

Abstract
The prefrontal cortex appears to contribute to the mnemonic retrieval of the context within which stimuli are experienced, but only under certain conditions that remain to be clarified. Patients with lesions to the frontal cortex, the temporal lobe and neurologically intact individuals were tested for context memory retrieval when verbal stimuli (words) had been experienced across multiple (unstable context condition) or unique (stable context condition) contexts; basic recognition memory of these words-in-contexts was also tested. Patients with lesions to the right ventrolateral prefrontal cortex (VLPFC) were impaired on context retrieval only when the words had been seen in multiple contexts, demonstrating that this prefrontal region is critical for active retrieval processing necessary to disambiguate memory items embedded across multiple contexts. Patients with lesions to the left dorsomedial prefrontal region were impaired on both context retrieval conditions, regardless of the stability of the stimulus-to-context associations. Conversely, prefrontal lesions sparing the ventrolateral and dorsomedial regions did not impair context retrieval. Only patients with temporal lobe excisions were impaired on basic recognition memory. The results demonstrate a basic contribution of the left dorsomedial frontal region to mnemonic context retrieval, with the VLPFC engaged, selectively, when contextual relations are unstable and require disambiguation.
Keywords: prefrontal cortex, context retrieval, memory retrieval, ventrolateral prefrontal cortex, dorsomedial frontal cortex, hippocampus
1. Introduction
There is consensus on the essential involvement of medial temporal lobe structures, such as the hippocampus and parahippocampal cortex, in various aspects of declarative memory, including context retrieval [1–5]. By contrast, the contribution of the frontal cortex in mnemonic context retrieval remains a matter of debate. There is functional neuroimaging and electrophysiological evidence for greater prefrontal cortex activation during the retrieval of the contextual information associated with an event compared with retrieval of the event itself [6–8]. Similarly, impairment in memory for source (i.e. context) but not for item has been reported following large frontal lesions [9–12], suggesting a critical role of the prefrontal cortex in context retrieval. However, source memory impairments have not always been reported in patients with lesions clearly restricted to the prefrontal cortex [13,14], raising the question of the precise conditions under which context retrieval depends on different parts of the large and anatomically heterogeneous prefrontal cortex.
It is often argued that the contribution of the prefrontal cortex in memory is indirect in the sense that it reflects various control processes that may be critical for memory retrieval under particular circumstances [15–19]. Specifically, it has been argued that the ventrolateral prefrontal region (areas 45 and 47/12) is critical for active controlled retrieval that would become increasingly important when stimuli are linked to multiple contexts with equal probability creating ambiguous relations between items and their contexts [15,16]. By contrast, the ventrolateral prefrontal region would not be necessary for mnemonic context retrieval that can be based on strong and stable stimulus-to-context relations [15,16]. Thus, the engagement of the prefrontal cortex in context/source retrieval is not obligatory and the ventrolateral prefrontal cortex (VLPFC) becomes necessary as ambiguity in item-to-context relations increases. This specific hypothesis was tested with functional neuroimaging, and evidence was provided for selective increases in activity in the VLPFC when human subjects were retrieving specific stimulus features that had occurred in association with multiple contexts [20–23], but direct evidence of the critical involvement of an area in such retrieval, which can only be provided by lesion studies, is not available.
There is, however, evidence that lesions that involve the ventrolateral region of the frontal cortex in the left hemisphere impair the retrieval of semantic information under conditions of high selection competition demands [24–26]. There is also functional neuroimaging evidence of the involvement of the left ventrolateral prefrontal region in verbal recall under conditions that require selective verbal retrieval, such as the free recall of words that appeared within particular contexts (lists) [27] and verbal fluency, which can be viewed as a form of selective verbal retrieval [28,29]. Another frontal region that has been implicated in verbal fluency is the left dorsomedial prefrontal region [30–32], raising the question whether this region may also be involved in the retrieval of stimulus-to-context relations.
This study tested the above predictions by examining the performance of patients with damage to the frontal cortex on three memory retrieval conditions, in which the level of ambiguity between stimulus items and their contexts was manipulated by varying the probability with which a stimulus (word) and a context (background) appeared in relation to one another [20–23]. Performance of patients with lesions to the frontal cortex was compared with that of patients with temporal lobe lesions that had involved the hippocampus and parahippocampal cortex, as well as healthy control subjects. It was predicted that patients with lesions invading the ventrolateral prefrontal region would perform normally in context retrieval if stimuli and their contexts were stably associated with each other, but that these patients would be impaired if stimuli were linked to multiple contexts and, thus, requiring top-down control to retrieve the relevant stimulus-to-context links.
2. Material and methods
(a). Subjects
(i). Patients
Forty-three patients with circumscribed brain lesions were included in the study and divided into two groups: 23 patients with lesions to the frontal cortex and 20 patients with lesions in the temporal lobe. Patients were tested from six months to 26 years and two months after the operation or incident, with an average of 4.00 years (s.d. = 4.29) for the patients with frontal lesions and 6.89 years (s.d. = 7.85) for the patients with temporal lesions. The frontal and temporal groups did not differ in terms of time elapsed since surgery (t41 = 1.523, p = 0.14). None of the patients had comorbid neurological or psychiatric disorders.
(ii). Frontal group
Patients included in the frontal group had damage restricted to the frontal cortex and no more than the immediately subjacent white matter. It consisted of 12 patients with lesions in the left hemisphere, 10 in the right hemisphere and one with a small bilateral frontal cortical excision of a tumour in the supplementary motor area (SMA). Of the 12 patients with left-sided frontal lesions, 10 had undergone neurosurgery for the resection of a tumour and one for the removal of epileptogenic tissue, and one patient had sustained a stroke. All surgical removals spared the precentral motor cortex, except for one patient (Patient F005). The patient with a stroke (Patient F024) sustained damage in Broca's region in the left hemisphere but was free of aphasic symptoms at the time of testing.
Seven of the 10 patients in the right frontal group had undergone resection of a cerebral tumour and two of epileptogenic tissue; one patient had a stroke. The left- and right-sided frontal lesions are shown in electronic supplementary material figures S1 and S2, respectively. The anatomical data were not available for four patients (F015, F022, F023 and F029), but we had confirmation from the neurosurgeon that the lesions were restricted to the frontal cortex. Patient F023 had a left-sided lesion and patients F015, F022 and F029 had right-sided lesions. The operation report for patient F015 specifies that she underwent a corticectomy of the mid-SMA on the medial aspect of the superior frontal gyrus, extending 4 cm in the rostral–caudal axis and 2.5 cm in the dorsal–ventral axis.
(iii). Temporal group
This group comprised 12 patients with left-sided lesions and eight with right-sided lesions. The left temporal group included three patients who had surgery for the relief of epilepsy, eight who had tumour resection and one who had a stroke. The three surgical removals of epileptogenic tissue consisted of either a selective amygdalo-hippocampectomy (n = 2) in which these two structures are resected with the surrounding cortex, or an anterior temporal lobectomy (n = 1) that also included the amygdala and the anterior part of the hippocampus. Of the eight tumour resections, two were standard anterior temporal lobectomies. Three tumour patients underwent selective amygdalo-hippocampectomy, with the additional excision of the middle temporal gyrus in one case. One tumour resection involved the posterior third of the inferior temporal gyrus, with slight extension on the middle temporal gyrus and the white matter underlying the cortical excision. The anatomical data for the remaining two temporal tumour excisions and for the only patient with a stroke were not available, but there was confirmation from the neurosurgeon or neurologist that the lesions were restricted to the temporal lobe.
The right temporal group included five patients with removal of epileptogenic tissue (four selective amygdalo-hippocampectomies and one anterior temporal lobectomy). There were three patients with tumour removal whose anatomical data were not available.
(iv). Healthy control participants
Twenty-three healthy control subjects were also tested. They were neurologically intact individuals with no history of traumatic brain injury or any neurological or psychiatric disorder. They were matched as closely as possible with the two patient groups for age and education.
Only participants (patients and healthy subjects) with a full-scale Wechsler IQ score above 79 were included in the study. There was no significant difference between the three groups for mean age (F2,63 = 0.342, p = 0.71), years of education (F2,63 = 0.284, p = 0.75) and IQ (F2,61 = 1.200, p = 0.31). All patients and participants were right-handed, with the exception of two left-handed patients with frontal lesions (Patients F015 and F018), one ambidextrous patient with temporal lesion (Patient T003) and two left-handed control participants. Characteristics of the participant groups are presented in table 1.
Table 1.
Characteristics of participant groups.
| gender |
||||||
|---|---|---|---|---|---|---|
| group | M | F | age mean (s.d.) | education mean (s.d.) | time since surgery (years) mean (s.d.) | Wechsler IQ mean (s.d.) |
| frontal | 7 | 16 | 48.83 (9.47) | 15.48 (3.33) | 4.00 (4.29) | 110.41 (13.76) |
| left | 5 | 7 | 46.17 (10.00) | 15.75 (3.44) | 3.20 (3.59) | 112.08 (11.20) |
| right | 2 | 8 | 51.50 (8.76) | 14.80 (3.26) | 4.17 (4.63) | 107.00 (17.12) |
| bilateral | 0 | 1 | 54 (–) | 19 (–) | 11.92 (–) | 121 (–) |
| temporal | 13 | 7 | 46.75 (11.34) | 14.85 (2.37) | 6.89 (7.85) | 107.00 (11.55) |
| left | 8 | 4 | 44.42 (11.70) | 15.5 (2.24) | 5.69 (6.36) | 107.73 (13.52) |
| right | 5 | 3 | 50.25 (10.50) | 13.88 (2.36) | 8.68 (9.88) | 106.00 (8.93) |
| control | 8 | 15 | 49.35 (11.44) | 15.39 (2.92) | — | 113.04 (12.22) |
(b). Experimental design
The logic of the experimental design was as follows. Individual words (i.e. items) were to be presented on particular abstract coloured rectangles (i.e. contexts). Participants were required to recall the words in their contexts. There were three retrieval conditions. In the control recognition memory condition, a series of these words-in-contexts were to be presented and, later, during memory testing, the subjects would be required to recognize these words in their contexts (targets) when paired with new words in new contexts (distracters).
This basic control recognition memory condition provides the background against which to assess specific memory retrieval of the context in which words had been experienced. It was expected that lateral frontal lesions, unlike medial temporal lobe lesions, would not impair basic recognition memory. Two context memory retrieval conditions were designed in order to test the specific hypothesis that the ventrolateral prefrontal region is necessary to retrieve the context of items if the items and their contexts are not strongly associated with each (e.g. when items have been experienced under multiple contexts with equal frequency), but it is not necessary if the items and their contexts are strongly associated with each other (e.g. unique item-to-context relations). In the stable context retrieval condition, subjects experienced words in unique contexts (as in the control condition), but in the unstable context retrieval condition words were experienced under multiple backgrounds with equal frequency and thus there were no strong item-to-context associations to support retrieval of the context of a word from memory.
Since the basic recognition memory control condition would be expected to be the easiest one, pilot research was carried out with normal subjects to increase its difficulty to levels close to those of the two context retrieval conditions by increasing the delay between the presentation of the stimuli and memory testing. Based on this pilot research, a delay of approximately 7 min in the recognition memory condition was determined to lead to a level of performance close to that of the other two retrieval conditions in which the delays were within 2–4.5 s. Thus, the difficulty of the control recognition memory condition emanated primarily from the number of stimuli and the length of the delay between the experience of the stimuli and the memory testing, while that of the context retrieval conditions emanated from the retrieval of the specific context of an event after very short delays.
(c). Experimental material
The stimuli were words appearing on particular coloured abstract backgrounds, here referred to as the contexts (figure 1). A total of 147 words each one appearing within one of 147 backgrounds were used in this experiment. The words were all emotionally neutral nouns balanced for frequency and imageability. The mean word frequency count [33] was 44.59 (s.d.: 47.60) for the recognition memory condition, 31.05 (s.d.: 33.77) for the stable context retrieval condition and 42.83 (s.d.: 56.64) for the unstable context retrieval condition (F2,144 = 1.143, p = 0.32). The mean imageability rating [34] was 586.17 (s.d.: 14.92) for the recognition memory condition, 592.95 (s.d.: 26.40) for the stable context retrieval condition and 586.00 (s.d.: 27.63) for the unstable context retrieval condition (F2,144 = 0.780, p = 0.46). The words were written in black in 72-point Arial font on a white background and each word was placed in the centre of a coloured abstract 15 × 10 cm rectangular context (figure 1). The rectangular contexts were abstract non-verbal coloured designs created for the purpose of this experiment using Adobe Illustrator and Photoshop. It has been demonstrated that semantic similarity, even between pictures and words [35], exerts an interfering influence on retrieval, as concepts compete with one another [36]. Because we wanted no pre-existing relationships (i.e. no potential conceptual interference) between words and backgrounds in order to manipulate experimentally the level of ambiguity between them, neutral abstract backgrounds were used. These word-in-context stimuli were presented on a laptop computer screen with E-prime (Psychology Software Tools, Inc.), a specialized psychology program for stimulus presentation and data collection.
Figure 1.

(a) Schematic diagram of the testing procedure of the recognition memory condition, (b) the stable context retrieval condition and (c) unstable context retrieval condition. Note that the size of the screen for the encoding and retrieval phases was exactly the same in the experiment. However, in order to make the words legible in the illustration, the screen is enlarged and the question is removed in the retrieval phase. ISI, interstimulus interval.
(d). Procedure
Testing on the experiment was preceded by a practice session during which instructions were presented on the screen and read to the participants, with examples of the stimuli. The subjects completed a few practice trials before the beginning of the testing session in order to familiarize themselves with the task and to make sure they understood and could perform it properly.
Each condition comprised different trials all of which included an encoding phase, a delay and a retrieval testing phase (figure 1). The experimental session started with the encoding phase of the recognition memory condition. For the recognition memory condition, there were two testing parts, each one consisting of the presentation of 24 word-in-context stimuli. Thus, a total of 48 stimuli were presented for encoding and 48 pairs of stimuli (target and distracter) for retrieval. In each part, 24 word-in-context stimuli were presented in a random order for 2000 ms each with an interstimulus interval of 1000 ms. Patients were instructed to memorize these stimuli. When all 24 word-in-context stimuli had been presented, a delay of approx. 7 min (mean: 7 min 34 s, s.d.: 47 s) was interposed between the presentation of the stimuli (encoding phase) and the presentation of the testing stimuli (retrieval testing phase). During this long delay, half of the trials from the stable and half of the trials from the unstable context retrieval conditions were administered in a counterbalanced order (see below). The second part of the recognition memory condition was then administered with the other half of unstable and stable context retrieval conditions administered during the approximate 7 min delay of the recognition memory condition.
At the end of the approximate 7 min delay of each part of the recognition memory condition, participants saw pairs of word-in-context stimuli on the screen, one on the left and one on the right (figure 1a). One of the stimuli had been presented in the encoding phase (target) and the other was a new word in a new context, a stimulus that the participant never saw before (distracter). Thus, there were 24 such pairs, one for each one of the 24 stimuli presented during the encoding phase. The left/right position of the target and distracter stimuli was randomly determined but in a balanced manner so that the target and the distracter appeared an equal number of times on the left and right sides. The following question was presented at the top of the screen: ‘Which of these two words did you see previously?’ Participants were instructed to select the stimulus they had seen during the encoding phase in a forced-choice paradigm by pressing on the appropriate key (‘1’ for the stimulus on the left or ‘0’ for the stimulus on the right) on the laptop computer keyboard. There was no limit on the time to respond.
In the stable context retrieval condition, there were 12 trials, each one consisting of the presentation of four word-in-context stimuli during the encoding phase and of four pairs of testing stimuli during the retrieval phase, for a total of 48 encoding stimuli and 48 pairs of testing stimuli (figure 1b). During the encoding phase of each trial, four word-in-context stimuli were presented one at a time, each one for 3000 ms with an interstimulus interval of 1000 ms. In this condition, participants were instructed to remember the association between each word and the context in which it had appeared. After a delay of 4500 ms, four pairs of testing stimuli were administered, one for each of the four stimuli just presented. The target was one combination of word and context presented during the encoding phase and the distracter was the same target word, but presented on a context associated with another word during the encoding phase. Thus, all four contexts served as distracter for a word in the same trial (figure 1b). Two stimuli were presented below the question (‘On which background was this word presented?’) and participants had to select the target context.
In the unstable context retrieval condition, only three words and three contexts were used and the word-in-context stimuli presented during the different trials were the nine possible combinations of these three words and three coloured abstract contexts (figure 1c). In the encoding phase of each trial, each one of the three words was presented once and in one of the three contexts, i.e. three of the nine possible combinations of the three words and three contexts were presented. Each stimulus was presented for 3500 ms with an interstimulus interval of 1000 ms. A delay of 2000 ms was interposed between the encoding phase and the retrieval phase which consisted of the presentation of three pairs of testing stimuli. Each pair consisted of one of the three word-in-context stimuli presented during encoding (target) together with the same target word but on one of the other two contexts (distracter). Recall that in this condition only three words and three coloured abstract contexts were used and the words and contexts were combined randomly but equiprobably across the whole experiment. During the retrieval phase, the following question was presented: ‘The last time you saw this word, on which background was it presented?’ with two stimulus-complexes below the question. Sixteen trials were administered, each trial presenting three word-in-context stimuli during encoding and three testing pairs, for a total of 48 stimulus-complexes for encoding and 48 pairs of stimulus-complexes during memory testing.
The experimenter ensured the participants understood what they had to memorize during the encoding phase (i.e. the word on the particular context background) and on what basis they had to respond during the testing phase (i.e. the targets were the words presented on the correct backgrounds for that particular trial) in the stable and unstable context retrieval conditions.
3. Results
A three group (frontal, temporal and healthy controls) by three retrieval conditions (unstable context retrieval, stable context retrieval and recognition memory) repeated measures ANOVA was carried out to examine group differences on memory retrieval performance. Figure 2 shows the mean retrieval performance in per cent correct responses for each group across the three retrieval conditions. The ANOVA yielded a significant interaction between participant group and retrieval condition (F4,126 = 3.15, p = 0.02), with Greenhouse–Geisser correction. Analyses of simple main effects of participant group were carried out within each retrieval condition, followed by Fisher least significant difference tests in order to make specific group comparisons. In the recognition memory condition, the patients with temporal lesions performed significantly worse than healthy controls (p = 0.03) and patients with frontal lesions (p = 0.03). There was no difference in performance between the frontal group and the normal control subjects (p = 0.96) in this condition. It is important to note that only in this recognition memory condition was the delay between the encoding phase and the memory test longer than a few seconds, i.e. approximately 7 min. Thus, the temporal lesions created sensitivity to delay, but the frontal lesions did not. In the stable context retrieval condition, the temporal group was significantly impaired in comparison with the control group (p = 0.03), and the frontal group was marginally impaired compared with the control group (p = 0.06). In the unstable context retrieval condition, patients with frontal cortical lesions performed significantly worse than control subjects (p = 0.03), whereas patients with temporal lesions did not differ from controls (p = 0.35).
Figure 2.
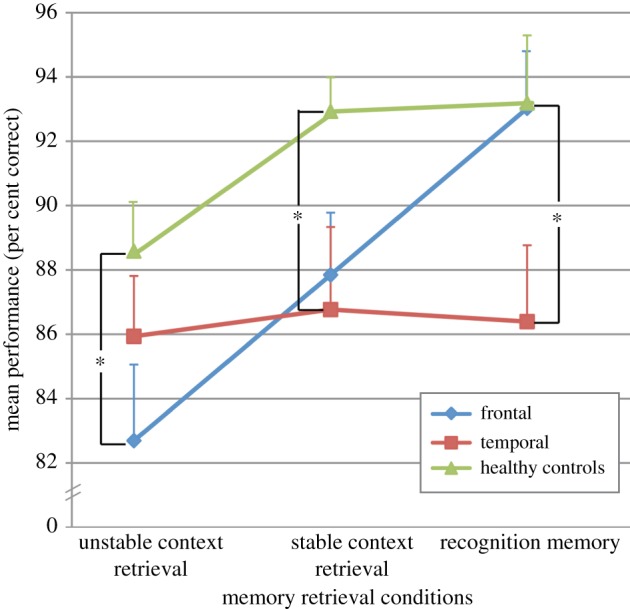
Graph showing the mean performance of each group across the three memory retrieval conditions. Error bars represent the standard error. *p < 0.05.
Note that the performance of the patients with temporal lesions was comparable across the three retrieval conditions and the absence of significant impairment in the temporal group on the unstable retrieval condition may have been due to the lower performance of the control subjects on this condition, thus decreasing the difference between the control and temporal groups, rather than to an absence of impairment per se in the temporal group. However, we must also note that only three stimuli were shown during the presentation phase in the unstable condition and the memory testing took place only 2 s later. Patients with temporal lesions perform well on short-term memory tasks that do not exceed their normal span [1–3]. Even the well-known patient H.M., who had a severe anterograde amnesia after bilateral excision of the limbic region of the medial temporal lobe, had no deficit on short-term memory with very short delays [1].
In the temporal group, the effect of laterality of the lesions was also examined by comparing patients with left- and right-sided temporal lesions within each retrieval condition. Simple main effect of lesion side yielded no significant difference for the stable context retrieval (F1,18 = 0.01, p = 0.92) and the recognition memory (F1,18 = 0.08, p = 0.78) conditions. In the unstable context retrieval condition, there was a trend for the patients with right temporal lesions to perform worse than patients with left temporal lesions (F1,18 = 3.24, p = 0.09).
In the preliminary analysis reported earlier, the patients with lesions anywhere in the frontal cortex were treated as a single group so as to examine overall differences between the effects of frontal and temporal lesions. However, the experiment was designed to test the hypothesis that the VLPFC is critical for the disambiguation of mnemonic traces when the relations between stimuli and their contexts are unstable [20–23]. Patients with lesions to the VLPFC were predicted to be selectively impaired on the unstable context retrieval condition which assesses active controlled retrieval. In addition, we had previously shown that lesions invading the left dorsomedial region of the frontal cortex (DMFC) yield impairment in verbal fluency [32]. One fundamental requirement in verbal fluency tasks is the retrieval from verbal long-term memory of words that meet certain requirements. Thus, the verbal fluency deficit after left DMFC lesions may be reflecting a more general retrieval impairment.
In a second series of analyses, we examined the above predictions within the frontal group. Patients with frontal cortical lesions were divided into the following subgroups: (i) patients with lesions invading only the left VLPFC (left VLPFC; n = 3); (ii) patients with lesions invading only the right VLPFC (right VLPFC; n = 3); (iii) patients with only left DMFC lesions (left DMFC; n = 4); (iv) patients with damage to the frontal cortex that did not include either the VLPFC or the left DMFC (other FC; n = 8). Patients included in the other FC subgroup had lesions that did not invade the mid-VLPFC, i.e. cytoarchitectonic areas 45 and 47/12 as defined by Petrides & Pandya [37]. In addition, these lesions had to spare the dorsomedial frontal cortex (DMFC) that in Chapados & Petrides [32] reduced verbal fluency: the DMFC anterior to the SMA, including the pre-SMA, the cingulate motor areas and medial areas 8, 9 and 32. Note that no patient had a lesion to the right dorsomedial frontal region analogous to the one found to be critical for verbal fluency. In addition, two patients with lesions that invaded both the left VLPFC and the left DMFC could not be assigned to either group. Their performance is discussed individually later. The three patients with frontal lesions whose anatomical data were not available were not included in this analysis. The mean scores on each retrieval condition for the four frontal sub-groups and the control group are presented in figure 3.
Figure 3.
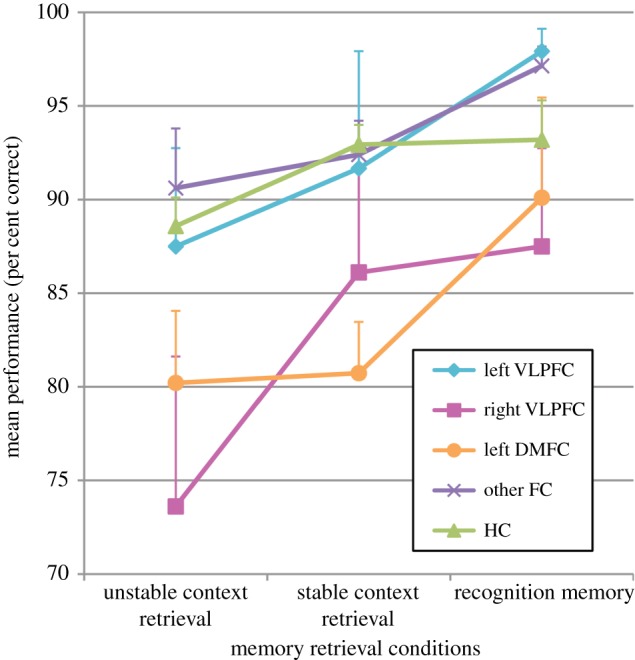
Graph showing the mean performance on the three memory retrieval conditions of the four frontal groups: patients with lesions (i) including the left ventrolateral prefrontal cortex (left VLPFC), (ii) including the right VLPFC, (iii) including the left dorsomedial frontal cortex (left DMFC), (iv) sparing both the VLPFC and left DMFC (other FC) and of normal control subjects (HC). Error bars represent the standard error.
To test the specific predictions about the individual frontal sub-groups, one-way ANOVAs were conducted to compare the five groups (left VLPFC, right VLPFC, left DMFC, other FC and healthy control) for each one of the three retrieval conditions, followed by Dunnett tests in which each frontal sub-group was compared with the healthy control group. Significant differences were found in the unstable (F4,36 = 3.23, p = 0.02) and stable (F4,36 = 4.31, p = 0.006) context retrieval conditions, but not in the recognition memory condition (F4,36 = 1.03, p = 0.41). Only the right ventrolateral prefrontal group was impaired on the unstable context retrieval condition in comparison with the control subjects (p = 0.02, two-tailed). The left VLPFC and the left DMFC groups were not significantly impaired relative to the control group in this condition. By contrast, on the stable context retrieval condition, only patients with left DMFC lesions were impaired in comparison with the healthy control subjects (p = 0.04, two-tailed). The performance of patients with frontal lesions sparing both the VLPFC and the left DMFC (other FC) was not significantly different from that of normal subjects on all three memory conditions (figure 3).
Thus, the essential finding was that lesions limited to the right VLPFC impaired performance only on the unstable context retrieval condition, as predicted. The left DMFC lesions clearly impaired performance on the stable context retrieval condition.
Finally, the scores of the two patients whose prefrontal lesions included both the left VLPFC and DMFC were low on the unstable context retrieval condition (68.75 and 72.92). On the stable context retrieval condition, one patient's performance (81.25) was similar to that of the left DMFC only group, but the other patient (93.75) performed like the normal control group. These double-lesion patients strengthen the argument that the left DMFC and perhaps also the left VLPFC play a role in the contextual retrieval of information.
4. Discussion
In this experiment, patients with frontal and temporal lesions were tested on three memory conditions that required retrieval of items and their contexts. In the control condition, subjects could perform well on the basis of mnemonic recognition of the previously experienced word-in-context stimuli. In both the stable and unstable context retrieval conditions, however, subjects had to retrieve explicitly the context associated with the words. The difference between the latter two conditions was the fact that, in the stable context condition, each word had been seen in only one context (unique and stable relations between items and contexts), while in the unstable context condition the words had been seen in all the contexts across trials (multiple relations of items and contexts). The first major finding of this study was a clear dissociation between the effect of lesions to the frontal cortex and the temporal lobe on basic recognition memory retrieval. In the recognition memory condition, the patients with temporal lesions performed significantly worse than the healthy controls, but the patients with frontal lesions performed as well as the control subjects. The difficulty in the recognition memory task emanates from the relatively large number of stimuli presented (24 stimuli versus four and three in the two context retrieval conditions) and the relatively long delay between stimulus presentation and memory testing (7 min versus a few seconds in the context retrieval conditions). These results are consistent with the well-established fact that the hippocampus and related structures in the medial temporal lobe are essential for item memory [1–5,38,39]. The normal performance of patients with frontal cortical lesions on the recognition memory condition confirms previous research that patients and monkeys with lesions of the lateral frontal cortex are not impaired on basic recognition memory [40–44].
In sharp contrast to their normal recognition memory with a relatively long list of stimuli (24 stimuli) and long delay (approx. 7 min), patients with lesions that invaded the right ventrolateral prefrontal region were impaired selectively on the unstable context retrieval condition despite the low number of stimuli presented during the encoding phase (three stimuli) and the very short delays between stimulus presentation and memory testing (2 s). Here it is important to note that patients with frontal cortical lesions that spared the ventrolateral prefrontal region and the left dorsomedial verbal fluency retrieval region [32] were not impaired on either of the two context retrieval conditions, emphasizing the regional specificity of frontal cortical impairments. Thus, consistent with the hypothesis tested in the present experiment, the patients who had lesions that included the right VLPFC were impaired on the context memory retrieval task in which stimulus-to-context relations were unstable, but were not impaired when these relations were stable (figure 3).
The lack of impairment in the left ventrolateral prefrontal group may have been due to the fact that the lesion of one patient in this group was not complete since area 47/12 was clearly spared (see Patient F024 in electronic supplementary material, figure S1). Alternatively, there may be a difference between the right and left ventrolateral prefrontal regions that remains to be explored further. For instance, the three words in the unstable context retrieval condition were not entering into variable relations with other words or semantic contexts, but rather with abstract visual backgrounds and this may have been the major source of ambiguity in the unstable context retrieval condition. Thus, the abstract visual contexts may have created a source of ambiguity that was demanding more processing from the right hemisphere as far as context disambiguation was concerned and, of course, the right hemisphere is dominant for the processing of abstract stimulus material [44]. This argument receives some support from the fact that patients with right temporal lesions tended to perform slightly worse than those with left temporal lesions on this condition.
The present results provide the first cause-and-effect confirmation of the essential role of the VLPFC in active retrieval processing necessary to disambiguate items and their context in memory, which had previously received support from neuroimaging studies on normal human subjects [15,20–23] and single neuron recording in monkeys [45]. Petrides [15,16] argued that the ventrolateral prefrontal cortical region (areas 45 and 47/12) may be a critical part of the prefrontal cortex for the active controlled retrieval of information in situations in which items of information exist in memory under multiple associations with one another and, therefore, top-down control processing is necessary to disambiguate the memory traces that are assumed to lie in the posterior association neocortex. It is important to note that the normal performance of the patients with ventrolateral prefrontal lesions on the stable context condition demonstrates that these patients can retrieve the context of a word recently experienced and can easily separate this context from other recently experienced contexts that were associated with other recently experienced words. Thus, the impairment on the unstable context condition stems not from a difficulty in discriminating between recent events (recency impairment), but rather from the need to retrieve the specific context under which a stimulus had been experienced on a particular trial when that stimulus had previously been experienced under multiple contexts.
The experimental design of this study permits a demonstration of the critical role of the right VLPFC in the disambiguation of item-to-context relations in memory when there are no strong unambiguous relations between items and their contexts to enable bottom-up retrieval. The finding that the ventrolateral prefrontal region is critical for selective retrieval under conditions of high selection demands is consistent with functional neuroimaging evidence of the involvement of the left ventrolateral prefrontal region in demanding verbal recall, such as the free recall of words that appeared on particular lists [27] and verbal fluency [28,29]. It is also consistent with demonstrations that the left ventrolateral prefrontal region plays a key role in the control of semantic retrieval [24–26], especially when a given stimulus activates the retrieval of many competing verbal response options.
The design of the present experiment provides some insight into the reason why demanding retrieval may require the ventrolateral prefrontal region. Theoretically, in the unstable context retrieval task, upon the presentation of each word on the test trials, any one of the three competing contexts has an equal chance of being retrieved. Successful retrieval of the required context on a particular trial depends on active disambiguation processing, namely enhancement of the appropriate context and suppression of the other contexts under which the word had also appeared, but on different trials. Single-unit recording in the macaque VLPFC, i.e. cytoarchitectonic areas 45 and 47/12, provided evidence of neuronal activity that can underlie this disambiguation process which is the basis of active controlled retrieval. After an instruction to retrieve a specific aspect of a recently memorized complex stimulus (e.g. to retrieve its colour, but not its shape), a class of neurons in the VLPFC responds selectively to the isolation of the instructed aspect of the memorized stimulus (see [45] for details). Thus, there is evidence of neuronal processing isolating particular aspects of memorized experiences.
Unlike the patients with right ventrolateral prefrontal lesions, the patients with left dorsomedial frontal lesions were clearly impaired on the stable context retrieval condition, and to a lesser extent, on the unstable context retrieval condition. This group had similar performance on both the stable and unstable context retrieval conditions, suggesting a difficulty in retrieving the item-to-context relations, regardless of their stability.
A plausible explanation for the finding that the left DMFC is involved in context retrieval comes from the idea that the dorsomedial prefrontal region plays a role in sustained attention, cognitive effort and ‘energization’ [46]. Energization refers to the process of sustaining a response after initiation. According to this view, the dorsomedial prefrontal region is recruited when a task requires maintaining a specific response over time, or producing new responses that are not overlearned. Energization is considered to be domain-general and could, therefore, also be recruited for memory retrieval tasks. In this sense, context retrieval is less automatic and requires more cognitive effort than basic recognition of an episode or an item, because a given context must not only be recognized, but also associated with the correct item and dissociated from incorrect items. Energization could thus be needed in order to sustain sufficient activation to complete this type of task. If the contextual information needs to be disambiguated because of unstable context-to-item relationships, the VLPFC would then be additionally recruited. These two regions would play two distinct executive roles at two different levels in context retrieval.
An alternative interpretation may be that the left dorsomedial prefrontal region plays a role in context retrieval in a more direct manner. Poor performance following lesions in this region on both context retrieval conditions could be driven by the basic requirement to retrieve the context within which particular words were embedded. Thus, the present findings could provide evidence, for the first time, that this region which had previously been linked with verbal fluency [30–32] is also involved in verbal item-to-context memory retrieval. This is consistent with the idea that the phonological verbal fluency task examines the capacity to retrieve from memory items that satisfy specific criteria, such as words beginning with a certain letter, to select the appropriate words and to differentiate them from words that do not meet the criteria.
The critical region for verbal fluency found by Chapados & Petrides [32] in the DMFC included the supplementary speech zone, the cingulate motor region, the paracingulate cortex and the medial prefrontal areas 8 and 9. This dorsomedial frontal region, which is linked with the hippocampal/parahippocampal region [47], may play a central role in episodic memory retrieval. Temporal lesions impair basic memory (i.e. both item recognition and retrieval of context), reflecting the necessary role of the hippocampus and parahippocampal cortex in forming new memories, such as binding an item with its context [2–4,48,49]. By contrast, the impairment after left dorsomedial frontal lesions appears in the context of normal recognition memory as shown by the present findings. Thus, the limbic medial temporal structures are necessary to encode and maintain strong memory representations of the words and their contexts, while the left dorsomedial frontal region contributes to the retrieval of words in their context. For both the temporal and the left dorsomedial frontal lesions, ambiguity of stimulus-to-context relations and the need to disambiguate these relations during memory retrieval was not a critical factor influencing performance. This effect of ambiguity was clearly observed only in patients with right ventrolateral prefrontal lesions. We know that anatomically the VLPFC is linked with the dorsomedial prefrontal region in both the monkey [37] and the human brain [50], although the meaning of these connections had remained unclear until now. This dorsomedial frontal region was also co-activated with the ventrolateral prefrontal region in the functional neuroimaging studies of controlled active memory retrieval (e.g. [22], see fig. 2b and table 1; [23], see fig. 2b,c and table 1). Thus, the present results suggest that the left dorsomedial frontal region is involved in context memory retrieval and its bi-directional link with the ventrolateral prefrontal region permits engagement of the latter region when the traces in memory are embedded in multiple contexts and, therefore, the relations among stimuli become ambiguous. Based on the present results, one could hypothesize the existence of a circuit comprising the hippocampus and adjacent medial temporal cortex, the DMFC and the VLPFC, each playing distinct roles in stimulus–context retrieval in episodic memory.
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Acknowledgements
We thank Rhonda Amsel for statistical advice.
Data accessibility
The performance data of individual patients with frontal cortical lesions are provided next to their anatomical data in electronic supplementary material, figures S1 and S2. For those frontal patients for whom no reconstruction of their lesions was available, the scores are provided in the legend of the electronic supplementary material, figures S1 and S2. The performance of the different participant groups is presented in electronic supplementary material, tables S3 and S4.
Funding statement
This work was supported by grant no: RGPIN-2014-04729 from NSERC to M.P. and CIHR and FRSQ Graduate Student Fellowships to C.C.
References
- 1.Scoville WB, Milner B. 1957. Loss of recent memory after bilateral hippocampal lesions. J. Neurol. Neurosurg. Psychiatry 20, 11–21. ( 10.1136/jnnp.20.1.11) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Smith ML, Milner B. 1981. The role of the right hippocampus in the recall of spatial location. Neuropsychologia 19, 781–93. ( 10.1016/0028-3932(81)90090-7) [DOI] [PubMed] [Google Scholar]
- 3.Smith ML, Milner B. 1989. Right hippocampal impairment in the recall of spatial location: encoding deficit or rapid forgetting? Neuropsychologia 27, 71–81. ( 10.1016/0028-3932(89)90091-2) [DOI] [PubMed] [Google Scholar]
- 4.Parkinson JK, Murray EA, Mishkin M. 1988. A selective mnemonic role for the hippocampus in monkeys: memory for the location of objects. J. Neurosci. 8, 4159–4167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nadel L, Moscovitch M. 2001. The hippocampal complex and long-term memory revisited. Trends Cogn. Sci. 5, 228–230. ( 10.1016/S1364-6613(00)01664-8) [DOI] [PubMed] [Google Scholar]
- 6.Dobbins IG, Foley H, Schacter DL, Wagner AD. 2002. Executive control during episodic retrieval: multiple prefrontal processes subserve source memory. Neuron 35, 98–996. ( 10.1016/S0896-6273(02)00858-9) [DOI] [PubMed] [Google Scholar]
- 7.Fujii T, Suzuki M, Okuda J, Ohtake H, Tanji K, Yamaguchi K, Itoh M, Yamadori A. 2004. Neural correlates of context memory with real-world events. Neuroimage 21, 1596–1603. ( 10.1016/j.neuroimage.2004.01.005) [DOI] [PubMed] [Google Scholar]
- 8.Mitchell KJ, Johnson MK. 2009. Source monitoring 15 years later: what have we learned from fMRI about the neural mechanisms of source memory? Psychol. Bull. 135, 638–677. ( 10.1037/a0015849) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shimamura AP, Squire LR. 1987. A neuropsychological study of fact memory and source amnesia. J. Exp. Psychol. Learn. Mem. Cogn. 13, 464–473. ( 10.1037/0278-7393.13.3.464) [DOI] [PubMed] [Google Scholar]
- 10.Janowsky JS, Shimamura AP, Squire LR. 1989. Source memory impairment in patients with frontal lobe lesions. Neuropsychologia 27, 1043–1056. ( 10.1016/0028-3932(89)90184-X) [DOI] [PubMed] [Google Scholar]
- 11.Shimamura AP, Janowsky JS, Squire LR. 1990. Memory for the temporal order of events in patients with frontal lobe lesions and amnesic patients. Neuropsychologia 28, 803–813. ( 10.1016/0028-3932(90)90004-8) [DOI] [PubMed] [Google Scholar]
- 12.Duarte A, Ranganath C, Knight RT. 2005. Effects of unilateral prefrontal lesions on familiarity, recollection, and source memory. J. Neurosci. 25, 8333–8337. ( 10.1523/JNEUROSCI.1392-05.2005) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Thaiss L, Petrides M. 2003. Source versus content memory in patients with a unilateral frontal cortex or a temporal lobe excision. Brain 126, 1112–1126. ( 10.1093/brain/awg112) [DOI] [PubMed] [Google Scholar]
- 14.Thaiss L, Petrides M. 2008. Autobiographical memory of the recent past following frontal cortex or temporal lobe excisions. Eur. J. Neurosci. 28, 829–840. ( 10.1111/j.1460-9568.2008.06381.x) [DOI] [PubMed] [Google Scholar]
- 15.Petrides M. 1996. Specialized systems for the processing of mnemonic information within the primate frontal cortex. Phil. Trans. R. Soc. Lond. B 351, 1455–1462. ( 10.1098/rstb.1996.0130) [DOI] [PubMed] [Google Scholar]
- 16.Petrides M. 2002. The mid-ventrolateral prefrontal cortex and active memory retrieval. Neurobiol. Learn. Mem. 78, 528–538. ( 10.1006/nlme.2002.4107) [DOI] [PubMed] [Google Scholar]
- 17.Moscovitch M. 1992. Memory and working-with-memory: a component process model based on modules and central systems. J. Cogn. Neurosci. 4, 257–267. ( 10.1162/jocn.1992.4.3.257) [DOI] [PubMed] [Google Scholar]
- 18.Stuss DT, Alexander MP. 2005. Does damage to the frontal lobes produce impairment in memory? Curr. Direct Psychol. Sci. 14, 84–88. ( 10.1111/j.0963-7214.2005.00340.x) [DOI] [Google Scholar]
- 19.Badre D, Wagner AD. 2007. Left ventrolateral prefrontal cortex and the cognitive control of memory. Neuropsychologia 45, 2883–2901. ( 10.1016/j.neuropsychologia.2007.06.015) [DOI] [PubMed] [Google Scholar]
- 20.Cadoret G, Pike GB, Petrides M. 2001. Selective activation of the ventrolateral prefrontal cortex in the human brain during active mnemonic processing. Eur. J. Neurosci. 14, 1164–1170. ( 10.1046/j.0953-816x.2001.01737.x) [DOI] [PubMed] [Google Scholar]
- 21.Kostopoulos P, Albanese MC, Petrides M. 2007. Ventrolateral prefrontal cortex and tactile memory disambiguation in the human brain. Proc. Natl Acad. Sci. USA 104, 10 223–10 228. ( 10.1073/pnas.0700253104) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kostopoulos P, Petrides M. 2003. The mid-ventrolateral prefrontal cortex: insights into its role in memory retrieval. Eur. J. Neurosci. 17, 1489–1497. ( 10.1046/j.1460-9568.2003.02574.x) [DOI] [PubMed] [Google Scholar]
- 23.Kostopoulos P, Petrides M. 2008. Left mid-ventrolateral prefrontal cortex: underlying principles of function. Eur. J. Neurosci. 27, 1037–1049. ( 10.1111/j.1460-9568.2008.06066.x) [DOI] [PubMed] [Google Scholar]
- 24.Thompson-Schill SL, D'Esposito M, Aguirre JK, Farah MJ. 1997. Role of the left inferior prefrontal cortex in retrieval of semantic knowledge: a reevaluation. Proc. Natl Acad. Sci. USA 94, 14 792–14 797. ( 10.1073/pnas.94.26.14792) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Robinson G, Blair J, Cipolotti L. 1998. Dynamic aphasia: an inability to select between competing verbal responses? Brain 121, 77–89. ( 10.1093/brain/121.1.77) [DOI] [PubMed] [Google Scholar]
- 26.Jefferies E, Lambon Ralph MA. 2006. Semantic impairment in stroke aphasia versus semantic dementia: a case-series comparison. Brain 129, 2132–2147. ( 10.1093/brain/awl153) [DOI] [PubMed] [Google Scholar]
- 27.Petrides M, Alivisatos B, Evans AC. 1995. Functional activation of the human ventrolateral frontal cortex during mnemonic retrieval of verbal information. Proc. Natl Acad. Sci. USA 92, 5803–5807. ( 10.1073/pnas.92.13.5803) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Phelps E, Hyder F, Blamire AM, Shulman RG. 1997. FMRI of the prefrontal cortex during overt verbal fluency. NeuroReport 8, 561–565. ( 10.1097/00001756-199701200-00036) [DOI] [PubMed] [Google Scholar]
- 29.Amunts K, et al. 2004. Analysis of neural mechanisms underlying verbal fluency in cytoarchitectonically defined stereotaxic space—the roles of Brodmann areas 44 and 45. Neuroimage 22, 42–56. ( 10.1016/j.neuroimage.2003.12.031) [DOI] [PubMed] [Google Scholar]
- 30.Robinson G, Shallice T, Bozzali M, Cipolotti L. 2012. The differing roles of the frontal cortex in fluency tests. Brain 135, 2202–2214. ( 10.1093/brain/aws142) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Stuss DT, Alexander MP, Hamer L, Palumbo C, Dempster R, Binns M, Levine B, Izukawa D. 1998. The effects of focal anterior and posterior brain lesions on verbal fluency. J. Int. Neuropsychol. Soc. 4, 265–278. [PubMed] [Google Scholar]
- 32.Chapados C, Petrides M. 2013. Impairments only on the verbal fluency subtest of the Frontal Assessment Battery after prefrontal lesions. Brain 136, 2966–2978. ( 10.1093/brain/awt228) [DOI] [PubMed] [Google Scholar]
- 33.Kucera H, Francis WN. 1967. Computational analysis of present-day American English. Providence, RI: Brown University Press. [Google Scholar]
- 34.Pavio A, Yuille JC, Madigan SA. 1968. Concreteness, imagery and meaningfulness values for 925 words. J. Exp. Psychol. 76(Monograph Suppl), 125. [DOI] [PubMed] [Google Scholar]
- 35.Lupker SJ, Katz AN. 1981. Input, decision, and response factors in picture–word interference. J. Exp. Psychol. Hum. Learn. Mem. 7, 269–282. ( 10.1037/0278-7393.7.4.269) [DOI] [Google Scholar]
- 36.Damian MF, Vigliocco G, Levelt WJM. 2001. Effects of semantic context in the naming of pictures and words. Cognition 81, B77–B86. ( 10.1016/S0010-0277(01)00135-4) [DOI] [PubMed] [Google Scholar]
- 37.Petrides M, Pandya DN. 2002. Comparative cytoarchitectonic analysis of the human and the macaque ventrolateral prefrontal cortex and corticocortical connection patterns in the monkey. Eur. J. Neurosci. 16, 291–310. ( 10.1046/j.1460-9568.2001.02090.x) [DOI] [PubMed] [Google Scholar]
- 38.Squire LR, Zola-Morgan S. 1991. The medial temporal lobe memory system. Science 253, 1380–1386. ( 10.1126/science.1896849) [DOI] [PubMed] [Google Scholar]
- 39.Bachevalier J, Nemanic S. 2008. Memory for spatial location and object–place associations are differently processed by the hippocampal formation, parahippocampal areas TH/TF and perirhinal cortex. Hippocampus 18, 64–80. ( 10.1002/hipo.20369) [DOI] [PubMed] [Google Scholar]
- 40.Bachevalier J, Mishkin M. 1986. Visual recognition impairment follows ventromedial but not dorsolateral prefrontal lesions in monkeys. Behav. Brain Res. 20, 249–261. ( 10.1016/0166-4328(86)90225-1) [DOI] [PubMed] [Google Scholar]
- 41.Petrides M. 1995. Impairments on nonspatial self-ordered and externally ordered working memory tasks after lesions of the mid-dorsal part of the lateral frontal cortex in the monkey. J. Neurosci. 15, 359–375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Petrides M. 2000. Frontal lobes and memory. In Handbook of neuropsychology, vol. 2 (eds Boller F, Grafman J.), pp. 67–84, 2nd edn Amsterdam, The Netherlands: Elsevier. [Google Scholar]
- 43.Petrides M. 2000. Dissociable roles of mid-dorsolateral prefrontal and anterior inferotemporal cortex in visual working memory. J. Neurosci. 20, 7496–7503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Milner B. 1971. Interhemispheric differences in the localization of psychological processes in man. Br. Med. Bull. 26, 272–277. [DOI] [PubMed] [Google Scholar]
- 45.Cadoret G, Petrides M. 2007. Ventrolateral prefrontal neuronal activity related to active controlled memory retrieval in nonhuman primates. Cereb. Cortex 17, i27–i40. ( 10.1093/cercor/bhm086) [DOI] [PubMed] [Google Scholar]
- 46.Stuss DT, Alexander MP. 2007. Is there a dysexecutive syndrome? Phil. Trans. R. Soc. B 362, 901–915. ( 10.1098/rstb.2007.2096) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Morris R, Pandya DN, Petrides M. 1999. Fiber system linking the middorsolateral frontal cortex with the retrosplenial/presubicular region in the rhesus monkey. J. Comp. Neurol. 407, 183–192. () [DOI] [PubMed] [Google Scholar]
- 48.Malkova L, Mishkin M. 2003. One-trial memory for object–place associations after separate lesions of hippocampus and posterior parahippocampal region in the monkey . J. Neurosci. 23, 1956–1965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Park H, Shannon V, Biggan J, Spann C. 2012. Neural activity supporting the formation of associative versus source memory. Brain Res. 1471, 81–92. ( 10.1016/j.brainres.2012.07.012) [DOI] [PubMed] [Google Scholar]
- 50.Margulies DS, Petrides M. 2013. Distinct parietal and temporal connectivity profiles of ventrolateral frontal areas involved in language production. J. Neurosci. 33, 16 846–16 852. ( 10.1523/JNEUROSCI.2259-13.2013) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The performance data of individual patients with frontal cortical lesions are provided next to their anatomical data in electronic supplementary material, figures S1 and S2. For those frontal patients for whom no reconstruction of their lesions was available, the scores are provided in the legend of the electronic supplementary material, figures S1 and S2. The performance of the different participant groups is presented in electronic supplementary material, tables S3 and S4.