

Abstract
Mycobacterium xenopi is an opportunistic mycobacterial pathogen of increasing clinical importance. Surveillance of M. xenopi is hampered by the absence of tools for genotyping and molecular epidemiology. In this study, we describe the development and evaluation of an effective multilocus sequence typing strategy for M. xenopi.
INTRODUCTION
Mycobacterium xenopi is an opportunistic mycobacterial pathogen of increasing clinical importance. M. xenopi disease is most common among patients with underlying medical conditions (e.g., chronic obstructive pulmonary disease, bronchiectasis, and immunodeficiencies) (1–4). Cases usually present as tuberculosis-like pulmonary disease, but extrapulmonary (e.g., bone, joint, and muscle) infections have also been reported (5).
In North America, M. xenopi disease is not reportable, so the actual incidence of infection is unknown. However, in a recent global survey of nontuberculous mycobacteria (NTM), M. xenopi accounted for 8% of all NTM species isolated from pulmonary samples (6). Although rarely seen in Asia, Australia, or South America, M. xenopi is common in North America and Europe. Isolation frequency is highest in central Europe (e.g., Germany, Italy, Switzerland, Hungary, Croatia, Poland, and the Czech Republic), the English Channel region (e.g., southeastern England, northern France, and Belgium), and the Great Lakes region of Canada and the United States (6, 7). In Hungary, M. xenopi is the dominant NTM species and accounts for almost half of all NTM isolated from pulmonary samples (6). In Ontario, Canada, M. xenopi prevalence is second only to that of the Mycobacterium avium complex (8). Moreover, population-based studies indicate that, in Ontario, the prevalence of M. xenopi pulmonary disease has increased from 7.4 cases/100,000 persons during 1998 to 2002 to 9.5 cases/100,000 persons during 2006 to 2010 (9).
Surveillance of M. xenopi is hampered by the absence of effective tools for molecular epidemiology. Restriction fragment length polymorphism (RFLP) methods have been described but are not in routine use, and no public IS1081 or IS1395 RFLP databases exist (10–12). Moreover, RFLP is time-consuming and labor-intensive and, like other gel electrophoresis-based typing methods, suffers from poor interlab reproducibility and portability. In contrast, multilocus sequence typing (MLST) is readily portable and reproducible. MLST is a DNA sequencing-based method that exploits nucleotide variations present in predefined sets of genetic loci (13, 14). The traditional approach involves Sanger sequencing of PCR amplicons, but MLST is compatible with whole-genome sequencing technologies and, thus, should remain a viable method. MLST schemes have been developed for dozens of bacterial species and some pathogenic fungi. Although not widely used for Mycobacterium species, an MLST scheme for the Mycobacterium abscessus complex has recently been described (15, 16). In this study, we describe the development and evaluation of an effective MLST strategy for M. xenopi.
MATERIALS AND METHODS
Bacterial strains.
A collection of 106 M. xenopi isolates was assembled for this study. Clinical isolates (n = 104) were provided by the Public Health Ontario Laboratories (PHOL; n = 37), the Laboratoire National de Santé Publique du Québec (LSPQ; n = 22), the Wisconsin State Laboratory of Hygiene (WSLH; n = 27), and the Wadsworth Center, New York State Department of Health (WCNY; n = 18). The clinical strains include respiratory isolates cultured from specimens submitted to the source laboratories for routine mycobacterial testing as well as referral isolates sent to the laboratories for identification. Two independent aliquots of the M. xenopi type strain ATCC 19250T, provided by the LSPQ and WSLH, were also examined. All M. xenopi isolates were submitted to the PHOL for analysis.
DNA extraction.
M. xenopi isolates obtained from the PHOL, LSPQ, WSLH, and WCNY were subcultured on Lowenstein-Jensen slants (Bio-media Unlimited Ltd., Toronto, ON, Canada) and incubated at 40°C until sufficient growth was observed (4 to 6 weeks). DNA was extracted using a modified boiling lysis method (17). Briefly, a loopful of culture was resuspended in 200 μl buffer (10 mM Tris, 1 mM EDTA, pH 8), vortexed to disperse any clumps, and then heated at 95°C for 20 min. Samples were centrifuged to pellet cellular debris, and the DNA-containing supernatant was transferred to a fresh tube and stored at −20°C until required for downstream applications.
PCR.
Six genes (atpD, fusA1, glnA1, pheT, secA1, and topA) plus the 16S-23S rRNA internal transcribed spacer (ITS) region were used for MLST of M. xenopi. Primer pairs for each MLST target are listed in Table 1. PCR was performed with the HotStarTaq Plus master mix kit (Qiagen, Louisville, KY, USA). Each 20-μl amplification reaction mixture contained 10 μl of 2× HotStartMM master mix, 2 μl of Q solution, 2 μl of 10× CoralLoad dye buffer, 3 μl of nuclease-free water, 1 μl (10 nmol) of each primer, plus 1 μl of crude DNA lysate. PCR was performed for 35 cycles using the following conditions: denaturation (94°C for 45 s), annealing (57°C for 45 s), and extension (72°C for 1 min), with a final extension cycle at 72°C for 10 min. Agarose gel electrophoresis was used to confirm amplification of each MLST target and estimate amplicon concentration.
TABLE 1.
Oligonucleotide primers for MLST analysis of Mycobacterium xenopi
| Target | Sequence |
Size (bp) | Function | |
|---|---|---|---|---|
| Primer 1 | Primer 2 | |||
| atpD | 5′-CGGTGTTCTCCAAGGGCATC-3′ | 5′-TTGTCGGTGTTGTCGGCCTG-3′ | 499 | ATP synthase (subunit b) |
| fusA1 | 5′-GCATCATGGCGCACATCGAC-3′ | 5′-TTCATCTCGACCAGGTCGACGACG-3′ | 505 | Elongation factor G |
| glnA1 | 5′-ATTCTGCACCATGCGCCTTC-3′ | 5′-ACACGTCGAAGTACAGCGCG-3′ | 511 | Glutamine synthetase |
| pheT | 5′-TACAGCTGGCTGCGCGAAGT-3′ | 5′-ATGGCCAGGTGGAACACCACGT-3′ | 494 | Phenylalanyl-tRNA synthetase |
| secA1 | 5′-CGAGCACCTCTACGAGATGG-3′ | 5′-TTAGCGCTTCTTGGCCGATT-3′ | 502 | Preprotein translocase |
| topA | 5′-AAAACCCGCTCGCCCCAAAC-3′ | 5′-GTGCCTTGCGGGTCGTCTTCTT-3′ | 515 | DNA topoisomerase I |
| ITS | 5′-TTGTACACACCGCCCGTCA-3′ | 5′-TCTCGATGCCAAGGCATCCACC-3′ | 445 | 16S-23S rRNA internal transcribed spacer |
DNA sequencing.
The BigDye Terminator v3.1 cycle sequencing kit (Life Technologies Inc., Burlington, ON, Canada) was used for DNA sequencing of amplicons for MLST. Labeling reactions were performed according to the manufacturer's instructions (Life Technologies). For each MLST target, bidirectional sequencing was performed. Unincorporated dyes were removed by ethanol-EDTA precipitation, and the labeled amplicons were resuspended in 10 μl of nuclease-free water. DNA sequencing was performed via capillary electrophoresis with an Applied Biosystems 3730xl DNA Analyzer. Editing, analysis, and alignment of DNA sequences were performed with MEGA 5.2 (18).
RESULTS
MLST.
DNA sequencing results for the MLST targets are summarized in Table 2 and also in Table S1 in the supplemental material. Individual genes were found to have two (atpD), three (fusA1 and topA), four (glnA1 and pheT), or five (secA1) allelic variants. All single nucleotide polymorphisms (SNPs) associated with the atpD, fusA1, glnA1, and topA1 alleles were synonymous. However, the pheT ST-2, pheT ST-3, secA1 ST-3, and secA1 ST-5 variants contained nonsynonymous SNPs. The six-locus MLST strategy (MLST-6) divided the 106 M. xenopi isolates into 15 distinct sequence types (STs). The MLST profile shared by the two aliquots of ATCC 19250T was designated ST-1. Most STs were observed only once or twice, but three STs were prevalent among the clinical isolates: ST-2 (n = 48; 46.1%), ST-5 (n = 23; 22.1%), and ST-9 (n = 20; 19.2%).
TABLE 2.
MLST analysis of Mycobacterium xenopi
| MLST-6 ST | No. of variants |
ITS | No. of isolatesa | |||||
|---|---|---|---|---|---|---|---|---|
| atpD | fusA1 | glnA1 | pheT | secA1 | topA | |||
| ST-1 | 1 | 1 | 1 | 1 | 1 | 1 | AA | 2 |
| ST-2 | 1 | 1 | 1 | 1 | 2 | 1 | Variableb | 48 |
| ST-3 | 1 | 1 | 1 | 1 | 3 | 1 | AB | 2 |
| ST-4 | 1 | 1 | 1 | 1 | 1 | 2 | CC | 1 |
| ST-5 | 1 | 1 | 1 | 1 | 2 | 2 | Variablec | 23 |
| ST-6 | 1 | 1 | 2 | 1 | 2 | 1 | BC | 1 |
| ST-7 | 1 | 1 | 3 | 1 | 2 | 2 | CC | 1 |
| ST-8 | 1 | 1 | 3 | 1 | 2 | 3 | CC | 1 |
| ST-9 | 1 | 1 | 2 | 2 | 2 | 1 | AAd | 20 |
| ST-10 | 1 | 1 | 4 | 3 | 4 | 1 | AC | 1 |
| ST-11 | 1 | 1 | 2 | 2 | 2 | 2 | AA | 1 |
| ST-12 | 1 | 1 | 4 | 4 | 5 | 3 | CC | 1 |
| ST-13 | 1 | 2 | 1 | 1 | 2 | 2 | Variablee | 2 |
| ST-14 | 1 | 3 | 1 | 1 | 2 | 1 | AB | 1 |
| ST-15 | 2 | 1 | 2 | 2 | 2 | 1 | AA | 1 |
Duplicate isolates (i.e., genotypically identical isolates from individual patients [Table 3]) were counted only once.
ST-2 isolates were associated with multiple ITS patterns: AA (n = 3), AB (n = 26), BB (n = 4), BC (n = 9), and nontypeable (n = 6).
ST-5 isolates were associated with multiple ITS patterns: BB (n = 1), BC (n = 16), CC (n = 4), and nontypeable (n = 2).
The ITS of one ST-9 isolate was nontypeable.
ST-13 isolates were associated with two ITS patterns: AB (n = 1) and BB (n = 1).
Previous studies have documented the existence of three distinct M. xenopi 16S-23S rRNA ITS alleles (i.e., Mxe-A, Mxe-B, and Mxe-C) (19). As such, the ITS region was selected as a seventh MLST target. However, for some isolates, sequencing of this target was confounded by the presence of multiple, nonidentical ITS sequences. Whereas genomes from the majority of “slow-growing” mycobacterial species feature a single rRNA operon, the M. xenopi genome contains two. Because the ITS PCR did not use operon-specific primers, the two ITS loci were amplified and sequenced together. Thus, homozygous strains (e.g., ITS A/A) generated “clean” sequences but heterozygous strains (e.g., ITS A/B) generated “mixed” electropherograms. By comparing the mixed electropherograms to published ITS sequences, it was possible to identify characteristic mixed-base patterns for heterozygous allele combinations (e.g., ITS A/B, ITS A/C, and ITS B/C) and assign ITS profiles to all but nine isolates. The seven-locus MLST strategy (MLST-7; MLST-6 plus ITS) divided the remaining 97 M. xenopi isolates into 21 STs (Table 2). In most cases, strains that clustered by six-locus MLST also clustered by ITS. For example, all isolates in the ST-9 group shared the same ITS A/A profile. In contrast, ST-2 was subdivided into four ITS clusters (A/A = 3, A/B = 26, B/B = 4, B/C = 9), ST-5 was subdivided into three clusters (B/B = 1, B/C = 16, C/C = 4), and the ST-13 pair included one A/B and one B/B isolate.
MLST of consecutive isolates from individual patients.
Table 3 includes MLST-6 results for a subset of PHOL strains: 17 consecutive isolates recovered from 7 patients. For five patients, only M. xenopi isolates of identical ST were recovered. However, the isolates from patient F were distinct (ST-11 in 2011 and ST-5 in 2012) and the four isolates recovered from patient G included one of ST-2 and three of ST-9.
TABLE 3.
MLST-6 analysis of consecutive isolates from individual patients
| Patient | Isolate | Date (mo/yr) | MLST-6 ST |
|---|---|---|---|
| A | 1 | 10/2010 | ST-5 |
| 2 | 11/2011 | ST-5 | |
| B | 1 | 08/2010 | ST-15 |
| 2 | 08/2012 | ST-15 | |
| C | 1 | 09/2010 | ST-5 |
| 2 | 10/2010 | ST-5 | |
| D | 1 | 09/2012 | ST-2 |
| 2 | 11/2012 | ST-2 | |
| E | 1 | 2009 | ST-5 |
| 2 | 10/2010 | ST-5 | |
| 3 | 01/2011 | ST-5 | |
| F | 1 | 10/2010 | ST-11 |
| 2 | 01/2012 | ST-5 | |
| G | 1 | 02/2012 | ST-2 |
| 2 | 04/2010 | ST-9 | |
| 3 | 08/2012 | ST-9 | |
| 4 | 10/2012 | ST-9 |
Geographic distribution of STs.
The geographic distribution of MLST-6-defined STs is summarized in Fig. 1 and also in Fig. S1 in the supplemental material. Most STs were observed only once or, in the case of the ST-3 and ST-13 pairs, were isolated by a single source laboratory. ST-9 strains were isolated at both the PHOL and LSPQ, and ST-5 strains were isolated at three facilities, but only ST-2 strains were isolated by all four source laboratories.
FIG 1.
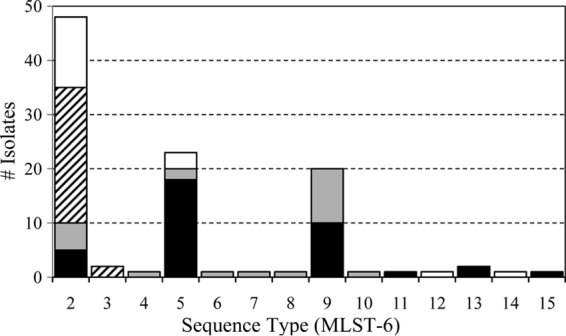
Geographic distribution of Mycobacterium xenopi MLST-6 sequence types. Isolates from PHOL (black), LSPQ (gray), WSLH (hatched), and WCNY (white) are shown.
DISCUSSION
Despite increasing recognition of M. xenopi as an important human pathogen, little is known about the genetic diversity of this species. MLST is a sequencing-based method that has been used for genotyping of many bacterial species. Traditionally, MLST strategies target “housekeeping” or “core” genes, loci that are conserved within the species of interest. For typing to be effective, these targets must also be heterogeneous, such that distinct isolates exhibit distinct sequence polymorphisms. To develop an MLST strategy for M. xenopi, we initially targeted 19 housekeeping genes: alaS, atpD, dnaK, ffh, ftsK, fum, fusA1, gap, gcvB, glnA1, gyrB, hsp65, infB, leuS, pheT, tpi, rpoB, secA1, and topA. Preliminary sequencing results, derived from a panel of 21 M. xenopi isolates, revealed that 13 of these loci were invariant and thus unsuitable for MLST (20).
The MLST-6 approach used in this study targets six polymorphic M. xenopi loci: atpD, fusA1, glnA1, pheT, secA1, and topA. The MLST-7 strategy includes one additional target, the M. xenopi ITS region. The two approaches were used to genotype a collection of 106 M. xenopi isolates provided by four mycobacteriology laboratories (Table 2 and Fig. 1; see also Fig. S1 in the supplemental material). The MLST-6 strategy divided the collection into 15 STs. The MLST-7 strategy provided greater resolution (21 STs) even though genotyping could not be completed for nine isolates. Despite the potential utility of the MLST-7 strategy, technical challenges, especially the requirement for interpretation of “mixed” ITS electropherograms, make the current MLST-7 approach less suitable for routine use than MLST-6. However, if the Sanger sequencing method used here was replaced by a next-generation approach, it would be possible to resolve distinct ITS loci and type additional targets. Tools to facilitate such analysis, sometimes called core genome multilocus sequence typing (cgMLST), have recently been described for M. tuberculosis (21).
For patients with multiple positive cultures or multiple episodes of disease, MLST can be used to distinguish cases of relapse from those of reinfection. MLST-6 analysis of 17 consecutive isolates obtained from 7 patients is summarized in Table 3. In six of the seven cases, multiple isolates of identical ST were obtained. These STs persisted over a period of months (patients C, D, and G) to 2 (patient A) and 3 (patients B and E) years. The two isolates from patient F were of different STs and isolated in different years, which is consistent with a case of reinfection. However, in the case of patient G, isolates of ST-2 and ST-9 were obtained within a few months of one another. Since subsequent isolates were also of ST-2, the situation may be indicative of strain replacement. Or, it may represent a polyclonal infection, a phenomenon that is common among patients with infections due to other nontuberculous mycobacteria, such as M. avium (22, 23).
MLST analysis demonstrates that M. xenopi isolates are genotypically heterogeneous. Nine of the MLST-6-defined STs, and 12 of the MLST-7-defined STs, were observed only once (Table 2 and Fig. 1). Because M. xenopi is an “environmental” mycobacterial species, it is possible that this genotypic diversity is related to geographic distribution. Although ST-2 strains were isolated by all four source laboratories, the distribution of other STs was restricted. For example, ST-9 strains were isolated only by the Canadian laboratories, while both ST-3 isolates were collected by WSLH. Most of the PHOL isolates were obtained from patients living in the greater Toronto area (GTA) of southern Ontario, but the genotypically distinct ST-15 strain was isolated from a patient in central Ontario. Analysis of the PHOL isolates suggests a geographic trend, with distribution of the ST-5, ST-9, and ST-2 strains restricted to the east/central, north/central, and west regions of the GTA, respectively. Additional isolates and environmental sampling are required to confirm this observation, but M. xenopi is typically associated with aquatic ecosystems of both natural and anthropogenic origin (24, 25). The geographic distribution of STs in the GTA may overlap patient exposure to specific engineered (e.g., domestic water reservoirs, distribution systems, and hot tubs) or natural (e.g., lakes, rivers, and aquifers) water sources.
The ST-1 profile assigned to M. xenopi type strain ATCC 19250T was not shared by any of the clinical isolates. Most likely, this is due to the fact that the type strain is from a different place and time (i.e., England, ca. 1958) (26). However, M. xenopi ATCC 19250T was isolated from the skin of an amphibian (Xenopus laevis), not the respiratory tract of a mammal. As such, the ST differences may be indicative of differences in the pathogenic potential and host preferences of genotypically distinct M. xenopi populations. Among other mycobacterial species, including the M. tuberculosis complex and M. avium complex, lineage-specific differences in pathogenicity and host range are well documented (27–29).
Despite its prevalence and increasing clinical importance, little is known about the genetic diversity and molecular epidemiology of M. xenopi. Gel electrophoresis-based methods are widely considered the gold standard for mycobacterial typing, and RFLP can be used to differentiate M. xenopi isolates. Unfortunately, for many laboratories, RFLP is not a practical option for routine typing. In contrast, in an era in which labor is expensive but sequencing is cheap, MLST may be a faster and more affordable alternative. With the development of an effective MLST scheme, it becomes possible to routinely examine the distribution, evolution, and population structure of M. xenopi. Future studies may provide insight into the geographic preferences of specific STs or uncover associations between M. xenopi genotype and disease phenotype, antibiotic susceptibility, and pathogenic potential.
Supplementary Material
ACKNOWLEDGMENTS
We acknowledge the skill and dedication of the laboratory staff from PHOL, LSPQ, WLSH, and WCNY responsible for the initial isolation and cultivation of the M. xenopi isolates used in the study.
Footnotes
Published ahead of print 10 September 2014
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JCM.01601-14.
REFERENCES
- 1.Griffith DE, Aksamit T, Brown-Elliott BA, Catanzaro A, Daley C, Gordin F, Holland SM, Horsburgh R, Huitt G, Iademarco MF, Iseman M, Olivier K, Ruoss S, von Reyn CF, Wallace RJ, Jr, Winthrop K, ATS Mycobacterial Diseases Subcommittee, American Thoracic Society, Infectious Disease Society of America 2007. An official ATS/IDSA statement: diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 175:367–416. 10.1164/rccm.200604-571ST. [DOI] [PubMed] [Google Scholar]
- 2.Marusic A, Katalinic-Jankovic V, Popovic-Grle S, Jankovic M, Mazuranic I, Puljic I, Sertic Milic H. 2009. Mycobacterium xenopi pulmonary disease—epidemiology and clinical features in non-immunocompromised patients. J. Infect. 58:108–112. 10.1016/j.jinf.2009.01.001. [DOI] [PubMed] [Google Scholar]
- 3.van Ingen J, Boeree MJ, de Lange WC, Hoefsloot W, Bendien SA, Magis-Escurra C, Dekhuijzen R, van Soolingen D. 2008. Mycobacterium xenopi clinical relevance and determinants, the Netherlands. Emerg. Infect. Dis. 14:385–389. 10.3201/eid1403.061393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Varadi RG, Marras TK. 2009. Pulmonary Mycobacterium xenopi infection in non-HIV-infected patients: a systematic review. Int. J. Tuberc. Lung Dis. 13:1210–1218. [PubMed] [Google Scholar]
- 5.Meybeck A, Fortin C, Abgrall S, Adle-Biassette H, Hayem G, Ruimy R, Yeni P. 2005. Spondylitis due to Mycobacterium xenopi in a human immunodeficiency virus type 1-infected patient: case report and review of the literature. J. Clin. Microbiol. 43:1465–1466. 10.1128/JCM.43.3.1465-1466.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hoefsloot W, van Ingen J, Andrejak C, Angeby K, Bauriaud R, Bemer P, Beylis N, Boeree MJ, Cacho J, Chihota V, Chimara E, Churchyard G, Cias R, Daza R, Daley CL, Dekhuijzen PN, Domingo D, Drobniewski F, Esteban J, Fauville-Dufaux M, Folkvardsen DB, Gibbons N, Gomez-Mampaso E, Gonzalez R, Hoffmann H, Hsueh PR, Indra A, Jagielski T, Jamieson F, Jankovic M, Jong E, Keane J, Koh WJ, Lange B, Leao S, Macedo R, Mannsaker T, Marras TK, Maugein J, Milburn HJ, Mlinko T, Morcillo N, Morimoto K, Papaventsis D, Palenque E, Paez-Pena M, Piersimoni C, Polanova M, Rastogi N, Richter E, Ruiz-Serrano MJ, Silva A, da Silva MP, Simsek H, van Soolingen D, Szabo N, Thomson R, Tortola Fernandez T, Tortoli E, Totten SE, Tyrrell G, Vasankari T, Villar M, Walkiewicz R, Winthrop KL, Wagner D, Nontuberculous Mycobacteria Network European Trials Group 2013. The geographic diversity of nontuberculous mycobacteria isolated from pulmonary samples: an NTM-NET collaborative study. Eur. Respir. J. 42:1604–1613. 10.1183/09031936.00149212. [DOI] [PubMed] [Google Scholar]
- 7.Martin-Casabona N, Bahrmand AR, Bennedsen J, Thomsen VO, Curcio M, Fauville-Dufaux M, Feldman K, Havelkova M, Katila ML, Koksalan K, Pereira MF, Rodrigues F, Pfyffer GE, Portaels F, Urgell JR, Rusch-Gerdes S, Tortoli E, Vincent V, Watt B, Spanish Group for Non-Tuberculosis Mycobacteria 2004. Non-tuberculous mycobacteria: patterns of isolation. A multi-country retrospective survey. Int. J. Tuberc. Lung Dis. 8:1186–1193. [PubMed] [Google Scholar]
- 8.Marras TK, Chedore P, Ying AM, Jamieson F. 2007. Isolation prevalence of pulmonary non-tuberculous mycobacteria in Ontario, 1997–2003. Thorax 62:661–666. 10.1136/thx.2006.070797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Marras TK, Mendelson D, Marchand-Austin A, May K, Jamieson FB. 2013. Pulmonary nontuberculous mycobacterial disease, Ontario, Canada, 1998–2010. Emerg. Infect. Dis. 19:1889–1891. 10.3201/eid1911.130737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Collins DM. 1994. DNA fingerprinting of Mycobacterium xenopi. Lett. Appl. Microbiol. 18:234–235. 10.1111/j.1472-765X.1994.tb00855.x. [DOI] [Google Scholar]
- 11.Lenart K, Sajduda A, Augustynowicz-Kopec E, Dziadek J. 2011. Profiling Mycobacterium xenopi with restriction fragment length polymorphism of insertion element IS1395. Int. J. Infect. Dis. 15:e321–e325. 10.1016/j.ijid.2010.10.008. [DOI] [PubMed] [Google Scholar]
- 12.Picardeau M, Varnerot A, Rauzier J, Gicquel B, Vincent V. 1996. Mycobacterium xenopi IS1395, a novel insertion sequence expanding the IS256 family. Microbiology 142:2453–2461. 10.1099/00221287-142-9-2453. [DOI] [PubMed] [Google Scholar]
- 13.Maiden MC. 2006. Multilocus sequence typing of bacteria. Annu. Rev. Microbiol. 60:561–588. 10.1146/annurev.micro.59.030804.121325. [DOI] [PubMed] [Google Scholar]
- 14.Perez-Losada M, Cabezas P, Castro-Nallar E, Crandall KA. 2013. Pathogen typing in the genomics era: MLST and the future of molecular epidemiology. Infect. Genet. Evol. 16:38–53. 10.1016/j.meegid.2013.01.009. [DOI] [PubMed] [Google Scholar]
- 15.Kim SY, Kang YA, Bae IK, Yim JJ, Park MS, Kim YS, Kim SK, Chang J, Jeong SH. 2013. Standardization of multilocus sequence typing scheme for Mycobacterium abscessus and Mycobacterium massiliense. Diagn. Microbiol. Infect. Dis. 77:143–149. 10.1016/j.diagmicrobio.2013.06.023. [DOI] [PubMed] [Google Scholar]
- 16.Macheras E, Konjek J, Roux AL, Thiberge JM, Bastian S, Leao SC, Palaci M, Sivadon-Tardy V, Gutierrez C, Richter E, Rusch-Gerdes S, Pfyffer GE, Bodmer T, Jarlier V, Cambau E, Brisse S, Caro V, Rastogi N, Gaillard JL, Heym B. 2014. Multilocus sequence typing scheme for the Mycobacterium abscessus complex. Res. Microbiol. 165:82–90. 10.1016/j.resmic.2013.12.003. [DOI] [PubMed] [Google Scholar]
- 17.Turenne CY, Tschetter L, Wolfe J, Kabani A. 2001. Necessity of quality-controlled 16S rRNA gene sequence databases: identifying nontuberculous Mycobacterium species. J. Clin. Microbiol. 39:3637–3648. 10.1128/JCM.39.10.3638-3648.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28:2731–2739. 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Roth A, Fischer M, Hamid ME, Michalke S, Ludwig W, Mauch H. 1998. Differentiation of phylogenetically related slowly growing mycobacteria based on 16S-23S rRNA gene internal transcribed spacer sequences. J. Clin. Microbiol. 36:139–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mirza S, Farrell DJ, Marras TK, Ma JH, Liu D, Alexander DC, Kus JV, Jamieson FB. 2013. Sequence-based typing of clinical isolates of Mycobacterium xenopi isolated from Ontario, Canada. J. Undergrad. Life Sci. 7:41–45. [Google Scholar]
- 21.Kohl TA, Diel R, Harmsen D, Rothganger J, Walter KM, Merker M, Weniger T, Niemann S. 2014. Whole-genome-based Mycobacterium tuberculosis surveillance: a standardized, portable, and expandable approach. J. Clin. Microbiol. 52:2479–2486. 10.1128/JCM.00567-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fujita K, Ito Y, Hirai T, Kubo T, Maekawa K, Togashi K, Ichiyama S, Mishima M. 2014. Association between polyclonal and mixed mycobacterial Mycobacterium avium complex infection and environmental exposure. Ann. Am. Thorac. Soc. 11:45–53. 10.1513/AnnalsATS.201309-297OC. [DOI] [PubMed] [Google Scholar]
- 23.Wallace RJ, Jr, Zhang Y, Brown BA, Dawson D, Murphy DT, Wilson R, Griffith DE. 1998. Polyclonal Mycobacterium avium complex infections in patients with nodular bronchiectasis. Am. J. Respir. Crit. Care Med. 158:1235–1244. 10.1164/ajrccm.158.4.9712098. [DOI] [PubMed] [Google Scholar]
- 24.Sebakova H, Kozisek F, Mudra R, Kaustova J, Fiedorova M, Hanslikova D, Nachtmannova H, Kubina J, Vraspir P, Sasek J. 2008. Incidence of nontuberculous mycobacteria in four hot water systems using various types of disinfection. Can. J. Microbiol. 54:891–898. 10.1139/W08-080. [DOI] [PubMed] [Google Scholar]
- 25.Vaerewijck MJ, Huys G, Palomino JC, Swings J, Portaels F. 2005. Mycobacteria in drinking water distribution systems: ecology and significance for human health. FEMS Microbiol. Rev. 29:911–934. 10.1016/j.femsre.2005.02.001. [DOI] [PubMed] [Google Scholar]
- 26.Schwabacher H. 1959. A strain of Mycobacterium isolated from skin lesions of a cold-blooded animal, Xenopus laevis, and its relation to atypical acid-fast bacilli occurring in man. J. Hyg. (Lond.) 57:57–67. 10.1017/S0022172400019896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Manabe YC. 2002. Naturally attenuated, orally administered Mycobacterium microti as a tuberculosis vaccine is better than subcutaneous Mycobacterium bovis BCG. Infect. Immun. 70:1566–1570. 10.1128/IAI.70.3.1566-1570.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Perez-Lago L, Navarro Y, Garcia-de-Viedma D. 2013. Current knowledge and pending challenges in zoonosis caused by Mycobacterium bovis: a review. Res. Vet. Sci. 10.1016/j.rvsc.2013.11.008. [DOI] [PubMed] [Google Scholar]
- 29.Turenne CY, Alexander DC. 2010. Mycobacterium avium complex, p 60–71 In Behr MA, Collins DM. (ed), Paratuberculosis: organism, disease, control. CAB International, Wallingford, Oxfordshire, United Kingdom. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.