

Abstract
Across species, complex circuits inside the basal ganglia (BG) converge on pallidal output neurons that exhibit movement-locked firing patterns. Yet the origins of these firing patterns remain poorly understood. In songbirds during vocal babbling, BG output neurons homologous to those found in the primate internal pallidal segment are uniformly activated in the tens of milliseconds prior to syllable onsets. To test the origins of this remarkably homogenous BG output signal, we recorded from diverse upstream BG cell types during babbling. Prior to syllable onsets, at the same time that internal pallidal segment-like neurons were activated, putative medium spiny neurons, fast spiking and tonically active interneurons also exhibited transient rate increases. In contrast, pallidal neurons homologous to those found in primate external pallidal segment exhibited transient rate decreases. To test origins of these signals, we performed recordings following lesion of corticostriatal inputs from premotor nucleus HVC. HVC lesions largely abolished these syllable-locked signals. Altogether, these findings indicate a striking homogeneity of syllable timing signals in the songbird BG during babbling and are consistent with a role for the indirect and hyperdirect pathways in transforming cortical inputs into BG outputs during an exploratory behavior.
Keywords: songbird, basal ganglia, globus pallidus, striatum
the basal ganglia (bg) are an evolutionarily conserved set of brain nuclei critical for motor learning (Graybiel et al. 1994). BG output neurons in the internal pallidal segment (GPi) and the substantia nigra pars reticulata control behavior through their projections to the brain stem and the ventral “motor” thalamus (Albin et al. 1989; Penney and Young 1983). Across behavioral paradigms and model systems, the activities of BG output neurons are time-locked to behavior in diverse ways: neurons may exhibit rate increases or decreases in response to task cues or reward, as well as prior to, during, or following movement initiation or termination (Anderson and Horak 1985; Anderson and Turner 1991; Bryden et al. 2011; Freeze et al. 2013; Georgopoulos et al. 1983; Gulley et al. 1999; Joshua et al. 2009; Kunimatsu and Tanaka 2010; Meyer-Luehmann et al. 2002; Nambu et al. 1990; Schmidt et al. 2013; Sheth et al. 2011; Turner and Anderson 2005; 1997; Yoshida and Tanaka 2009).
The behavior-locked firing patterns of GPi neurons are controlled by multiple parallel circuits inside the BG (Fig. 1A) (Tachibana et al. 2008), including the hyperdirect pathway, as well as the direct and indirect pathways, which themselves can be modulated by striatal interneurons (Fig. 1A) (Gittis et al. 2010; Hikosaka and Wurtz 1989). Songbirds provide a unique opportunity to determine how these pathways shape behavior-locked firing of BG outputs. First, songbird area X is a striato-pallidal nucleus homologous to the mammalian BG (Fig. 1B) (Doupe et al. 2005; Farries et al. 2005; Farries and Perkel 2002; Goldberg et al. 2010; Goldberg and Fee 2010). Second, during vocal babbling, BG outputs discharge with a peculiar homogeneity that, to our knowledge, has not been observed in other model systems or behaviors: GPi-like output neurons in area X exhibit brief peaks in activity in the milliseconds prior to syllable onsets and brief rate decreases prior to offsets. Pre-onset decreases or pre-offset increases are never observed (Goldberg et al. 2012; Goldberg and Fee 2012). The striking homogeneity of these signals appears only during the babbling stage of vocal development. At later plastic and adult song stages, each GPi-like neuron exhibits its own idiosyncratic timing relationship to song, increasing or decreasing its firing rate at onsets or offsets or at reproducible time points in between (Goldberg et al. 2010; Hessler and Doupe 1999; Iwasaki et al. 2013; Woolley et al. 2014).
Fig. 1.

Basal ganglia (BG) circuitry in mammals and songbirds. A: three main pathways run in parallel in mammalian BG circuits and converge on BG output neurons in the globus pallidus, internal segment (GPi). First, in the direct pathway, medium spiny neurons (MSNs) directly inhibit BG outputs via the MSN → GPi projection. Second, in the indirect pathway, the external pallidal segment (GPe), an inhibitory nucleus, is interposed between the MSNs and the GPi, such that indirect pathway MSNs may disinhibit GPi neurons through the MSN → GPe → GPi projection. Third, in the hyperdirect pathway, cortical inputs can activate GPi neurons through the glutamatergic subthalamic nucleus (Nambu et al. 2002). Additionally, BG output neurons may directly suppress each other's firing through extensive collaterals (Brown et al. 2014). Finally, three classes of striatal interneuron, low-threshold spiking (LTS), fast spiking (FS) and tonically active neurons (TAN), modulate MSN firing, but their roles in shaping BG output signals remain unknown (English et al. 2012; Gittis and Kreitzer 2012; Szydlowski et al. 2013). B: similar circuits exist in area X nucleus of songbirds, but there are also important differences. First, no homolog of the mammalian subthalamic nucleus has yet to be reported inside area X. A hyperdirect pathway is manifest via inputs from HVC and lateral magnocellular nucleus of the anterior nidopallium (LMAN) directly to pallidal output neurons, but this pathway lacks recurrent subthalamic nucleus-GPe connections important in mammals. Second, it remains unknown if there are different area X MSN subtypes that give rise to direct and indirect pathways via distinct outputs to GPi- and GPe-like neurons, respectively.
To determine the origins of this “known” BG output signal, we recorded area X neurons during vocal babbling. We find that putative medium spiny neurons (MSNs) and fast spiking (FS) and tonically active (TAN) interneurons exhibit similar song-locked discharge as the GPi neurons: rate increases at onsets and decreases at offsets. Meanwhile, external pallidal segment (GPe)-like neurons exhibit rate decreases at onsets. These syllable locked signals were abolished by lesion of the premotor cortical nucleus HVC. Together, these findings are consistent with roles for the indirect and hyperdirect pathways in shaping BG output signals during early stages of vocal development.
MATERIALS AND METHODS
Animals.
Subjects were seven control juvenile male zebra finches, 35–50 days posthatch, and four HVC-lesioned zebra finches, 40–55 days posthatch. Birds were obtained from the Cornell zebra finch breeding facility (Ithaca, NY). The care and experimental manipulation of the animals were carried out in accordance with guidelines of the National Institutes of Health and were reviewed and approved by the Cornell Committee on Animal Care. All data are from juvenile birds singing undirected song.
Chronic neural recordings and histology.
Recordings were carried out using a modified version of a motorized microdrive described previously (Fee and Leonardo 2001) mounted with arrays of 5–10 single electrodes (3–5 MΩ, Pt/Ir and Tungsten from microprobes.com). During microdrive implant surgery, the array was stereotactically targeted to area X based on previously established coordinates (5.8 mm anterior, 1.5 mm lateral, and 2.8 mm ventral), at a head angle 20° displaced from the flat groove on the anterior border of the bird's skull. Electrophysiological data were band-pass filtered in custom analog circuits (0.25–15 kHz) and were acquired at 40 kHz using custom Matlab acquisition software. Units accepted for analysis had signal-to-noise ratios (average spike peak amplitude compared with SD of noise) greater than 10:1 and were sorted by threshold crossings and template matching implemented in custom Matlab code, as previously described (Goldberg and Fee 2012). Small electrolytic lesions (20 μA for 15 s) were made through the electrodes at the conclusion of experiments for histological verification of electrode position.
Data analysis.
Spikes were sorted offline using custom Matlab software. We represented neural activities as instantaneous firing rates (IFR), R(t), defined at each time point as the inverse of the enclosed interspike interval as follows (Eq. 1):
where ti is the time of the ith spike. Peak firing rates (99th percentile rate) were calculated for each neuron as the inverse of its 1st percentile interspike interval. Nonsinging firing rates were calculated from spiking activity during silent periods more than 10 s separated from singing.
Identification of cell types.
BG cell types were identified on the basis of singing-related neural activity [for putative GPe, GPi, MSN, TAN, and low-threshold spike (LTS)] and spike waveform (for putative FS cells) as described previously (Goldberg et al. 2010; Goldberg and Fee 2010). For each neuron, the spike width was calculated as the half-width of the average of 50 spike waveform examples. Units with spike widths less than 0.06 ms were part of a distinct cluster and were identified as putative FS neurons (Goldberg and Fee 2010). Neurons with mean firing rate during singing less than 5 Hz and spike width greater than 0.06 ms were classified as putative MSNs; neurons with mean firing rate during singing greater than 10 Hz and peak firing rate (inverse of the 1st percentile interspike interval) less than 600 Hz were identified as putative TANs; those with peak firing rate greater than 600 Hz were identified as LTS neurons. Pallidal neurons were distinguished from the above striatal cell types by mean firing rate during singing greater than 50 Hz. Furthermore, to distinguish GPe neurons from GPi pallidal neurons, we used previously established criteria that leverage the distinct spectral properties of GPe and GPi firing: GPe-like neurons exhibit slow rate modulations with long burst and pauses, whereas GPi-like neurons exhibit high-frequency modulations without long bursts or pauses. To quantitatively distinguish these neurons, we smoothed the IFR with an finite impulse response equiripple filter with a pass band below 25 Hz and stop band greater than 75 Hz, with 80-dB attenuation in the stop band. To remove the DC offset (high average firing rate), IFRs were mean-subtracted using an infinite impulse response 1-Hz high-pass filter generated with coefficients b = [1 −1] and a = [1 −0.9988]. The peak of this smoothed IFR produces two distinct clusters that separate GPe-like neurons, which exhibit peaks in smoothed IFR greater than 250 Hz, from GPi-like neurons, which exhibit peaks less than 250 Hz (Goldberg et al. 2010).
Analysis of correlations of neural activity to syllable onsets and offsets.
Statistical significance of rate increases and decreases in syllable onset (or offset) aligned histograms was calculated as described previously (Goldberg and Fee 2012). Neural activity was aligned to the 200 ms preceding and following all syllable onsets and offsets. Only cells recorded for greater than 50 syllables were included in the study. To determine the significance of firing-rate peaks and troughs, a rate histogram (bin size, 10 ms) was generated of the real data. Then a surrogate histogram was generated in which each trial of syllable-aligned neural activity was time-shifted by a uniformly distributed random amount over a range equal to the duration of the histogram (400 ms). The shift was circular, such that spikes wrapped around to the beginning of the histogram, preserving the overall spike statistics of the data. The minimum and maximum of the surrogate rate histogram was then obtained with 1,000 repetitions of randomly shifted data. P values for the rate minimum and maximum of the real data set were calculated by analyzing the frequency with which shifted data sets generated larger maxima or smaller minima in firing rates. Maximal rate increases and decreases with P values <0.05 were considered significant. The amplitude of the rate modulation was computed as the maximal deviation from the average rate during the ±200 ms around syllable onset (or offset). The timing of the rate change relative to the syllable onset (or offset) was computed as the time of maximal rate change above (for rate increases) or below (for rate decreases) the baseline rate. To construct population rate histograms (as in Fig. 2D), histograms from individual cells were mean-subtracted and then averaged across the group.
Fig. 2.
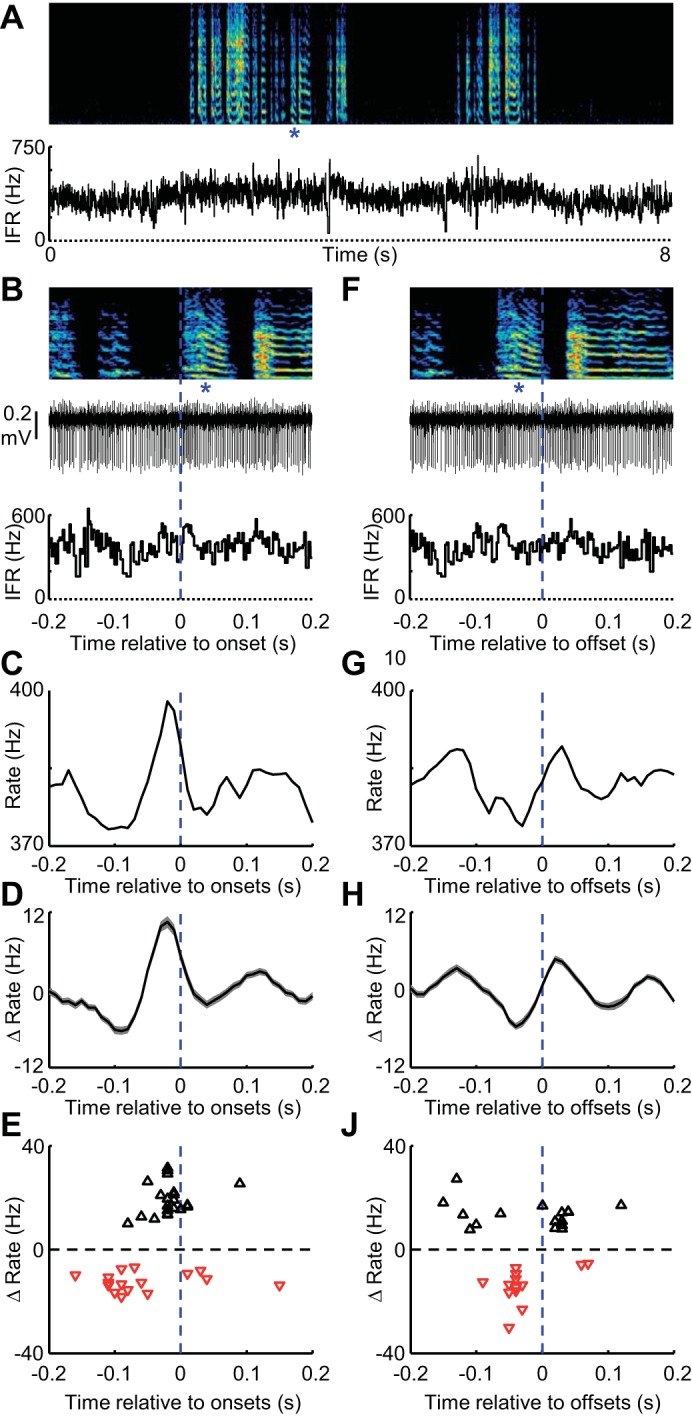
GPi-like neurons in area X exhibit rate peaks at syllable onsets during vocal babbling. A: spectrogram showing vocal babbling of a juvenile bird is shown above the instantaneous firing rate (IFR) of a single GPi axon terminal recorded in the medial nucleus of the dorsolateral thalamus (DLM). B: expanded view of song (top), spiking activity (middle) and IFR (bottom) aligned to the onset of a single syllable (marked with blue asterisks). C and D: syllable onset-aligned rate histograms for the neuron in shown in A and B (C) and for all GPi-like neurons recorded in area X and axon terminals recorded in DLM (D) (shading indicates ±SE) (n = 26). E: scatter plot showing the magnitude of statistically significant maximal rate increases (black triangles) and decreases (red inverted triangles) plotted against the time, relative to syllable onset, at which they occurred. Note that each cell can contribute at most one increase and decrease to this plot. Note also that the x-axis is the same for B–E. F–J: data are plotted exactly as in B–E for syllable offsets. Δ, Change.
Most syllables during vocal babbling are less than 200 ms in duration, and, because syllable onsets are preceded by syllable offsets and syllable offsets are followed by syllable onsets, average rate histograms of activity ±200 ms around a syllable boundary include firing rate changes, not only at the zero time point of the boundary of interest, but also over the preceding and following onsets and offsets (Aronov et al. 2008). For this reason, rate histograms of neurons with onset- and offset-locked firing appear periodic and exhibit side peaks and troughs. Thus, for each histogram with a significant rate change, we defined the “dominant” rate change (either an increase or a decrease) as the one that exhibited the largest deviation from the neuron's average firing rate (Table 1). For each dominant rate change, we computed five values: 1) its magnitude (in spike rate, Hz); 2–4) the timing of its peak (2), onset (3), and offset (4) (in milliseconds relative to syllable boundary, as in Fig. 2, E and J, and reported in Table 1); and 5) its modulation width (defined as the duration between its offset and onset). The onset was defined as the bin that exceeded 2 SDs from the mean in the baseline rate. The offset was the bin that returned to below 2 SDs from mean. The modulation width (ms) was the time separating the onset and offset of the rate change (Table 1).
Table 1.
The timing and magnitude of syllable-locked firing patterns across putatively identified cell classes during vocal babbling
| GPi | MSN | GPe | FS | TAN | LTS | |
|---|---|---|---|---|---|---|
| Dominant rate change, syllable onset, Hz | 18.9 ± 6.1 | 4.3 ± 1.7 | −19.8 ± 7.4 | 15.9 ± 14.0 | 20.0 ± 8.2 | N/S |
| Peak of dominant rate change, syllable onset, ms | −16 ± 32 | −9 ± 32 | −18 ± 30 | −8 ± 10 | −15 ± 13 | N/S |
| Onset of dominant rate change, syllable onset, ms | −45 ± 20 | −3 ± 60 | −37 ± 30 | −27 ± 17 | −43 ± 13 | N/S |
| Width of dominant rate change, onset, ms | 41 ± 20 | 30 ± 13*† | 47 ± 11 | 38 ± 13 | 53 ± 14 | N/S |
| No. cells with dominant rate increase at onset | 22/26 | 15/20 | 0/15 | 4/5 | 11/13 | 0/10 |
| No. cells with dominant rate decrease at onset | 0/26 | N/A | 12/15 | N/A | 0/13 | 0/10 |
| Onset reliability, %syllable onsets exhibiting dominant rate change | 60 ± 6 | N/A | 59 ± 5 | N/A | 67 ± 7*‡ | N/S |
| Dominant rate change, syllable offset, Hz | −13.7 ± 6.6 | N/A | 15.0 ± 5.0 | N/A | −17.2 ± 12.7 | N/S |
| Peak of dominant rate change, syllable offset, ms | −29 ± 42 | N/A | −1 ± 60 | N/A | −17 ± 16 | N/S |
| Onset of dominant rate change, syllable offset, ms | −43 ± 46 | N/A | −27 ± 67 | N/A | −43 ± 13 | N/S |
| Width of dominant rate change, offset, ms | 37 ± 14 | N/A | 38 ± 16 | N/A | −53 ± 13 | N/S |
| No. cells with dominant rate increase at offset | 40/26 | 0/20 | 6/15 | 0/5 | 0/13 | 0/10 |
| No. cells with dominant rate decrease at offset | 14/26 | N/A | 0/15 | N/A | 7/13 | 0/10 |
| Offset reliability, %syllable offsets exhibiting dominant rate change | 55 ± 15 | N/A | 55 ± 17 | N/A | 56 ± 3 | N/S |
Values are means ± SD across cells within a cell class.
GPi, globus pallidus internal segment; MSN, medium spiny neurons; GPe, globus pallidus external segment; FS, fast spiking; TAN, tonically active; LTS, low-threshold spiking; N/A, not applicable.
For each cell class, the magnitude of the dominant maximal rate change (Hz), its timing relative to syllable onset and offset, its reliability across syllables, and the number of cells with statistically significant dominant rate increases and decreases in syllable onset- and offset-aligned histograms are shown (see materials and methods). Note that these data were acquired from syllable-locked rate histograms that exhibited statistically significant rate changes. N/S indicates that there were no such histograms in that cell class dataset. Note also that rate decreases in rate histograms on onset/offset reliability were not computed in MSN and FS cell classes due to their sparse firing. A different metric, the percentage of syllables with a spike, is shown in Table 3. See materials and methods for analysis details.
P < 0.05 in unpaired t-test compared with the cell classes indicated.
<GPe and TAN.
>GPi and GPe.
For MSNs and putative FS cells with very low baseline firing rates and strong peaks at syllable onsets, significant peaks were also observed in offset-aligned histograms; closer inspection of offset-aligned rasters showed that these offset-peaks were due to high probability of firing in relation to the following (e.g., Fig. 3H) or preceding (e.g., see Fig. 5I) syllable onset. Thus, for MSNs and FS interneurons, we also defined a dominant activation as occurring at either onset or offset, the dominant one being the one with the larger peak. In all cases, this dominant rate change occurred in the syllable onset aligned histograms. Note that, because these cell classes exhibit such low baseline firing rates, we did not test for significance of rate decreases in syllable-aligned histograms of MSN or FS activity.
Fig. 3.
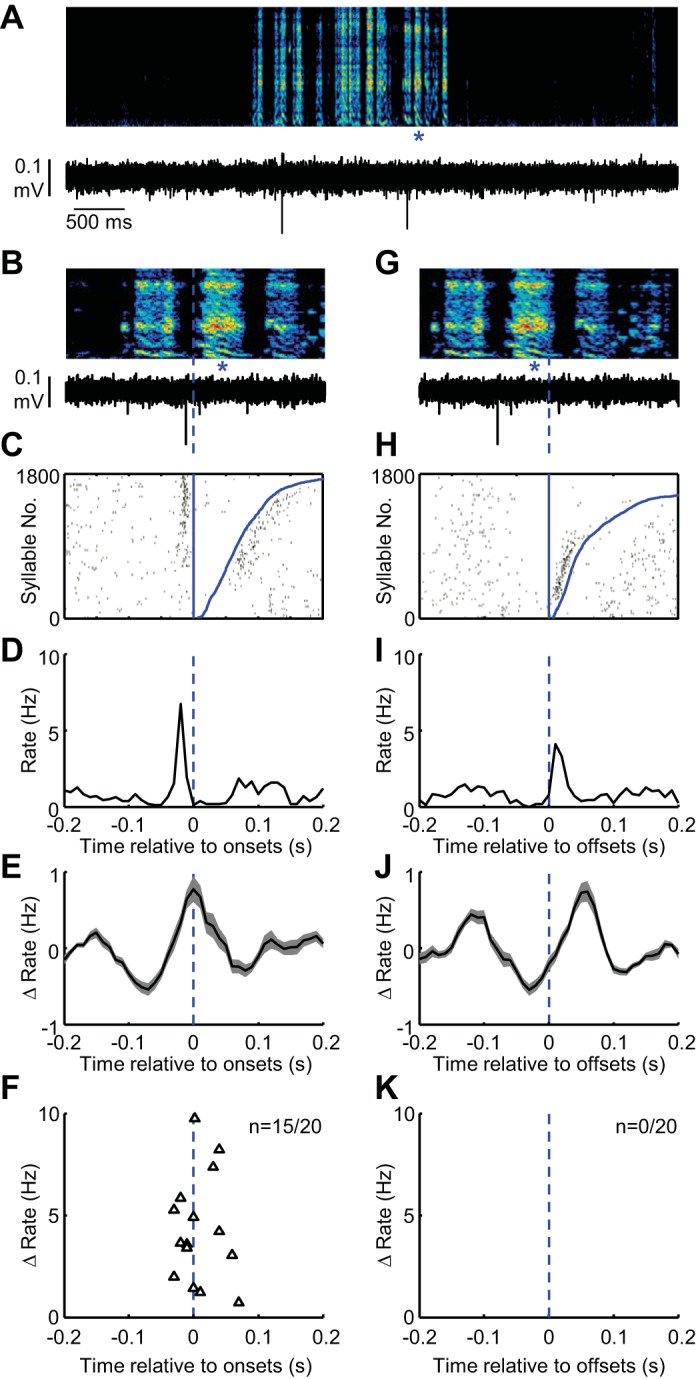
Putative MSNs in area X exhibit rate peaks at syllable onsets during vocal babbling. A: spectrogram showing vocal babbling of a juvenile bird is shown above the spiking activity of a single MSN in area X. B: expanded view of song (top) and spiking activity (bottom) aligned to the onset of a single syllable (marked with blue asterisks). C: syllable onset aligned raster plot showing the spiking activity of this neuron during 1,800 syllable renditions. The raster rows are sorted by the duration to the syllable offset (curved blue line at right of raster). To visualize sparse spikes as raster tics, tic height was 5 syllables. D and E: syllable onset-aligned rate histograms for the neuron in shown in A–C (D) and for all MSNs recorded in area X (E) (shading indicates ± SE) (n = 20). F: scatter plot showing the magnitude of significant dominant rate increases (black triangles) plotted against the time, relative to syllable onset, at which they occurred. Note that the x-axis is the same for B–F. G–K: data are plotted exactly as in B–E for syllable offsets. In the raster in H, the data are sorted by the arrival time of the following syllable onset. Note that the post-offset spiking is aligned to the following syllable onset. K: 0/20 MSNs exhibited their dominant rate peak at syllable offsets.
Fig. 5.
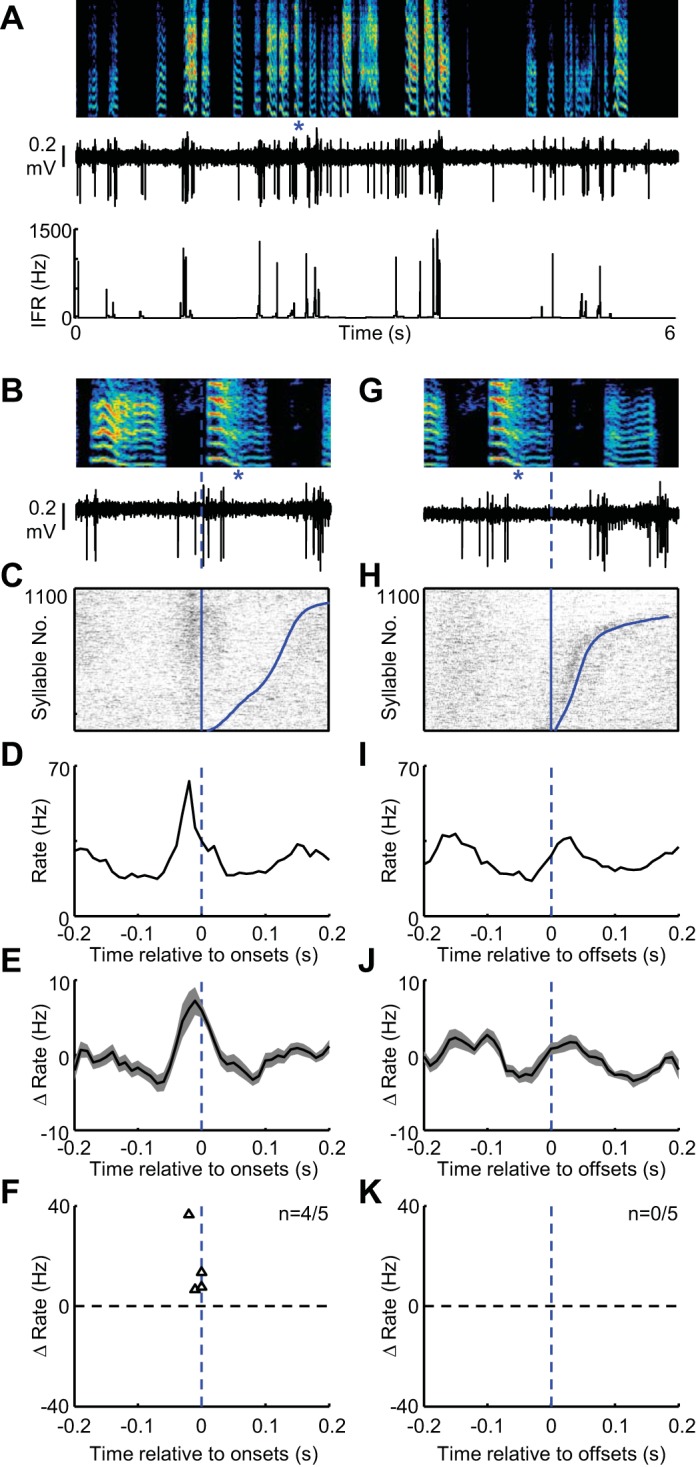
Putative FS interneurons in area X exhibit rate increases at syllable onsets during vocal babbling. A: spectrogram showing vocal babbling of a juvenile bird is shown above the spiking activity (middle) and IFR (bottom) of a single FS neuron recorded in area X. B: expanded view of song (top) and spiking activity (bottom) aligned to the onset of a single syllable (marked with blue asterisks). C: syllable onset aligned raster plot showing the spiking activity of this neuron during 1,100 syllable renditions. The data are sorted by the duration to the syllable offset (curved blue line at right of raster). Tic height in the raster is 3 syllables. Syllable onset-aligned rate histograms for the neuron in shown in A–C (D) and for all FS interneurons recorded in area X (shading indicates ± SE) (n = 5) (E). F: scatter plot showing the magnitude of dominant rate increases (black triangles, n = 4/5 FS neurons exhibited significant dominant rate peaks at onset) plotted against the time, relative to syllable onset, at which they occurred. Note that the x-axis is the same for B–F. G–K: data are plotted exactly as in B–E for syllable offsets. H: the syllable-offset aligned raster is sorted by the arrival time of the following syllable onset (curved blue line at right of raster). K: 0/5 FS neurons exhibited significant dominant rate peaks at offset.
To quantitatively test if average firing rate peaks (and dips) were clustered at syllable boundaries beyond predicted by chance, we determined, for each neuron, the timing of the dominant rate change at onset and offset, as described above. Next, for each cell class, we constructed a “real data” histogram that plotted the number of cells exhibiting a dominant rate change as a function of time relative to the syllable boundary (−0.3 to +0.3 s relative to syllable boundary; 0.2-s bins). We also computed 1,000 surrogate histograms with the same data, but with each cell's timing randomly shuffled. We computed the P value of the peak observed in the real data by analyzing the frequency with which shuffled data sets generated a larger peak than what was observed. P values <0.01 were considered significant.
To determine the reliability of neuronal firing rate changes across syllables, we computed the percentage of syllables in which a neuron exhibited an increased rate at the time of its dominant rate change (quantified in the average syllable onset aligned rate histogram). Specifically, for each neuron with a dominant rate change, we examined the mean IFR that occurred in a 60-ms time window centered at the time of its dominant rate change and then determined the frequency with which this mean was greater than that observed in the preceding 60-ms window, as previously described (Goldberg and Fee 2012).
HVC lesions.
HVC was lesioned bilaterally using large electrolytic lesions, as previously described in detail (Aronov et al. 2008; Goldberg and Fee 2011). Lesions were confirmed histochemically using fluorescent antibody to neuronal nuclei (Mouse anti-Neu-N; Millipore, Temecula, CA) and/or the absence of retrograde label (10,000 mol wt Alexa dextran; www.lifetechnologies.com) of HVC following injection into area X during the microdrive implant surgery. In control birds, tracer injection into area X results in pronounced labeling of HVC. These neurons were confirmed to be absent following HVC lesion.
RESULTS
GPi-like neurons in areka X, as well as their axon terminals recorded in the medial nucleus of the dorsolateral thalamus (DLM) exhibit increased firing rates in the millisecond prior to onsets and decreases prior to offsets (Fig. 2, Table 1; rate changes were clustered at syllable boundaries, P < 0.001, see materials and methods) (Goldberg et al. 2012; Goldberg and Fee 2012). Different pathways inside area X could “construct” these GPi output signals. For example, if the direct pathway is primarily involved, one would expect direct pathway MSNs to exhibit decreased and increased probability of firing prior to onsets and offsets, respectively. If the MSN-GPe-GPi indirect pathway is involved, one might expect GPe neurons to exhibit rate decreases at onsets, and increases at offsets. Hyperdirect pathway involvement, via direct cortical projections to area X output neurons (Farries et al. 2005), might bypass intrinsic BG circuits altogether and makes no clear prediction about their firing. Finally, the relationship between striatal interneuronal firing to BG output signals remains poorly understood. To test these distinct scenarios, we recorded upstream cell types inside area X.
Putative MSNs are distinguished by their very low firing rate (<5 Hz during singing) and their ultra-sparse song-locked firing (Goldberg and Fee 2010; Woolley et al. 2014). MSNs fired similarly sparsely in babbling birds, exhibiting a small but significant increase in firing rate during singing (mean rate during silent periods: 0.22 ± 0.8 Hz vs. mean rate during singing 0.80 ± 0.5 Hz, P < 0.05, paired t-test, n = 20 neurons) and discharging during only 10.6 ± 6.0% of syllables (mean ± SD). Inspection of syllable-locked firing revealed that most exhibited a significantly higher probability of spiking at syllable onsets (rate increase of 4.3 ± 1.7 Hz occurring at −9.0 ± 32 ms relative to syllable onsets, statistically significant rate increases in n = 15/20 neurons, Table 1, Fig. 3F, see materials and methods). MSNs with onset peaks also exhibited peaks in offset-aligned histograms. However, examination of syllable offset-aligned spike rasters that were sorted by the arrival time of the preceding or following syllable onset clearly showed that these peaks were attributable to the discharge related to the neighboring onsets (e.g., see Fig. 3, H and I). Thus for each MSN we defined the dominant peak as occurring in either onset or offset aligned histograms as the one with the larger rate change from baseline (see materials and methods). Of the 15/20 neurons that exhibited statistically significant syllable-locked activations, all exhibited dominant peaks at syllable onsets, and none of them at offsets (Fig. 3, Table 1, peaks were clustered at onsets, P < 0.001, see materials and methods).
The timing of MSN firing does not support a direct-pathway origin of GPi onset peaks. If anything, MSN-GPi inhibition would produce pauses prior to syllable onsets and peaks prior to syllable offset, the opposite of what was observed in the GPi-like neurons. Yet MSNs also target GPe-like neurons, which can suppress firing of GPi neurons. If this pathway contributes to syllable timing signals in GPi neurons, one would expect GPe neurons to exhibit rate decreases prior to onsets, and increases prior to offsets. To test this hypothesis, we recorded GPe-like neurons in area X. As reported previously, these neurons discharged like many primate GPe neurons, exhibiting large bursts and long pauses during singing, a type of firing never observed in the thalamus-projecting GPi-like outputs in area X (Goldberg et al. 2010; Woolley et al. 2014) (Table 2, see materials and methods). Consistent with a role for the indirect pathway in shaping GPi firing, GPe neurons exhibited, on average, rate decreases at syllable onsets, and some also exhibited rate increases prior to offsets (n = 12/15 and 6/15, respectively; Fig. 4, Table 1, rate changes were significantly clustered at syllable boundaries, P < 0.001, see materials and methods). Together, these findings are consistent with a scenario in which MSN rate increases suppress GPe firing at syllable onsets, which in turn may disinhibit GPi firing at this time.
Table 2.
Cell-class specific spiking characteristics of pallidal neurons persist following HVC lesion
| GPi | GPi (No HVC) | GPe | GPe (No HVC) | |
|---|---|---|---|---|
| Rate, nonsinging, Hz | 119.5 ± 48.0 | 122.1 ± 27.8 | 113.5 ± 28.8 | 108.2 ± 37.6 |
| Rate, singing, Hz | 263.1 ± 67.5 | 249.2 ± 58.3 | 125.5 ± 33.3 | 152.3 ± 49.3 |
| CV, nonsinging | 0.33 ± 0.13 | 0.30 ± 0.09 | 0.36 ± 0.08 | 0.36 ± 0.10 |
| CV, singing | 0.50 ± 0.10 | 0.43 ± 0.09 | 0.87 ± 0.27 | 0.85 ± 0.35 |
| 99th percentile ISI, nonsinging, ms | 16.4 ± 6.8 | 15.5 ± 6.7 | 20.2 ± 7.8 | 22.9 ± 11.7 |
| 99th percentile ISI, singing, ms | 11.2 ± 4.4 | 12.1 ± 5.6 | 42.5 ± 30.0 | 34.9 ± 22.5 |
| Peak in smoothed firing rate (range: min–max) | 195 ± 21 (153–228) | 189 ± 28 (151–249) | 404 ± 88 (266–583) | 404 ± 112 (253–587) |
Values are means ± SD across cells within a cell class. Singing-related changes in firing rates and interspike interval (ISI) statistics were not affected by HVC lesion. The peak in the smoothed instantaneous firing rate, the metric used to distinguish GPe- from GPi-like neurons, was also not affected (see materials and methods).
CV, coefficient of variation.
Fig. 4.
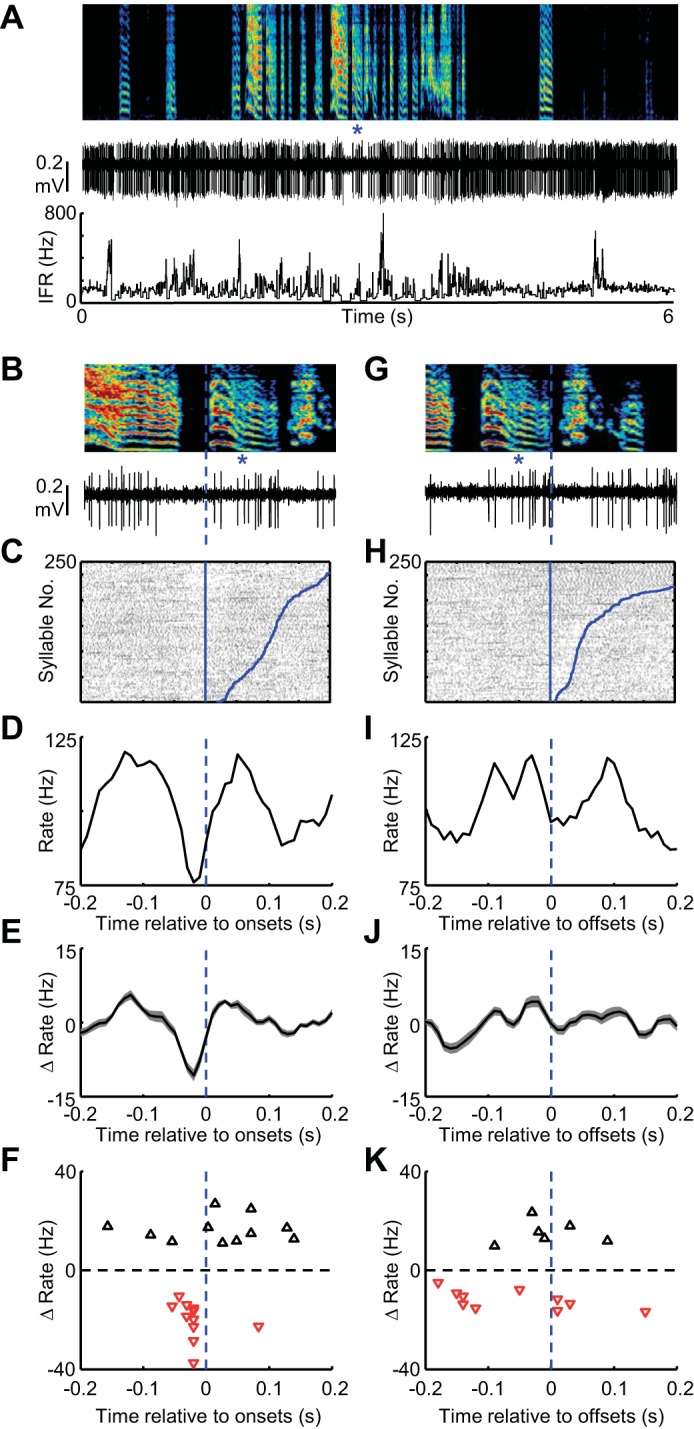
GPe-like neurons in area X exhibit rate decreases at syllable onsets during vocal babbling. A: spectrogram showing vocal babbling of a juvenile bird is shown above the spiking activity (middle) and IFR (bottom) of a single GPe-like neuron recorded in area X. B: expanded view of song (top) and spiking activity (bottom) aligned to the onset of a single syllable (marked with blue asterisks). C: syllable onset aligned raster plot showing the spiking activity of this neuron during 250 syllable renditions. The data are sorted by the duration to the syllable offset (curved blue line at right of raster). Tic height in the raster is 1 syllable. Syllable onset-aligned rate histograms for the neuron in shown in A–C (D) and for all GPe neurons recorded in area X (shading indicates ± SE) (n = 15) (E). F: scatter plot showing the magnitude of significant rate increases (black triangles) and rate decreases (red inverted triangles) plotted against the time, relative to syllable onset, at which they occurred. Note that the x-axis is the same for B–F. G–K: data are plotted exactly as in B–E for syllable offsets. K: raster rows are sorted by the duration to the next syllable onset.
Three types of striatal interneuron are conserved between songbirds and mammals, FS, LTS and TAN, yet it remains unclear how their activities shape MSN and GP firing. We recorded units putatively identified as these subclasses and found that both FS and TAN cell types also exhibited rate peaks at syllable onsets and decreases at syllable offsets (Figs. 5 and 6, Table 1). Meanwhile, LTS neurons exhibited highly randomly timed high-frequency bursts, with weak, if any, relationship to syllable timing (Fig. 7, Table 1).
Fig. 6.
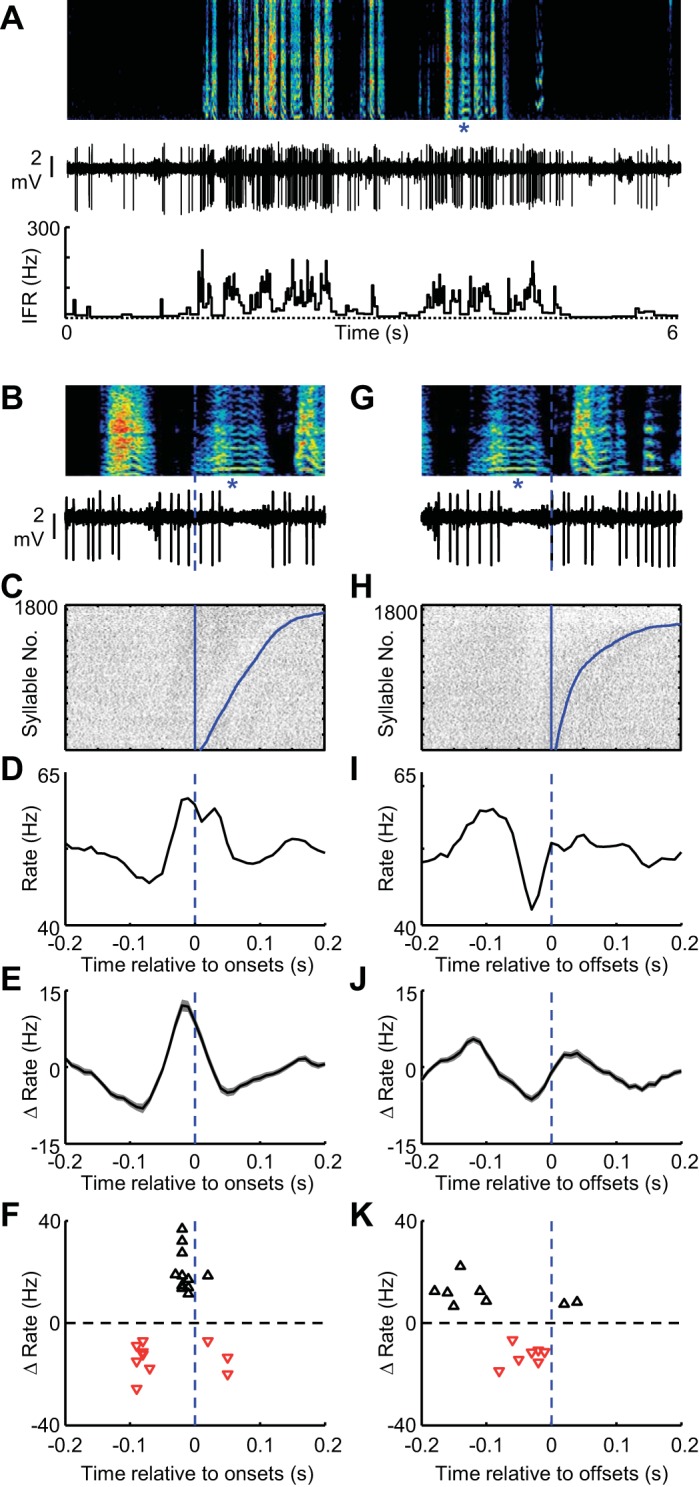
Putative TAN interneurons in area X exhibit rate increases at syllable onsets during vocal babbling. A: spectrogram showing vocal babbling of a juvenile bird is shown above the spiking activity (middle) and IFR (bottom) of a putative TAN interneuron recorded in area X. B: expanded view of song (top) and spiking activity (bottom) aligned to the onset of a single syllable (marked with blue asterisks). C: syllable onset aligned raster plot showing the spiking activity of this neuron during 1,800 syllable renditions. The data are sorted by the duration to the syllable offset (curved blue line at right of raster). Tic height in the raster is 1 syllable. Syllable onset-aligned rate histograms for the neuron shown in A–C (D) and for all TAN interneurons recorded in area X (shading indicates ± SE) (n = 13) (E). F: scatter plot showing the magnitude of significant rate increases (black triangles) and rate decreases (red inverted triangles) plotted against the time, relative to syllable onset, at which they occurred. Note that the x-axis is the same for B–F. G–K: data are plotted exactly as in B–E for syllable offsets. H: the syllable-offset aligned raster is sorted by the arrival time of the following syllable onset (curved blue line at right of raster).
Fig. 7.
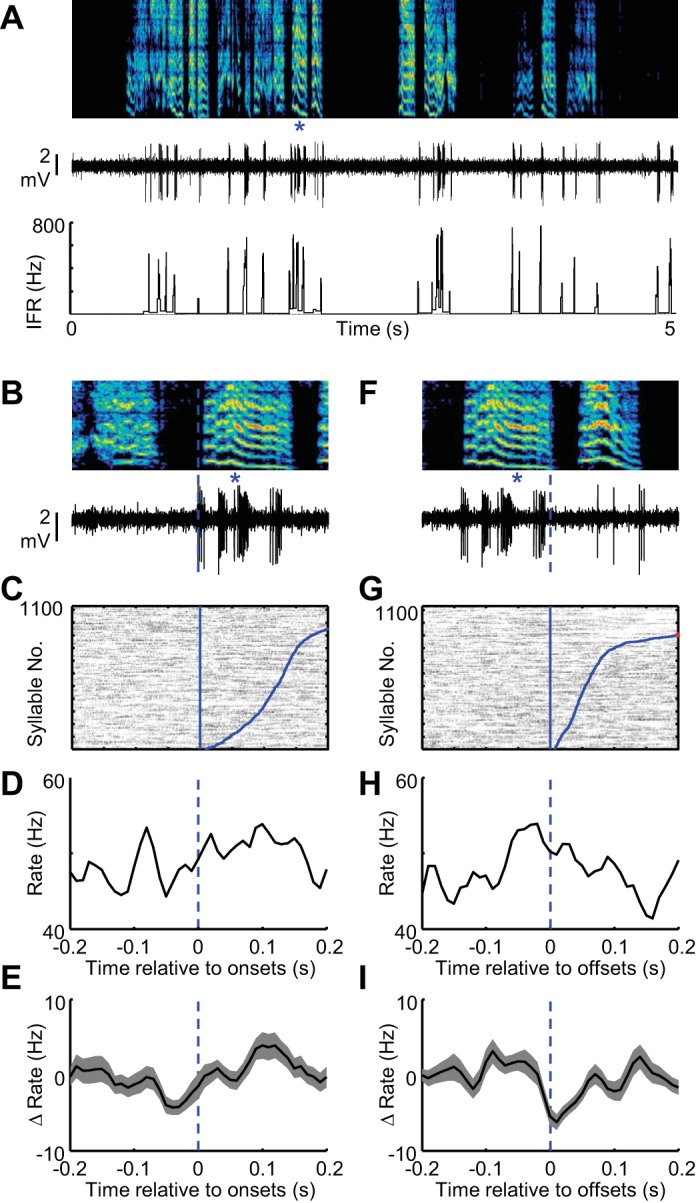
Putative LTS interneurons in area X do not exhibit strong syllable-locked firing during vocal babbling. A: spectrogram showing vocal babbling of a juvenile bird is shown above the spiking activity (middle) and IFR (bottom) of a putative LTS interneuron recorded in area X. B: expanded view of song (top) and spiking activity (bottom) aligned to the onset of a single syllable (marked with blue asterisks). C: syllable onset aligned raster plot showing the spiking activity of this neuron during 1,100 syllable renditions. The data are sorted by the duration to the syllable offset (curved blue line at right of raster). Tic height in the raster is 4 syllables. Syllable onset-aligned rate histograms for the neuron shown in A–C (D) and for all LTS interneurons recorded in area X (shading indicates ± SE) (n = 10) (E). Note that the x-axis is the same for B–E. F–I: data are plotted exactly as in B–E for syllable offsets. H: the syllable-offset aligned raster is sorted by the arrival time of the following syllable onset (curved blue line at right of raster).
Note that all of the syllable-locked signals reported thus far are computed from syllable-aligned rate histograms, which reflect the average firing of a neuron across syllables. Yet a neuron need not increase its discharge at every syllable to yield a strong peak in the histogram. For each neuron in the GPi, GPe and TAN cell classes, we defined its “reliability” as the percentage of syllables during which an observed rate change occurred in the direction of the dominant rate change (see materials and methods). At syllable onsets, TAN neurons exhibited increased firing rates prior to 67 ± 7% of syllables (P < 0.01 compared with GPi: 60 ± 6%; and to GPe: 59 ± 5%, Table 1). There were no significant intergroup differences in reliability of syllable-offset locked rate changes (Table 1).
We next wondered what input to area X might cause these remarkably homogenous firing patterns. The two main cortical inputs to area X, HVC and lateral magnocellular nucleus of the anterior nidopallium (LMAN), play different roles in vocal babbling. LMAN lesions abolish singing altogether in juvenile babbling birds, and abolish vocal variability in older plastic song birds, causing premature song crystallization (Aronov et al. 2008; Bottjer et al. 1984; Brainard and Doupe 2001; Olveczky et al. 2005; Scharff and Nottebohm 1991). Meanwhile, HVC lesions have little, if any, impact on song structure during vocal babbling (Aronov et al. 2008). Thus HVC lesions provide an opportunity to record area X neurons during a singing behavior that is highly similar to vocal babbling.
To test how syllable onset and offset aligned firing patterns depend on inputs from HVC, we recorded GPi, GPe and MSN cell classes after complete bilateral HVC lesion in juvenile birds (see materials and methods). As reported previously, song after HVC lesion resembled vocal babbling (Figs. 8–10) (Aronov et al. 2008; Chen et al. 2014b; Goldberg and Fee 2011). We also found that the basic spiking statistics of pallidal neurons were not significantly affected by HVC lesion, and the same analyses that distinguished these cell types in intact birds distinguished them in HVC-lesioned birds (Table 2). In MSNs, average firing rate during singing, but not during silent periods, was slightly but significantly elevated in HVC-lesioned birds (average rate during singing, control: 0.88 ± 0.50 Hz vs. HVC-lesioned: 1.81 ± 1.8 Hz, P < 0.05 Wilcoxon rank-sum test, n = 13 MSNs). MSNs in HVC-lesioned birds also discharged during a higher percentage of syllables (percentage of syllables with MSN spike, control: 10.6 ± 6.0%, vs. HVC lesioned: 18.1 ± 7.7%, P < 0.05 Wilcoxon rank-sum test) (Table 3).
Fig. 8.
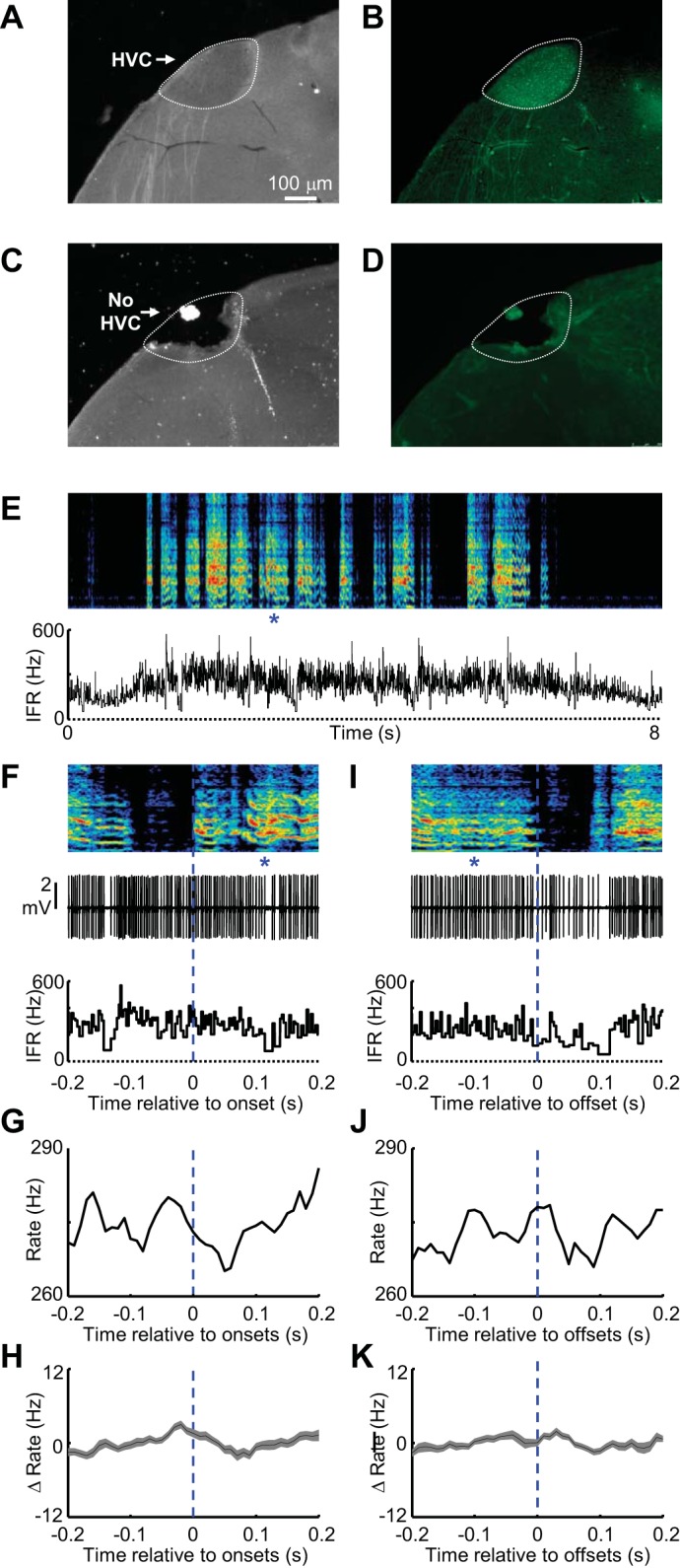
HVC lesions abolish syllable-locked firing in area X GPi neurons. A–D: histological verification of complete HVC lesion. A and B: in unlesioned birds, HVC is clearly visible in dark-field (A) and via fluorescent cell bodies retrogradely labeled from injection of Alexa dextran tracer into area X (white dotted line, see materials and methods) (B). C and D: electrolytic lesion of HVC was confirmed by visible destruction of the structure visible in dark-field (C) and in the failure to observe retrogradely labeled neurons following injection of tracer into area X (D). Scale bar in A applies to A–D. E: spectrogram showing post-HVC lesion song of the juvenile bird whose brain is shown in C and D. This song is shown above the IFR of a GPi neuron in area X. F: expanded view of song (top), spiking activity (middle) and IFR (bottom) aligned to the onset of a single syllable (marked with blue asterisks). Syllable onset-aligned rate histograms for the neuron are shown in E and F (G) and for all GPi-like neurons recorded in area X after HVC lesion (shading indicates ± SE) (n = 17) (H). Note that the x-axis is the same for F–H. I–K: data are plotted exactly as in F–H for syllable offsets. In G and H and J and K, the ordinate scale was matched to the scale used in homologous plots for control GPi neurons in Fig. 2.
Fig. 10.
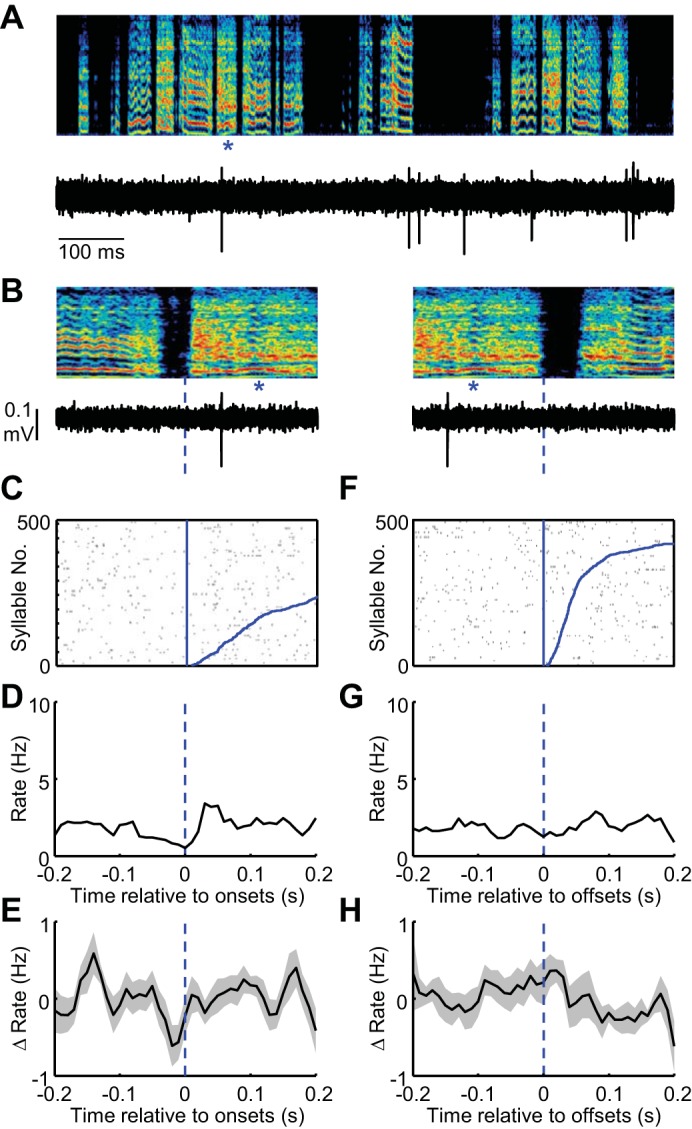
HVC lesions abolish syllable-locked firing in area X MSNs. A: spectrogram showing post-HVC lesion song of a juvenile bird is shown above the spiking activity of a single MSN in area X. B: expanded view of song (top) and spiking activity (bottom) aligned to the onset of a single syllable (marked with blue asterisks). C: syllable onset aligned raster plot showing the spiking activity of this neuron during 500 syllable renditions. The data are sorted by the duration to the syllable offset (curved blue line at right of raster). Tic height in the raster was 5 syllables. D and E: syllable onset-aligned rate histograms for the neuron in shown in A–C (D) and for all MSNs recorded in area X (E) (shading indicates ± SE) (n = 20). F–H: data are plotted exactly as in B–E for syllable offsets. H: in the raster, the data are sorted by the arrival time of the following syllable onset.
Table 3.
Spiking characteristics of putative MSNs following HVC lesion
| MSN | MSN (No HVC) | |
|---|---|---|
| n | 20 | 13 |
| Rate, nonsinging, Hz | 0.22 ± 0.79 | 0.64 ± 0.72 |
| Rate, singing, Hz | 0.88 ± 0.50 | 1.81 ± 1.18* |
| Syllables with spike, % | 10.6 ± 6.0 | 18.1 ± 7.7* |
Values are means ± SD across cells within a cell class; n, no. of medium spiny neurons (MSNs).
P < 0.05 in Wilcoxon rank sum tests.
HVC lesions largely abolished syllable-locked firing across cell types. In HVC-lesioned birds producing babbling-like vocalizations, only 2/15 GPi-like neurons had significant rate peaks at syllable onsets, and none had significant rate decreases at offsets (Fig. 8). Only 2/19 GPe-like neurons had rate decreases at syllable onsets, and 2/19 had peaks at offsets (Fig. 9). Only 2/13 MSNs exhibited rate peaks in syllable onset aligned histogram (Fig. 10), and none exhibited significant rate changes at offsets. Together, these data suggest that HVC is a principal driver of syllable timing-related firing in area X of babbling birds.
Fig. 9.
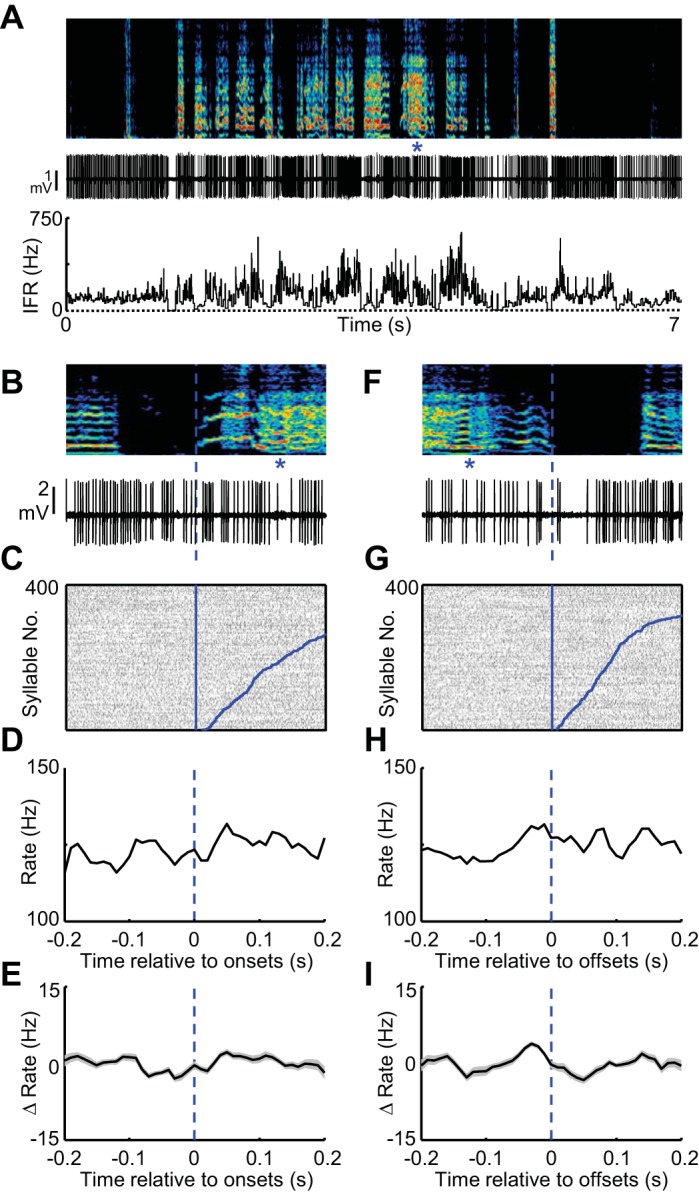
HVC lesions abolish syllable-locked firing in area X GPe neurons. A: spectrogram showing post-HVC lesion song of a juvenile bird is plotted above the spiking activity (middle) and IFR (bottom) of a single GPe-like neuron recorded in area X of a bird with complete bilateral HVC lesion. B: expanded view of song (top) and spiking activity (bottom) aligned to the onset of a single syllable (marked with blue asterisks). C: syllable onset aligned raster plot showing the spiking activity of this neuron during 400 syllable renditions. The data are sorted by the duration to the syllable offset (curved blue line at right of raster). Tic height in the raster is 1 syllable. Syllable onset-aligned rate histograms for the neuron in shown in A–C (D) and for all GPe neurons recorded in area X of HVC-lesioned birds (shading indicates ± SE) (n = 19) (E). F–I: data are plotted exactly as in B–E for syllable offsets.
The importance of HVC in driving syllable-locked firing raises the specific prediction that timing signals in area X should be syllable specific. Specifically, in subsong and early plastic songbirds, short syllables less than 50 ms are abolished by LMAN lesion, elongated by mild LMAN cooling, but not strongly affected by HVC lesion or cooling (Aronov et al. 2011; Veit et al. 2011). To test if area X neurons exhibit diminished timing-related signals at short syllables, we examined onset- and offset-locked firing at short (<50 ms) syllables. The magnitude of dominant peaks and dips at short-syllable onsets was reduced in all cell types (Table 4), consistent with a diminished role of HVC in contributing to the initiation of brief syllables.
Table 4.
Syllable-locked rate modulations during syllables <50 ms in duration
| GPi | MSN | GPe | FS | TAN | LTS | |
|---|---|---|---|---|---|---|
| Dominant rate change, short syllable onsets, Hz | 11.3 ± 15.0* | 0.9 ± 2.4* | −10.8 ± 10.0* | 4.8 ± 6.9 | 10.8 ± 14.0* | N/S |
| Dominant rate change, short syllable offsets, Hz | −14.2 ± 19.0 | N/A | 14.4 ± 22.0 | N/A | −8.6 ± 6.8 | N/S |
Values are means ± SD across cells within a cell class. Data are plotted as in Table 1 showing magnitude of the dominant rate change for short syllables.
P < 0.05, indicates that the rate change was significantly less than that observed for all-duration syllables, shown in Table 1.
DISCUSSION
We studied the origins of BG output signals during vocal babbling in juvenile zebra finches. Previous studies showed that, at this early stage of vocal development, the GPi-like BG output neurons in the songbird area X exhibit rate increases prior to syllable onsets and decreases prior to syllable offsets. Here we report that syllable-locked firing is similarly homogenous for upstream BG cell classes. Putative MSN, TAN and FS cell classes exhibited transient rate increases at syllable onsets, and rate decreases at offsets, while GPe-like neurons exhibit transient decreases at onsets. Meanwhile, putative LTS neurons were unmodulated by syllable timing. HVC lesions did not significantly affect babbling behavior but largely abolished these song timing signals inside area X.
Together, these findings are consistent with a role for the MSN-GPe-GPi indirect pathway inside area X in transforming cortically derived timing signals into a BG output signal. Yet several caveats are relevant to this interpretation, and other scenarios cannot be ruled out. First, HVC projects directly onto pallidal neurons in area X (Farries et al. 2005). Thus the peak in GPi firing at syllable onsets might also arise from hyperdirect inputs form HVC (Fig. 1B). Support for a role of the hyperdirect pathway is that many MSNs exhibited onset peaks (Fig. 3F) after the occurrence of GPe onset troughs (Fig. 4E) and GPi onset peaks (Fig. 2E; Table 1). These “late-peaking” MSN neurons are therefore unlikely to play a role in sculpting the average BG output signal and may reflect more complex processing inside the BG, perhaps related to syllable phonology or elongation. Note that, for this hyperdirect pathway alone to explain the syllable locked firing of the other neurons in area X, GPi-GPe-MSN feedback would be required, which, although not impossible, is opposite the direction of signal propagation described in this circuit in mammals (Hikosaka 2007; Jaeger and Kita 2011; Nambu et al. 2002).
A second caveat is that we were unable to determine in our recordings which “type” of MSN we recorded and thus have no direct evidence for their participation in direct or indirect pathways or both. Specifically, area X MSNs may express exclusively D1 receptors or D2 receptors, but, unlike mammalian MSNs, most songbird ones express both (Kubikova et al. 2010). Based on evolutionarily conserved patterns (Stephenson-Jones et al. 2012), it is possible that these distinct subtypes project differentially to GPe- and GPi-like neurons in area X, but this remains unknown. Assuming that we randomly sampled MSN subtypes in area X, it is likely that the 20 we recorded represented a mix of MSN cell types. If this is the case, then this would suggest that both direct and indirect pathway MSNs synchronously exhibit rate increases at syllable initiation, similar to what was recently observed in mice in the context of locomotor initiation (Cui et al. 2013). However, we cannot rule out the possibility that the 5/20 MSNs that did not exhibit strong syllable-locked firing were a specific MSN subtype.
A third caveat is that, while GPi firing increases, on average, prior to syllable onsets, rate increases were not observed prior to each individual syllable (see reliability in Table 1) (Goldberg and Fee 2012). By focusing on average syllable locked rate histograms, we did not emphasize neural firing that varies on a syllable-by-syllable basis; instead, we focused on possible sources of the robust GPi onset signal only observable by averaging across syllables. Thus, while our data are consistent with a role for the indirect and hyperdirect pathways in shaping this average BG output signal, on a trial-by-trial basis, signal propagation is likely to be more complex.
A final caveat relates to our assignment of cell class type in our area X recordings. In past work, we discovered six distinct singing-related firing patterns in area X (Goldberg et al. 2010; Goldberg and Fee 2010), and we attempted to map their identities to the six cell classes that were characterized in area X in vitro (Carrillo and Doupe 2004; Farries et al. 2005; Perkel et al. 2002). This mapping was based on the similarities to well-characterized firing patterns of potentially homologous mammalian BG cell types. Yet, because extracellular recording alone is not sufficient to unambiguously identify a neuron type, our cell type assignments in this paper represent our current best hypotheses for cell identity. This caveat is particularly important for the striatal interneurons whose firing patterns during behavior remain poorly understood.
An outstanding question is the functional significance of the syllable timing signals inside area X during babbling. The production of vocal babbling is not strongly affected by lesions of HVC or of area X (Aronov et al. 2008, 2011; Chen et al. 2014a; Goldberg and Fee 2011); therefore, these signals are not necessary for vocal motor production and are unlikely to be actually driving syllable onsets and offsets. Instead, because area X lesions block learning (Goldberg and Fee 2011; Scharff and Nottebohm 1991), syllable timing signals in area X might reflect a learning signal important for advancing the subsong-plastic song transition. Specifically, this transition is the earliest manifestation of vocal motor learning in birdsong and is characterized by the emergence of a “protosyllable” of discernable phonological and temporal structure that is repeated in a rhythmic fashion (Aronov et al. 2011; Tchernichovski et al. 2001). The critical neural event underlying protosyllable emergence is the linkage of a syllable offset to its own onset. This linkage enables a syllable to become stereotyped in duration and to be repetitively produced: two events that underlie the first emergence of song rhythm (Aronov et al. 2011; Lipkind et al. 2013; Saar and Mitra 2008; Veit et al. 2011). Given the importance of syllable onset and offset times in this process, an intriguing hypothesis is that syllable boundary signals observed inside area X in this study are important for the early emergence of temporal structure in the song itself. To test this and other hypotheses, it will be necessary to manipulate activity in area X firing during babbling, for example, with opto- or chemogenetics. Manipulations that influence song in “real time” would support a model where area X is an “actor” that biases vocalizations. Manipulations that only affect protosyllable learning would support a model where area X may function more like a “critic” (Doya and Sejnowski 1995; Fee and Goldberg 2011; Sutton and Barto 1998). Notably, protosyllables resemble “reduplication” events that occur early in human babbling (Doupe and Kuhl 1999; Oller 1980). Because BG disorders impair vocal learning in humans (Enard 2011), determining the functional significance of BG timing signals in babbling birds may illuminate neural mechanisms by which the earliest exploratory utterances acquire nascent temporal patterns during learning.
GRANTS
Funding to J. H. Goldberg was provided by the National Institute of Neurological Disorders and Stroke (Grant R00-NS-067062), the Klingenstein Neuroscience Foundation, and the Pew Biomedical Scholars Award.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: M.P. and J.H.G. conception and design of research; M.P., T.B., T.R., and J.H.G. performed experiments; M.P., T.R., and J.H.G. analyzed data; M.P., T.R., and J.H.G. interpreted results of experiments; M.P., T.R., and J.H.G. prepared figures; M.P., T.B., T.R., and J.H.G. approved final version of manuscript; T.B., T.R., and J.H.G. edited and revised manuscript; J.H.G. drafted manuscript.
REFERENCES
- Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Trends Neurosci 12: 366–375, 1989. [DOI] [PubMed] [Google Scholar]
- Anderson ME, Horak FB. Influence of the globus pallidus on arm movements in monkeys. III. Timing of movement-related information. J Neurophysiol 54: 433–448, 1985. [DOI] [PubMed] [Google Scholar]
- Anderson ME, Turner RS. A quantitative analysis of pallidal discharge during targeted reaching movement in the monkey. Exp Brain Res 86: 623–632, 1991. [DOI] [PubMed] [Google Scholar]
- Aronov D, Andalman AS, Fee MS. A specialized forebrain circuit for vocal babbling in the juvenile songbird. Science 320: 630–634, 2008. [DOI] [PubMed] [Google Scholar]
- Aronov D, Veit L, Goldberg JH, Fee MS. Two distinct modes of forebrain circuit dynamics underlie temporal patterning in the vocalizations of young songbirds. J Neurosci 31: 16353–16368, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bottjer SW, Miesner EA, Arnold AP. Forebrain lesions disrupt development but not maintenance of song in passerine birds. Science 224: 901–903, 1984. [DOI] [PubMed] [Google Scholar]
- Brainard MS, Doupe AJ. Postlearning consolidation of birdsong: stabilizing effects of age and anterior forebrain lesions. J Neurosci 21: 2501–2517, 2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown J, Pan WX, Dudman JT. The inhibitory microcircuit of the substantia nigra provides feedback gain control of the basal ganglia output. Elife 3: e02397, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bryden DW, Johnson EE, Diao X, Roesch MR. Impact of expected value on neural activity in rat substantia nigra pars reticulata. Eur J Neurosci 33: 2308–2317, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrillo GD, Doupe AJ. Is the songbird Area X striatal, pallidal, or both? An anatomical study. J Comp Neurol 473: 415–437, 2004. [DOI] [PubMed] [Google Scholar]
- Chen JR, Stepanek L, Doupe AJ. Differential contributions of basal ganglia and thalamus to song initiation, tempo, and structure. J Neurophysiol 111: 248–257, 2014a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen JR, Stepanek L, Doupe AJ. Differential contributions of basal ganglia and thalamus to song initiation, tempo, and structure. J Neurophysiol 111: 248–257, 2014b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui G, Jun SB, Jin X, Pham MD, Vogel SS, Lovinger DM, Costa RM. Concurrent activation of striatal direct and indirect pathways during action initiation. Nature 494: 238–242, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doupe AJ, Kuhl PK. Birdsong and human speech: common themes and mechanisms. Annu Rev Neurosci 22: 567–631, 1999. [DOI] [PubMed] [Google Scholar]
- Doupe AJ, Perkel DJ, Reiner A, Stern EA. Birdbrains could teach basal ganglia research a new song. Trends Neurosci 28: 353–363, 2005. [DOI] [PubMed] [Google Scholar]
- Doya K, Sejnowski T. A novel reinforcement model of birdsong vocalization learning. Adv Neural Inf Process Syst 7: 101–108, 1995. [Google Scholar]
- Enard W. FOXP2 and the role of cortico-basal ganglia circuits in speech and language evolution. Curr Opin Neurobiol 21: 415–424, 2011. [DOI] [PubMed] [Google Scholar]
- English DF, Ibanez-Sandoval O, Stark E, Tecuapetla F, Buzsaki G, Deisseroth K, Tepper JM, Koos T. GABAergic circuits mediate the reinforcement-related signals of striatal cholinergic interneurons. Nat Neurosci 15: 123–130, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farries MA, Ding L, Perkel DJ. Evidence for “direct” and “indirect” pathways through the song system basal ganglia. J Comp Neurol 484: 93–104, 2005. [DOI] [PubMed] [Google Scholar]
- Farries MA, Perkel DJ. A telencephalic nucleus essential for song learning contains neurons with physiological characteristics of both striatum and globus pallidus. J Neurosci 22: 3776–3787, 2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fee MS, Goldberg JH. A hypothesis for basal ganglia-dependent reinforcement learning in the songbird. Neuroscience 198: 152–170, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fee MS, Leonardo A. Miniature motorized microdrive and commutator system for chronic neural recording in small animals. J Neurosci Method 112: 83–94, 2001. [DOI] [PubMed] [Google Scholar]
- Freeze BS, Kravitz AV, Hammack N, Berke JD, Kreitzer AC. Control of basal ganglia output by direct and indirect pathway projection neurons. J Neurosci 33: 18531–18539, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Georgopoulos AP, DeLong MR, Crutcher MD. Relations between parameters of step-tracking movements and single cell discharge in the globus pallidus and subthalamic nucleus of the behaving monkey. J Neurosci 3: 1586–1598, 1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gittis AH, Kreitzer AC. Striatal microcircuitry and movement disorders. Trends Neurosci 35: 557–564, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gittis AH, Nelson AB, Thwin MT, Palop JJ, Kreitzer AC. Distinct roles of GABAergic interneurons in the regulation of striatal output pathways. J Neurosci 30: 2223–2234, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg JH, Adler A, Bergman H, Fee MS. Singing-related neural activity distinguishes two putative pallidal cell types in the songbird basal ganglia: comparison to the primate internal and external pallidal segments. J Neurosci 30: 7088–7098, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg JH, Farries MA, Fee MS. Integration of cortical and pallidal inputs in the basal ganglia-recipient thalamus of singing birds. J Neurophysiol 108: 1403–1429, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg JH, Fee MS. A cortical motor nucleus drives the basal ganglia-recipient thalamus in singing birds. Nat Neurosci 15: 620–627, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg JH, Fee MS. Singing-related neural activity distinguishes four classes of putative striatal neurons in the songbird basal ganglia. J Neurophysiol 103: 2002–2014, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldberg JH, Fee MS. Vocal babbling in songbirds requires the basal ganglia-recipient motor thalamus but not the basal ganglia. J Neurophysiol 105: 2729–2739, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graybiel AM, Aosaki T, Flaherty AW, Kimura M. The basal ganglia and adaptive motor control. Science 265: 1826–1831, 1994. [DOI] [PubMed] [Google Scholar]
- Gulley JM, Kuwajima M, Mayhill E, Rebec GV. Behavior-related changes in the activity of substantia nigra pars reticulata neurons in freely moving rats. Brain Res 845: 68–76, 1999. [DOI] [PubMed] [Google Scholar]
- Hessler NA, Doupe AJ. Singing-related neural activity in a dorsal forebrain-basal ganglia circuit of adult zebra finches. J Neurosci 19: 10461–10481, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hikosaka O. GABAergic output of the basal ganglia. Prog Brain Res 160: 209–226, 2007. [DOI] [PubMed] [Google Scholar]
- Hikosaka O, Wurtz RH. The basal ganglia. Rev Oculomot Res 3: 257–281, 1989. [PubMed] [Google Scholar]
- Iwasaki M, Poulsen TM, Oka K, Hessler NA. Singing-related activity in anterior forebrain of male zebra finches reflects courtship motivation for target females. PLos One 8: e81725, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaeger D, Kita H. Functional connectivity and integrative properties of globus pallidus neurons. Neuroscience 198: 44–53, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joshua M, Adler A, Rosin B, Vaadia E, Bergman H. Encoding of probabilistic rewarding and aversive events by pallidal and nigral neurons. J Neurophysiol 101: 758–772, 2009. [DOI] [PubMed] [Google Scholar]
- Kubikova L, Wada K, Jarvis ED. Dopamine receptors in a songbird brain. J Comp Neurol 518: 741–769, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kunimatsu J, Tanaka M. Roles of the primate motor thalamus in the generation of antisaccades. J Neurosci 30: 5108–5117, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipkind D, Marcus GF, Bemis DK, Sasahara K, Jacoby N, Takahasi M, Suzuki K, Feher O, Ravbar P, Okanoya K, Tchernichovski O. Stepwise acquisition of vocal combinatorial capacity in songbirds and human infants. Nature 498: 104–108, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meyer-Luehmann M, Thompson JF, Berridge KC, Aldridge JW. Substantia nigra pars reticulata neurons code initiation of a serial pattern: implications for natural action sequences and sequential disorders. Eur J Neurosci 16: 1599–1608, 2002. [DOI] [PubMed] [Google Scholar]
- Nambu A, Tokuno H, Takada M. Functional significance of the cortico-subthalamo-pallidal “hyperdirect” pathway. Neurosci Res 43: 111–117, 2002. [DOI] [PubMed] [Google Scholar]
- Nambu A, Yoshida S, Jinnai K. Discharge patterns of pallidal neurons with input from various cortical areas during movement in the monkey. Brain Res 519: 183–191, 1990. [DOI] [PubMed] [Google Scholar]
- Oller DK. The emergence of the sounds of speech in infancy. In: Child Phonology, edited by Yeni-Komshian GH, Kavanaugh JF, and Ferguson CA. New York: Academic, 1980, p. 93–112. [Google Scholar]
- Olveczky BP, Andalman AS, Fee MS. Vocal experimentation in the juvenile songbird requires a basal ganglia circuit. PLoS Biol 3: e153, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Penney JB Jr, Young AB. Speculations on the functional anatomy of basal ganglia disorders. Annu Rev Neurosci 6: 73–94, 1983. [DOI] [PubMed] [Google Scholar]
- Perkel DJ, Farries MA, Luo M, Ding L. Electrophysiological analysis of a songbird basal ganglia circuit essential for vocal plasticity. Brain Res Bull 57: 529–532, 2002. [DOI] [PubMed] [Google Scholar]
- Saar S, Mitra PP. A technique for characterizing the development of rhythms in bird song. PLos One 3: e1461, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scharff C, Nottebohm F. A comparative study of the behavioral deficits following lesions of various parts of the zebra finch song system: implications for vocal learning. J Neurosci 11: 2896–2913, 1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt R, Leventhal DK, Mallet N, Chen F, Berke JD. Canceling actions involves a race between basal ganglia pathways. Nat Neurosci 16: 1118–1124, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheth SA, Abuelem T, Gale JT, Eskandar EN. Basal ganglia neurons dynamically facilitate exploration during associative learning. J Neurosci 31: 4878–4885, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stephenson-Jones M, Ericsson J, Robertson B, Grillner S. Evolution of the basal ganglia: dual output pathways conserved throughout vertebrate phylogeny. J Comp Neurol 520: 2957–2973, 2012. [DOI] [PubMed] [Google Scholar]
- Sutton RS, Barto AG. Reinforcement learning: an introduction. IEEE Trans Neural Netw 9: 1054, 1998. [Google Scholar]
- Szydlowski SN, Pollak Dorocic I, Planert H, Carlen M, Meletis K, Silberberg G. Target selectivity of feedforward inhibition by striatal fast-spiking interneurons. J Neurosci 33: 1678–1683, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tachibana Y, Kita H, Chiken S, Takada M, Nambu A. Motor cortical control of internal pallidal activity through glutamatergic and GABAergic inputs in awake monkeys. Eur J Neurosci 27: 238–253, 2008. [DOI] [PubMed] [Google Scholar]
- Tchernichovski O, Mitra PP, Lints T, Nottebohm F. Dynamics of the vocal imitation process: how a zebra finch learns its song. Science 291: 2564–2569, 2001. [DOI] [PubMed] [Google Scholar]
- Turner RS, Anderson ME. Context-dependent modulation of movement-related discharge in the primate globus pallidus. J Neurosci 25: 2965–2976, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner RS, Anderson ME. Pallidal discharge related to the kinematics of reaching movements in two dimensions. J Neurophysiol 77: 1051–1074, 1997. [DOI] [PubMed] [Google Scholar]
- Veit L, Aronov D, Fee MS. Learning to breathe and sing: development of respiratory-vocal coordination in young songbirds. J Neurophysiol 106: 1747–1765, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolley SC, Rajan R, Joshua M, Doupe AJ. Emergence of context-dependent variability across a basal ganglia network. Neuron 82: 208–223, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida A, Tanaka M. Enhanced modulation of neuronal activity during antisaccades in the primate globus pallidus. Cereb Cortex 19: 206–217, 2009. [DOI] [PubMed] [Google Scholar]