

Abstract
Adjuvants improve the adaptive immune response to a vaccine antigen by modulating innate immunity or facilitating transport and presentation. The selection of an appropriate adjuvant has become vital as new vaccines trend toward narrower composition, expanded application, and improved safety. Functionally, adjuvants act directly or indirectly on antigen presenting cells (APCs) including dendritic cells (DCs) and are perceived as having molecular patterns associated either with pathogen invasion or endogenous cell damage (known as pathogen associated molecular patterns [PAMPs] and damage associated molecular patterns [DAMPs]), thereby initiating sensing and response pathways. PAMP-type adjuvants are ligands for toll-like receptors (TLRs) and can directly affect DCs to alter the strength, potency, speed, duration, bias, breadth, and scope of adaptive immunity. DAMP-type adjuvants signal via proinflammatory pathways and promote immune cell infiltration, antigen presentation, and effector cell maturation. This class of adjuvants includes mineral salts, oil emulsions, nanoparticles, and polyelectrolytes and comprises colloids and molecular assemblies exhibiting complex, heterogeneous structures. Today innovation in adjuvant technology is driven by rapidly expanding knowledge in immunology, cross-fertilization from other areas including systems biology and materials sciences, and regulatory requirements for quality, safety, efficacy and understanding as part of the vaccine product. Standardizations will aid efforts to better define and compare the structure, function and safety of adjuvants. This article briefly surveys the genesis of adjuvant technology and then re-examines polyionic macromolecules and polyelectrolyte materials, adjuvants currently not known to employ TLR. Specific updates are provided for aluminum-based formulations and polyelectrolytes as examples of improvements to the oldest and emerging classes of vaccine adjuvants in use.
Keywords: Immunologic adjuvant, Innate immunity, Toll like receptors, Pattern recognition receptors, Nalp3 protein, Alum, Aluminum hydroxide, Polymers, Chitosan, Polyphosphazene, Polyoxidonium
Introduction
Vaccination is among humanity's greatest achievements for reducing death, disability, and inequity caused by infectious diseases. The net benefit to health and well-being afforded by vaccination has been triumphant across nearly 300 years of use. A recent analysis of public records in the United States concluded that vaccines prevented 103.1 million cases of communicable and environmental disease over the past 90 years [1]. This accomplishment supported another benefit, namely, the increase in life expectancy of 30.4 years over the same time period [2]. Today, the global mortality prevention rate is about 2.5 million lives saved worldwide each year by immunization programs. The World Health Organization (WHO) immunization program includes the original six childhood vaccines-against diphtheria, tetanus, pertussis (whooping cough), measles, polio, and tuberculosis-plus more recently introduced vaccines against hepatitis B virus, Haemophilus influenzae type b, mumps, pneumococcal disease, rotavirus and rubella (German measles) [3]. Regional, national, and local or high-risk occupational immunization programs have added vaccines for varicella (chickenpox), meningococcal, typhoid fever, herpes zoster (shingles), human papillomavirus, yellow fever, Japanese encephalitis, as well as a list of investigational status biodefense vaccines [3,4,5]. Due in large part to worldwide vaccination programs, the global annualized rate of under-five-year-old deaths was reduced in half, from 12.6 million in 1990 to 6.6 million in 2012 [6]. Nevertheless, such reports also reveal that low income families represented 13 times the average case rate, immunization coverage is incomplete in many regions, vaccine stocks shortages are reported, and case rates have recently been increasing for some targeted diseases [6]. Beyond the remarkable impact on mortality rates, vaccines have also helped raise life quality, work productivity, and social equity by averting untold numbers of disabling post-infection disease sequelae, most notably blindness, deafness, neurological disorders [7,8].
Citing a list of accomplishments-the global eradication of smallpox in 1979 [9,10,11,12], an active endgame strategic plan for the eradication of polio [3,13,14], regional initiatives to eliminate or control several remaining vaccine-preventable childhood diseases, plus robust international post-marketing surveillance programs and periodic benefit-to-cost assessments [15,16,17,18,19,20,21,22,23]-the value of prophylactic vaccines against infectious diseases remains undisputed in the scientific, medical, and public policy communities as well as the general public at large [9,10,24,25,26,27]. However, significant advances in knowledge and technology are now helping to rapidly expand product development goals for immunization beyond the traditional and proven capabilities of vaccinology. Challenging new targets include vaccines for recent and ancient pathogens (human immunodeficiency virus [HIV], pandemic influenza, and malaria), improved capabilities to extend immunity to the very young, elderly, and immune compromised populations, for post-exposure prophylaxis indications in preparedness against biological threat agents (anthrax and Ebola virus) and emerging pandemic strains (H5N1 influenza, Middle East respiratory syndrome coronavirus, and methicillin-resistant Staphylococcus aureus), as well as immune treatments and protections against important non-infectious targets including cancers, autoimmune diseases, neurological disorders, and other medical conditions.
Adjuvants and the broader phenomenon of adjuvantation have been partner to vaccines successes throughout this history. The proportion of vaccines formulated with added adjuvant has increased over time and now comprises about half of the vaccines either licensed or in clinical testing (Fig. 1). A more detailed understanding of adjuvants has only recently appeared with the discovery and deepening characterization of immune signaling by biomolecules and physicochemical structures derived from pathogens or related to cell damage, now referred to as pathogen associated molecular patterns (PAMPs) and damage associated molecular patterns (DAMPs). This growth in understanding recently includes contributions from structural vaccinology and synthetic biology. Initially added to vaccines through empirical discovery, adjuvants are presently developed through more deductive ("rational") design and testing. The informed incorporation of adjuvant materials will continue to be an essential component of forthcoming vaccines, particularly as vaccines are engineered to meet rising global demand for greater potency using less antigen, wider disease and population coverage in fewer formulations, easier administration, and reduced logistical costs.
Fig. 1.

Proportion of human vaccines containing adjuvant through stages of history. Circles depict periods in vaccine development with fractional amount containing adjuvant shaded in blue, and diameter proportional to the log of number of different vaccines. A, up to 1899; B, 1900 to 1949; C, 1950 to 2012; D, 2014 U.S. licensed as listed by Food and Development Administration (FDA); E, 2014 in clinical testing as listed by HuVax (http://www.violinet.org). Note that the licensed vaccines group D contains many more multiple products for similar or overlapping indications, most of which are non-adjuvanted, while the experimental group E includes all existing and new candidate adjuvants reported in clinical testing.
Here we first look back at the origin of adjuvants, and then focus on a few classical and recent adjuvants as examples of the category of ionic polymers and ionic materials. Recent advances in immunology and materials science have improved the understanding and capability of aluminum salts which remain the most widely used adjuvant, and of synthetic polymers which represent a newer material showing wide ability and promise.
A Synopsis of Adjuvant History
Adjuvant technology emerged from medical necessity, regulatory pressures, and scientific discovery. Recounted here are a few of the seminal events leading to the development and use of adjuvants as a medical tool that continues to evolve in capability and importance, now more rapidly than ever before. This history shows that empirical observation and testing were critical to the discovery and development of the first immunologic adjuvants.
Recognition of a need for supplemental processing or formulation to improve immunization arose with the expansion of vaccine design beyond the attenuated and inactivated or killed strain preparations used since the dawn of vaccinology [28,29]. In the early 20th century, difficulties accompanied the initial toxoid type vaccines which comprised dilute active toxin administered with antitoxin (horse anti-serum). Discovered forty years earlier by Kitasato and Von Behring, for which Von Behring later won the first Nobel Prize in medicine [30], these toxin/serum type vaccines were marketed boldly and were desired for the protection they afforded against diphtheria and tetanus, commonly fatal childhood diseases. Unlike most other endemic and epidemic diseases awaiting effective vaccines, the bacterial exotoxin proteins associated with these diseases reproduced disease pathologies in isolation of and apart from the etiological, live infectious organisms. The activity of exotoxins could induce neutralizing antibodies, as demonstrated first in animal testing and later by passive transfer in humans, and small doses could induce immunity if co-administration with protective antiserum [29,31,32]. Diphtheria was a leading cause of illness and death among children then, estimated at 200,000 cases and 16,000 deaths annually, and various toxin/antitoxin products afforded prophylaxis. However, in 1901 tetanus-contaminated diphtheria antiserum caused the death of 13 children in St. Louis, and soon afterward 9 children died in New Jersey from tetanus contaminated smallpox vaccine [33,34]. These tragedies underscored a need for regulation, standardization, and quality control and drove legislative action for public safety. The Biologics Control Act of 1902 launched federal requirements for annual licensing and inspections of biologics manufacturers, rules for product testing, purity standards and quality control, truth in labeling, and also created the Hygienic Laboratory of the U.S. Public Health Service which eventually became the National Institutes of Health [34]. Subsequent acts and amendments added and consolidated regulations, then eventually transferred oversight of biologics to the Food and Drug Administration (FDA) in 1972. Thus, the events of 1902 were groundbreaking as "the first enduring scheme of national regulation for any pharmaceutical product...the very first premarket approval statute in history," and set precedent for "shifting from retrospective post-market to prospective pre-market government review" [35]. This general period and area of research also yielded the first biological standard, established by Paul Ehrlich in working with Von Behring's antiserum to control for variations in content and temperature in proposing universal units of activity.
The genesis of immunologic adjuvants emerged within the same setting of medical and regulatory pressure surrounding toxoid vaccines and antisera, though detailed knowledge of the underlying science about how adjuvants enhance immunity would not coalesce until the 1990s. A series of discoveries which focused on reducing the toxicity of diphtheria toxin while improving its immunity as an antigen illustrate the inception of adjuvant science and methodology:
1921: Inactivated toxin (toxoid) is immunogenic and protective against toxin [29].
1923: Bread crumbs, tapioca starch, agar, oil or other compounds increased antiserum titer and slowed the release and elimination of toxoid [36].
1927: Fractional purification of toxoid by precipitation with aluminum salts provided robust immunization and protection in guinea pig [37].
1932: Alum greatly enhanced the value of diphtheria toxoid immunization in humans [38].
1933: A single dose of alum-precipitated toxoid afforded protective immunity in humans, assessed by the Director of the National Institutes of Health (NIH) as being much more effective and tolerable compared to toxoid alone [39].
1946: Aluminum phosphate improves precipitation of diphtheria toxoid [40].
1947: Pre-formed aluminum acts as an adsorbent and functions as a robust adjuvant [41].
The last achievement listed, on the effectiveness of pre-formed aluminum salts compared to co-precipitation of antigen with aluminum phosphate or alum (potassium aluminum sulfate) before that, connects this line of development to modern aluminum gels which are mostly employed today. Often considered together, research has shown that the various aluminum salt gels do not exhibit universally equivalent attributes as adjuvant materials. The full chemical formula and process formulation of aluminum salt adjuvants for some licensed vaccines are not publically released. The discovery and evolution of aluminum salt adjuvants surveyed here is just one thread of several adjuvants in a deep history of vaccine development, and illustrates that empirical findings were foundational both in general and specifically to the largest class of adjuvant still in use today. A depiction of the development of several licensed vaccines and their adjuvant is shown in Fig. 2. This illustration includes timelines for the common vaccine classes (attenuated or killed, serum or toxoid, conjugate, etc.) and a few technologies supporting new development of vaccines and adjuvants, as reprinted from Rappuoli et al. [2].
Fig. 2.

Timeline of vaccines and adjuvants development. Shown is an historical view of the origin and development for some early and currently licensed human vaccines, the time needed to translate a scientific discovery (shown in red) into a safe and scalable vaccine technology (orange bars), the evolution of adjuvants, and of some supporting areas of science and technology. The blue bars represent the time needed to develop and implement one or more vaccines derived from the same technology. IPV, inactivated polio virus; OPV, oral polio virus; Hib, Haemophilus influenzae type b; HBV, hepatitis B virus; HPV, human papillomavirus. Reproduced from Rappuoli et al. (2014), with permission of the author and publisher, Proceedings of the National Academy of Sciences [2].
The only other adjuvant classes used in currently licensed vaccines include emulsions systems, liposomes and toll-like receptor (TLR)-based compounds. As with aluminum salts, their discoveries have been critical to the general development of adjuvant science and technology. Their introduction as modern version adjuvants is also noted in the adjuvants timeline of Fig. 2. Currently licensed emulsion systems and liposome type adjuvant formulations used in vaccines for seasonal flu, pandemic influenza, and hepatitis A [42] trace back to Freund's complete adjuvant (1951) [43]. Complete Freund's adjuvant (CFA) is an emulsion system of soluble antigen mixed into mineral oil (water-in-oil [w/o]) with surfactant and inactivated dried mycobacteria. Though CFA became popular because of its potency [43], it was deemed too toxic for human use as evidenced by severe inflammatory effects and granulomatous reactions [44]. Reversed emulsion formulations containing an oil-in-water (o/w) mixture without mycobacterium, including incomplete Freund's adjuvant (IFA), were less viscous so easier to prepare and inject as well as less reactogenic [28,45]. Squalene, a biodegradable oil that is still extracted from shark liver, emerged as part of effective o/w emulsion adjuvants used in few licensed vaccine formulations today including MF59 (squalene, Tween-80, and sorbitan trioleate) [46], AS03 (squalene, Tween-80, and α-tocopherol) [47], as well as AF03, a candidate pandemic flu vaccine not yet licensed [48]. Several candidate emulsion and liposome adjuvants are also being evaluated in preclinical and clinical testing and are not yet licensed, including the GlaxoSmithKline "adjuvant system" formulations AS01 (liposome, 3-O-desacyl-4'-monophosphoryl lipid A [MPL], and saponin) and AS02 (o/w, MPL, and saponin). No TLR or other pattern recognition receptor (PRR) mechanism of action has been indicated for squalene [49] or other emulsion and liposome type adjuvants though these are being investigated as vehicles to combine with known TLR ligands.
The synthetic compound MPL is the only TLR class of adjuvant presently used in a US licensed vaccine. This is a detoxified version of the gram-negative bacterial endotoxin lipopolysaccharide (LPS) which signals innate immunity via the TLR4 complex. LPS was the first TLR ligand out of many PAMPs now intensively investigated for development as potential adjuvants with new capabilities to improve vaccines. Discovery of bacterial endotoxin in the late 1890s by Richard Pfieffer, and subsequent demonstrations that this heat stable substance could evoke neutralizing antibody which suppressed the poisonous effect [50] initiated a long line of research that underlies current knowledge of PAMPs and TLR-directed innate immune response. Among several other discoveries that also spurred the advance of immunologic adjuvants were medical reports of "Coley's toxins," extracts from infectious bacteria that successfully treated cancer for almost 50 years (1895-1944) [51]. The wide study of immune-stimulating bacterial components converged in the 1990s with discoveries and understanding that certain microbial substructures embody PAMPS which serve as ligands for immune cell PRRs (aka PRRs) that signal the innate immune system [52,53]. The identification of TLRs (10 in human), nod-like receptors (NLRs, 3 classes), and other PRRs has revolutionized the science and practice of vaccine adjuvant characterization and development [54,55,56,57,58,59,60,61,62].
It is evident from this synopsis that the early period of vaccine development was determinative both for the science of adjuvantation and for the standards of biologics regulation. Since then the variety of vaccine types has expanded, and adjuvants have co-developed with each vaccines type licensed or under investigation today with a few exceptions, principally whole organism vaccines and carbohydrate vaccines.
With a large body of cumulative knowledge, the nature of adjuvant development and testing has changed from one of empirical, trial-and-error testing toward one more akin to rational design [55,63], though much is left to be understood about adjuvant mechanism of action, and particularly for DAMP-type class of substances. Several fields of science intersect as drivers for the co-evolution of vaccine and adjuvant for improved immunization capability [54,55]. The largest single driver of innovation and improvements in adjuvant technology is now scientific knowledge and information, for the rapid growth in primary data and the improved, more accessible means of connecting and cross-referencing information. Due to the high interdependency and rapid pace of development, this shared endeavor will benefit from comprehensive, descriptive reporting and networking of data and results from testing that is designed and performed with standards then annotated and communicated with universal scientific meaning.
Adjuvant Roles in Immunization
Adjuvants improve vaccine effectiveness by modulating the immunogenicity of antigen delivered in a vaccine product. In addition to the original goal of increasing antibody titer, immunologic adjuvants are now being engineered to alter the natural adaptive immune response to an antigen for increased potency and duration, shortened time to immunity, altered immune polarization, increased scope and breadth for disease target and patient population (Table 1).
Table 1.
Immunological roles for adjuvants in vaccine development
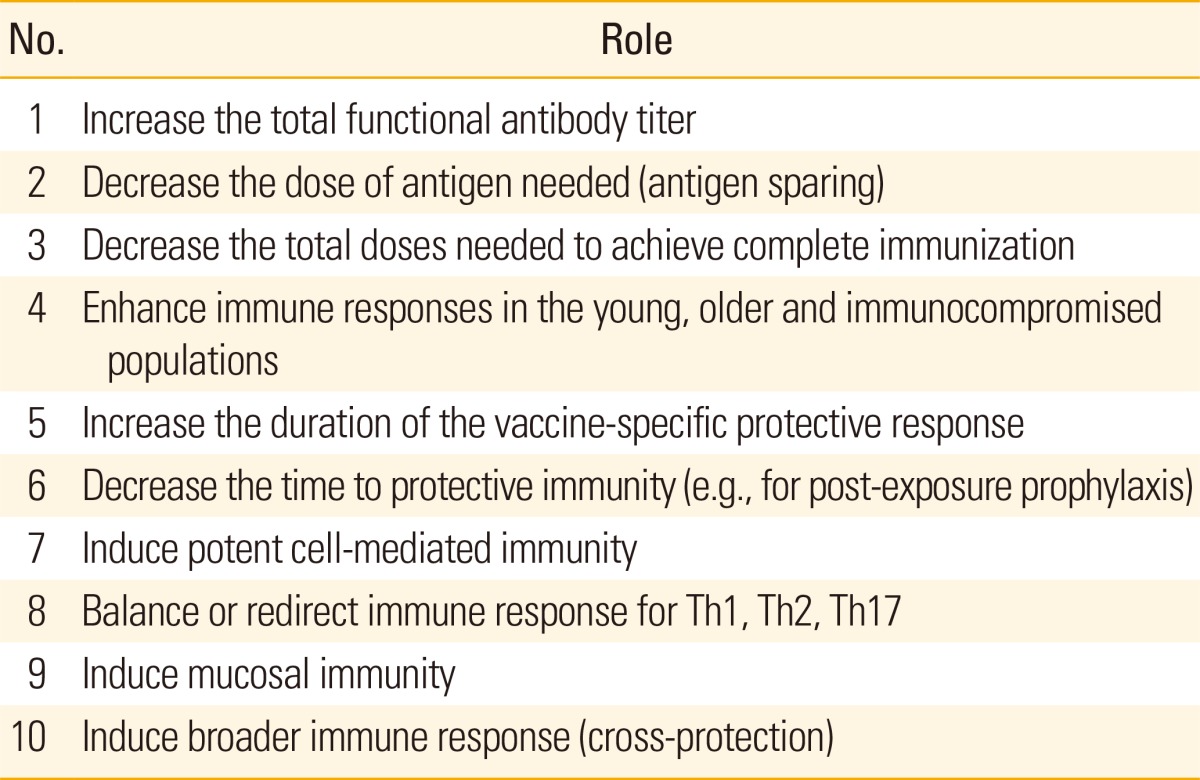
These goals are sought while also aligning with efforts to improve vaccine tolerability and safety, product manufacture and stability, as well as to aid the cost and global logistics of transport, distribution, stockpiling and administration of new vaccines, including to underdeveloped and challenging environments.
A few types of vaccine do not require adjuvant, though their total product number for licensed indications exceeds those of vaccines containing adjuvant (Fig. 1). These include the live attenuated vaccines (adenovirus, Bacillus Calmette-Guerin [BCG], flu, mumps and rubella vaccine, oral polio virus polio, rotavirus, smallpox, Salmonella Typhi Ty21a, varicella, and yellow fever), split virion vaccines (flu), some inactivated vaccines (rabies and inactivated poliovirus virus [IPV] polio), and carbohydrate vaccines (pneumococcal, meningococcal, and Typhium Vi). While the live attenuated, inactivated and split virion vaccines do not require adjuvant because they include sufficient PAMPs for self-adjuvantation, the carbohydrate vaccines do not require adjuvant because they are T-cell independent. Carbohydrate antigens such as capsular polysaccharide directly bind B-cells to elicit activation and proliferation without MHC-restricted T-cell help [64]. This response generally is of short duration with no affinity maturation and fails to elicit immunologic memory. For this reason, the newer carbohydrate-based vaccines consist of carbohydrates conjugated to protein which provide T-cell epitopes for a T-cell dependent response capable of antibody class switching, affinity maturation, immunologic memory, and longer duration. Although adjuvants are often added to conjugate vaccines to enhance the T-cell dependent response, some candidate conjugate proteins also appear to have the capacity to function simultaneously as true adjuvants through TLR interaction. For example, neisserial porins signal via a TLR1/TLR2 complex [65,66]. In addition, other proteins may provide TLR-based adjuvant effects when fused with a target protein antigen. As an example, salmonella flagellin interacts with TLR5 when genetically fused with influenza hemagglutinin [67].
How adjuvants promote immune stimulating effects is an active and growing area of research in which ongoing discoveries are helping to build areas of understanding in vaccine adjuvant immunology, as is briefly discussed next.
Adjuvant Mechanisms of Action
Today, information about the mechanisms of immune potentiation is accumulating quite rapidly, and mechanisms are frequently challenged or supported by experimental evidence. Consensus exits for several pathways involved in adjuvant mechanisms of action, the most detailed of which are for PRRs and the TRL class of signal receptors that helped revolutionize understandings of immune phenomenology [68,69]. In summary, TLRs1-6 located on the plasma membrane of immune cells (macrophages, dendritic cells [DC], and neutrophils, etc.) recognize PAMPs from the surface of bacterial and fungal pathogens such as LPS and flagellin, while endosomally located TLR3 and TLRs7-9 recognize viral and microbial RNA and DNA [70]. Several other PRRs are described, such as NLRs, RIG-1 like helicases, extracellular C-type lectin receptors, all of which function via adaptor proteins (such as MyD88, TIRAP, TRIF, and ASC, etc.) to coordinate signaling between and among PAMP pathways. PRRs including some TLRs (e.g., TLR2/4) also recognize endogenous DAMP ligands like heat shock proteins 60 and 70, fibrinogen, and fibronectin produced in cell damage, injury, stress, or metabolic imbalance [71,72]. On activation, TLRs in neutrophils can activate NADPH oxidase to produce reactive oxygen species (ROS) which mediate release of other DAMPs driving an inflammatory response [71,73,74]. In this way TLRs, NLRs and other PRRs transmit and help coordinate immunomodulatory responses signaled by pathogens and endogenous stressors [73,75]. The net immediate effect of responses involving the inflammasome is to activate the inflammatory process by promoting the processing and secretion of the proinflammatory cytokines interleukin (IL)-1β and IL-18. This initiates processes driving the recruitment, activation, and maturation of immune cells to direct innate and adaptive immune functions, processes at the heart of functional mechanisms of immunologic adjuvants. Excellent reviews have been published with great detail about mechanisms for TLR-dependent and TLR-independent adjuvants [44,54,55,61,64,68,69,70,75,76,77]. Many of these reviews provide generalized summary illustrations which are instructive in developing, challenging, and refining models toward an understanding for immune regulation and adjuvant function. Naturally, concepts and models occasionally require adjustment to adapt as knowledge evolves.
Current evidence does not yet connect the functioning of aluminum salt gels and polyelectrolyte adjuvants, topics of this review, to known TLR pathways. However, the activity of some TLR-independent adjuvant materials such as aluminum and oil emulsions are linked to DAMP pathways, including activation of the inflammasome by ROS [78] and deformation or re-structuring of the plasma membrane [79,80,81]. The NLRP3-inflammasome plays an important role, though it has also been shown not to be universally essential in DAMP-signaling by adjuvants. Interestingly, an NLRP3-oligemeric complex also acts as an extracellular DAMP particle itself that disperses to neighboring macrophages to amplify the inflammatory response [82]. Generalized roles for cells, pathways, and the NLRP3-inflammasome are depicted in Fig. 3, reproduced and modified slightly from De Gregorio et al. [68]. This illustration shows how some TLR and non-TLR adjuvant activities focus on the DC as the central controller of adaptive immunity.
Fig. 3.

A model of multiple mechanisms of adjuvanticity that converge toward dendritic cell (DC) activation. Adjuvants activate DCs either through direct interactions or through cellular intermediaries. toll-like receptor (TLR)-dependent adjuvants (in black) act directly on DCs; however, they can also activate other cells expressing the responding TLR. TLR-independent adjuvants (in red) can act directly either on DCs or on accessory cells. Aluminum salts, PLG, MSU, and Quil-A activate the NLRP3 inflammasome. Aluminum salts and MF59 act on monocytes, macrophages, or granulocytes to induce cytokines that generate a local immunostimulatory environment eventually leading to DC activation. In addition they also promote monocyte differentiation into DCs. MF59 can also activate muscle cells at injection site. It has been suggested that aluminum salts causes local necrosis of stromal cells leading to the release of uric acid, an endogenous danger signal that activates the inflammasome. Mast cell activators such as c48/80 can also act as adjuvants in a DC-dependent mechanism. iNKT activation by a-GalCer presented on CD1d leads to DC activation. Beta glucans activate DC through dectin-1 and ISCOMATRIX directly promotes cross-presentation in DCs [68]. So far, a role by TRLs has not been shown for the adjuvant action of polyelectrolytes. Reproduced from De Greggorio et al. (2009) with slight update and with permission of the author and publisher, Elsevier, Current Opinion in Immunology [68].
For many years, particulate and colloidal adjuvants such as aluminum salts and polymers were considered to function mainly as inert delivery vehicles, assisting in antigen stability, uptake and delivery. The original depot model for slow-release of antigen by aluminum adjuvants has been challenged [83] with experimental evidence that some antigens desorb from some aluminum adjuvants soon after injection [84]. Nevertheless, the carrier or transport role is not debunked. Antigen stability in both structure and potency can be greatly enhanced as bound to aluminum salt gels compared to their free, soluble form [85,86,87]. Importantly, recent work demonstrated that the simple inclusion of 4 mM phosphate with Alhydrogel (aluminum oxyhydroxide) saline formulation resulted in hydrated antigen and re-established soluble-like structure compared to formulation with plain Alhydrogel [85]. There have also been contrasting conclusions about whether aluminum salts adjuvants are internalized by antigen presenting cells [88,89,90]. Recent reports provide convincing evidence of internalization [89,91]. In particular, the fluorescent molecular probe for aluminum, lumogallion, was used to confirm intracellular location of aluminum oxyhydroxide adjuvants taken up by a monocytic T helper 1 cell line [91]. Thus, immune activation is supported both with and without internalization of aluminum adjuvant particles.
Adjuvant Classification
What began as a handful of materials to boost antigen titer 90 years ago has now become a short list of licensed vaccine adjuvants and a very long list of potential adjuvant materials under preclinical and clinical testing. Adjuvants are commonly divided into two broad functional categories as delivery vehicles or immunopoteniators, though many show both functions including the mineral salts and polyelectrolytes discussed below. Subcategories of licensed vaccine adjuvants include mineral salts (alum and aluminum hydroxide, etc.), oil-in-water emulsions (MF59 and AS03), liposomes (virosome), and a combination adjuvant (AS04) [42]. On the other hand, there are very many adjuvants and combination adjuvant formulations under exploration or development, including many in clinical testing. The Vaxjo web-based database tool for vaccine adjuvant informatics (http://www.violinet.org/vaxjo/adjuvant) lists 103 different vaccine adjuvant compounds [92,93]. The Huvax database, another vaccine informatics tool within the VIOLONET suite, lists 101 human vaccines or vaccine candidates containing adjuvant. The ClinicalTrials.gov registry database [94,95] lists 869 clinical trials globally that involve vaccine adjuvant. The number and proposed indications for vaccine adjuvants continues to expand as increasing knowledge and information enables rational justification for new combination products or testing a new potential ligand, receptor, or pathway [96]. The remainder of this review focusses on polyionic classes of adjuvants.
Updates on Polyionic Adjuvant Materials
Aluminum salt adjuvants
Aluminum salt gels represent the vast majority of mineral salt compounds used as vaccine adjuvants. Calcium phosphate is the only non-aluminum mineral salt adjuvant that has been used in human vaccines [97], though several other mineral hydroxides have also been tested systematically as model nanomaterials for measuring immune effects by in vitro and animal models as described below.
Aluminum salts have a long history of use and the most comprehensive safety record of all adjuvants incorporated into human vaccines, beginning with potassium alum in the diphtheria and tetanus vaccines of the 1930s. Aluminum salt adjuvants used today in human vaccines are all gels comprising an oxide, phosphate, or sulfate salt of aluminum. In the past, particularly on discovery and during the subsequent few decades, an aluminum oxide sulfate or phosphate was salted out together with antigen in creating toxoid preparation. While preformed gels are commonly used now because of convenience and arguable control in product quality, some proprietary formulations today may still employ in situ aluminum salt precipitation. The atomic and macromolecular structures and biological activities of aluminum salts was studied intensively in the Stanley Hem laboratory [77,98,99] and has seen a renascence with interest from the areas of nano/mesotechnology, material sciences, and the rapid advance of immunology as supported by systems biology, bioinformatics and new capabilities in cell biology, microscopy, recombinant and mouse genetics and labeling.
Cumulative experience has established that aluminum salt adjuvants enhance vaccines to induce high titer IgG with a relatively long lasting immunity, ease of formulation, and a long record of general safety [44,100]. They elicit strong Th2 immune polarization which has been effective in creating protective immunity against extracellular pathogens. However, aluminum-based adjuvants can also induce IgE antibodies and stimulate little or no cell-mediated Th1 or cytotoxic CD8 T-cell responses. If injected via subcutaneous or intradermal route, or administered repeatedly, their use has also been associated with the occurrence of granulomas [44,100]. Because of these effects, the use of aluminum salt adjuvants has adapted. A simple, effective and specific example of this was altering the route of injection for the anthrax vaccines which historically were injected subcutaneously and had a clearly associated record of adverse events [101]. In 2008, the advised route of administration changed from subcutaneous to intramuscular route for both the US and UK licensed anthrax adsorbed vaccines (BioThrax, Emergent Biosolutions Inc., Rockville, MD, USA; AVP, Public Health England, London, UK). More generally, combination adjuvant formulations have been developed to impart additional Th1 and cell mediated immunity. The AS03 and AS04 products, which combine aluminum hydroxyphosphate with other adjuvants to create Th1 immunity have been used in Europe for decades and recently received US licensing for use in the vaccines. AS03 is a squalene oil emulsion while AS04 contains MPL adsorbed on to aluminum hydroxide salt. Beyond these combinations, the aluminum salt compounds themselves have been investigated further both in counter ion formulations and structurally as defined nanomaterials and shown new capabilities [96,99]. The most common mineral salt adjuvants used in licensed human vaccines are described next.
Alum
"Alum" defines a heterogeneous class of hydrated aluminum double salts, composed of aluminum sulfate plus a cation, typically potassium, sodium, or ammonium. Today, alums used in vaccine formulations commonly refer to the potassium salt of aluminum sulfate, KAl2(SO4)2·12H2O. However, the salts of other triply charged metal cations may replace or be mixed with aluminum sulfate, including iron, magnesium, or cobalt, and can be present depending on the identity and purity of source materials. Potassium aluminum sulfate forms in situ on mixing with component salts, precipitating as K(SO4)·Al2(SO4)3·24H2O with antigen if included, and then dehydrates to KAl2(SO4)2·12H2O on processing during preparation. Unfortunately, the terms "alum" and "Al(OH)3" have been frequently interchanged with aluminum hydroxide and other aluminum salts. Generic, lenient use of either "alum" or "AL(OH)3" should be avoided as it propagates confusion. Notably, inaccurate use of the term "alum" confounds efforts to compare vaccine adjuvant safety and efficacy between different vaccine formulations. This precaution is becoming more important as aluminum salts are used as benchmark for the testing and development of new adjuvants including aluminum salt gels of specific size and surface chemistry.
Aluminum oxyhydroxide
The "aluminum hydroxide" gels used in vaccines is chemically defined as aluminum oxyhydroxide, AlO(OH)·nH2O. Alhydrogel (BRENNTAG Biosector, Frederikssund, Denmark) is a GMP example of this adjuvant subclass. Aluminum hydroxide gels are described as poorly crystalline and amorphous with a pseudo-boehmite mineral structure [98], and comprise a mixture of micron- and nano-sized particles, formed from aggregates of primary crystals up to 10 nm in length. Aluminum oxyhydroxide has a very high surface area (500 m2/g), and only the outer layer binds antigen. Binding is driven principally by electrostatic forces though other interactions occur including hydrogen-bonding and hydrophobic bonding. The point of zero charge (PZC) is 11.4, creating a positively charged surface at neutral pH to which antigens bind principally via their negative charges. However, the PZC and binding of antigen can be altered to substantial effect in both potency and stability by formulation with specific concentration of phosphate counter ion, as is summarized below [85]. The ability to simply alter antigen binding affinity, stability and potency via ligand exchange of the surface hydroxyls for phosphate, without altering the antigen, provides an inexpensive and rapid route to improving the capability of this adjuvant for other protein antigens [85]. As with other aluminum salt gels, the protein/aluminum bond tightens over prolonged storage, though this effect is also partially mitigated by formulation with 4 mM phosphate [85].
Aluminum phosphate
"Aluminum phosphate" used in vaccines are gels made of aluminum hydroxyphosphate, Al(OH)x(PO4)y. Adju-Phos (BRENNTAG) is a GMP example of this adjuvant subclass. Aluminum hydroxyphosphate has an amorphous mineral structure, with planar particles of 3 mm average diameter. The specific ratio of surface phosphate to hydroxyl depends on manufacturing conditions. With PZC ranging 4.5 to 5.5, the surface charge is positive at neutral pH in contrast to unmodified aluminum oxyhydroxide. Aluminum hydroxyphosphate dissolves more readily than does aluminum oxyhydroxide following injection, releasing about three times as much radiolabeled aluminum over a month as measured in rabbits [102].
Calcium phosphate
Commercial calcium phosphate used in vaccines is a non-stoichiometric hydroxyapatite. Although the name implies a formula of Ca3(PO4)2, study has identified the structure as Ca10x·(HPO4)x·(PO4)6x·(OH)2x, where x varies from 0 to 2 [103]. Therefore, these aluminum salts are not equivalent in surface chemistry or particulate size, and should not be considered as interchangeable adjuvant materials in direct testing or as reference for comparison to other adjuvants. Studies have established involvement of the NLRP3 inflammasome for adjuvant activity by aluminum salts [104,105], and that the role may not be not essential [83]. There is evidence for roles by other signaling pathways [79,88,89,91,102]. Importantly, it has been reported that aluminum salt adjuvants interact directly with DCs by binding plasma membrane lipids with substantial force [79]. Surface binding induces lipid sorting that involves aggregation of immunoreceptor tyrosine-based activation motif signaling-containing receptors which cascades to phosphoinositide 3 kinase-mediated responses, resulting in high affinity, stable binding to CD4+ T cells in the absence of further adjuvant interaction [79]. This is similar to the DAMP cascade caused by monosodium urate crystals, an original candidate model for aluminum mode of action, and helps to explain conflicting results for urate crystals and aluminum salt adjuvants. Evidence supports both models by which aluminum adjuvant is internalized [91] and by which it does not enter the cell but instead delivers admixed antigen for passage across the membrane [90].
Recent studies on aluminum and other model particulate materials have revealed reproducible effects of material size and surface chemistry on cell responses using aluminum salts and metal oxide materials and systems immunology to investigate immune signaling by in vitro and in vivo studies which also explains part of their Th2 immune polarization [79,106,107,108,109]. Particle size is shown to be an important parameter for vaccine adjuvants, and the nanometer size range of 150-200 nm is particularly active, which may be due to due to cell presentation and effects on cellular uptake or membrane perturbation [79]. Larger particles exert different effects on innate immune signaling and activation of inflammasome [110]. Crystalline particles exceeding a 10 nm threshold are not fully engulfed by macrophages in vitro and create a "frustrated phagocytosis" effect that activates proinflammatory responses whose chronic stimulation in the lung may lead to medical complications including fibrosis and cancer. Studies using silver nanowires confirmed the effect in vivo implying that chronic exposure may contribute to the toxicity of fibrous materials such as asbestos [106]. Of relevance to adjuvant immune signaling by particulate adjuvants including aluminum salts is their finding of a gradation in effect in vivo, with full phagocytosis of materials <3 nm giving no proinflammatory signaling, yet complete phagocytosis and proinflammatory signaling by material sizes ≥5 mm. Thus, specific size of a relatively inert crystalline material will signal innate immunity without antigen, and the effect can vary from no proinflammatory response, to functional adjuvanticity, to frustrated phagocytosis. These findings can be relevant to aluminum salt gels which are partly crystalline and populated in the same range of sizes. Thus, it is becoming evident that physical dimension may be a useful attribute to measure and control for an aluminum-based adjuvant materials. Studies have also demonstrated strong effect by surface chemistry of aluminum salts adjuvants on their immune potentiating activities [107]. In particular, the surface density of hydroxyl groups correlate positively with proinflammatory responses [108].
This is in agreement with prior findings regarding the effects of phosphate counter ion on antigen binding and biological function of the commercially available aluminum oxyhydroxide, Alhydrogel. Watkinson et al. [85] used recombinant anthrax protective antigen (rPA) as a model antigen to investigate aluminum structure-function effects because of its dependency on adjuvant, a track record of aluminum oxyhydroxide use, and a history of differing potencies and stabilities by various producers. Through systematic investigation of phosphate binding on thermodynamics and kinetics of Alhydrogel/rPA interaction, phosphate at 4 mM was shown to convert thermodynamic control of binding of antigen from endothermic to exothermic and restore the thermostability of bound antigen so that it resembled that of the soluble form [85,87]. This was due to altered binding created via lignd exchange of phosphate for surface hydroxyls, with optimum effect below 10 mM concentration [85]. The 4 mM phosphate Alhydrogel formulation increased potency more than 5-fold as compared to the 0.25 mM phosphate concentration of conventional phosphate buffered saline formulations of rPA on Alhydrogel. Through stability trending analysis, using pooled mouse potency data from three batches of 4 mM phosphate formulated vaccine compared to a freshly formulated standard, the phosphate formulated vaccine showed an estimated 53 months shelf life at 4℃ based on its 95% confidence interval crossing a relative potency lower limit of 2 standard deviations. Thus, a substantial improvement in potency and stability was engineered into a conventional Alhydrogel vaccine simply using a defined, small change in phosphate counter ion concentration. Alternatively, improved efficacy afforded by phosphate-based binding to aluminum salt adjuvant may also be achieved by re-engineering the antigen with covalent phosphonate linker [111].
These studies establish that surface chemistry, particle size, and structure are all important attributes of aluminum gels [107] which should be measured and controlled. A commonly-known example of this is that autoclave treatment changes the physicochemical structure of aluminum gels and affects their adjuvant properties [112]. Since all current commercial aluminum based gels are heterogeneous in size as well as chemical composition to some degree, their specific effects as adjuvants cannot be expected a priori to be replicated across chemical class, manufacturer, or lot and require experimental confirmation. Thus, content and size matter in leveraging the capability, familiarity and safety of aluminum salt gels by mixing with other adjuvants for extended vaccine capability. Controls therein will aid further comparative study and improvement of this oldest and most frequently used class of adjuvants.
Ionic Polymer Adjuvants
It has been generally known for at least few decades that polyelectrolytes-ionic macromolecules of either synthetic or natural origin-serve as immunostimulants when introduced as mixtures with typical antigens, thereby enhancing immune responses by several fold [113]. Moreover, this effect is increased by orders of magnitude if the antigen is chemically bound to synthetic polyelectrolytes [113]. It was suggested that membrane activity and immunostimulation are inherent to polyelectrolytes with different chemical structures of the repeating units as a class and depend critically on molecular size, self-assembly, and other characteristics of the system. Although, there are examples of licensed products and clinical use of such materials [113,114], this family of polymers is generally reviewed less frequently in immunoadjuvants literature. This may have been partly due to challenges related to isolation, purification, and characterization of natural and some synthetic polyelectrolytes In addition, their ability to self-assemble in aqueous solutions under certain conditions can often lead to poor reproducibility, conflicting results, and difficulties in interpretation. Nevertheless, the synthetic and purification approaches to this class of polymers are greatly improving and the body of literature related to these compounds is rapidly growing. Natural and synthetic polyelectrolytes possess a number of common features and parameters affecting their biological function in solution. A few important parameters are: length and conformation of the polymer chains, only a few types of repeating units, flexibility of the backbone, ionic density, tacticity (regularity of repeating units), solvent interaction parameters, and hydrodynamic behavior. Thus, in summarizing reports related to immunoadjuvant properties of these polymers, it was important to review literature related to both natural and synthetic types of these long-chain, polyionic macromolecules.
Chitosan
Chitin and its derivative, chitosan, are linear polysaccharides which having increased attention as biomaterials, drug delivery carriers and immunomodulators [115,116,117]. Chitin is a long-chain polymer of a N-acetyl-D-glucosamine. While it is one of the most abundant polysaccharides in nature, its life sciences applications are somewhat limited due to insolubility in water. In contrast, chitosan, which is produced by partial deacetylation of chitin, is a copolymer consisting of randomly distributed ionizable D-glucosamine (deacetylated unit) and N-acetyl-D-glucosamine (acetylated unit) (Fig. 4), and consequently is more amendable for water-soluble and hydrogel nanoparticulate formulations. Moreover, cationic derivatives of chitosan provide improved water-solubility and have been also widely tested [118].
Fig. 4.
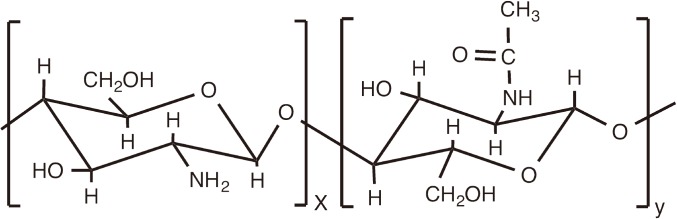
Schematic chemical structure of chitosan.
There are multiple reports that chitin stimulates macrophages to produce cytokines that confer anti-tumor activity and non-specific host resistance against bacterial and viral infections [119]. It is conceivable that chitin could be a potent innate immune stimulator of macrophages and other innate immune cells. There are reports that it stimulates macrophages by interacting with different cell surface receptors such as macrophage mannose receptor, TLR-2, C-type lectin receptor Dectin-1, and leukotriene B4 receptor (BLT1) [120]. It must be emphasized that that isolated chitin presents two difficulties, both related to its water-insolubility. Due to this insolubility, the chemical structure of chitin cannot be defined under physiological conditions, and may involve concurrent destruction. Formulations have always been evaluated as particulates and the above mentioned host responses vary dramatically depending on its size and possibly the nature of the chitin employed as well as the methods of preparation, timing, and route of administration [121].
Contrary to chitin, chitosan formulations are prepared by hydrolysis and are either water-soluble mixed polymers or hydrogel nanoparticulates, therefore more amenable to characterization than chitin. Unless the polymer was additionally modified, the water-solubility of chitosan is typically limited to slightly acidic to acidic conditions. Unfortunately, upon review of the literature, it is not always possible to elucidate which physical or even chemical form of chitosan was evaluated. The assessment of activity then become further complicated as various routes of administrations are used and chitosan was shown to be mucoadhesive, can promote prolonged retention times following mucosal vaccination, and additionally acts as a permeation enhancer by opening tight junctions [116]. However, there have been a number of studies reported recently focusing on the evaluation of well-characterized original materials, but not necessarily detailing the formulation itself.
However, one well defined chitosan, when tested for parenteral immunization in C57BL/6 mice using β-galactosidase as a model antigen, was shown to enhance antigen specific antibody titers over five-fold and antigen-specific splenic CD4+ proliferation over six-fold [122]. According to the authors, strong increases in antibody titers together with robust delayed-type hypersensitivity responses revealed that chitosan induced a mixed Th1/Th2 humoral and cell-mediated immune responses. By this study, chitosan was demonstrated to be equipotent to IFA and superior to Imject Alum (aluminum hydroxide + aluminum magnesium hydroxide). Mechanistic studies revealed that this viscous chitosan solution created an antigen depot when injected subcutaneously and it also induced a transient 67% cellular expansion in draining lymph nodes [122]. It was also shown that chitosan can synergistically enhance the immunoadjuvant properties of granulocyte-macrophage colony-stimulating factor, a pleiotropic, proinflammatory cytokine that was initially discovered as a growth factor capable of generating granulocyte and macrophage colonies from bone marrow precursor cells [123].
In a separate study, evaluation of three different well-defined types of chitosan for parenteral administration of model antigen, ovalbumin, in mice showed adjuvant activity for all of them, in some cases exceeding that of "alum" (Alu-S-Gel aluminum hydroxide sensitive to phosphate) [124]. However, the adjuvant activity varied significantly depending on the formulation. Authors conclude that this was probably a result of interplay between molecular weight, particle size, preparation technique, solubility and viscosity, as well as degree of deacetylation, whereas impurities were found to play a minor role. Nevertheless, authors also note that potential activation of PRR on cells of the innate immune system, which was reported for some chitin derivatives, can be potentially explained by the presence of bacterial and fungal contaminants and endotoxins since these materials are of natural origin and their ionic nature can complicate the purification efforts [124], as already noted.
These observations are mirrored in a recently published review, which emphasizes that in the majority of publications on the adjuvant activity of chitosan to date, the molecular weight, viscosity, deacetylation degree and purity level, especially endotoxins, are not provided for the chitosan under investigation and final formulation and preparation procedures are not sufficiently detailed. Due to this, evaluation of adjuvant properties is challenging, especially as it concerns elucidation of the mechanism of action or exclusion of potential interferences due to impurities [125].
Alginic Acids and Salts
Alginic acid is a linear copolymer containing (1-4)-linked β-D-mannuronate (M) and its C-5 epimer α-L-guluronate (G) residues, covalently linked together in different sequences or blocks (Fig. 5). Although alginates are widely used for the preparation of hydrogel microspheres and nanospheres for encapsulation of biological agents [126], there have been also reports on the adjuvant activity of soluble salts of alginic acid.
Fig. 5.
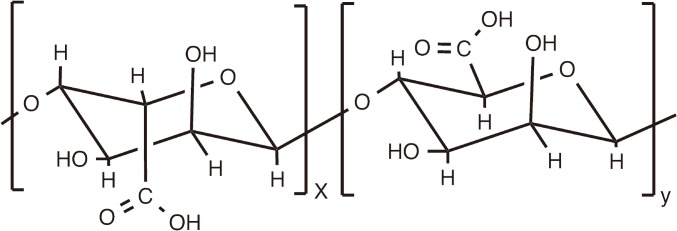
Schematic chemical structure of alginic acid.
Adjuvant effect of sterile sodium alginate was investigated in a subcutaneous immunization with BCG vaccine in BALB/c mice [127]. Proliferative and delayed-type hypersensitivity responses, interferon (IFN)-g, specific anti-mycobacterium total IgG, IgG1, and IgG2a production were found all significantly higher in mice immunized with adjuvanted formulation. In addition, following systemic infection with BCG, mice vaccinated with BCG plus alginate had lower mean bacterial count compared to those vaccinated with BCG alone [127].
The effect of sodium alginate was also evaluated in combination with microparticle carriers fabricated using biodegradable polymer-poly(lactic-co-glycolic acid) (PLGA) with two malaria synthetic peptides, SPf66 and S3 [128]. Despite the fact that the formulations were practically indistinguishable in terms of size and morphology, the addition of alginate improved both encapsulation efficiency and immune responses. Immunization studies in BALB/c mice by intradermal route demonstrated that incorporation of alginate elicited higher humoral and cellular immune responses leading to more balanced Th1/Th2 responses. Furthermore, administration of microparticles containing RGD-modified alginate showed evidence of cell targeting by enhancing immunogenicity of microparticles, in particular with regard to cellular responses such as IFN-c secretion and lymphoproliferation [128].
Polyoxidonium
Polyoxidonium-a water-soluble cationic polymer is a polyelectrolyte, which was specifically designed as an immunomodulator for use in vaccines [113,129]. It is a ternary copolymer of 1,4-ethylenepiperazine, 1,4-ethylenepiperazine-N-oxide, and (N-carboxymethylene)-1,4-ethylenepiperazinium bromide with a molecular weight of 60-100 kDa (Fig. 6). A derivative of poly(1,4-ethylenepiperazine), it is synthesized by a partial oxidation of the parent polymer with hydrogen peroxide to introduce N-oxide groups followed by the quaternization of some of the nonoxidized aminogroups with bromoacetic acid. The introduction of N-oxide groups is critically important as the optimal composition was selected in order to minimize toxicity, typically inherent to polyamines, and to maintain an appropriate level of immunostimulation [113]. In addition, the N-oxide units in the backbone are capable of degradation at elevated temperatures rearranging to oximes and then to amine and aldehyde groups. As a result, the copolymer chains are cleaved to shorter fragments, which then can be released from the body [113].
Fig. 6.
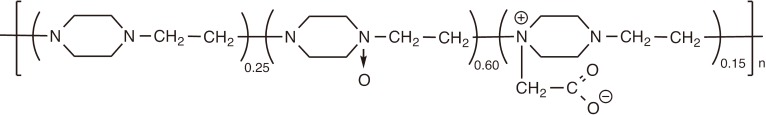
Schematic chemical structure of polyoxidonium.
The analysis of immunomodulating effect induced by polyoxidonium proved its stimulating activity on proinflammatory cytokines production in vitro, such as IL-1H, tumor necrosis factor (TNF)-α and IL-6 [129]. A dose-dependent increase in the intracellular killing by blood phagocytes was also observed for this polymer. In another study, it was established that polyoxidonium could affect the bactericidal activity of leukocytes [130]. Many chronic infectious inflammatory diseases are characterized by a sluggish, recurrent course, resistant to adequate therapy and requiring additional immunostimulation. It was established that a one-hour incubation of human peripheral blood leukocytes with polyoxidonium increased the ability of leukocytes to kill the ingested Staphylococcus aureus in a dose-dependent manner [130]. This increase was observed with leukocytes obtained both from healthy persons and from patients with chronic granulomatous disease. Polyoxidonium was also shown to have antioxidant activity at all dose range of 100 to 500 µg/mL [130]. Polyoxidonium displayed ability to enhance immune responses to the live brucellosis vaccine, Brucella abortus strain 82-PS (penicillin sensitive) in a guinea pig model [131].
One of the first clinical applications of polyoxidonium was with the commercial influenza vaccine, in which polyoxidonium was covalently conjugated to antigenic components of the vaccine-hemagglutinin and neuraminidase [113]. Reportedly, about 50 million recipients were vaccinated, and extensive data indicating the efficacy and safety character of the preparation was obtained [113]. In a clinical study polyoxidonium was also evaluated with trivalent live attenuated measles, mumps and rubella vaccine [132]. Although findings indicate that healthy children needed no fortification of their immune responses on the vaccination as they can produce a high level of specific antibodies, children with previous exposure to harmful factors affecting normal T cell content (viral and other diseases) may benefit from the use of polyoxidonium. However, authors also note that the detected increase in TNF-β level and skewing the dominant immune responses from Th1 to Th2 type could not be appreciated as positive effect of polyoxidonium in this environment [132].
Polyphosphazenes
Introduction to polyphosphazenes
Polyphosphazene immunoadjuvants are well-defined macromolecules, which are designed around the biodegradable phosphorus-nitrogen backbone and organic side groups containing anionic moieties. These macromolecules are uniquely positioned among other classes of synthetic and natural polyelectrolytes due to a number of structural and chemical features. Among them are hydrolytic degradation and benign degradation products, which can be controlled via selection of the appropriate side groups, flexibility of the backbone and high ionic density, which are critical for self-assembly with biological targets, and tremendous diversity and synthetic versatility of the class allowing the creation of unprecedented macromolecular structures. Well established synthetic route, reproducibility, high level of purity, ease of characterization and quality control, adequate stability, and "mix and fix" aqueous formulation approach, which does not involve covalent conjugation with antigen, all render these polymers attractive for commercial development [133,134,135,136].
Poly[di(carboxylatophenoxy)phosphazene], PCPP (Fig. 7) is by far the most investigated representative of this class, which has been successfully advanced to the development state. A potent immunoadjuvant effect of PCPP has been well-documented when administered with a variety of bacterial and viral antigens in a more than a dozen of animal models [137,138,139,140,141,142,143]. Its track record in human clinical trials includes significant improvement in seroconversion and seroprotection rates for a seasonal influenza vaccine [144] and four-fold rise in neutralizing antibodies against respiratory syncytial virus strains in greater than 75% of participants [145]. Vaccine formulations containing PCPP have been reported to be safe and immunogenic [146,147].
Fig. 7.

Schematic chemical structure of polyphosphazenes: PCPP (A), PCPP copolymer with oxyethylene groups (B), and PCEP (C). PCPP, poly[di(carboxylatophenoxy) phosphazene]; PCEP, poly[di(carboxylatoethylphenoxy)phosphazene].
Immunoadjuvant effect of polyphosphazenes can be characterized by modulations in the onset, magnitude, quality, and duration of immune responses. It has been reported that polyphosphazene adjuvants induced significant increases in antibody responses as early as 2 weeks after immunization for a number of antigen including influenza and hepatitis B [139,141,143,148]. Sustained levels of antibody titers for the length of the experiment (up to 41 weeks) were observed for X:31 influenza antigen and hepatitis B surface antigen (HBsAg) [139,140,141]. Another important feature of PCPP-antigen sparing-PCPP adjuvanted X:31 influenza formulations injected in mice were as potent as their non-adjuvanted counterparts containing 25 times higher dose of the antigen [141]. This important feature has been also demonstrated in lethal challenge studies with H5N1 influenza vaccine in ferrets, which is a highly relevant preclinical model. In these experiments PCPP formulated vaccine afforded 100% protection from mortality, whereas non-adjuvanted formulation was not protective at a dose of at least 10-fold higher [138]. Benchmarking of polyphosphazene adjuvanted vaccines against those containing "alum" (aluminum phosphate), suggest either greatly superior responses for polyphosphazenes in animal studies [140,141] or at least equal performance in humans [145]. PCPP was also proved to work well with gentle antigens, such as virus-like particles (VLPs). A single intramuscular dose of PCPP formulated rotavirus VLPs induced rotavirus-specific serum IgG and IgA, fecal IgG titers that were enhanced 5- to 90-fold by the adjuvant [149].
Polyphosphazenes Mechanism of Action
The mechanism of action of polyphosphazene immunoadjuvants is not yet fully understood. It has been demonstrated, that on the molecular level, the parameters of interactions between polyphosphazene and the antigen play an important role. Similarly to other polyelectrolyte adjuvants [113], biological activity of PCPP strongly depends on its association with the antigen, however, contrary to their conventional counterparts, PCPP can form stable water-soluble, non-covalent complexes with antigenic molecules spontaneously and thus do not require chemical conjugation [133,150]. The molecular size and antigen loading of the complex have been shown to correlate with its activity in vivo.
Recent study explored the effect of PCPP and its multimeric formulation with recombinant Gag-HIV antigen on the maturation, activation and antigen presentation by human adult and newborn DCs in vitro [150]. PCPP treatment induced DCs activation and maturation as assessed by upregulation of co-stimulatory molecules and cytokine production. The effect of PCPP was found to be more robust than that of "alum" (Adju-Phos) and resulted in the release of mixed Th1/Th2 cytokine responses, promoting both cellular and potentially humoral responses to the formulated antigen. PCPP also induced an innate immune transcriptome in human adult and newborn DCs [150]. Authors note that the ability of PCPP to activate neonatal monocyte derived DCs to levels similar to their adult counterparts is particularly noteworthy and suggests that this adjuvantation platform may have additional applicability for early life immunization, a key point of healthcare contact at which impaired immune responses are noted to many current vaccine formulations.
"Next Generation" Polyphosphazene Adjuvants
Much attention has been also focused on the 'next generation' polyphosphazene adjuvants. It has been clearly elucidated that structure and composition of polyphosphazenes play an important role in their immunopotentiation effect and novel molecules can differentiate significantly from PCPP in their activity and mechanism of action. Although about two dozen "new generation" polyphosphazenes, have been synthesized, only few have been explored for their immunoadjuvants properties, namely or PCPP copolymers containing oxyethelyne side groups (Fig. 7B) [151] poly[di(carboxylatoethylphenoxy)phosphazene], and PCEP (Fig. 7C) [152]. It was found that depending on the type of polyphosphazene the IgG isotype profiles of the immune response can vary indicating potential differences in the mechanism of action. PCPP primarily enhances IgG1 antibody responses, which are typically associated with Th2-type response, whereas its sister polymers have been shown to also enhance IgG2a [141,151], which can be associated with Th1-type immune responses providing protection against intracellular pathogens [153].
It was reported recently that PCEP can induce time-dependent changes in the gene expression of many "adjuvant core response genes" including cytokines, chemokines, innate immune receptors, IFN-induced genes, adhesion molecules and antigen-presentation genes, upregulate the gene expression of the inflammasome receptor, NLRP3, and induce the production of pro-inflammatory cytokines IL-1β, and IL-18 at the site of injection [154]. Since the secretion of these cytokines is predominantly a result of activation of the inflammasome, which leads to the processing and secretion of proinflammatory cytokines, authors suggest that PCEP may modulate antigen-specific immune responses by activating early innate immune responses and promoting a strong immuno-stimulatory environment at the site of injection [154]. Furthermore, intramuscular injection of mice with PCEP induced significant recruitment of neutrophils, macrophages, monocytes, DCs, and lymphocytes at the site of injection as well as in the draining lymph nodes [155]. The results of the confocal analysis revealed intracytoplasmic lysosomal localization of PCEP in recruited immune cells [155].
Although recent publications, especially on a newer generation polyphosphazenes, clearly shed light on their mechanism of action, it should be noted that very few of these papers detail synthesis, characterization and formulation studies. This certainly needs to be addressed to allow the exclusion of the impact of the secondary factors, such as molecular size of the polymer, potential self-assembly in solution, and even the effect of any structural irregularities in the polymer, on the immunological effects.
Polyphosphazene Nanoparticulates, Microneedles, and Synergy with Other Immunoadjuvants
Solubility of polyphosphazene immunoadjuvants in neutral and basic aqueous solutions provide for their straightforward formulation, however conformation-activity and molecular weight-activity relationships [133,139], as well as ionic sensitivity [136], may play an important roles in biological activity. Therefore, formulations must be characterized, tested for compatibility with a specific antigen, and optimized accordingly to achieve superior results. Importantly, due to their water-solubility and well-defined structures, polyphosphazenes provide an attractive basis for the development of combination adjuvants. In fact, a synergistic effect between PCPP, PCEP and some important adjuvants, such as CpG has been well established [148].
Although the majority of results on polyphosphazene adjuvants have been reported for water-soluble formulations, the versatility of these polymers allows for their use as nanoparticulate delivery systems or microneedles for intradermal immunization. In fact, PCPP and some of its copolymers can be easily conformed into hydrogel nano- or microparticles in aqueous solutions using ionic complexation processes with benign agents, such as calcium chloride or spermine. Contrary to nanoparticles comprised from hydrophobic biodegradable nanoparticles like PLGA, these aqueous technologies are highly protein compatible and don't require the use of organic solvents, elevated temperatures, or complex manufacturing equipment. Although the resulting systems are well characterized, allow control of size and surface characteristics, and provide for a slow release of both antigen and immunoadjuvants, their immunological behavior and antigen delivery capabilities are yet to be studied systematically.
Another attractive feature of polyphosphazene adjuvants is in its compatibility and with various intradermal immunization techniques [156,157]. The concept of fabricating microneedles using PCPP as a construction material stems out of the macromolecular nature of this adjuvant, which determines the required set of mechanical properties, such as strength, film forming characteristics, and adhesion. These features, combined with excellent water-solubility, make polyphosphazene adjuvant an attractive platform for vaccine transdermal patch technology. Polyphosphazenes enable straightforward production of coated or dissolvable microneedles with excellent skin penetration capabilities and fast formulation dissolution profiles once microneedles are applied to the skin. Immunization with microneedles containing recombinant HBsAg and PCPP in pigs resulted in an increase in antibody titers of at least 10-fold higher than intramuscular group with similar formulation, but injected intramuscularly as a solution, and three orders of magnitude higher than non-adjuvanted group. In these studies PCPP microneedles also showed a substantial potential for antigen dose sparing.
Overall, well-defined nature of this synthetic class of adjuvants, its biodegradable nature, ability to fine-tune the structure to achieve optimal immune response and modulate its quality, compatibility with other adjuvants, and versatility of formulations, makes this class of adjuvants especially attractive for challenging applications.
Stakeholders, standards, and considerations
Continued advancement on harmonization of international regulations will aid the development of adjuvanted vaccines. Adjuvants are licensed with the whole vaccine product, strictly as a specific adjuvant-antigen formulation for each vaccine and not as a separate adjuvant entity. The testing and documentation of an adjuvant is considered only in relation to the whole vaccine formulation as reviewed for safety, tolerability, toxicity, potency and efficacy. While efforts toward harmonization in biological drug product regulation between the United States, Europe and Japan have been in progress since 1990, there is presently no mutual recognition of licensing or lot release for vaccines. A recent biennial survey reported that Europe manufactured more than 45,000 doses, which made 79% of the world market in vaccine production. North America manufactured 13% of world supply, with China and India at 8% [158]. Therefore, full alignment of regulatory rules and guidelines will aid the development, review and licensing of adjuvants as components of vaccines entering global commerce.
Standardization of scientific nomenclature and laboratory practices will aid the ongoing development and application of vaccine adjuvants, particularly as new combination adjuvants are devised and compared [96]. This is important for research and development efforts as well as in manufacture and preclinical testing. As noted above, wider use of standardizations will help animal and in vitro models are of special concern due to variations between cell lines and laboratory strains. Efforts to completely as possible describe and communicate the with regard germ line immune system receptor alleles (TLR, CD, and lectin-binding, etc.) and other alleles impact their use as surrogates for disease indications that cannot be directly tested in people, such as the anthrax vaccine for biodefense.
As mentioned above, characterization of compound structure and use of relevant naming is of critical importance to comparing the effectiveness and safety of vaccine adjuvants. Generous use of terms such as alum and Al(PO4)3 can cause confusion and confound science-based communication of safety. Aluminum gels are heterogeneous in composition and higher order structure, and polymers may be copolymers and homopolymers with residual groups, so it no longer serves common interest in the science or technology of adjuvants to provide only a common name: best efforts for accurate naming and experimental characterization of composition and structure are needed.
The advancement of polymers in the field of vaccine delivery and immunopotentiation is largely explained by their physicochemical properties allowing their use as materials for the preparation of nanoparticles and encapsulation of antigens. Polymers and copolymers of lactic and glycolic acid are one of the most common choices for such applications. Ionic macromolecules, both synthetic and natural, provide an important alternative to this approach. They afford similar physical properties, which stem out of their macromolecular nature, but offer protein friendly aqueous environment for antigen encapsulation. In addition, they can integrate physical characteristics of vaccine carriers with powerful immunopotentiating properties. It is clear that such systems begin attracting increasing attention from both academic and industrial researchers. Much progress has been made in terms of studying their biologically relevant properties and mechanism of action. However, future development can be impeded by the lack of standard approaches to basic characterization of such system. This is especially relevant to macromolecules from natural sources. Although excellent immunological research has been conducted on chitin and chitosan, the absolute majority of papers lack fundamental description of materials used in these studies. This is especially concerning as methods for isolation of chitin as well as its purity level vary dramatically and its insolubility under the conditions of use poses severe restrictions on the analytical approaches. Chitosan, although providing some water-solubility, is a mixture of copolymers with varying compositions, random distribution of repeating units, and molecular sizes. Proper analysis of polymer compositions and molecular characterization using common analytical techniques is clearly an absolute necessity for establishing proper structure activity relationship and future development of the field. The situation becomes even more challenging when the lack of the appropriate analysis itself is further complicated by insufficient characterization of nanoparticles.
In this regard, synthetic polyelectrolytes, such as polyphosphazenes and polyoxidonium, are better positioned due to their well-defined production processes and simplicity of molecular structures. Nevertheless, it has to be emphasized that even these molecules can contain structural irregularities and contaminants, which has been shown to affect their performance in vivo. There a clear need for a thorough characterization of such systems in order to avoid any potential misinterpretation of the results. Fortunately, the well-defined structures and water-solubility of such macromolecules allow the application of a whole array of powerful analytical methods currently employed in polymer science. Nuclear magnetic resonance, size exclusion chromatography, field flow fractionation, capillary electrophoresis, dynamic and static light scattering are among the techniques that can be and should be successfully applied to avoid undesired ambiguity in the interpretation of the results. Further development of this important class of immunopotentiators and vaccine delivery vehicles builds on the reliability of their production and characterization methods.
Conclusions
Adjuvants have established value for vaccinology and the vaccines industry and emerged as a sub-discipline of knowledge and technology, at the crossroads of immunology, cell biology, bioinformatics, nanotechnology and materials sciences, public health, and several other areas of scientific knowledge and practice. This review has updated just a few aspects of adjuvants and adjuvantation and traced the current state of knowledge back to the origins of adjuvant use and science. We focused on particles and colloids of polyionic composition-aluminum salt gels and polyelectrolyte polymers-which are historical and remain important to vaccine product development. Evidence is now showing that nanoparticulate structure and surface chemistry of aluminum salt compounds are critical determinants of adjuvant activity. Therefore, measurement and control of such attributes can improve the comparability, capability, quality and further product development for this oldest of vaccine adjuvants. Definition and control of surface chemistry and molecular structure are also important to polyelectrolyte polymer adjuvants. Recent information has improved the understanding of their molecular structure and interactions with immune cells, and their mechanism of action continues to be the subject of intensive study in the quest to expand applicability and utility for vaccine technology. Adjuvants including polyionic materials will likely gain greater importance in the development of newer vaccines of greater capability and safety.
Footnotes
We thank Wayne Morges for critical reading of the manuscript and Jill Sharp for help in background research. Bradford S. Powell and Peter C. Fusco received compensation from PharmAthene as employees during manuscript preparation. Alexander Andrianov received compensation from the University of Maryland IBBR as employee during manuscript preparation. Alexander K. Andrianov is a co-inventor of patents on polyphosphazene technology. PharmAthene is developing a next generation anthrax vaccine, SparVax, described by published work cited in this review.
References
- 1.van Panhuis WG, Grefenstette J, Jung SY, et al. Contagious diseases in the United States from 1888 to the present. N Engl J Med. 2013;369:2152–2158. doi: 10.1056/NEJMms1215400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rappuoli R, Pizza M, Del Giudice G, De Gregorio E. Vaccines, new opportunities for a new society. Proc Natl Acad Sci U S A. 2014;111:12288–12293. doi: 10.1073/pnas.1402981111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.World Health Organization. Global Vaccine Action Plan 2011-2020. Geneva: WHO; 2013. [Google Scholar]
- 4.Black RE, Cousens S, Johnson HL, et al. Global, regional, and national causes of child mortality in 2008: a systematic analysis. Lancet. 2010;375:1969–1987. doi: 10.1016/S0140-6736(10)60549-1. [DOI] [PubMed] [Google Scholar]
- 5.Rusnak JM, Kortepeter MG, Hawley RJ, Anderson AO, Boudreau E, Eitzen E. Risk of occupationally acquired illnesses from biological threat agents in unvaccinated laboratory workers. Biosecur Bioterror. 2004;2:281–293. doi: 10.1089/bsp.2004.2.281. [DOI] [PubMed] [Google Scholar]
- 6.World Health Organization. Global vaccine action plan: monitoring, evaluation and accountability annual report. Geneva: WHO; 2014. [Google Scholar]
- 7.Mahadevan M, Navarro-Locsin G, Tan HK, et al. A review of the burden of disease due to otitis media in the Asia-Pacific. Int J Pediatr Otorhinolaryngol. 2012;76:623–635. doi: 10.1016/j.ijporl.2012.02.031. [DOI] [PubMed] [Google Scholar]
- 8.Memish ZA. Meningococcal disease and travel. Clin Infect Dis. 2002;34:84–90. doi: 10.1086/323403. [DOI] [PubMed] [Google Scholar]
- 9.Hinman A. Eradication of vaccine-preventable diseases. Annu Rev Public Health. 1999;20:211–229. doi: 10.1146/annurev.publhealth.20.1.211. [DOI] [PubMed] [Google Scholar]
- 10.Hennessey K, Schluter WW, Wang X, et al. Are we there yet? Assessing achievement of vaccine-preventable disease goals in WHO's Western Pacific Region. Vaccine. 2014;32:4259–4266. doi: 10.1016/j.vaccine.2014.02.093. [DOI] [PubMed] [Google Scholar]
- 11.Riedel S. Edward Jenner and the history of smallpox and vaccination. Proc (Bayl Univ Med Cent) 2005;18:21–25. doi: 10.1080/08998280.2005.11928028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Henderson DA. The eradication of smallpox: an overview of the past, present, and future. Vaccine. 2011;29(Suppl 4):D7–D9. doi: 10.1016/j.vaccine.2011.06.080. [DOI] [PubMed] [Google Scholar]
- 13.Anand A, Pallansch MA, Estivariz CF, Gary H, Wassilak SG. Estimating the likely coverage of inactivated poliovirus vaccine in routine immunization: evidence from demographic and health surveys. J Infect Dis. 2014;210(Suppl 1):S465–S474. doi: 10.1093/infdis/jiu343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Klepac P, Metcalf CJ, McLean AR, Hampson K. Towards the endgame and beyond: complexities and challenges for the elimination of infectious diseases. Philos Trans R Soc Lond B Biol Sci. 2013;368:20120137. doi: 10.1098/rstb.2012.0137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lievano F, Galea SA, Thornton M, et al. Measles, mumps, and rubella virus vaccine (M-M-RII): a review of 32 years of clinical and postmarketing experience. Vaccine. 2012;30:6918–6926. doi: 10.1016/j.vaccine.2012.08.057. [DOI] [PubMed] [Google Scholar]
- 16.Alicino C, Merlano C, Zappettini S, et al. Routine surveillance of adverse events following immunization as an important tool to monitor vaccine safety: the two-years' experience of the Liguria Region, Italy. Hum Vaccin Immunother. 2015;11:91–94. doi: 10.4161/hv.34360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Arguedas A, Soley C, Abdelnour A, et al. Assessment of the safety, tolerability and kinetics of the immune response to A/H1N1v vaccine formulations with and without adjuvant in healthy pediatric subjects from 3 through 17 years of age. Hum Vaccin. 2011;7:58–66. doi: 10.4161/hv.7.1.13411. [DOI] [PubMed] [Google Scholar]
- 18.Chang S, O'Connor PM, Slade BA, Woo EJ. U.S. Postlicensure safety surveillance for adolescent and adult tetanus, diphtheria and acellular pertussis vaccines: 2005-2007. Vaccine. 2013;31:1447–1452. doi: 10.1016/j.vaccine.2012.10.097. [DOI] [PubMed] [Google Scholar]
- 19.Cottin P, Niedrig M, Domingo C. Safety profile of the yellow fever vaccine Stamaril(R): a 17-year review. Expert Rev Vaccines. 2013;12:1351–1368. doi: 10.1586/14760584.2013.836320. [DOI] [PubMed] [Google Scholar]
- 20.DeStefano F Vaccine Safety Datalink Research Group. The Vaccine Safety Datalink project. Pharmacoepidemiol Drug Saf. 2001;10:403–406. doi: 10.1002/pds.613. [DOI] [PubMed] [Google Scholar]
- 21.Lee CJ, Lee LH, Lu CH, Huang YJ, Chu ML. Safety monitoring in vaccine development and immunization. Acta Paediatr Taiwan. 2006;47:7–13. [PubMed] [Google Scholar]
- 22.Macartney KK, Chiu C, Georgousakis M, Brotherton JM. Safety of human papillomavirus vaccines: a review. Drug Saf. 2013;36:393–412. doi: 10.1007/s40264-013-0039-5. [DOI] [PubMed] [Google Scholar]
- 23.Tseng HF, Liu A, Sy L, et al. Safety of zoster vaccine in adults from a large managed-care cohort: a Vaccine Safety Datalink study. J Intern Med. 2012;271:510–520. doi: 10.1111/j.1365-2796.2011.02474.x. [DOI] [PubMed] [Google Scholar]
- 24.Nguyen TH, Vu MH, Nguyen VC, et al. A reduction in chronic hepatitis B virus infection prevalence among children in Vietnam demonstrates the importance of vaccination. Vaccine. 2014;32:217–222. doi: 10.1016/j.vaccine.2013.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Teleb N, Lebo E, Ahmed H, et al. Progress toward measles elimination: Eastern Mediterranean Region, 2008-2012. MMWR Morb Mortal Wkly Rep. 2014;63:511–515. [PMC free article] [PubMed] [Google Scholar]
- 26.Kapoor R, Kottilil S. Strategies to eliminate HBV infection. Future Virol. 2014;9:565–585. doi: 10.2217/fvl.14.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Andre FE, Booy R, Bock HL, et al. Vaccination greatly reduces disease, disability, death and inequity worldwide. Bull World Health Organ. 2008;86:140–146. doi: 10.2471/BLT.07.040089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Garcon N, Hem S, Friede M. Evolution of adjuvants across the centuries. In: Plotkin S, Ornstein W, Offit P, editors. Vaccines. 6th ed. Philadelphia: Saunders Elsevier; 2013. pp. 58–70. [Google Scholar]
- 29.Glenny AT, Sudmersen HJ. Notes on the production of immunity to diphtheria toxin. J Hyg (Lond) 1921;20:176–220. doi: 10.1017/s0022172400033945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Emil von Behring: the founder of serum therapy [Internet] Nobel Foundation; 2001. [cited 2014 Nov 18]. Available from: http://www.nobelprize.org/nobel_prizes/medicine/laureates/1901/behring-article.html. [Google Scholar]
- 31.Kantha SS. The legacy of von Behring and Kitasato. Immunol Today. 1992;13:374. doi: 10.1016/0167-5699(92)90177-9. [DOI] [PubMed] [Google Scholar]
- 32.Plotkin SL, Plotkin SA. A short history of vaccination. In: Plotkin S, Ornstein W, Offit P, editors. Vaccines. 6th ed. Philadelphia: Saunders Elsevier; 2013. pp. 1–13. [Google Scholar]
- 33.The tetanus cases in Camden, N.J. JAMA. 1901;37:1539–1540. [Google Scholar]
- 34.U.S. Food and Drug Administration. Biologics centennial: 100 years of biologics regulation [Internet] Silver Spring: FDA; 2002. [cited 2014 Nov 18]. Available from: http://www.fda.gov/AboutFDA/WhatWeDo/History/ProductRegulation/SelectionsFromFDLIUpdateSeriesonFDAHistory/ucm091754.htm. [Google Scholar]
- 35.Dudzinski DM. Reflections on historical, scientific, and legal issues relevant to designing approval pathways for generic versions of recombinant protein-based therapeutics and monoclonal antibodies. Food Drug Law J. 2005;60:143–260. [PubMed] [Google Scholar]
- 36.Ramon G. Regarding the flocculant ability and immunizing properties of a diphtheria toxin rendered atoxic. C R Hebd Seances Acad Sci. 1923;177:1338–1340. [Google Scholar]
- 37.Glenny AT, Pope CG, Waddington H, Wallace U. Immunological notes: XVII-XXIV. J Pathol Bacteriol. 1926;29:31–40. [Google Scholar]
- 38.Park WH, Schroder MC. Diphtheria toxin-antitoxin and toxoid: a comparison. Am J Public Health Nations Health. 1932;22:7–16. doi: 10.2105/ajph.22.1.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Baker JN, Gill DG. Precipitated toxoid as an immunizing agent against diphtheria. Am J Public Health Nations Health. 1934;24:22–24. doi: 10.2105/ajph.24.1.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Ericsson H. Purification and adsorption of diphtheria toxoid. Nature. 1946;158:350. doi: 10.1038/158350a0. [DOI] [PubMed] [Google Scholar]
- 41.Holt LB. Purified precipitated diphtheria toxoid of constant composition. Lancet. 1947;1:282–285. doi: 10.1016/s0140-6736(47)90094-9. [DOI] [PubMed] [Google Scholar]
- 42.Rappuoli R, Mandl CW, Black S, De Gregorio E. Vaccines for the twenty-first century society. Nat Rev Immunol. 2011;11:865–872. doi: 10.1038/nri3085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Freund J. The effect of paraffin oil and mycobacteria on antibody formation and sensitization: a review. Am J Clin Pathol. 1951;21:645–656. doi: 10.1093/ajcp/21.7.645. [DOI] [PubMed] [Google Scholar]
- 44.Petrovsky N, Aguilar JC. Vaccine adjuvants: current state and future trends. Immunol Cell Biol. 2004;82:488–496. doi: 10.1111/j.0818-9641.2004.01272.x. [DOI] [PubMed] [Google Scholar]
- 45.Stuart-Harris CH. Adjuvant influenza vaccines. Bull World Health Organ. 1969;41:617–621. [PMC free article] [PubMed] [Google Scholar]
- 46.O'Hagan DT, Ott GS, Nest GV, Rappuoli R, Giudice GD. The history of MF59((R)) adjuvant: a phoenix that arose from the ashes. Expert Rev Vaccines. 2013;12:13–30. doi: 10.1586/erv.12.140. [DOI] [PubMed] [Google Scholar]
- 47.Morel S, Didierlaurent A, Bourguignon P, et al. Adjuvant System AS03 containing alpha-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine. 2011;29:2461–2473. doi: 10.1016/j.vaccine.2011.01.011. [DOI] [PubMed] [Google Scholar]
- 48.Caillet C, Piras F, Bernard MC, et al. AF03-adjuvanted and non-adjuvanted pandemic influenza A (H1N1) 2009 vaccines induce strong antibody responses in seasonal influenza vaccine-primed and unprimed mice. Vaccine. 2010;28:3076–3079. doi: 10.1016/j.vaccine.2010.02.050. [DOI] [PubMed] [Google Scholar]
- 49.Fox CB, Baldwin SL, Duthie MS, Reed SG, Vedvick TS. Immunomodulatory and physical effects of oil composition in vaccine adjuvant emulsions. Vaccine. 2011;29:9563–9572. doi: 10.1016/j.vaccine.2011.08.089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Rietschel ET, Cavaillon JM. Richard Pfeiffer and Alexandre Besredka: creators of the concept of endotoxin and anti-endotoxin. Microbes Infect. 2003;5:1407–1414. doi: 10.1016/j.micinf.2003.10.003. [DOI] [PubMed] [Google Scholar]
- 51.Nauts HC, Swift WE, Coley BL. The treatment of malignant tumors by bacterial toxins as developed by the late William B. Coley, M.D., reviewed in the light of modern research. Cancer Res. 1946;6:205–216. [PubMed] [Google Scholar]
- 52.Medzhitov R. Toll-like receptors and innate immunity. Nat Rev Immunol. 2001;1:135–145. doi: 10.1038/35100529. [DOI] [PubMed] [Google Scholar]
- 53.Medzhitov R, Janeway CA., Jr Innate immunity: impact on the adaptive immune response. Curr Opin Immunol. 1997;9:4–9. doi: 10.1016/s0952-7915(97)80152-5. [DOI] [PubMed] [Google Scholar]
- 54.Reed SG, Orr MT, Fox CB. Key roles of adjuvants in modern vaccines. Nat Med. 2013;19:1597–1608. doi: 10.1038/nm.3409. [DOI] [PubMed] [Google Scholar]
- 55.De Gregorio E, Rappuoli R. From empiricism to rational design: a personal perspective of the evolution of vaccine development. Nat Rev Immunol. 2014;14:505–514. doi: 10.1038/nri3694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Petrovsky N. Vaccine adjuvants: in search of new paradigms. Interview by Jenaid Rees. Expert Rev Vaccines. 2013;12:723–726. doi: 10.1586/14760584.2013.811197. [DOI] [PubMed] [Google Scholar]
- 57.Koff WC, Burton DR, Johnson PR, et al. Accelerating next-generation vaccine development for global disease prevention. Science. 2013;340:1232910. doi: 10.1126/science.1232910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Brito LA, Malyala P, O'Hagan DT. Vaccine adjuvant formulations: a pharmaceutical perspective. Semin Immunol. 2013;25:130–145. doi: 10.1016/j.smim.2013.05.007. [DOI] [PubMed] [Google Scholar]
- 59.Li L, Saade F, Petrovsky N. The future of human DNA vaccines. J Biotechnol. 2012;162:171–182. doi: 10.1016/j.jbiotec.2012.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Williams A, Flavell RA, Eisenbarth SC. The role of NOD-like receptors in shaping adaptive immunity. Curr Opin Immunol. 2010;22:34–40. doi: 10.1016/j.coi.2010.01.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Coffman RL, Sher A, Seder RA. Vaccine adjuvants: putting innate immunity to work. Immunity. 2010;33:492–503. doi: 10.1016/j.immuni.2010.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Carter D, Reed SG. Role of adjuvants in modeling the immune response. Curr Opin HIV AIDS. 2010;5:409–413. doi: 10.1097/COH.0b013e32833d2cdb. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Rappuoli R. Reverse vaccinology. Curr Opin Microbiol. 2000;3:445–450. doi: 10.1016/s1369-5274(00)00119-3. [DOI] [PubMed] [Google Scholar]
- 64.Vos Q, Lees A, Wu ZQ, Snapper CM, Mond JJ. B-cell activation by T-cell-independent type 2 antigens as an integral part of the humoral immune response to pathogenic microorganisms. Immunol Rev. 2000;176:154–170. doi: 10.1034/j.1600-065x.2000.00607.x. [DOI] [PubMed] [Google Scholar]
- 65.Wetzler LM. Innate immune function of the neisserial porins and the relationship to vaccine adjuvant activity. Future Microbiol. 2010;5:749–758. doi: 10.2217/fmb.10.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Massari P, Gunawardana J, Liu X, Wetzler LM. Meningococcal porin PorB prevents cellular apoptosis in a toll-like receptor 2- and NF-kappaB-independent manner. Infect Immun. 2010;78:994–1003. doi: 10.1128/IAI.00156-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Taylor DN, Treanor JJ, Sheldon EA, et al. Development of VAX128, a recombinant hemagglutinin (HA) influenza-flagellin fusion vaccine with improved safety and immune response. Vaccine. 2012;30:5761–5769. doi: 10.1016/j.vaccine.2012.06.086. [DOI] [PubMed] [Google Scholar]
- 68.De Gregorio E, D'Oro U, Wack A. Immunology of TLR-independent vaccine adjuvants. Curr Opin Immunol. 2009;21:339–345. doi: 10.1016/j.coi.2009.05.003. [DOI] [PubMed] [Google Scholar]
- 69.Kay E, Scotland RS, Whiteford JR. Toll-like receptors: Role in inflammation and therapeutic potential. Biofactors. 2014;40:284–294. doi: 10.1002/biof.1156. [DOI] [PubMed] [Google Scholar]
- 70.Cervantes JL, Weinerman B, Basole C, Salazar JC. TLR8: the forgotten relative revindicated. Cell Mol Immunol. 2012;9:434–438. doi: 10.1038/cmi.2012.38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Carta S, Tassi S, Pettinati I, Delfino L, Dinarello CA, Rubartelli A. The rate of interleukin-1beta secretion in different myeloid cells varies with the extent of redox response to Toll-like receptor triggering. J Biol Chem. 2011;286:27069–27080. doi: 10.1074/jbc.M110.203398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Isakov E, Weisman-Shomer P, Benhar M. Suppression of the pro-inflammatory NLRP3/interleukin-1beta pathway in macrophages by the thioredoxin reductase inhibitor auranofin. Biochim Biophys Acta. 2014;1840:3153–3161. doi: 10.1016/j.bbagen.2014.07.012. [DOI] [PubMed] [Google Scholar]
- 73.Dasu MR, Isseroff RR. Toll-like receptors in wound healing: location, accessibility, and timing. J Invest Dermatol. 2012;132:1955–1958. doi: 10.1038/jid.2012.208. [DOI] [PubMed] [Google Scholar]
- 74.Chen GY, Nunez G. Sterile inflammation: sensing and reacting to damage. Nat Rev Immunol. 2010;10:826–837. doi: 10.1038/nri2873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Muralidharan S, Mandrekar P. Cellular stress response and innate immune signaling: integrating pathways in host defense and inflammation. J Leukoc Biol. 2013;94:1167–1184. doi: 10.1189/jlb.0313153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Mount A, Koernig S, Silva A, Drane D, Maraskovsky E, Morelli AB. Combination of adjuvants: the future of vaccine design. Expert Rev Vaccines. 2013;12:733–746. doi: 10.1586/14760584.2013.811185. [DOI] [PubMed] [Google Scholar]
- 77.Kuroda E, Coban C, Ishii KJ. Particulate adjuvant and innate immunity: past achievements, present findings, and future prospects. Int Rev Immunol. 2013;32:209–220. doi: 10.3109/08830185.2013.773326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Zhou R, Yazdi AS, Menu P, Tschopp J. A role for mitochondria in NLRP3 inflammasome activation. Nature. 2011;469:221–225. doi: 10.1038/nature09663. [DOI] [PubMed] [Google Scholar]
- 79.Flach TL, Ng G, Hari A, et al. Alum interaction with dendritic cell membrane lipids is essential for its adjuvanticity. Nat Med. 2011;17:479–487. doi: 10.1038/nm.2306. [DOI] [PubMed] [Google Scholar]
- 80.Seubert A, Calabro S, Santini L, et al. Adjuvanticity of the oil-in-water emulsion MF59 is independent of Nlrp3 inflammasome but requires the adaptor protein MyD88. Proc Natl Acad Sci U S A. 2011;108:11169–11174. doi: 10.1073/pnas.1107941108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Wilson NS, Duewell P, Yang B, et al. Inflammasome-dependent and -independent IL-18 production mediates immunity to the ISCOMATRIX adjuvant. J Immunol. 2014;192:3259–3268. doi: 10.4049/jimmunol.1302011. [DOI] [PubMed] [Google Scholar]
- 82.Baroja-Mazo A, Martin-Sanchez F, Gomez AI, et al. The NLRP3 inflammasome is released as a particulate danger signal that amplifies the inflammatory response. Nat Immunol. 2014;15:738–748. doi: 10.1038/ni.2919. [DOI] [PubMed] [Google Scholar]
- 83.McKee AS, Munks MW, MacLeod MK, et al. Alum induces innate immune responses through macrophage and mast cell sensors, but these sensors are not required for alum to act as an adjuvant for specific immunity. J Immunol. 2009;183:4403–4414. doi: 10.4049/jimmunol.0900164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Hutchison S, Benson RA, Gibson VB, Pollock AH, Garside P, Brewer JM. Antigen depot is not required for alum adjuvanticity. FASEB J. 2012;26:1272–1279. doi: 10.1096/fj.11-184556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Watkinson A, Soliakov A, Ganesan A, et al. Increasing the potency of an alhydrogel-formulated anthrax vaccine by minimizing antigen-adjuvant interactions. Clin Vaccine Immunol. 2013;20:1659–1668. doi: 10.1128/CVI.00320-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Iyer V, Hu L, Liyanage MR, et al. Preformulation characterization of an aluminum salt-adjuvanted trivalent recombinant protein-based vaccine candidate against Streptococcus pneumoniae. J Pharm Sci. 2012;101:3078–3090. doi: 10.1002/jps.23175. [DOI] [PubMed] [Google Scholar]
- 87.Soliakov A, Kelly IF, Lakey JH, Watkinson A. Anthrax sub-unit vaccine: the structural consequences of binding rPA83 to Alhydrogel(R) Eur J Pharm Biopharm. 2012;80:25–32. doi: 10.1016/j.ejpb.2011.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Exley C, Siesjo P, Eriksson H. The immunobiology of aluminium adjuvants: how do they really work. Trends Immunol. 2010;31:103–109. doi: 10.1016/j.it.2009.12.009. [DOI] [PubMed] [Google Scholar]
- 89.Lu F, Hogenesch H. Kinetics of the inflammatory response following intramuscular injection of aluminum adjuvant. Vaccine. 2013;31:3979–3986. doi: 10.1016/j.vaccine.2013.05.107. [DOI] [PubMed] [Google Scholar]
- 90.Rimaniol AC, Gras G, Verdier F, et al. Aluminum hydroxide adjuvant induces macrophage differentiation towards a specialized antigen-presenting cell type. Vaccine. 2004;22:3127–3135. doi: 10.1016/j.vaccine.2004.01.061. [DOI] [PubMed] [Google Scholar]
- 91.Mold M, Eriksson H, Siesjo P, Darabi A, Shardlow E, Exley C. Unequivocal identification of intracellular aluminium adjuvant in a monocytic THP-1 cell line. Sci Rep. 2014;4:6287. doi: 10.1038/srep06287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.He Y. Vaccine adjuvant informatics: from data integration and analysis to rational vaccine adjuvant design. Front Immunol. 2014;5:32. doi: 10.3389/fimmu.2014.00032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Sayers S, Ulysse G, Xiang Z, He Y. Vaxjo: a web-based vaccine adjuvant database and its application for analysis of vaccine adjuvants and their uses in vaccine development. J Biomed Biotechnol. 2012;2012:831486. doi: 10.1155/2012/831486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Hartung DM, Zarin DA, Guise JM, McDonagh M, Paynter R, Helfand M. Reporting discrepancies between the ClinicalTrials.gov results database and peer-reviewed publications. Ann Intern Med. 2014;160:477–483. doi: 10.7326/M13-0480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Zarin DA, Tse T, Williams RJ, Califf RM, Ide NC. The ClinicalTrials.gov results database--update and key issues. N Engl J Med. 2011;364:852–860. doi: 10.1056/NEJMsa1012065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Levast B, Berri M, Wilson HL, Meurens F, Salmon H. Development of gut immunoglobulin A production in piglet in response to innate and environmental factors. Dev Comp Immunol. 2014;44:235–244. doi: 10.1016/j.dci.2013.12.012. [DOI] [PubMed] [Google Scholar]
- 97.Gupta RK, Rost BE, Relyveld E, Siber GR. Adjuvant properties of aluminum and calcium compounds. Pharm Biotechnol. 1995;6:229–248. doi: 10.1007/978-1-4615-1823-5_8. [DOI] [PubMed] [Google Scholar]
- 98.Hem SL, White JL. Structure and properties of aluminum-containing adjuvants. Pharm Biotechnol. 1995;6:249–276. doi: 10.1007/978-1-4615-1823-5_9. [DOI] [PubMed] [Google Scholar]
- 99.Li X, Aldayel AM, Cui Z. Aluminum hydroxide nanoparticles show a stronger vaccine adjuvant activity than traditional aluminum hydroxide microparticles. J Control Release. 2014;173:148–157. doi: 10.1016/j.jconrel.2013.10.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Barbaud A, Deschildre A, Waton J, Raison-Peyron N, Trechot P. Hypersensitivity and vaccines: an update. Eur J Dermatol. 2013;23:135–141. doi: 10.1684/ejd.2012.1842. [DOI] [PubMed] [Google Scholar]
- 101.Pittman PR, Kim-Ahn G, Pifat DY, et al. Anthrax vaccine: immunogenicity and safety of a dose-reduction, route-change comparison study in humans. Vaccine. 2002;20:1412–1420. doi: 10.1016/s0264-410x(01)00462-5. [DOI] [PubMed] [Google Scholar]
- 102.Hogenesch H. Mechanism of immunopotentiation and safety of aluminum adjuvants. Front Immunol. 2012;3:406. doi: 10.3389/fimmu.2012.00406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Jiang D, Premachandra GS, Johnston C, Hem SL. Structure and adsorption properties of commercial calcium phosphate adjuvant. Vaccine. 2004;23:693–698. doi: 10.1016/j.vaccine.2004.06.029. [DOI] [PubMed] [Google Scholar]
- 104.Eisenbarth SC, Colegio OR, O'Connor W, Sutterwala FS, Flavell RA. Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature. 2008;453:1122–1126. doi: 10.1038/nature06939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Sharp FA, Ruane D, Claass B, et al. Uptake of particulate vaccine adjuvants by dendritic cells activates the NALP3 inflammasome. Proc Natl Acad Sci U S A. 2009;106:870–875. doi: 10.1073/pnas.0804897106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Schinwald A, Donaldson K. Use of back-scatter electron signals to visualise cell/nanowires interactions in vitro and in vivo: frustrated phagocytosis of long fibres in macrophages and compartmentalisation in mesothelial cells in vivo. Part Fibre Toxicol. 2012;9:34. doi: 10.1186/1743-8977-9-34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Williams GR, Fierens K, Preston SG, et al. Immunity induced by a broad class of inorganic crystalline materials is directly controlled by their chemistry. J Exp Med. 2014;211:1019–1025. doi: 10.1084/jem.20131768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Sun B, Ji Z, Liao YP, et al. Engineering an effective immune adjuvant by designed control of shape and crystallinity of aluminum oxyhydroxide nanoparticles. ACS Nano. 2013;7:10834–10849. doi: 10.1021/nn404211j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Mori A, Oleszycka E, Sharp FA, et al. The vaccine adjuvant alum inhibits IL-12 by promoting PI3 kinase signaling while chitosan does not inhibit IL-12 and enhances Th1 and Th17 responses. Eur J Immunol. 2012;42:2709–2719. doi: 10.1002/eji.201242372. [DOI] [PubMed] [Google Scholar]
- 110.Neumann S, Burkert K, Kemp R, Rades T, Rod Dunbar P, Hook S. Activation of the NLRP3 inflammasome is not a feature of all particulate vaccine adjuvants. Immunol Cell Biol. 2014;92:535–542. doi: 10.1038/icb.2014.21. [DOI] [PubMed] [Google Scholar]
- 111.Lu F, Boutselis I, Borch RF, Hogenesch H. Control of antigen-binding to aluminum adjuvants and the immune response with a novel phosphonate linker. Vaccine. 2013;31:4362–4367. doi: 10.1016/j.vaccine.2013.07.019. [DOI] [PubMed] [Google Scholar]
- 112.Lindblad EB. Aluminium compounds for use in vaccines. Immunol Cell Biol. 2004;82:497–505. doi: 10.1111/j.0818-9641.2004.01286.x. [DOI] [PubMed] [Google Scholar]
- 113.Kabanov VA. From synthetic polyelectrolytes to polymer-subunit vaccines. Pure Appl Chem. 2004;76:1659–1677. [Google Scholar]
- 114.Andrianov AK. Polyphosphazene vaccine delivery vehicles: state of development and perspectives. In: Andrianov AK, editor. Polyphosphazenes for biomedical applications. Hoboken: John Wiley & Sons Inc.; 2009. pp. 47–63. [Google Scholar]
- 115.Muzzarelli RA. Chitins and chitosans as immunoadjuvants and non-allergenic drug carriers. Mar Drugs. 2010;8:292–312. doi: 10.3390/md8020292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Amidi M, Mastrobattista E, Jiskoot W, Hennink WE. Chitosan-based delivery systems for protein therapeutics and antigens. Adv Drug Deliv Rev. 2010;62:59–82. doi: 10.1016/j.addr.2009.11.009. [DOI] [PubMed] [Google Scholar]
- 117.Jayakumar R, Prabaharan M, Sudheesh Kumar PT, Nair SV, Tamura H. Biomaterials based on chitin and chitosan in wound dressing applications. Biotechnol Adv. 2011;29:322–337. doi: 10.1016/j.biotechadv.2011.01.005. [DOI] [PubMed] [Google Scholar]
- 118.You J, Li W, Yu C, et al. Amphiphilically modified chitosan cationic nanoparticles for drug delivery. J Nanopart Res. 2013;15:2123. [Google Scholar]
- 119.Nishimura K, Nishimura S, Nishi N, Saiki I, Tokura S, Azuma I. Immunological activity of chitin and its derivatives. Vaccine. 1984;2:93–99. doi: 10.1016/s0264-410x(98)90039-1. [DOI] [PubMed] [Google Scholar]
- 120.Lee CG. Chitin, chitinases and chitinase-like proteins in allergic inflammation and tissue remodeling. Yonsei Med J. 2009;50:22–30. doi: 10.3349/ymj.2009.50.1.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Lee CG, Da Silva CA, Dela Cruz CS, et al. Role of chitin and chitinase/chitinase-like proteins in inflammation, tissue remodeling, and injury. Annu Rev Physiol. 2011;73:479–501. doi: 10.1146/annurev-physiol-012110-142250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Zaharoff DA, Rogers CJ, Hance KW, Schlom J, Greiner JW. Chitosan solution enhances both humoral and cell-mediated immune responses to subcutaneous vaccination. Vaccine. 2007;25:2085–2094. doi: 10.1016/j.vaccine.2006.11.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Zaharoff DA, Rogers CJ, Hance KW, Schlom J, Greiner JW. Chitosan solution enhances the immunoadjuvant properties of GM-CSF. Vaccine. 2007;25:8673–8686. doi: 10.1016/j.vaccine.2007.10.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Scherliess R, Buske S, Young K, Weber B, Rades T, Hook S. In vivo evaluation of chitosan as an adjuvant in subcutaneous vaccine formulations. Vaccine. 2013;31:4812–4819. doi: 10.1016/j.vaccine.2013.07.081. [DOI] [PubMed] [Google Scholar]
- 125.Vasiliev YM. Chitosan-based vaccine adjuvants: incomplete characterization complicates preclinical and clinical evaluation. Expert Rev Vaccines. 2015;14:37–53. doi: 10.1586/14760584.2015.956729. [DOI] [PubMed] [Google Scholar]
- 126.Murano E. Use of natural polysaccharides in the microencapsulation techniques. J Appl Ichthyol. 1998;14:245–249. [Google Scholar]
- 127.Dobakhti F, Naghibi T, Taghikhani M, et al. Adjuvanticity effect of sodium alginate on subcutaneously injected BCG in BALB/c mice. Microbes Infect. 2009;11:296–301. doi: 10.1016/j.micinf.2008.12.003. [DOI] [PubMed] [Google Scholar]
- 128.Mata E, Igartua M, Patarroyo ME, Pedraz JL, Hernandez RM. Enhancing immunogenicity to PLGA microparticulate systems by incorporation of alginate and RGD-modified alginate. Eur J Pharm Sci. 2011;44:32–40. doi: 10.1016/j.ejps.2011.05.015. [DOI] [PubMed] [Google Scholar]
- 129.Dyakonova VA, Dambaeva SV, Pinegin BV, Khaitov RM. Study of interaction between the polyoxidonium immunomodulator and the human immune system cells. Int Immunopharmacol. 2004;4:1615–1623. doi: 10.1016/j.intimp.2004.07.015. [DOI] [PubMed] [Google Scholar]
- 130.Dambaeva SV, Mazurov DV, Golubeva NM, D'Yakonova VA, Pinegin BV, Khaitov RM. Effect of polyoxidonium on the phagocytic activity of human peripheral blood leukocytes. Russ J Immunol. 2003;8:53–60. [PubMed] [Google Scholar]
- 131.Denisov AA, Korobovtseva YS, Karpova OM, et al. Immunopotentiation of live brucellosis vaccine by adjuvants. Vaccine. 2010;28(Suppl 5):F17–F22. doi: 10.1016/j.vaccine.2010.03.054. [DOI] [PubMed] [Google Scholar]
- 132.Toptygina A, Semikina E, Alioshkin V. Influence of an immunopotentiator polyoxidonium on cytokine profile and antibody production in children vaccinated with Priorix. Arch Physiol Biochem. 2012;118:197–203. doi: 10.3109/13813455.2012.659669. [DOI] [PubMed] [Google Scholar]
- 133.Andrianov AK, Marin A, Roberts BE. Polyphosphazene polyelectrolytes: a link between the formation of noncovalent complexes with antigenic proteins and immunostimulating activity. Biomacromolecules. 2005;6:1375–1379. doi: 10.1021/bm049329t. [DOI] [PubMed] [Google Scholar]
- 134.Andrianov AK, Chen J, LeGolvan MP. Poly(dichlorophosphazene) as a precursor for biologically active polyphosphazenes: synthesis, characterization, and stabilization. Macromolecules. 2004;37:414–420. [Google Scholar]
- 135.Andrianov AK, LeGolvan MP. Characterization of poly [di(carboxylatophenoxy)-phosphazene] by an aqueous gel permeation chromatography. J Appl Polym Sci. 1996;60:2289–2295. [Google Scholar]
- 136.Andrianov AK, Svirkin YY, LeGolvan MP. Synthesis and biologically relevant properties of polyphosphazene polyacids. Biomacromolecules. 2004;5:1999–2006. doi: 10.1021/bm049745d. [DOI] [PubMed] [Google Scholar]
- 137.Andrianov AK, DeCollibus DP, Gillis HA, Kha HH, Marin A. Polyphosphazene immunoadjuvants for intradermal vaccine delivery. In: Andrianov AK, editor. Polyphosphazene for biomedical applications. Hoboken: John Wiley & Sons Inc.; 2009. pp. 101–116. [Google Scholar]
- 138.Andrianov AK, Decollibus DP, Marin A, Webb A, Griffin Y, Webby RJ. PCPP-formulated H5N1 influenza vaccine displays improved stability and dose-sparing effect in lethal challenge studies. J Pharm Sci. 2011;100:1436–1443. doi: 10.1002/jps.22367. [DOI] [PubMed] [Google Scholar]
- 139.Payne LG, Jenkins SA, Woods AL, et al. Poly[di(carboxylatophenoxy)phosphazene] (PCPP) is a potent immunoadjuvant for an influenza vaccine. Vaccine. 1998;16:92–98. doi: 10.1016/s0264-410x(97)00149-7. [DOI] [PubMed] [Google Scholar]
- 140.Payne LG, Van Nest G, Barchfeld GL, Siber GR, Gupta RK, Jenkins SA. PCPP as a parenteral adjuvant for diverse antigens. Dev Biol Stand. 1998;92:79–87. [PubMed] [Google Scholar]
- 141.Mutwiri G, Benjamin P, Soita H, et al. Poly[di(sodium carboxylatoethylphenoxy)phosphazene] (PCEP) is a potent enhancer of mixed Th1/Th2 immune responses in mice immunized with influenza virus antigens. Vaccine. 2007;25:1204–1213. doi: 10.1016/j.vaccine.2006.10.011. [DOI] [PubMed] [Google Scholar]
- 142.Lu Y, Salvato MS, Pauza CD, et al. Utility of SHIV for testing HIV-1 vaccine candidates in macaques. J Acquir Immune Defic Syndr Hum Retrovirol. 1996;12:99–106. doi: 10.1097/00042560-199606010-00001. [DOI] [PubMed] [Google Scholar]
- 143.Wu JY, Wade WF, Taylor RK. Evaluation of cholera vaccines formulated with toxin-coregulated pilin peptide plus polymer adjuvant in mice. Infect Immun. 2001;69:7695–7702. doi: 10.1128/IAI.69.12.7695-7702.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Le Cam NN, Ronco J, Francon A, Blondeau C, Fanget B. Adjuvants for influenza vaccine. Res Immunol. 1998;149:19–23. [Google Scholar]
- 145.Plotkin SA. Vaccines: past, present and future. Nat Med. 2005;11(4 Suppl):S5–11. doi: 10.1038/nm1209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Ison MG, Mills J, Openshaw P, Zambon M, Osterhaus A, Hayden F. Current research on respiratory viral infections: Fourth International Symposium. Antiviral Res. 2002;55:227–278. doi: 10.1016/S0166-3542(02)00055-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Kim JH, Kirsch EA, Gilliam B. Abstracts of the 37th Annual Meeting of the Infectious Diseases Society of America. Philadelphia, PA, USA: 1999. Nov 18 21, A phase I, open label, dose ranging trial of the Pasteur Merieux Connaught (PMC) oligomeric HIV-1 Gp160mn/LAI-2 vaccine in HIV seronegative adults. [Google Scholar]
- 148.Mutwiri G, Benjamin P, Soita H, Babiuk LA. Co-administration of polyphosphazenes with CpG oligodeoxynucleotides strongly enhances immune responses in mice immunized with hepatitis B virus surface antigen. Vaccine. 2008;26:2680–2688. doi: 10.1016/j.vaccine.2008.03.031. [DOI] [PubMed] [Google Scholar]
- 149.Istrate C, Hinkula J, Charpilienne A, et al. Parenteral administration of RF 8-2/6/7 rotavirus-like particles in a one-dose regimen induce protective immunity in mice. Vaccine. 2008;26:4594–4601. doi: 10.1016/j.vaccine.2008.05.089. [DOI] [PubMed] [Google Scholar]
- 150.Palmer CD, Ninkovic J, Prokopowicz ZM, et al. The effect of stable macromolecular complexes of ionic polyphosphazene on HIV Gag antigen and on activation of human dendritic cells and presentation to T-cells. Biomaterials. 2014;35:8876–8886. doi: 10.1016/j.biomaterials.2014.06.043. [DOI] [PubMed] [Google Scholar]
- 151.Andrianov AK, Sargent JR, Sule SS, et al. Synthesis, physico-chemical properties and immunoadjuvant activity of water-soluble phosphazene polyacids. J Bioact Compat Polym. 1998;13:243–256. [Google Scholar]
- 152.Andrianov AK, Marin A, Chen J. Synthesis, properties, and biological activity of poly[di(sodium carboxylatoethylphenoxy)phosphazene] Biomacromolecules. 2006;7:394–399. doi: 10.1021/bm050790a. [DOI] [PubMed] [Google Scholar]
- 153.Johansson E, Istrate C, Charpilienne A, et al. Amount of maternal rotavirus-specific antibodies influence the outcome of rotavirus vaccination of newborn mice with virus-like particles. Vaccine. 2008;26:778–785. doi: 10.1016/j.vaccine.2007.11.089. [DOI] [PubMed] [Google Scholar]
- 154.Awate S, Wilson HL, Lai K, Babiuk LA, Mutwiri G. Activation of adjuvant core response genes by the novel adjuvant PCEP. Mol Immunol. 2012;51:292–303. doi: 10.1016/j.molimm.2012.03.026. [DOI] [PubMed] [Google Scholar]
- 155.Awate S, Wilson HL, Singh B, Babiuk LA, Mutwiri G. The adjuvant PCEP induces recruitment of myeloid and lymphoid cells at the injection site and draining lymph node. Vaccine. 2014;32:2420–2427. doi: 10.1016/j.vaccine.2014.03.014. [DOI] [PubMed] [Google Scholar]
- 156.Andrianov AK, DeCollibus DP, Gillis HA, et al. Poly[di (carboxylatophenoxy)phosphazene] is a potent adjuvant for intradermal immunization. Proc Natl Acad Sci U S A. 2009;106:18936–18941. doi: 10.1073/pnas.0908842106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Andrianov AK, Marin A, DeCollibus DP. Microneedles with intrinsic immunoadjuvant properties: microfabrication, protein stability, and modulated release. Pharm Res. 2011;28:58–65. doi: 10.1007/s11095-010-0133-7. [DOI] [PubMed] [Google Scholar]
- 158.Vaccines Europe. The vaccine industry in figures [Internet]. Brussels: EFPIA/Vaccines Europe. 2014. [cited 2014 Nov 18]. Available from: http://www.vaccineseurope.eu/about-vaccines-europe/vaccines-europe-in-figures/