMCV IRES or by the FMDV 2A peptide, the authors detected 42% of Sox-EGFP fusion protein, reflecting an inefficient ribosome skipping mechanism[86]. Formation of such fusion proteins often occurs with proteins bearing N-terminal signal sequences[87]. In addition, no information is available about the 2A peptides tissue-specificity or behavior in response to stress, in contrast to IRESs.
CLINICAL APPLICATIONS OF IRES-BASED VECTORS TO GENE THERAPY
All the preclinical studies mentioned above show that IRES-based vectors represent an exciting tool to be used for combined gene therapy. Nowadays, very little clinical trials with such vectors have been reported. The first trial to be cited is the tricistronic IL12-expressing retrovirus, which gave significant decrease of tumor sizes on a few patients with melanoma or head and neck cancer[88,89].
A bicistronic IRES-based vector co-expressing FGF2 and VEGFA has been assessed in a clinical assay of gene therapy on patients with refractory coronary disease[90] (Table 3). The protocol corresponded to intramyocardial transfer of a plasmid expressing the bicistronic cassette. This study showed no improvement in myocardial perfusion, but treated patients exhibited improved exercice tolerance and clinical symptoms. Furthermore the bicistronic gene transfer was safe. This moderate benefit, although encouraging, may be due to the use of a plasmid, which does not provide long term expression in contrast to viral vectors, and also to the choice of the EMCV IRES which is not optimal to drive gene expression in hypoxic conditions[31,36].
Table 3.
Clinical studies of combined gene therapy
| Pathology | Therapeutic genes | IRES | Vector type | Outcome | Ref. |
| Ischemic heart disease | VEGFA + FGF2 | EMCV | Plasmid | Moderate benefits | Kukula et al[90], 2011 |
| Parkinson | TH + AADC + CH1 | EMCV | Lentivector | Benefits for 15/15 patients | Palfi et al[91], 2014 |
| Mucopolysaccharidosis type IIIA | SGSH + SUMF1 | EMCV | Lentivector | Benefits for 1/4 patients, stabilization for 3/4 | Tardieu et al[71], 2014 |
TH: Tyrosine hydroxylase; AADC: Aromatic L-amino acid decarboxylase; CH1: GTP cyclohydrolase-1; SGSH: N-sulfoglycosamine sulfohydrolase; SUMF1: Sulfatase-modifying factor.
Very recently, two gene therapy clinical trials successfully used multi-cistronic IRES-based viral vectors. On the one hand, a gene therapyI/II phase clinical trial on patients with mucopolysaccharidosis type IIIA, a severe degenerative disease, has displayed neurocognitive benefits[71]. Four children received intracerebral injections of a bicistronic AAV vector expressing the SGSH and SUMF1 genes separated by the EMCV IRES. Neurocognitive evaluations suggest a cognitive benefit on the youngest patient, where as the other ones are stabilized. Importantly, the treatment was safe and well tolerated after 1 year in all the patients, validating the surgical approach for direct AAV delivery in the brain parenchyma. On the other hand, a phaseI/II assay was performed on 15 patients with Parkinson’s disease using Prosavin (see above), a tricistronic lentivector with EMCV IRESs administrated by intrastriatal delivery[91]. A significant improvement of motor scores was recorded in all patients at 6 mo. This is the first-in-man use of a lentiviral-based gene therapy vector for a neurodegenerative disease. These studies validate the clinical use of IRES-based viral vectors.
CONCLUSION
Many reports have shown that combined gene therapy is an attractive approach in animal models. This observation has justified extensive research on optimization of gene transfer vectors able to co-express several proteins. In this context, IRES-based vectors have now been validated in pre-clinical as well as in clinical studies by showing their safety and ability to generate therapeutic benefits.
In addition, the data available on IRES tissue-specificity and activation in response to stress provide promising perspectives of vector improvement, which may result in better efficiency of gene therapy.
Footnotes
Conflict-of-interest: The authors declare they have no conflicting interests (including but not limited to commercial, personal, political, intellectual, or religious interests) related to the present work.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: October 18, 2014
First decision: November 20, 2014
Article in press: December 19, 2014
P- Reviewer: Midoux P, Samulski RJ S- Editor: Ji FF L- Editor: A E- Editor: Wu HL
References
- 1.Campochiaro PA. Gene transfer for ocular neovascularization and macular edema. Gene Ther. 2012;19:121–126. doi: 10.1038/gt.2011.164. [DOI] [PubMed] [Google Scholar]
- 2.Couderc B, Zitvogel L, Douin-Echinard V, Djennane L, Tahara H, Favre G, Lotze MT, Robbins PD. Enhancement of antitumor immunity by expression of CD70 (CD27 ligand) or CD154 (CD40 ligand) costimulatory molecules in tumor cells. Cancer Gene Ther. 1998;5:163–175. [PubMed] [Google Scholar]
- 3.Ponnazhagan S, Mahendra G, Kumar S, Shaw DR, Stockard CR, Grizzle WE, Meleth S. Adeno-associated virus 2-mediated antiangiogenic cancer gene therapy: long-term efficacy of a vector encoding angiostatin and endostatin over vectors encoding a single factor. Cancer Res. 2004;64:1781–1787. doi: 10.1158/0008-5472.can-03-1786. [DOI] [PubMed] [Google Scholar]
- 4.Scappaticci FA, Smith R, Pathak A, Schloss D, Lum B, Cao Y, Johnson F, Engleman EG, Nolan GP. Combination angiostatin and endostatin gene transfer induces synergistic antiangiogenic activity in vitro and antitumor efficacy in leukemia and solid tumors in mice. Mol Ther. 2001;3:186–196. doi: 10.1006/mthe.2000.0243. [DOI] [PubMed] [Google Scholar]
- 5.Chae JK, Kim I, Lim ST, Chung MJ, Kim WH, Kim HG, Ko JK, Koh GY. Coadministration of angiopoietin-1 and vascular endothelial growth factor enhances collateral vascularization. Arterioscler Thromb Vasc Biol. 2000;20:2573–2578. doi: 10.1161/01.atv.20.12.2573. [DOI] [PubMed] [Google Scholar]
- 6.Cao R, Bråkenhielm E, Pawliuk R, Wariaro D, Post MJ, Wahlberg E, Leboulch P, Cao Y. Angiogenic synergism, vascular stability and improvement of hind-limb ischemia by a combination of PDGF-BB and FGF-2. Nat Med. 2003;9:604–613. doi: 10.1038/nm848. [DOI] [PubMed] [Google Scholar]
- 7.Małecki J, Wesche J, Skjerpen CS, Wiedłocha A, Olsnes S. Translocation of FGF-1 and FGF-2 across vesicular membranes occurs during G1-phase by a common mechanism. Mol Biol Cell. 2004;15:801–814. doi: 10.1091/mbc.E03-08-0589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fussenegger M, Moser S, Bailey JE. pQuattro vectors allow one-step multigene metabolic engineering and auto-selection of quattrocistronic artificial mammalian operons. Cytotechnology. 1998;28:229–235. doi: 10.1023/A:1008014706196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Allera-Moreau C, Delluc-Clavières A, Castano C, Van den Berghe L, Golzio M, Moreau M, Teissié J, Arnal JF, Prats AC. Long term expression of bicistronic vector driven by the FGF-1 IRES in mouse muscle. BMC Biotechnol. 2007;7:74. doi: 10.1186/1472-6750-7-74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rayssac A, Neveu C, Pucelle M, Van den Berghe L, Prado-Lourenco L, Arnal JF, Chaufour X, Prats AC. IRES-based vector coexpressing FGF2 and Cyr61 provides synergistic and safe therapeutics of lower limb ischemia. Mol Ther. 2009;17:2010–2019. doi: 10.1038/mt.2009.211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kozak M. Migration of 40 S ribosomal subunits on messenger RNA when initiation is perturbed by lowering magnesium or adding drugs. J Biol Chem. 1979;254:4731–4738. [PubMed] [Google Scholar]
- 12.Kozak M. The scanning model for translation: an update. J Cell Biol. 1989;108:229–241. doi: 10.1083/jcb.108.2.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jang SK, Kräusslich HG, Nicklin MJ, Duke GM, Palmenberg AC, Wimmer E. A segment of the 5’ nontranslated region of encephalomyocarditis virus RNA directs internal entry of ribosomes during in vitro translation. J Virol. 1988;62:2636–2643. doi: 10.1128/jvi.62.8.2636-2643.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Pelletier J, Sonenberg N. Internal initiation of translation of eukaryotic mRNA directed by a sequence derived from poliovirus RNA. Nature. 1988;334:320–325. doi: 10.1038/334320a0. [DOI] [PubMed] [Google Scholar]
- 15.Macejak DG, Sarnow P. Internal initiation of translation mediated by the 5’ leader of a cellular mRNA. Nature. 1991;353:90–94. doi: 10.1038/353090a0. [DOI] [PubMed] [Google Scholar]
- 16.Coldwell MJ, Mitchell SA, Stoneley M, MacFarlane M, Willis AE. Initiation of Apaf-1 translation by internal ribosome entry. Oncogene. 2000;19:899–905. doi: 10.1038/sj.onc.1203407. [DOI] [PubMed] [Google Scholar]
- 17.Holcik M, Lefebvre C, Yeh C, Chow T, Korneluk RG. A new internal-ribosome-entry-site motif potentiates XIAP-mediated cytoprotection. Nat Cell Biol. 1999;1:190–192. doi: 10.1038/11109. [DOI] [PubMed] [Google Scholar]
- 18.Nanbru C, Lafon I, Audigier S, Gensac MC, Vagner S, Huez G, Prats AC. Alternative translation of the proto-oncogene c-myc by an internal ribosome entry site. J Biol Chem. 1997;272:32061–32066. doi: 10.1074/jbc.272.51.32061. [DOI] [PubMed] [Google Scholar]
- 19.Stoneley M, Paulin FE, Le Quesne JP, Chappell SA, Willis AE. C-Myc 5’ untranslated region contains an internal ribosome entry segment. Oncogene. 1998;16:423–428. doi: 10.1038/sj.onc.1201763. [DOI] [PubMed] [Google Scholar]
- 20.Vagner S, Gensac MC, Maret A, Bayard F, Amalric F, Prats H, Prats AC. Alternative translation of human fibroblast growth factor 2 mRNA occurs by internal entry of ribosomes. Mol Cell Biol. 1995;15:35–44. doi: 10.1128/mcb.15.1.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Berlioz C, Darlix JL. An internal ribosomal entry mechanism promotes translation of murine leukemia virus gag polyprotein precursors. J Virol. 1995;69:2214–2222. doi: 10.1128/jvi.69.4.2214-2222.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Vagner S, Waysbort A, Marenda M, Gensac MC, Amalric F, Prats AC. Alternative translation initiation of the Moloney murine leukemia virus mRNA controlled by internal ribosome entry involving the p57/PTB splicing factor. J Biol Chem. 1995;270:20376–20383. doi: 10.1074/jbc.270.35.20376. [DOI] [PubMed] [Google Scholar]
- 23.Jackson RJ. mRNA translation. Initiation without an end. Nature. 1991;353:14–15. doi: 10.1038/353014a0. [DOI] [PubMed] [Google Scholar]
- 24.Audigier S, Guiramand J, Prado-Lourenco L, Conte C, Gonzalez-Herrera IG, Cohen-Solal C, Récasens M, Prats AC. Potent activation of FGF-2 IRES-dependent mechanism of translation during brain development. RNA. 2008;14:1852–1864. doi: 10.1261/rna.790608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bernstein J, Sella O, Le SY, Elroy-Stein O. PDGF2/c-sis mRNA leader contains a differentiation-linked internal ribosomal entry site (D-IRES) J Biol Chem. 1997;272:9356–9362. doi: 10.1074/jbc.272.14.9356. [DOI] [PubMed] [Google Scholar]
- 26.Conte C, Ainaoui N, Delluc-Clavières A, Khoury MP, Azar R, Pujol F, Martineau Y, Pyronnet S, Prats AC. Fibroblast growth factor 1 induced during myogenesis by a transcription-translation coupling mechanism. Nucleic Acids Res. 2009;37:5267–5278. doi: 10.1093/nar/gkp550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gonzalez-Herrera IG, Prado-Lourenco L, Pileur F, Conte C, Morin A, Cabon F, Prats H, Vagner S, Bayard F, Audigier S, et al. Testosterone regulates FGF-2 expression during testis maturation by an IRES-dependent translational mechanism. FASEB J. 2006;20:476–478. doi: 10.1096/fj.04-3314fje. [DOI] [PubMed] [Google Scholar]
- 28.Cornelis S, Bruynooghe Y, Denecker G, Van Huffel S, Tinton S, Beyaert R. Identification and characterization of a novel cell cycle-regulated internal ribosome entry site. Mol Cell. 2000;5:597–605. doi: 10.1016/s1097-2765(00)80239-7. [DOI] [PubMed] [Google Scholar]
- 29.Pyronnet S, Pradayrol L, Sonenberg N. A cell cycle-dependent internal ribosome entry site. Mol Cell. 2000;5:607–616. doi: 10.1016/s1097-2765(00)80240-3. [DOI] [PubMed] [Google Scholar]
- 30.Marcel V, Ghayad SE, Belin S, Therizols G, Morel AP, Solano-Gonzàlez E, Vendrell JA, Hacot S, Mertani HC, Albaret MA, et al. p53 acts as a safeguard of translational control by regulating fibrillarin and rRNA methylation in cancer. Cancer Cell. 2013;24:318–330. doi: 10.1016/j.ccr.2013.08.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bornes S, Prado-Lourenco L, Bastide A, Zanibellato C, Iacovoni JS, Lacazette E, Prats AC, Touriol C, Prats H. Translational induction of VEGF internal ribosome entry site elements during the early response to ischemic stress. Circ Res. 2007;100:305–308. doi: 10.1161/01.RES.0000258873.08041.c9. [DOI] [PubMed] [Google Scholar]
- 32.Braunstein S, Karpisheva K, Pola C, Goldberg J, Hochman T, Yee H, Cangiarella J, Arju R, Formenti SC, Schneider RJ. A hypoxia-controlled cap-dependent to cap-independent translation switch in breast cancer. Mol Cell. 2007;28:501–512. doi: 10.1016/j.molcel.2007.10.019. [DOI] [PubMed] [Google Scholar]
- 33.Conte C, Riant E, Toutain C, Pujol F, Arnal JF, Lenfant F, Prats AC. FGF2 translationally induced by hypoxia is involved in negative and positive feedback loops with HIF-1alpha. PLoS One. 2008;3:e3078. doi: 10.1371/journal.pone.0003078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Damiano F, Alemanno S, Gnoni GV, Siculella L. Translational control of the sterol-regulatory transcription factor SREBP-1 mRNA in response to serum starvation or ER stress is mediated by an internal ribosome entry site. Biochem J. 2010;429:603–612. doi: 10.1042/BJ20091827. [DOI] [PubMed] [Google Scholar]
- 35.Holcik M, Sonenberg N. Translational control in stress and apoptosis. Nat Rev Mol Cell Biol. 2005;6:318–327. doi: 10.1038/nrm1618. [DOI] [PubMed] [Google Scholar]
- 36.Morfoisse F, Kuchnio A, Frainay C, Gomez-Brouchet A, Delisle MB, Marzi S, Helfer AC, Hantelys F, Pujol F, Guillermet-Guibert J, et al. Hypoxia induces VEGF-C expression in metastatic tumor cells via a HIF-1α-independent translation-mediated mechanism. Cell Rep. 2014;6:155–167. doi: 10.1016/j.celrep.2013.12.011. [DOI] [PubMed] [Google Scholar]
- 37.Pestova TV, Shatsky IN, Fletcher SP, Jackson RJ, Hellen CU. A prokaryotic-like mode of cytoplasmic eukaryotic ribosome binding to the initiation codon during internal translation initiation of hepatitis C and classical swine fever virus RNAs. Genes Dev. 1998;12:67–83. doi: 10.1101/gad.12.1.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Pilipenko EV, Gmyl AP, Maslova SV, Svitkin YV, Sinyakov AN, Agol VI. Prokaryotic-like cis elements in the cap-independent internal initiation of translation on picornavirus RNA. Cell. 1992;68:119–131. doi: 10.1016/0092-8674(92)90211-t. [DOI] [PubMed] [Google Scholar]
- 39.Moss B, Elroy-Stein O, Mizukami T, Alexander WA, Fuerst TR. Product review. New mammalian expression vectors. Nature. 1990;348:91–92. doi: 10.1038/348091a0. [DOI] [PubMed] [Google Scholar]
- 40.Allera-Moreau C, Chomarat P, Audinot V, Cogé F, Gillard M, Martineau Y, Boutin JA, Prats AC. The use of IRES-based bicistronic vectors allows the stable expression of recombinant G-protein coupled receptors such as NPY5 and histamine 4. Biochimie. 2006;88:737–746. doi: 10.1016/j.biochi.2006.05.019. [DOI] [PubMed] [Google Scholar]
- 41.Morgan RA, Couture L, Elroy-Stein O, Ragheb J, Moss B, Anderson WF. Retroviral vectors containing putative internal ribosome entry sites: development of a polycistronic gene transfer system and applications to human gene therapy. Nucleic Acids Res. 1992;20:1293–1299. doi: 10.1093/nar/20.6.1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zitvogel L, Tahara H, Cai Q, Storkus WJ, Muller G, Wolf SF, Gately M, Robbins PD, Lotze MT. Construction and characterization of retroviral vectors expressing biologically active human interleukin-12. Hum Gene Ther. 1994;5:1493–1506. doi: 10.1089/hum.1994.5.12-1493. [DOI] [PubMed] [Google Scholar]
- 43.Arnaud E, Touriol C, Boutonnet C, Gensac MC, Vagner S, Prats H, Prats AC. A new 34-kilodalton isoform of human fibroblast growth factor 2 is cap dependently synthesized by using a non-AUG start codon and behaves as a survival factor. Mol Cell Biol. 1999;19:505–514. doi: 10.1128/mcb.19.1.505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Créancier L, Morello D, Mercier P, Prats AC. Fibroblast growth factor 2 internal ribosome entry site (IRES) activity ex vivo and in transgenic mice reveals a stringent tissue-specific regulation. J Cell Biol. 2000;150:275–281. doi: 10.1083/jcb.150.1.275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Créancier L, Mercier P, Prats AC, Morello D. c-myc Internal ribosome entry site activity is developmentally controlled and subjected to a strong translational repression in adult transgenic mice. Mol Cell Biol. 2001;21:1833–1840. doi: 10.1128/MCB.21.5.1833-1840.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Bosch MK, Nerbonne JM, Ornitz DM. Dual transgene expression in murine cerebellar Purkinje neurons by viral transduction in vivo. PLoS One. 2014;9:e104062. doi: 10.1371/journal.pone.0104062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Martineau Y, Le Bec C, Monbrun L, Allo V, Chiu IM, Danos O, Moine H, Prats H, Prats AC. Internal ribosome entry site structural motifs conserved among mammalian fibroblast growth factor 1 alternatively spliced mRNAs. Mol Cell Biol. 2004;24:7622–7635. doi: 10.1128/MCB.24.17.7622-7635.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Delluc-Clavières A, Le Bec C, Van den Berghe L, Conte C, Allo V, Danos O, Prats AC. Efficient gene transfer in skeletal muscle with AAV-derived bicistronic vector using the FGF-1 IRES. Gene Ther. 2008;15:1090–1098. doi: 10.1038/gt.2008.49. [DOI] [PubMed] [Google Scholar]
- 49.Spriggs KA, Bushell M, Mitchell SA, Willis AE. Internal ribosome entry segment-mediated translation during apoptosis: the role of IRES-trans-acting factors. Cell Death Differ. 2005;12:585–591. doi: 10.1038/sj.cdd.4401642. [DOI] [PubMed] [Google Scholar]
- 50.Thakor N, Holcik M. IRES-mediated translation of cellular messenger RNA operates in eIF2α- independent manner during stress. Nucleic Acids Res. 2012;40:541–552. doi: 10.1093/nar/gkr701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Lang KJ, Kappel A, Goodall GJ. Hypoxia-inducible factor-1alpha mRNA contains an internal ribosome entry site that allows efficient translation during normoxia and hypoxia. Mol Biol Cell. 2002;13:1792–1801. doi: 10.1091/mbc.02-02-0017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Schepens B, Tinton SA, Bruynooghe Y, Beyaert R, Cornelis S. The polypyrimidine tract-binding protein stimulates HIF-1alpha IRES-mediated translation during hypoxia. Nucleic Acids Res. 2005;33:6884–6894. doi: 10.1093/nar/gki1000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Romanelli MG, Diani E, Lievens PM. New insights into functional roles of the polypyrimidine tract-binding protein. Int J Mol Sci. 2013;14:22906–22932. doi: 10.3390/ijms141122906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Akiri G, Nahari D, Finkelstein Y, Le SY, Elroy-Stein O, Levi BZ. Regulation of vascular endothelial growth factor (VEGF) expression is mediated by internal initiation of translation and alternative initiation of transcription. Oncogene. 1998;17:227–236. doi: 10.1038/sj.onc.1202019. [DOI] [PubMed] [Google Scholar]
- 55.Huez I, Créancier L, Audigier S, Gensac MC, Prats AC, Prats H. Two independent internal ribosome entry sites are involved in translation initiation of vascular endothelial growth factor mRNA. Mol Cell Biol. 1998;18:6178–6190. doi: 10.1128/mcb.18.11.6178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Stein I, Itin A, Einat P, Skaliter R, Grossman Z, Keshet E. Translation of vascular endothelial growth factor mRNA by internal ribosome entry: implications for translation under hypoxia. Mol Cell Biol. 1998;18:3112–3119. doi: 10.1128/mcb.18.6.3112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Morfoisse F, Renaud E, Hantelys F, Prats AC, Garmy-Susini B. Lymphangiogenic gene expression adaptation in tumor hypoxic environment. Med Sci (Paris) 2014;30:506–508. doi: 10.1051/medsci/20143005010. [DOI] [PubMed] [Google Scholar]
- 58.Kupatt C, Hinkel R, Pfosser A, El-Aouni C, Wuchrer A, Fritz A, Globisch F, Thormann M, Horstkotte J, Lebherz C, et al. Cotransfection of vascular endothelial growth factor-A and platelet-derived growth factor-B via recombinant adeno-associated virus resolves chronic ischemic malperfusion role of vessel maturation. J Am Coll Cardiol. 2010;56:414–422. doi: 10.1016/j.jacc.2010.03.050. [DOI] [PubMed] [Google Scholar]
- 59.Lee JS, Kim JM, Kim KL, Jang HS, Shin IS, Jeon ES, Suh W, Byun J, Kim DK. Combined administration of naked DNA vectors encoding VEGF and bFGF enhances tissue perfusion and arteriogenesis in ischemic hindlimb. Biochem Biophys Res Commun. 2007;360:752–758. doi: 10.1016/j.bbrc.2007.06.120. [DOI] [PubMed] [Google Scholar]
- 60.Ohlfest JR, Demorest ZL, Motooka Y, Vengco I, Oh S, Chen E, Scappaticci FA, Saplis RJ, Ekker SC, Low WC, et al. Combinatorial antiangiogenic gene therapy by nonviral gene transfer using the sleeping beauty transposon causes tumor regression and improves survival in mice bearing intracranial human glioblastoma. Mol Ther. 2005;12:778–788. doi: 10.1016/j.ymthe.2005.07.689. [DOI] [PubMed] [Google Scholar]
- 61.Zhang X, Xu J, Lawler J, Terwilliger E, Parangi S. Adeno-associated virus-mediated antiangiogenic gene therapy with thrombospondin-1 type 1 repeats and endostatin. Clin Cancer Res. 2007;13:3968–3976. doi: 10.1158/1078-0432.CCR-07-0245. [DOI] [PubMed] [Google Scholar]
- 62.Abmayr S, Gregorevic P, Allen JM, Chamberlain JS. Phenotypic improvement of dystrophic muscles by rAAV/microdystrophin vectors is augmented by Igf1 codelivery. Mol Ther. 2005;12:441–450. doi: 10.1016/j.ymthe.2005.04.001. [DOI] [PubMed] [Google Scholar]
- 63.Douin V, Bornes S, Creancier L, Rochaix P, Favre G, Prats AC, Couderc B. Use and comparison of different internal ribosomal entry sites (IRES) in tricistronic retroviral vectors. BMC Biotechnol. 2004;4:16. doi: 10.1186/1472-6750-4-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Jazwa A, Tomczyk M, Taha HM, Hytonen E, Stoszko M, Zentilin L, Giacca M, Yla-Herttuala S, Emanueli C, Jozkowicz A, et al. Arteriogenic therapy based on simultaneous delivery of VEGF-A and FGF4 genes improves the recovery from acute limb ischemia. Vasc Cell. 2013;5:13. doi: 10.1186/2045-824X-5-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zhang C, Wang KZ, Qiang H, Tang YL, Li Q, Li M, Dang XQ. Angiopoiesis and bone regeneration via co-expression of the hVEGF and hBMP genes from an adeno-associated viral vector in vitro and in vivo. Acta Pharmacol Sin. 2010;31:821–830. doi: 10.1038/aps.2010.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Isayeva T, Ren C, Ponnazhagan S. Recombinant adeno-associated virus 2-mediated antiangiogenic prevention in a mouse model of intraperitoneal ovarian cancer. Clin Cancer Res. 2005;11:1342–1347. [PubMed] [Google Scholar]
- 67.Li ZH, Wen XY, Mandelbaum S, Falcioni N, Hawley TS, Hawley RG, Stewart AK. Improved therapeutic outcome following combination immunogene vaccination therapy in murine myeloma. Leuk Lymphoma. 2003;44:1775–1784. doi: 10.1080/1042819031000119208. [DOI] [PubMed] [Google Scholar]
- 68.Prats AC, Van den Berghe L, Rayssac A, Ainaoui N, Morfoisse F, Pujol F, Legonidec S, Bikfalvi A, Prats H, Pyronnet S, et al. CXCL4L1-fibstatin cooperation inhibits tumor angiogenesis, lymphangiogenesis and metastasis. Microvasc Res. 2013;89:25–33. doi: 10.1016/j.mvr.2013.05.005. [DOI] [PubMed] [Google Scholar]
- 69.Wen XY, Mandelbaum S, Li ZH, Hitt M, Graham FL, Hawley TS, Hawley RG, Stewart AK. Tricistronic viral vectors co-expressing interleukin-12 (1L-12) and CD80 (B7-1) for the immunotherapy of cancer: preclinical studies in myeloma. Cancer Gene Ther. 2001;8:361–370. doi: 10.1038/sj.cgt.7700321. [DOI] [PubMed] [Google Scholar]
- 70.Kachi S, Binley K, Yokoi K, Umeda N, Akiyama H, Muramatu D, Iqball S, Kan O, Naylor S, Campochiaro PA. Equine infectious anemia viral vector-mediated codelivery of endostatin and angiostatin driven by retinal pigmented epithelium-specific VMD2 promoter inhibits choroidal neovascularization. Hum Gene Ther. 2009;20:31–39. doi: 10.1089/hum.2008.046. [DOI] [PubMed] [Google Scholar]
- 71.Tardieu M, Zérah M, Husson B, de Bournonville S, Deiva K, Adamsbaum C, Vincent F, Hocquemiller M, Broissand C, Furlan V, et al. Intracerebral administration of adeno-associated viral vector serotype rh.10 carrying human SGSH and SUMF1 cDNAs in children with mucopolysaccharidosis type IIIA disease: results of a phase I/II trial. Hum Gene Ther. 2014;25:506–516. doi: 10.1089/hum.2013.238. [DOI] [PubMed] [Google Scholar]
- 72.Derrington EA, López-Lastra M, Darlix JL. Dicistronic MLV-retroviral vectors transduce neural precursors in vivo and co-express two genes in their differentiated neuronal progeny. Retrovirology. 2005;2:60. doi: 10.1186/1742-4690-2-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Azzouz M, Martin-Rendon E, Barber RD, Mitrophanous KA, Carter EE, Rohll JB, Kingsman SM, Kingsman AJ, Mazarakis ND. Multicistronic lentiviral vector-mediated striatal gene transfer of aromatic L-amino acid decarboxylase, tyrosine hydroxylase, and GTP cyclohydrolase I induces sustained transgene expression, dopamine production, and functional improvement in a rat model of Parkinson’s disease. J Neurosci. 2002;22:10302–10312. doi: 10.1523/JNEUROSCI.22-23-10302.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Stewart HJ, Fong-Wong L, Strickland I, Chipchase D, Kelleher M, Stevenson L, Thoree V, McCarthy J, Ralph GS, Mitrophanous KA, et al. A stable producer cell line for the manufacture of a lentiviral vector for gene therapy of Parkinson’s disease. Hum Gene Ther. 2011;22:357–369. doi: 10.1089/hum.2010.142. [DOI] [PubMed] [Google Scholar]
- 75.Mathison M, Singh VP, Gersch RP, Ramirez MO, Cooney A, Kaminsky SM, Chiuchiolo MJ, Nasser A, Yang J, Crystal RG, et al. “Triplet” polycistronic vectors encoding Gata4, Mef2c, and Tbx5 enhances postinfarct ventricular functional improvement compared with singlet vectors. J Thorac Cardiovasc Surg. 2014;148:1656–1664.e2. doi: 10.1016/j.jtcvs.2014.03.033. [DOI] [PubMed] [Google Scholar]
- 76.Martin P, Albagli O, Poggi MC, Boulukos KE, Pognonec P. Development of a new bicistronic retroviral vector with strong IRES activity. BMC Biotechnol. 2006;6:4. doi: 10.1186/1472-6750-6-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Jeong YH, Park CH, Jang GH, Jeong YI, Hwang IS, Jeong YW, Kim YK, Shin T, Kim NH, Hyun SH, et al. Production of multiple transgenic Yucatan miniature pigs expressing human complement regulatory factors, human CD55, CD59, and H-transferase genes. PLoS One. 2013;8:e63241. doi: 10.1371/journal.pone.0063241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Koh EY, Ho SC, Mariati Z, Bi X, Bardor M, Yang Y. An internal ribosome entry site (IRES) mutant library for tuning expression level of multiple genes in mammalian cells. PLoS One. 2013;8:e82100. doi: 10.1371/journal.pone.0082100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Scappaticci FA, Contreras A, Smith R, Bonhoure L, Lum B, Cao Y, Engleman EG, Nolan GP. Statin-AE: a novel angiostatin-endostatin fusion protein with enhanced antiangiogenic and antitumor activity. Angiogenesis. 2001;4:263–268. doi: 10.1023/a:1016067717433. [DOI] [PubMed] [Google Scholar]
- 80.Tysome JR, Wang P, Alusi G, Briat A, Gangeswaran R, Wang J, Bhakta V, Fodor I, Lemoine NR, Wang Y. Lister vaccine strain of vaccinia virus armed with the endostatin-angiostatin fusion gene: an oncolytic virus superior to dl1520 (ONYX-015) for human head and neck cancer. Hum Gene Ther. 2011;22:1101–1108. doi: 10.1089/hum.2010.172. [DOI] [PubMed] [Google Scholar]
- 81.Cepko CL, Roberts BE, Mulligan RC. Construction and applications of a highly transmissible murine retrovirus shuttle vector. Cell. 1984;37:1053–1062. doi: 10.1016/0092-8674(84)90440-9. [DOI] [PubMed] [Google Scholar]
- 82.Korman AJ, Frantz JD, Strominger JL, Mulligan RC. Expression of human class II major histocompatibility complex antigens using retrovirus vectors. Proc Natl Acad Sci USA. 1987;84:2150–2154. doi: 10.1073/pnas.84.8.2150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Fallot S, Ben Naya R, Hieblot C, Mondon P, Lacazette E, Bouayadi K, Kharrat A, Touriol C, Prats H. Alternative-splicing-based bicistronic vectors for ratio-controlled protein expression and application to recombinant antibody production. Nucleic Acids Res. 2009;37:e134. doi: 10.1093/nar/gkp716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Doronina VA, Wu C, de Felipe P, Sachs MS, Ryan MD, Brown JD. Site-specific release of nascent chains from ribosomes at a sense codon. Mol Cell Biol. 2008;28:4227–4239. doi: 10.1128/MCB.00421-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.de Felipe P, Luke GA, Hughes LE, Gani D, Halpin C, Ryan MD. E unum pluribus: multiple proteins from a self-processing polyprotein. Trends Biotechnol. 2006;24:68–75. doi: 10.1016/j.tibtech.2005.12.006. [DOI] [PubMed] [Google Scholar]
- 86.Chan HY, V S, Xing X, Kraus P, Yap SP, Ng P, Lim SL, Lufkin T. Comparison of IRES and F2A-based locus-specific multicistronic expression in stable mouse lines. PLoS One. 2011;6:e28885. doi: 10.1371/journal.pone.0028885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.de Felipe P, Luke GA, Brown JD, Ryan MD. Inhibition of 2A-mediated ‘cleavage’ of certain artificial polyproteins bearing N-terminal signal sequences. Biotechnol J. 2010;5:213–223. doi: 10.1002/biot.200900134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Lotze MT, Zitvogel L, Campbell R, Robbins PD, Elder E, Haluszczak C, Martin D, Whiteside TL, Storkus WJ, Tahara H. Cytokine gene therapy of cancer using interleukin-12: murine and clinical trials. Ann N Y Acad Sci. 1996;795:440–454. doi: 10.1111/j.1749-6632.1996.tb52715.x. [DOI] [PubMed] [Google Scholar]
- 89.Tahara H, Zitvogel L, Storkus WJ, Zeh HJ, McKinney TG, Schreiber RD, Gubler U, Robbins PD, Lotze MT. Effective eradication of established murine tumors with IL-12 gene therapy using a polycistronic retroviral vector. J Immunol. 1995;154:6466–6474. [PubMed] [Google Scholar]
- 90.Kukuła K, Chojnowska L, Dąbrowski M, Witkowski A, Chmielak Z, Skwarek M, Kądziela J, Teresińska A, Małecki M, Janik P, et al. Intramyocardial plasmid-encoding human vascular endothelial growth factor A165/basic fibroblast growth factor therapy using percutaneous transcatheter approach in patients with refractory coronary artery disease (VIF-CAD) Am Heart J. 2011;161:581–589. doi: 10.1016/j.ahj.2010.11.023. [DOI] [PubMed] [Google Scholar]
- 91.Palfi S, Gurruchaga JM, Ralph GS, Lepetit H, Lavisse S, Buttery PC, Watts C, Miskin J, Kelleher M, Deeley S, et al. Long-term safety and tolerability of ProSavin, a lentiviral vector-based gene therapy for Parkinson’s disease: a dose escalation, open-label, phase 1/2 trial. Lancet. 2014;383:1138–1146. doi: 10.1016/S0140-6736(13)61939-X. [DOI] [PubMed] [Google Scholar]