

Abstract
Animal–vehicle collisions cause high levels of vertebrate mortality worldwide, and what goes wrong when animals fail to escape and ultimately collide with vehicles is not well understood. We investigated alert and escape behaviours of captive brown-headed cowbirds (Molothrus ater) in response to virtual vehicle approaches of different sizes and at speeds ranging from 60 to 360 km h−1. Alert and flight initiation distances remained similar across vehicle speeds, and accordingly, alert and flight initiation times decreased at higher vehicle speeds. Thus, avoidance behaviours in cowbirds appeared to be based on distance rather than time available for escape, particularly at 60–150 km h−1; however, at higher speeds (more than or equal to 180 km h−1) no trend in response behaviour was discernible. As vehicle speed increased, cowbirds did not have enough time to assess the approaching vehicle, and cowbirds generally did not initiate flight with enough time to avoid collision when vehicle speed exceeded 120 km h−1. Although potentially effective for evading predators, the decision-making process used by cowbirds in our study appears maladaptive in the context of avoiding fast-moving vehicles. Our methodological approach and findings provide a framework to assess how novel management strategies could affect escape rules, and the sensory and cognitive abilities animals use to avoid vehicle collisions.
Keywords: animal–vehicle collisions, antipredator behaviour, disturbance, escape response, flight initiation distance, video playback
1. Introduction
When approached by predators and other potential threats, animals must decide when to initiate an escape response. This decision is informed by characteristics of the oncoming object, such as size, speed and directness of approach [1], as well as the state or condition of the animal being threatened, including hunger level, experience and variation in risk-taking behaviours and personalities [2,3]. Animals combine sensory inputs with behavioural rules to assess the costs and benefits of fleeing and thus determine the timing of escape responses [4–6].
Animals appear to react to oncoming automobiles, aircraft and other non-biological threats in a qualitatively similar manner to predators [7,8]. During these encounters, animals use some variation of their antipredator repertoire [9], possibly because the evolutionary novelty of modern vehicles precludes more specialized responses [10]. However, vehicles and natural predators often differ in several important ways, including speed, size and consistency of approach. Such differences can lead to maladaptive (and often fatal) responses when faced with an oncoming vehicle, such as deer ‘freezing’ and turtles withdrawing into their shells while still on the road [10,11].
Animal–vehicle collisions, which kill hundreds of millions of birds and other animals each year [12–14], can negatively impact populations [15,16] and pose substantial safety risks to humans [17,18]. Yet, it is unclear what goes wrong when individual animals fail to escape and eventually collide with vehicles [10]. For instance, Legagneux & Ducatez [19] demonstrated that several species of birds escaped earlier from oncoming vehicles as the posted speed limit increased, but the actual speed of vehicles had no effect on escape behaviours. DeVault et al. [11] found that near-collisions with turkey vultures (Cathartes aura) increased with vehicle speed, suggesting that animals may have difficulty assessing the threat from high-speed vehicles. Another recent study, employing vehicle approaches at night, found that white-tailed deer (Odocoileus virginianus) did not adjust escape responses across vehicle speeds [20]. However, understanding the behavioural rules involved in these collisions at the individual level is challenging, because it is difficult to safely simulate the high speeds typical of automobiles on modern highways (ca 120 km h−1) or large jet aircraft during take-off and landing (ca 240 km h−1) without causing actual collisions.
We investigated the antipredator behavioural rules used for initiating alert and escape responses when animals are exposed to high-speed vehicle approaches. We developed an experimental protocol where individuals were exposed to video playback of an approaching vehicle in a manner similar to that used to investigate human pedestrian responses to cars in road-crossing scenarios [21]. Specifically, our goal was to examine responses of brown-headed cowbirds (Molothrus ater) to virtual vehicles of different sizes (standard and extended) appearing to approach at speeds from 60 to 360 km h−1. We were particularly interested in whether simulated ‘collisions’ occurred, and if so, at which vehicle size and speeds such collisions were common. Prey generally perceive enhanced threat and thus increase flight initiation (escape) distance when exposed to larger predator sizes and faster approach speeds [1,22], and humans are more cautious in road-crossing situations when faced with large oncoming vehicles than smaller ones [23].
We also examined which (if any) of the following behavioural rules might be in effect as birds attempt to avoid collisions with vehicles: (i) a temporal margin of safety, (ii) a fixed spatial margin of safety or (iii) a dynamic spatial margin of safety [24]. We considered each rule as a stand-alone hypothesis. The first rule is based on temporal decisions, whereas the other two are based on distance decisions. These behavioural rules comprise three key phases of animal–object interactions (figure 1): sensory detection (i.e. the vehicle becomes salient to the animal from background sensory noise), alert behaviour (i.e. the animal shows alert postures in response to the approaching threat) and flight initiation behaviour (i.e. the animal initiates escape by running or flying from the threat).
Figure 1.
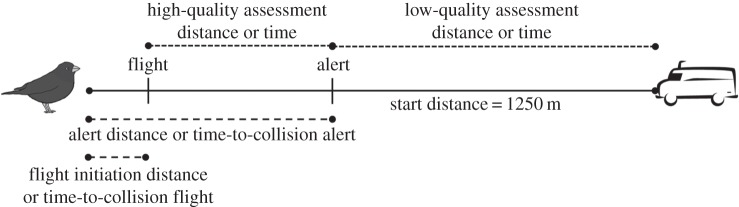
Response metrics of animals avoiding oncoming vehicles.
If flight initiation distance (FID) increases with vehicle speed but flight initiation time prior to collision remains constant, then a temporal margin of safety rule is apparent (i.e. the animal maintains a fixed amount of time for escape). If flight initiation time decreases as vehicle speed increases but FID remains constant, then a fixed spatial margin of safety is apparent (i.e. escape occurs at a fixed or threshold vehicle–animal separation distance). A dynamic spatial margin of safety is apparent when the animal allows a vehicle to approach to a fixed percentage of the alert distance (AD) before initiating escape [24]. Based on findings by DeVault et al. [11], we expected that cowbirds would use a spatial decision rule, at least at the lower vehicle speeds.
Finally, animals may not follow any of these behavioural rules because the speed of the vehicle may not provide them with enough time or distance to process information about the looming object [25]. There are two phases in which animals assess the risk of an oncoming object after sensory detection (figure 1): low-quality assessment and high-quality assessment [26]. Under low-quality assessment (between detection and alert; figure 1), the object is far away (i.e. low risk) and is monitored with parts of the visual field (e.g. retinal periphery) and body postures (e.g. head-down) that do not necessarily provide high-quality information [27]. Under high-quality assessment (between alert and flight; figure 1), the object is closer and is monitored directly with the centre of visual attention (e.g. fovea) and in information-gathering body postures (e.g. head-up; [26]). We could not evaluate low-quality assessment in our study, because we lacked the behavioural assays necessary to determine when the study animals detected the oncoming vehicle. However, we estimated empirically the minimum time and distance necessary for animals to determine the risk of an oncoming vehicle under the high-quality assessment phase and contrasted them with the time and distance available at the different vehicle speeds. Speeds at which the time available was less than the minimum time necessary for a decision would indicate that the animal's processing of the approaching threat during high-quality assessment was overwhelmed.
2. Material and methods
(a). Bird capture and care
We conducted our experiment at the National Aeronautics and Space Administration Plum Brook Station, Erie County, OH, USA (41°22′ N, 82°41′ W). Male brown-headed cowbirds (hereafter, cowbirds) were chosen as our model species (as in our previous work [28,29]) because they are plentiful in our area and relatively easy to capture in large numbers, respond well to being held in captivity, are regularly involved in vehicle collisions [14] and have a well-described visual system [28].
Cowbirds were captured continuously from late March through May 2012 using six 3.6 × 3.6 × 2.0 m modified Australian crow traps [30] located throughout the 2200-ha Plum Brook Station and held in 2.4 × 2.4 × 1.8 m cages in an indoor aviary illuminated with natural lighting. They were fed a mixture of white millet and sunflower seeds ad libitum and were given meal worms (Tenebrio molitor) once per week. Individuals were used within 7 to 14 days of capture and released unharmed after all experimental trials were completed. Because we have held cowbirds in our aviary for up to eight weeks with no apparent ill effects [29], we are confident that their behaviour in our experiment was not unduly affected by their confinement.
(b). Video playback
We used video playback as a means of simulating consistent treatment scenarios involving an approaching vehicle. Video playback has a long history in ornithological research [31,32] and is especially suitable for studies such as ours [33,34]. We provide details concerning video editing, validation and equipment in the electronic supplementary material.
We video-recorded approaches of a directly oncoming 2003 Ford F250 pickup truck using a Sony HD Handycam video camera with a resolution of 1080 × 1920 pixels (high definition) and a recording speed of 30 frames s−1 in MPEG-4 format. These recordings were used in playback experiments, although videos were played-back at double speed and 60 frames s−1 (see electronic supplementary material, table S1). We chose a consistently sunny and calm day (28 February 2012) to record vehicle approaches to minimize differences in ambient light and movement of vegetation from wind among recordings at different vehicle speeds. The camera was placed directly on the pavement in the middle of a closed, straight and flat road at Plum Brook Station (thus from the perspective of a bird on the road; [35]), facing the direction of the oncoming vehicle (electronic supplementary material, figure S1). We recorded approaches towards the video camera at seven actual vehicle speeds: 30, 45, 60, 75, 90, 105 and 120 km h−1. We began each vehicle approach from a distance of 1.25 km from the camera, within the range of cowbird visual perception [28]. During each approach, the driver (T.L.D.) quickly accelerated to the predetermined speed, and then set the vehicle speed control to maintain a constant speed until driving directly over the camera. The driver kept the vehicle in the middle of the road during the entire approach.
We made two video recordings at each vehicle speed: one using the vehicle with its standard frontal area, and one using the vehicle with an extended (2×) frontal area (except for the recording of vehicle approach at 120 km h−1, which presented logistical issues; see electronic supplementary material). The visual area of the truck was expanded by covering the front of the vehicle with a flat, dark-green fabric stretched over a plastic frame which extended laterally from the front of the truck. To control for vehicle colour across sizes, the same fabric was also used for the standard frontal area (electronic supplementary material, figure S2). We viewed all videos immediately after recording and repeated those in which a potential distraction was present (e.g. a bird flying across the scene, insects on the pavement in front of the camera) so that, to the best of our ability, all videos were identical except for the speed and size of the oncoming vehicle.
Our video editing resulted in eight apparent speed categories used during playback experiments: 60, 90, 120, 150, 180, 210, 240 and 360 km h−1 (details in electronic supplementary material, table S1). Given our previous estimate of the visual acuity of male cowbirds [28], we assumed that cowbirds would be able to detect the vehicle at the beginning of each virtual approach (1.25 km away). The vehicle was discernable from the background to us (although very small) on the TV monitor at the beginning of the approach.
We designed a video chamber (102 × 61 × 70 cm; electronic supplementary material, figure S3) to expose birds to virtual vehicle approach. The video chamber had three walls and a ceiling of plywood painted flat grey; the fourth wall consisted of a high-definition TV monitor with a visual surface of 102.0 × 57.5 cm (details are provided in the electronic supplementary material). The combination of the location of our camera recording the vehicle approach (i.e. at the level of a cowbird on the road; electronic supplementary material, figure S1) and the position of the video monitor (electronic supplementary material, figure S3) allowed birds a natural viewing angle of the approaching vehicle. All playbacks were silent, and the video chamber was located in a closed room and thus acoustically isolated.
(c). Experimental protocol
Cowbirds were placed in the video chamber and exposed to video playback in groups of three individuals (using groups provides a calming influence for social species [36,37]). Each combination of speed and size treatments included 10 groups (i.e. replicates); group served as the experimental unit (see below). Each cowbird group was exposed to only one vehicle approach, thus every trial used naive individuals to avoid habituation or sensitization effects. To begin a trial, a paused video file of the empty road was first placed on the monitor in ‘full-screen’ mode. We then released a group of three cowbirds inside the video chamber, closed the door to the chamber and played the video. During each video playback trial, the empty road was visible to cowbirds for about 11 min (before the vehicle approach began), and each video playback ended when the vehicle virtually passed over the video chamber, 11.5 min after birds were placed in the video chamber.
Because of initial limitations on bird availability, we first conducted video playback trials from 9 to 13 April 2012 using apparent speed categories 60, 120 and 180 km h−1 with the standard and extended vehicle sizes and 240 km h−1 at standard size. We added the seven remaining apparent speed/size categories (90, 150 and 210 km h−1 at standard and extended sizes and 360 km h−1 at standard size) from 18 April through 3 May 2012. We adjusted our analyses to incorporate a potential ordinal date effect on response behaviours (see below). We conducted video playbacks on 11 days, with an average of 12.7 trials per day (s.d. = 5.3). Notably, our virtual video chamber yielded results on FID (median = 28 m) that were similar to previous outdoor experiments with male cowbirds in enclosures responding to a real vehicle approach (ca 36 m ± s.d. of 8–10 across a variety of experimental conditions; [28,29]), providing confidence that our simulated vehicle approaches gave realistic results (sensu [31,38]).
(d). Behavioural metrics
We video-recorded cowbird response behaviours from three directions (through the rear wall and both sides of the video chamber; electronic supplementary material, figure S3) from the moment the birds were placed inside the video chamber until the vehicle passed over the camera (when the trial ended). We examined responses of each of the three cowbirds to the virtual oncoming vehicle frame-by-frame on a TV monitor and recorded behaviours to the nearest 1/15 s relative to the instant when the vehicle passed over the camera (the potential point of collision). At least two observers (T.L.D. and T.W.S.) viewed all recordings of cowbird reactions to interpret and quantify behaviours. We recorded the time (in seconds, before potential collision) that each individual showed an alert response and an escape response.
We defined an alert response to the vehicle approach as a marked transition in an individual's baseline behaviour (e.g. pecking, preening or loafing) to behaviours that might include head-up with neck extended, sudden and increased scanning behaviour, feather compression or sudden crouching [39]. Flight response was defined as an obvious intent to ‘escape’ the simulated oncoming vehicle, and generally entailed running or flying towards the back of the video chamber. During some trials, the birds quickly turned towards the back of the video chamber just before taking flight or running; on those occasions, we scored flight initiation time at the instant when they began their turn.
We scored individual alert time (time-to-collision alert; TTCalert) and flight initiation time (time-to-collision flight; TTCflight) as the time (s) required for the vehicle to reach the birds' location at the onset of the behaviour in question. Greater values of TTCalert and TTCflight indicate an earlier response to the approaching vehicle. In instances where an individual showed no alert behaviour, we scored TTCalert as equivalent to TTCflight. We converted TTCalert and TTCflight to AD and FID, respectively, by incorporating vehicle speed using the following equations:
| 2.1 |
and
| 2.2 |
Here, VAS = vehicle approach speed (km h−1) and the constant (0.2778) is the conversion factor used when AD and FID are expressed in metres [11].
In addition to becoming alert to vehicle approach and initiating the escape response, a bird must respond with enough time to cover the distance necessary to avoid the oncoming vehicle. We thus conducted a field experiment in which we measured the time necessary for cowbirds to travel 3 m (roughly, the width of one lane in a standard road) from a stationary position (see the electronic supplementary material). This allowed us to estimate the time (0.80 s) cowbirds need to clear the path of an oncoming vehicle and thus avoid a ‘collision’. We also established (post hoc) the minimum high-quality assessment times (using only birds that exhibited an alert response) that cowbirds showed in avoiding a ‘collision’ with the virtual vehicle. We sorted the trials where both alert and flight responses were recorded and classified the outcome of each trial as birds escaping or (virtually) colliding with the vehicle based on the time necessary to avoid the collision (0.80 s; see the electronic supplementary material). The minimum high-quality assessment time that led to successful escapes was 0.06 s.
(e). Analyses
We selected the median response (alert and flight) within a group of three cowbirds (i.e. the second bird to respond) as the more accurate measurement of central tendency than the mean (see also [11]), although birds within a group usually reacted at about the same time and results using mean values were qualitatively similar. We first assessed whether behavioural escape rules followed temporal or distance decisions by pooling information from all speeds. A significant effect of speed on FID, but a non-significant effect of speed on TTCflight, would support the idea that birds maintained a temporal margin of safety. The reverse scenario (speed significantly affecting TTCflight, but no effect on FID) would support the idea that birds maintained a fixed or dynamic spatial margin of safety. To accomplish this, we used linear mixed models with vehicle speed, vehicle size and their interaction as fixed independent variables, ordinal date as a random effect, and TTCalert, TTCflight, AD and FID as the response variables. The inclusion of ordinal date allowed us to assess effects on alert and flight behaviours that might be associated with progression of the breeding season. We assumed a Gaussian distribution (via PROC GLIMMIX; SAS v. 9.2, SAS, Inc., Cary, NC, USA), examined model residuals and found that they were normally distributed.
A temporal response was not evident (see below), therefore we explored further whether our data reflected a fixed spatial margin of safety or a dynamic spatial margin of safety by conducting analyses at each speed, because the same behavioural escape rule might not be used across all vehicle speeds [11]. Specifically, we examined the response of the ratio FID/AD relative to AD (see the electronic supplementary material for details) for each vehicle speed category 60–210 km h−1 (n = 20 trials per speed). We then fitted a line describing the relationship between FID/AD and AD, including a second-order polynomial term (to allow for nonlinearity), and compared those relationships to those expected for the three distance rules (electronic supplementary material, figure S4). Specifically, for a fixed spatial margin of safety (in which the animal allows the threat to approach to a fixed separation distance independent of AD), the ratio FID/AD would decrease with AD to an asymptote approaching the x-axis as AD increases. For a dynamic spatial margin of safety, FID/AD would exhibit a (horizontal) line of zero slope, likely near 0.5 [24,40].
However, it is conceivable that vehicle speed could hinder an animal's successful adherence to an escape rule, such that there is not enough time or distance to process information about the looming object [25]. For example, an alert response might provide ample time for a subsequent flight response, but vehicle speed could still overwhelm the time necessary to process and act on the stimulus. To assess at which point of the approach animals might be so limited in time or distance, we examined the effects of vehicle speed, size, speed × size (fixed effects) and ordinal date (random effect) on high-quality assessment time and distance using linear mixed models and assuming a Gaussian distribution. We examined model residuals and found they were normal. For each vehicle speed, high-quality assessment time was defined as the elapsed time from alert behaviour until flight. If no alert was apparent, high-quality assessment time was zero. Further, we defined the high-quality assessment distance as the distance between the vehicle's location at the points of alert and flight behaviour (figure 1). As with high-quality assessment time, if no alert was apparent, high-quality assessment distance was zero.
Finally, in addition to understanding how escape decisions are made, we determined whether the animal would have enough time to successfully escape at different speeds during the high-quality assessment phase. In this analysis, we used a subset of our data for which we were able to measure alert responses (i.e. those data points for which we were able to calculate the high-quality assessment time). This post hoc analysis therefore does not include the instances where alert responses were scored as zero, but birds ‘survived’ (see below), thus inference is limited to this subset of the data. We classified each trial as ‘not enough time’ or ‘enough time’ to successfully escape by subtracting the minimum time estimated to avoid a collision (see Behavioural metrics) from the recorded high-quality assessment time. We then calculated at each speed the proportion of trials in which the animals had or did not have enough time to escape and analysed the data with a log-linear analysis.
Because we did not have trials with the extended vehicle for apparent speed categories 240 and 360 km h−1 (see the electronic supplementary material for details), all linear mixed models included only apparent speed categories 60–210 km h−1 (n = 20 trials per speed). However, we present all response data in our descriptive statistics.
3. Results
Each group of three cowbirds generally appeared calm after being placed into the video chamber. Most alert reactions were obvious, although for 28.6% of groups (n = 40) no alert behaviours were observed before initiation of escape. However, all groups showed obvious escape behaviour in response to a playback of the oncoming vehicle. Across treatments, median TTCalert was 1.02 s (mode = 0.73; range = 0.20–5.30 s), median TTCflight was 0.67 s (mode = 0.34; range = 0.12–3.00 s), median AD was 43.3 m (mode = 58.3 m; range = 10.2–100.0 m) and median FID was 28.0 m (mode = 28.0; range = 5.1–81.3 m).
Times-to-collision were not constant across speeds. TTCalert (F5,99 = 31.00; p < 0.001) and TTCflight (F5,99 = 33.02; p < 0.001; electronic supplementary material, table S2) significantly decreased as vehicle speed increased (figure 2a,b). Cowbirds generally did not initiate flight with enough time to avoid collision (0.8 s needed) when vehicle speed exceeded 120 km h−1 (figure 2b). On the other hand, alert and escape distances remained relatively constant across speeds. AD (F5,99 = 0.91; p = 0.479) and FID (F5,99 = 1.41; p = 0.230; electronic supplementary material, table S2) were not significantly affected by vehicle speed (figure 2c,d). Vehicle size, the interaction of vehicle speed × vehicle size and ordinal date did not influence TTCalert, TTCflight, AD or FID (electronic supplementary material, table S2). These analyses indicate that cowbirds followed a distance-based rather than temporal escape rule.
Figure 2.

Reactions of brown-headed cowbirds to virtual oncoming vehicles approaching at apparent speeds of 60–360 km h−1. Panels (a) and (b) represent alert and flight behaviours, respectively, expressed as time-to-collision (s); panels (c,d) represent the same behaviours expressed as distance to collision (m). Two vehicle sizes (standard and extended) are combined for apparent speeds 60–210 (n = 20 cowbird groups for each), but only standard vehicles were used for approaches at 240 and 360 km h−1 (n = 10 groups for each). Bars represent means ± 1 s.e. The horizontal dotted line in the time-to-collision panels represents the time necessary (0.8 s) for cowbirds to travel 3 m from a resting position (i.e. successfully escape the oncoming vehicle; see the electronic supplementary material).
At the lower vehicle speeds (less than 180 km h−1), we found that the relationships between FID/AD and AD were generally negative (figure 3), arguing against a dynamic spatial margin of safety rule. These negative relationships suggest that birds which quickly became alert to the oncoming vehicle generally delayed escape behaviour relative to birds that became alert to the vehicle later in the approach. Further, the FID/AD relationships at vehicle speeds less than 180 k h−1 best approximated that expected from a fixed spatial margin of safety (electronic supplementary material, figure S4), although there was no apparent asymptotic relationship with AD (i.e. to the x-axis).
Figure 3.
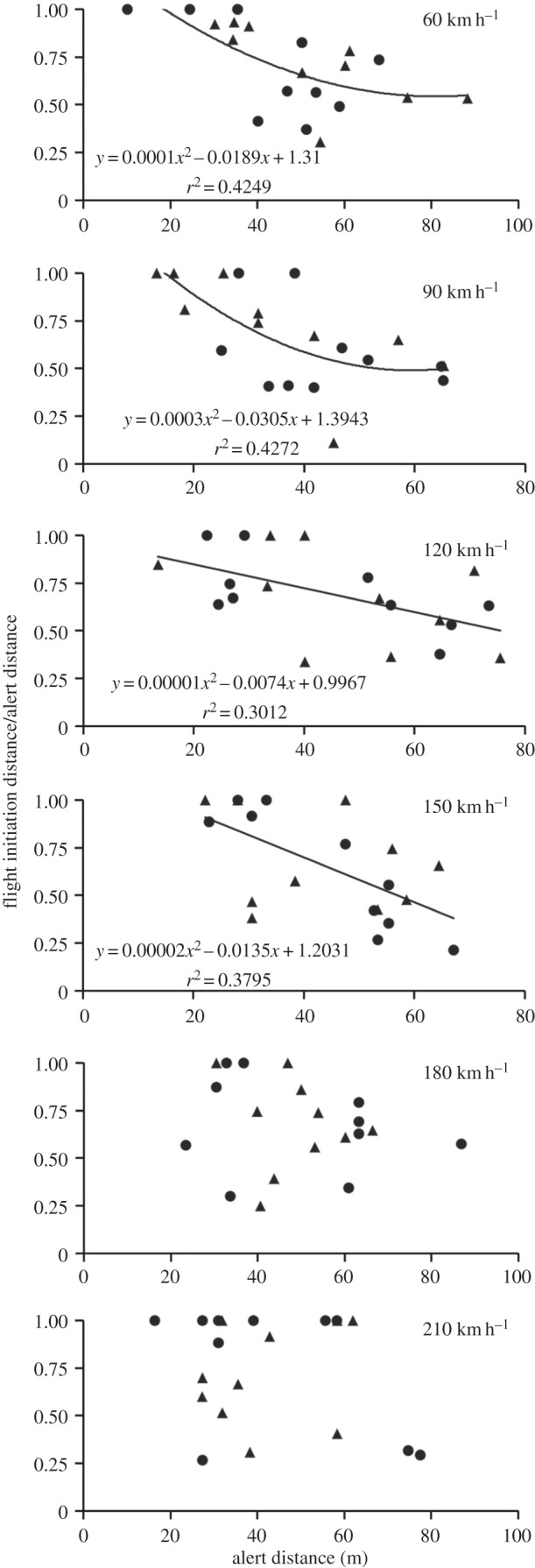
Scatterplots depicting the ratio of FID to AD versus AD across vehicle speed treatments. Circles represent trials using the standard truck size; triangles represent trials using the extended truck size. Regression lines, equations and r2 values are shown where relationships are significant (p ≤ 0.05). A second-order polynomial term was included in all models to account for potential asymptotic relationships characteristic of that expected from a fixed spatial margin of safety (see also electronic supplementary material, figure S4).
Also, our data indicate that cowbirds might not successfully use the same escape strategy across all vehicle speeds. For example, the relationships between FID/AD and AD at 180 and 210 km h−1 were negative but non-significant (p > 0.33). This lack of significance may reflect a general breakdown of all relevant behavioural rules at very high vehicle speeds (see below). We also note that the proportion of trials for which FID/AD = 1 (trials for which no alert behaviour was apparent) increased with vehicle speed (figure 4). Because AD does not vary with vehicle speed (figure 2c), the time available for escape is increasingly limited at higher speeds. As a result, we were unable, in these instances, to distinguish an alert from flight response as per our definitions.
Figure 4.
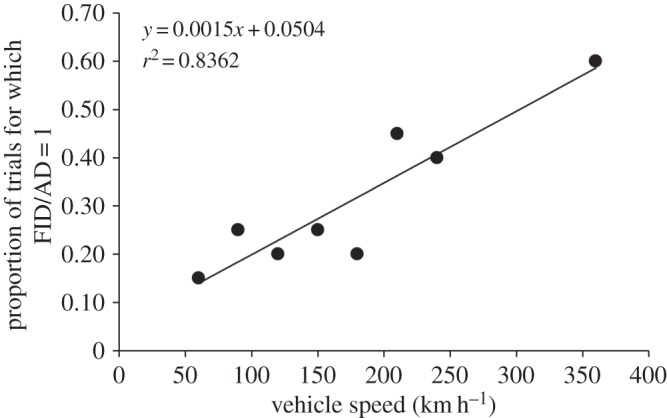
Proportion of trials, by vehicle speed, for which FID divided by AD = 1 (i.e. trials for which no alert behaviour was apparent).
High-quality assessment time (F5,99 = 6.13; p < 0.001; electronic supplementary material, table S3) was significantly affected by vehicle speed, decreasing by 91% from 60 to 360 km h−1 (figure 5). High-quality assessment distance (F5,99 = 0.34; p = 0.887; electronic supplementary material, table S3) was not affected by vehicle speed. Vehicle size, the interaction of vehicle speed × vehicle size and ordinal date did not influence high-quality assessment time or distance (electronic supplementary material, table S3).
Figure 5.
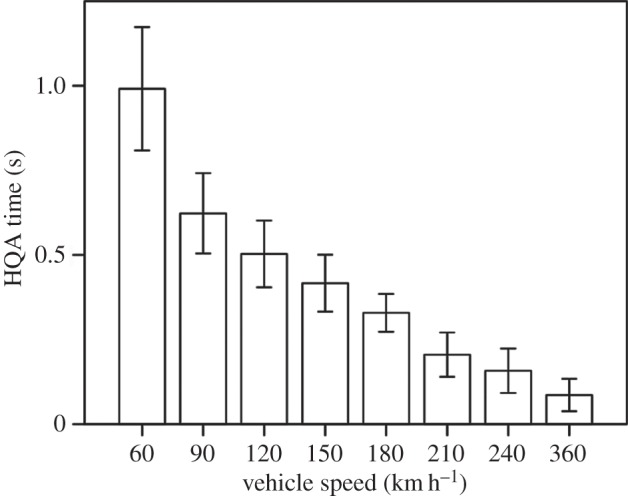
Bar chart of high-quality assessment (HQA) time (time difference between alert and flight behaviours) of brown-headed cowbirds in response to virtual vehicles approaching at apparent speeds of 60–360 km h−1. Two vehicle sizes (standard and extended) are combined for apparent speeds 60–210 (n = 20 cowbird groups for each), but only standard vehicles were used for approaches at 240 and 360 km h−1 (n = 10 groups for each). Bars represent means ± 1 s.e.
Finally, we found that animals generally did not have enough time for assessing risk in the high-quality assessment phase at speeds greater than 180 km h−1 (Pearson 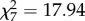 , p = 0.012; figure 6) suggesting that vehicle speed might have overwhelmed processing of the approaching threat during the high-quality assessment phase.
, p = 0.012; figure 6) suggesting that vehicle speed might have overwhelmed processing of the approaching threat during the high-quality assessment phase.
Figure 6.
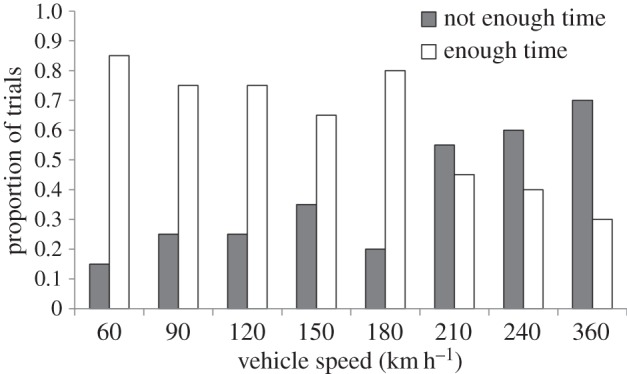
Proportion of trials for high-quality assessment time in which the estimated minimum time threshold for a successful escape was (enough time) or was not (not enough time) met. See text for details.
4. Discussion
Brown-headed cowbirds in our study usually managed to respond quickly enough to avoid virtual collisions during simulated low-speed vehicle approaches, but they were often overwhelmed by high-speed approaches. Cowbirds did not adjust escape responses for differences in vehicle speed, but instead appeared to use a distance rule. At high simulated vehicle speeds, the lack of adjustment in FID resulted in little time for high-quality assessment of the approaching vehicle, as well as its avoidance at the onset of escape responses. For example, cowbirds usually initiated escape responses with less than 0.8 s until collision (time needed for escape) when the vehicle approached at speeds greater than 120 km h−1 (figure 2b). This type of late response could result in a high risk of collision when birds are faced with highway traffic or aircraft during take-off and landing.
Our study has limitations that should be taken into account. First, because we used only brown-headed cowbirds in our experiments, we caution that our results might not be representative of all species. Vehicle avoidance behaviour is known to differ across bird species due to differences such as relative brain size [41], and solitary birds might react differently than individuals in a group that can observe alert behaviours of conspecifics. Also, a recent study demonstrated that female cowbirds have lower visual resolution than male cowbirds [42], thus it is possible that female cowbirds might respond to vehicle approach differently than the males used in our study. Second, the absence of noise during video playback reduced the sensory cues available to birds during vehicle avoidance behaviours. The somewhat greater FIDs observed for cowbirds responding to a real vehicle approach [28,29] may reflect this difference.
Despite these limitations, our study is the first to provide direct evidence that the behavioural escape rules used by birds are overwhelmed at higher vehicle speeds. Because flight initiation time (TTCflight) decreased as vehicle speed increased but FID remained constant, cowbirds appeared to monitor aspects of distance rather than time when attempting to avoid oncoming vehicles. More specifically, at vehicle speeds less than 180 km h−1 cowbird responses approximated that expected from a fixed spatial margin of safety, i.e. reacting at a threshold distance. However, at the highest two vehicle speeds considered (180 and 210 km h−1), the escape strategy employed (if any) was unclear. Thus, our findings complement those reported by DeVault et al. [11], who found that FIDs of turkey vultures responding to an approaching vehicle were similar at vehicle speeds of 30 and 60 km h−1, although FIDs became highly variable at 90 km h−1 (the highest speed tested). It is conceivable that the physiological mechanisms or behavioural strategies used by birds (and potentially other animals) to avoid oncoming objects might ‘break down’ as object speed increases and exceeds those naturally encountered [11]. Indeed, the emergence of high-speed vehicles over the past 100 years as a novel threat challenges the effectiveness of distance-based escape strategies for all dangerous oncoming objects.
Our analysis indicated that birds which showed earlier alert behaviours allowed for a proportionally longer approach before initiating escape than birds that became alert to the vehicle later in the approach (figure 3). This result appears to oppose that predicted by the ‘flush early and avoid the rush’ hypothesis [6,43,44], which holds that FID increases in proportion to AD because of the attentional costs associated with monitoring oncoming predators. However, in some situations, animals might perceive vehicles differently than natural predators [1,10]. Vehicle approach does not involve the full complement of predator approach stimuli (e.g. form, appendage movement and eye direction) that is characteristic of humans or predators [45]. Thus, it is unclear how well previous studies on predator avoidance (e.g. [6]) apply to the problem of vehicle collisions, which often entail, among these other differences, much higher object speeds.
The apparent inadequacy of cowbird responses relative to vehicle speed is further demonstrated in our finding of shorter high-quality assessment times as vehicle speed increased. Vehicle speed could compromise survival through the inability of individuals to adjust their high-quality assessment time, mostly at speeds more than 180 km h−1 (figure 6). Therefore, at speeds of typical automobiles on highways (ca 120 km h−1) or jet aircraft during take-off (ca 240 km h−1), the risk of collision might be affected by a lack of time for assessment.
Vehicle size (frontal area) had no effect on cowbird alert and escape responses. This was an unexpected result, because a larger oncoming object should produce an expanded looming image and thus trigger a quicker escape response [7,25]. The lack of a size effect may be due to our stimulus being only laterally enlarged. Because of safety concerns (i.e. visibility of the driver), we were unable to expand the frontal area of the truck vertically (electronic supplementary material, figure S2). Another possibility is that the looming stimulus was well above the threshold size that would lead to a neuronal response [25,46].
We envision an application of our findings in aspects of road ecology [47] where speed limits within critical conservation areas might be reconsidered [20], as well as in efforts to enhance detection of vehicles where lowering vehicle speed is not logistically possible (e.g. bird–aircraft collisions, some highways). In the latter scenario, it is conceivable that by enhancing detection of an approaching vehicle, avoidance behaviours could be initiated sooner [28,48,49]. For example, the activation of pulsating lights on aircraft could lengthen AD [49], possibly leading to an increase in FID and thus an enhanced probability of survival during encounters with vehicles.
In some species, an escape strategy based on distance rather than time might be beneficial (i.e. when predators approach at predictable speeds) to reliably elicit avoidance manoeuvres with sufficient time for escape [6,7,25]. However, our study suggests that the distance rule used by cowbirds is generally ineffective for avoiding high-speed vehicles. This may result from information-processing mechanisms [25] not being well suited to avoiding vehicles. Alternatively, cowbirds might establish consistent FIDs based on their prior experience with vehicles (sensu [19]), and thus have difficulty adjusting to faster, unfamiliar vehicle speeds. Irrespective of the mechanism employed, our findings provide a framework and methodological approach to further investigate how different management strategies (e.g. lights on vehicles, vehicle coloration; [49,50]) affect the sensory and behavioural abilities of animals to assess risk and initiate escape in time to avoid collisions with modern vehicles.
Supplementary Material
Acknowledgements
We thank L. Tyson for help with bird care and M. Majoy for technical assistance with our video system. B. Washburn and two anonymous reviewers provided helpful comments on the manuscript.
Ethics statement
The Institutional Animal Care and Use Committee of the US Department of Agriculture, Wildlife Services, National Wildlife Research Center approved all procedures used in this study (QA-1961).
Data accessibility
Data from this study are archived with data dryad: doi:10.5061/dryad.7154q.
Funding statement
The US Department of Agriculture and Federal Aviation Administration funded this research.
References
- 1.Stankowich T, Blumstein DT. 2005. Fear in animals: a meta-analysis and review of risk assessment. Proc. R. Soc. B 272, 2627–2634. ( 10.1098/rspb.2005.3251) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lima SL, Dill LM. 1990. Behavioral decisions made under the risk of predation: a review and prospectus. Can. J. Zool. 68, 619–640. ( 10.1139/z90-092) [DOI] [Google Scholar]
- 3.Carrete M, Tella JL. 2010. Individual consistency in flight initiation distances in burrowing owls: a new hypothesis on disturbance-induced habitat selection. Biol. Lett. 6, 167–170. ( 10.1098/rsbl.2009.0739) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Blumstein DT. 2003. Flight initiation distance in birds is dependent on intruder starting distance. J. Wildl. Manag. 67, 852–857. ( 10.2307/3802692) [DOI] [Google Scholar]
- 5.Cooper WE, Jr, Frederick WG. 2007. Optimal flight initiation distance. J. Theor. Biol. 244, 59–67. ( 10.1016/j.jtbi.2006.07.011) [DOI] [PubMed] [Google Scholar]
- 6.Cooper WE, Jr, Blumstein DT. 2014. Novel effects of monitoring predators on costs of fleeing and not fleeing explain flushing early in economic escape theory. Behav. Ecol. 25, 44–52. ( 10.1093/beheco/art083) [DOI] [Google Scholar]
- 7.Dill LM. 1974. The escape response of the zebra danio (Brachydanio rerio) I. The stimulus for escape. Anim. Behav. 22, 711–722. ( 10.1016/S0003-3472(74)80022-9) [DOI] [Google Scholar]
- 8.Bernhardt GE, Blackwell BF, DeVault TL, Kutchbach-Brohl L. 2010. Fatal injuries to birds from collisions with aircraft reveal antipredator behaviours. Ibis 152, 830–834. ( 10.1111/j.1474-919X.2010.01043.x) [DOI] [Google Scholar]
- 9.Frid A, Dill L. 2002. Human-caused disturbance stimuli as a form of predation risk. Conserv. Ecol. 6, 11. [Google Scholar]
- 10.Lima SL, Blackwell BF, DeVault TL, Fernández-Juricic E. 2014. Animal reactions to oncoming vehicles: a conceptual review. Biol. Rev. ( 10.1111/brv.12093) [DOI] [PubMed] [Google Scholar]
- 11.DeVault TL, Blackwell BF, Seamans TW, Lima SL, Fernández-Juricic E. 2014. Effects of vehicle speed on flight initiation by turkey vultures: implications for bird-vehicle collisions. PLoS ONE 9, e87944 ( 10.1371/journal.pone.0087944) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Erritzoe J, Mazgajski TD, Rejt L. 2003. Bird casualties on European roads—a review. Acta Ornithol. 38, 77–93. ( 10.3161/068.038.0204) [DOI] [Google Scholar]
- 13.Bishop CA, Brogan JM. 2013. Estimates of avian mortality attributed to vehicle collisions in Canada. Avian Conserv. Ecol. 8, 2 ( 10.5751/ACE-00604-080202) [DOI] [Google Scholar]
- 14.Loss SR, Will T, Marra PP. 2014. Estimation of bird-vehicle collision mortality on U.S. roads. J. Wildl. Manag. 78, 763–771. ( 10.1002/jwmg.721) [DOI] [Google Scholar]
- 15.Fahrig L, Rytwinski T. 2009. Effects of roads on animal abundance: an empirical review and synthesis. Ecol. Soc. 14, 21. [Google Scholar]
- 16.Kociolek AV, Clevenger AP, St Clair CC, Proppe DS. 2011. Effects of road networks on bird populations. Conserv. Biol. 25, 241–249. [DOI] [PubMed] [Google Scholar]
- 17.Conover MR. 2002. Resolving human–wildlife conflicts: the science of wildlife damage management. Boca Raton, FL: CRC Press. [Google Scholar]
- 18.DeVault TL, Blackwell BF, Belant JL. (eds). 2013. Wildlife in airport environments: preventing animal–aircraft collisions through science-based management. Baltimore, MD: Johns Hopkins University Press. [Google Scholar]
- 19.Legagneux P, Ducatez S. 2013. European birds adjust their flight initiation distance to road speed limits. Biol. Lett. 9, 20130417 ( 10.1098/rsbl.2013.0417) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Blackwell BF, Seamans TW, DeVault TL. 2014. White-tailed deer response to vehicle approach: evidence of unclear and present danger. PLoS ONE 9, e109988 ( 10.1371/journal.pone.0109988) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Neider MB, McCarley JS, Crowell JA, Kaczmarski H, Kramer AF. 2010. Pedestrians, vehicles, and cell phones. Accid. Anal. Prev. 42, 589–594. ( 10.1016/j.aap.2009.10.004) [DOI] [PubMed] [Google Scholar]
- 22.Cooper WE, Jr, Stankowich T. 2010. Prey or predator? Body size of an approaching animal affects decisions to attack or escape. Behav. Ecol. 21, 1278–1284. ( 10.1093/beheco/arq142) [DOI] [Google Scholar]
- 23.Das S, Manski CF, Manuszak MD. 2005. Walk or wait? An empirical analysis of street crossing decisions. J. Appl. Econ. 20, 529–548. ( 10.1002/jae.791) [DOI] [Google Scholar]
- 24.Cárdenas YL, Shen B, Zung L, Blumstein DT. 2005. Evaluating temporal and spatial margins of safety in galahs. Anim. Behav. 70, 1395–1399. ( 10.1016/j.anbehav.2005.03.022) [DOI] [Google Scholar]
- 25.Sun H, Frost BJ. 1998. Computation of different optical variables of looming objects in pigeon nucleus rotundus neurons. Nat. Neurosci. 1, 296–303. ( 10.1038/1110) [DOI] [PubMed] [Google Scholar]
- 26.Tyrrell LP, Fernández-Juricic E. In press Sensory systems and escape behavior. In Escaping from predators: an integrative view of escape decisions (eds Cooper WE, Blumstein DT.). Cambridge, UK: Cambridge University Press. [Google Scholar]
- 27.Lima SL, Bednekoff PA. 1999. Back to the basics of anti-predatory vigilance: can non-vigilant animals detect attack? Anim. Behav. 58, 537–543. ( 10.1006/anbe.1999.1182) [DOI] [PubMed] [Google Scholar]
- 28.Blackwell BF, Fernández-Juricic E, Seamans TW, Dolan T. 2009. Avian visual system configuration and behavioural response to object approach. Anim. Behav. 77, 673–684. ( 10.1016/j.anbehav.2008.11.017) [DOI] [Google Scholar]
- 29.Blackwell BF, Bernhardt GE. 2004. Efficacy of aircraft landing lights in stimulating avoidance behavior in birds. J. Wildl. Manag. 68, 725–732. ( 10.2193/0022-541X(2004)068[0725:EOALLI]2.0.CO;2) [DOI] [Google Scholar]
- 30.Dolbeer RA, Holler NR, Hawthorne DW. 1995. Identification and control of wildlife damage. In Research and management techniques for wildlife and habitats (ed. Bookhout TA.), pp. 474–506. Bethesda, MD: The Wildlife Society. [Google Scholar]
- 31.D'Eath RB. 1998. Can video images imitate real stimuli in animal behavior experiments? Biol. Rev. 73, 267–292. ( 10.1017/S0006323198005179) [DOI] [Google Scholar]
- 32.Woo KL, Rieucau G. 2011. From dummies to computer-animated stimuli: a synthesis of techniques employed to stage animal interactions. Behav. Ecol. Sociobiol. 65, 1671–1685. ( 10.1007/s00265-011-1226-y) [DOI] [Google Scholar]
- 33.Gibson JJ. 1986. The ecological approach to visual perception. New York, NY: Taylor and Francis Group. [Google Scholar]
- 34.Lea SEG, Dittrich WH. 1999. What do birds see in moving video images? Curr. Psychol. Cogn. 18, 765–803. [Google Scholar]
- 35.Zeil J. 2000. Depth cues, behavioural context, and natural illumination: some potential limitations of video playback techniques. Acta Ethol. 3, 39–48. ( 10.1007/s102110000021) [DOI] [Google Scholar]
- 36.Fernández-Juricic E, Siller S, Kacelnik A. 2004. Flock density, social foraging, and scanning: an experiment with starlings. Behav. Ecol. 15, 371–379. ( 10.1093/beheco/arh017) [DOI] [Google Scholar]
- 37.Lowther PE. 1993. Brown-headed cowbird (Molothrus ater). In The birds of North America online (ed. Poole A.), monograph No. 47 Ithaca, NY: Cornell Lab of Ornithology; ( 10.2173/bna.47) [DOI] [Google Scholar]
- 38.Fleishman LJ, Endler JA. 2000. Some comments on visual perception and the use of video playback in animal behavior studies. Acta Ethol. 3, 15–27. ( 10.1007/s102110000025) [DOI] [Google Scholar]
- 39.Fernández-Juricic E, Jimenez MD, Lucas E. 2001. Alert distance as an alternative measure of bird tolerance to human disturbance: implications for park design. Environ. Conserv. 28, 263–269. ( 10.1017/S0376892901000273) [DOI] [Google Scholar]
- 40.Gulbransen D, Segrist T, del Castillo P, Blumstein DT. 2006. The fixed-slope rule: an inter-specific study. Ethology 112, 1056–1061. ( 10.1111/j.1439-0310.2006.01265.x) [DOI] [Google Scholar]
- 41.Husby A, Husby M. 2014. Interspecific analysis of vehicle avoidance behavior in birds. Behavioral Ecology. 25, 504–508. ( 10.1093/beheco/aru011) [DOI] [Google Scholar]
- 42.Fernández-Juricic E, Ojeda A, Deisher M, Burry B, Baumhardt P, Stark A, Elmore AG, Ensminger AL. 2013. Do male and female cowbirds see their world differently? Implications for sex differences in the sensory system of an avian brood parasite. PLoS ONE 8, e58985 ( 10.1371/journal.pone.0058985) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Samia DSM, Nomura F, Blumstein DT. 2013. Do animals generally flush early and avoid the rush? A meta-analysis. Biol. Lett. 9, 20130016 ( 10.1098/rsbl.2013.0016) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Blumstein DT. 2010. Flush early and avoid the rush: a general rule of antipredator behavior? Behav. Ecol. 21, 440–442. ( 10.1093/beheco/arq030) [DOI] [Google Scholar]
- 45.Whittington J, St Clair CC, Mercer G. 2004. Path tortuosity and the permeability of roads and trails to wolf movement. Ecol. Soc. 9, 4. [Google Scholar]
- 46.Stankowich T, Coss RG. 2005. Effects of predator behavior and proximity on risk assessment by Columbian black-tailed deer. Behav. Ecol. 17, 246–254. ( 10.1093/beheco/arj020) [DOI] [Google Scholar]
- 47.Forman RTT, et al. 2003. Road ecology: science and solutions. Washington, DC: Island Press. [Google Scholar]
- 48.Blackwell BF, Seamans TW. 2009. Enhancing the perceived threat of vehicle approach to deer. J. Wildl. Manag. 73, 128–135. ( 10.2193/2008-014) [DOI] [Google Scholar]
- 49.Blackwell BF, DeVault TL, Seamans TW, Lima SL, Baumhardt P, Fernández-Juricic E. 2012. Exploiting avian vision with aircraft lighting to reduce bird strikes. J. Appl. Ecol. 49, 758–766. ( 10.1111/j.1365-2664.2012.02165.x) [DOI] [Google Scholar]
- 50.Fernández-Juricic E, Gaffney J, Blackwell BF, Baumhardt P. 2011. Bird strikes and aircraft fuselage color: a correlational study. Hum. Wildl. Interact. 5, 224–234. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data from this study are archived with data dryad: doi:10.5061/dryad.7154q.