

Abstract
Exercise is a well-established tool to prevent and combat type 2 diabetes. Exercise improves whole body metabolic health in people with type 2 diabetes, and adaptations to skeletal muscle are essential for this improvement. An acute bout of exercise increases skeletal muscle glucose uptake, while chronic exercise training improves mitochondrial function, increases mitochondrial biogenesis, and increases the expression of glucose transporter proteins and numerous metabolic genes. This review focuses on the molecular mechanisms that mediate the effects of exercise to increase glucose uptake in skeletal muscle.
Keywords: exercise, type 2 diabetes
diabetes is a complex disease that affects millions of people worldwide. There are 23.6 million people in the United States or ∼8% of the population that have diabetes, a number that has doubled over the last 15 yr and is continuing to increase at epidemic rates (56). It is predicted that 1 in 3 adults in the United States will have diabetes by 2050, yielding enormous tolls at individual, public health, and economic levels (56).
Type 2 diabetes arises from a combination of genetic susceptibility and environmental factors including physical inactivity and poor nutrition (25). Obesity plays a role in the majority of cases of type 2 diabetes (7a). The progression of obesity increases the risk for the development of type 2 diabetes; as a person becomes more obese, they enter a more insulin-resistant state, leading to impaired glucose tolerance, which can potentially lead to the onset of type 2 diabetes. As the disease progresses, the risk of complications, such as retinopathy, nephropathy, and neuropathy, increases. In fact, type 2 diabetes is the leading cause of blindness, kidney failure, and amputations and a major cause of heart attacks and strokes (56).
In people with type 2 diabetes, insulin levels are normal or high, but tissues such as liver, skeletal muscle, and adipose tissue become resistant to insulin. The pancreas compensates by producing large amounts of insulin, and this overproduction of insulin can eventually fail. This increase in circulating insulin can result in impaired glucose transport into liver, skeletal muscle, and adipose tissue (11). While type 2 diabetes is usually adult onset, the number of children and adolescents afflicted by this disease is increasing, likely as a result of increased rates of obesity and poor nutrition and an increasingly sedentary lifestyle.
Exercise Training Improves Metabolic Health in People With Type 2 Diabetes
While rates of diabetes are on the rise, it has long been recognized that exercise has important health benefits for people with type 2 diabetes. One of the environmental factors considered a risk for the development of insulin resistance and type 2 diabetes is a lack of physical activity, and regular physical exercise can delay or prevent the onset of this disease (18, 58, 78).
Randomized trials have found that lifestyle interventions including ∼150 min/wk of physical activity, combined with diet-induced weight loss, reduced the risk of type 2 diabetes by 58% in an at-risk population (58, 78). Interventions of exercise alone have proved to be just as effective in terms of prevention of the progression of type 2 diabetes as programs of diet alone or diet and exercise combined (9). Exercise training in type 2 diabetic patients improves management of blood glucose levels, body weight, lipids, blood pressure, cardiovascular disease, mortality, and overall quality of life (13, 19, 38, 47, 48, 62, 82). The more recent Look AHEAD study has shown that combined weight loss and physical activity in people with type 2 diabetes resulted in modest weight loss of ∼6%, improved glycolated hemoglobin, improved mobility, and improved kidney function, but no improvement in cardiovascular disease over a 10-yr period (13, 47, 48, 62). However, since the level of fitness was only assessed through year 4 of the study, conclusions on the effects of fitness level on cardiovascular disease cannot be made (13, 47, 48, 62). Another recent study (23) has shown that 6 mo of moderate-intensity exercise training decreased visceral fat mass and decreased hepatic triglyceride content in people with type 2 diabetes and that this program of exercise alone was more effective than programs of diet alone. Another recent study (19) showed that increasing physical activity in adults with type 2 diabetes resulted in partial or complete remission of type 2 diabetes in 11.5% of subjects within the first year of intervention and an additional 7% had partial or complete remission of type 2 diabetes after 4 yr of exercise intervention. Complete remission was defined by glucose normalization without need for drugs, and partial remission was defined as a transition to prediabetic or normal glucose levels without drug treatment (19). Taken together, these data demonstrate the beneficial effects of exercise training to combat type 2 diabetes.
One of the most well-established mechanisms through which type 2 diabetics improve metabolic health with exercise is through adaptations to skeletal muscle, which, in turn, decreases skeletal muscle insulin resistance. Here, we will discuss the effects of exercise on skeletal muscle, because skeletal muscle is responsible for the majority of glucose uptake in the postprandial state (6, 11). In the following sections, we will discuss specific adaptations of skeletal muscle to both acute and exercise training on skeletal muscle glucose uptake and metabolism.
Effects of Acute Exercise on Skeletal Muscle Glucose Uptake
It is well established that insulin is a potent simulator of glucose transport in skeletal muscle. In people with type 2 diabetes, insulin-stimulated glucose uptake in skeletal muscle is impaired. However, exercise-stimulated glucose uptake in people with type 2 diabetes is normal or at near normal levels (51). Because exercise-stimulated glucose uptake is normal in people with type 2 diabetes, defining insulin-independent mechanisms in the control of exercise-stimulated skeletal muscle glucose uptake is of critical importance as a potential means to treat diabetes. During the last several years, researchers have learned much about the signaling mechanisms that regulate exercise-induced glucose transport. There are many lines of evidence that show that exercise activates molecular signals that bypass defects in insulin action in skeletal muscle.
Both insulin and exercise increase skeletal muscle glucose uptake by translocation of glucose transporter 4 (GLUT4), the predominant GLUT in muscle, from an intracellular location to the plasma membrane. Insulin and exercise stimulate GLUT4 translocation through distinct signaling mechanisms. Insulin signaling involves rapid phosphorylation of the insulin receptor, insulin receptor substrate-1/2 on tyrosine residues, and the activation of phosphatidylinositol 3-kinase (14, 17). Exercise, however, has no effect on insulin receptor and insulin receptor substrate-1/2 tyrosine phosphorylation or on phosphatidylinositol 3-kinase activity (17, 75). In fact, mice that lack insulin receptors in skeletal muscle [muscle-specific insulin receptor knockout (KO) mice] have normal exercise-stimulated glucose uptake (85). These data clearly demonstrate that insulin and exercise mediate GLUT4 translocation in skeletal muscle through distinct proximal signaling mechanisms.
Acute exercise activates multiple signaling pathways, but the activated signaling pathways necessary for increased glucose uptake and GLUT4 translocation are not well understood. Muscle contraction involves changes in energy status (i.e., increased AMP/ATP), increases in intracellular Ca2+ concentration, increased ROS, and PKC. These changes activate various signaling cascades, some of which likely work to phosphorylate Tre-2/USP6, BUB2, cdc16 domain family member 1 (TBC1D1) and Akt substrate of 160 kDa (AS160) and activate GLUT4 translocation. Here, we will discuss some of the various signaling cascades that have been implicated in exercise-stimulated glucose uptake (Fig. 1) (63, 66).
Fig. 1.
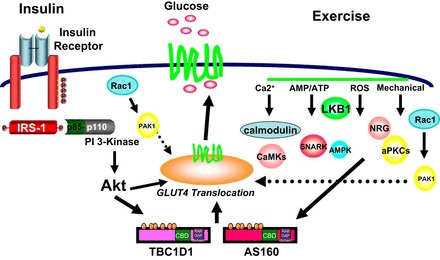
Exercise and insulin regulation of glucose transport. A proposed model for the signaling pathways mediating exercise- and insulin-induced skeletal muscle glucose transport is shown. IRS-1, insulin receptor substrate-1; PAK, p21 protein (Cdc42/Rac)-activated kinase 1; LKB1, liver kinase B1; PI3K, phosphatidylinositol 3-kinase; CaMK, Ca2+/calmodulin-dependent protein kinase; SNARK, sucrose nonfermenting AMP-dependent protein kinase (AMPK)-related kinase; NRG, neuroglian; aPKCs, atypical PKCs; GLUT, glucose transporter; TBC1D1, Tre-2/USP6, BUB2, cdc16 domain family member 1; AS160, Akt substrate of 160 kDa; CBD, calmodulin-binding domain. [Adapted from Ref. 66.]
AMP-Activated Protein Kinase
AMP-activated protein kinase (AMPK) and its primary upstream kinase, liver kinase B1 (LKB1), are the most widely studied proteins implicated in skeletal muscle glucose transport in response to exercise. AMPK is a heterotrimeric protein composed of a catalytic α-subunit and regulatory β- and γ-subunits. The α- and β-subunits each exist in two isoforms (α1, α2, β1, and β2), and the γ-subunit exists in three isoforms (γ1, γ2, and γ3). AMPK is activated by phosphorylation by one or more upstream kinases, including LKB1 (20, 39). A previous study (53) involving incubation with the AMP-analog 5-aminoimidazole-4-carboxamide ribonucleoside (AICAR) have shown that activation of AMPK is positively correlated with increased skeletal muscle glucose uptake. This increase in glucose uptake by AICAR stimulation is lost in mouse models deficient in AMPK α2- or γ3-subunits (10, 33). Some studies (1, 28, 30, 33, 57, 69) have shown that mice overexpressing a dominant negative AMPK-α2 construct in muscle or AMPK-α1 and -β KO mice have impaired exercise-stimulated glucose uptake. However, other studies using mouse models with ablated AMPK activity have demonstrated that inhibition of AMPK has little or no effect on exercise-induced glucose uptake (15, 33, 55) or exercise-stimulated glucose uptake in vivo (50). Thus, whether AMPK is necessary for exercise-stimulated glucose uptake is not fully understood. In a mouse muscle-specific LKB1 KO model, AMPK-α2 activation was completely inhibited and exercise-stimulated glucose uptake was severely blunted (41, 67). This impairment could be due to decreased activation of AMPK and one or more of the AMPK-related kinases that are substrates of LKB1, for example, sucrose nonfermenting AMPK-related kinase, which is involved in exercise-induced glucose uptake (42). However, the role of LKB-1 in exercise-stimulated glucose uptake is debatable (31). Whereas previous studies (41, 67) have shown that exercise-stimulated glucose uptake was impaired in LKB-1 KO mice, another recent study (31) showed that glucose uptake during treadmill running was similar, if not higher, in LKB-1 KO mice compared with wild-type control mice. An additional study (23) has also shown that muscle-specific deletion of LKB1 only partially inhibits exercise-stimulated glucose transport. These data suggest that while AMPK and LKB1 are important in the regulation of exercise-stimulated glucose uptake, there are several potential mechanisms involved in the control of exercise-stimulated glucose transport in skeletal muscle.
Ca2+/Calmodulin-Dependent Protein Kinases
A fundamental part of skeletal muscle contraction is the increase in intracellular Ca2+ concentration. More recently, studies have indicated Ca2+/calmodulin signaling and Ca2+/calmodulin-dependent protein kinases (CaMKs) as critical components of Ca2+- and exercise-stimulated skeletal muscle glucose uptake. Incubation of rat skeletal muscle with the Ca2+/calmodulin inhibitor KN-93 decreased skeletal muscle glucose transport in response to contraction (86). Incubation with KN-93 also significantly inhibited exercise-induced CaMKII phosphorylation in the absence of AMPK inhibition. This suggests that CaMKs regulate glucose uptake independently of AMPK signaling (83, 86). These studies also showed that overexpression of constitutively active CaMKKα in mouse skeletal muscle increased AMPK Thr172 phosphorylation and skeletal muscle glucose uptake (83). However, this increase in glucose uptake was also observed in muscle overexpressing dead AMPK-α2 and thus is likely independent of AMPK activation. In contrast, another study (30) using isolated skeletal muscle showed that inhibition of CaMK signaling with KN-93 inhibited contraction-induced skeletal muscle glucose uptake through an AMPK-dependent signaling pathway. These data suggest that CaMKs play important roles in the regulation of contraction-induced skeletal muscle glucose uptake, but the role of AMPK in the regulation of Ca2+/calmodulin-mediated increases in skeletal muscle glucose uptake is unclear. Recently, electroporation of a specific CaMKII inhibitor into mouse tibialis anterior muscle reduced exercise-stimulated glucose uptake by 30% (84). However, a separate study (29) found that increases in Ca2+ concentration in muscle caused very little increase in glucose uptake when the contractile response of the muscle was impaired. These data point to an indirect effect of Ca2+ on muscle glucose uptake.
Downstream Targets of Insulin- and Exercise-Stimulated Contraction (AS160, TBC1D1, and Rac1)
The downstream signaling pathways for insulin and exercise have revealed a converging signal for insulin- and the exercise-stimulated glucose uptake in skeletal muscle. Some of the molecules involved in this point of convergence are AS160 and TBC1D1. The link between AS160 and TBC1D1 as well as GLUT4 translocation involves Rab proteins. Rab proteins are members of the Ras small GTPase superfamily (81). These proteins are involved in many membrane trafficking events, and active Rabs recruit various effectors that are involved in vesicle budding, tethering, and fusion and therefore also in GLUT4 translocation (34, 63, 81). In addition to the well-established roles of Rab proteins, there is evidence that the Rho family GTPase Rac1 is involved in both insulin- and exercise-stimulated GLUT4 translocation (71, 72). Mice deficient in Rac1 (Rac1 KO mice) have decreased insulin-stimulated GLUT4 translocation (71, 83), and Rac1 inhibition decreased contraction-stimulated glucose uptake in mouse skeletal muscle (72).
AS160.
AS160 regulates insulin-stimulated GLUT4 translocation (5, 68, 74). It is phosphorylated on six different phospho-Akt-substrate (PAS) sites in response to both insulin and exercise in skeletal muscle (5, 43, 76). Studies have shown that AS160 PAS phosphorylation is increased after prolonged exercise in both humans (12, 70, 76) and rats (6, 14). Studies have shown that AMPK phosphorylates AS160 (PAS site) in response to AICAR and exercise in skeletal muscle (43) and that mutation of four PAS sites significantly inhibits both insulin and exercise-induced glucose uptake (44). Additional data have demonstrated that mutation of the calmodulin-binding domain on AS160 significantly inhibits exercise-stimulated glucose uptake but not insulin-stimulated glucose uptake (45). Interestingly, whole body KO of AS160 does not result in a significant increase in glucose transport in skeletal muscle (8). These data clearly show that both phosphorylation and calmodulin binding on AS160 are involved in the regulation of exercise-stimulated glucose uptake. These data also suggest that while AS160 may serve as a point of convergence for both insulin- and exercise-dependent signaling in the regulation of glucose uptake, other proteins may be involved in this regulation of glucose uptake.
TBC1D1.
Recent studies (5, 36, 37, 43, 64, 73, 77) have identified TBC1D1, the paralog of AS160, as another potential molecular link among signaling pathways converging on GLUT4 translocation in skeletal muscle. TBC1D1 and AS160 are 47% identical overall and have several comparable structural features. TBC1D1 was first identified in adipocytes in culture but is highly expressed in skeletal muscle (64). Insulin increases TBC1D1 PAS phosphorylation in skeletal muscle (2, 73, 80), but, unlike AS160, TBC1D1 can regulate insulin-stimulated glucose transport through a PAS-independent mechanism (2). Studies (2, 80) have shown that distinct mutations of TBC1D1 differentially regulate insulin- and exercise-stimulated glucose transport in skeletal muscle. These data suggest that TBC1D1 regulates both insulin- and exercise-stimulated glucose uptake through distinct phosphorylation sites. Taken together, these data suggest that TBC1D1, along with AS160, may also serve as a point of convergence for the regulation of GLUT4 translocation via both insulin- and exercise-stimulated glucose uptake in skeletal muscle.
Effects of Chronic Exercise Whole Body Metabolic Health
Exercise training has important therapeutic implications for people with type 2 diabetes. Here, we will discuss several adaptations of skeletal muscle to chronic exercise and how these adaptations can improve metabolic health in people with type 2 diabetes, specifically by improving mitochondrial function and increasing GLUT4 protein expression.
Effects of Chronic Exercise on Skeletal Muscle Mitochondria
People with type 2 diabetes have been reported to have smaller, damaged, or dysfunctional mitochondria (59) and decreased expression of peroxisome proliferator-activated receptor-γ coactivator-1α, a marker of mitochondrial biogenesis (47, 55). Skeletal muscle mitochondrial dysfunction has also been linked with insulin resistance and type 2 diabetes (59), although more studies are needed to directly connect mitochondrial deficits with impaired muscle glucose metabolism. It is also unclear if the mitochondrial defect precedes the development of type 2 diabetes or vice versa.
Endurance exercise training has been shown to increase mitochondrial content and activity (22, 52). A recent study (52) examined the effects of exercise training in people with type 2 diabetes and determined if exercise could restore mitochondrial content and function and insulin sensitivity in a patient population. Eighteen male subjects underwent 12 wk of exercise training. Subjects exercised 3 times/wk, 2 times/wk on a cycle ergometer (55% of their maximal O2 consumption), and 1 time/wk with resistance exercise (at 75% maximal voluntary contraction). After 12 wk of exercise, the defects previously observed in skeletal muscle mitochondrial activity in people with type 2 diabetes were completely negated. Interestingly, this restored mitochondrial function (Fig. 2) was correlated to improved insulin-stimulated glucose disposal (52). These data indicate that the restoration of mitochondrial function in type 2 diabetic patients accompanies the improved skeletal muscle insulin sensitivity.
Fig. 2.
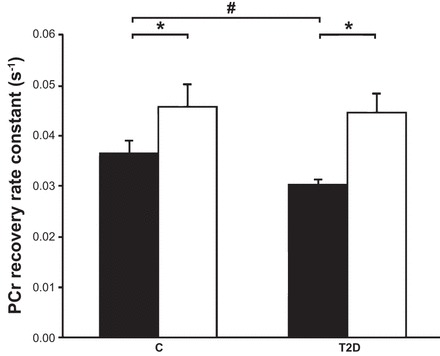
Exercise restores mitochondrial function in type 2 diabetic (T2D) subjects. In vivo mitochondrial function was measured in vastus lateralis muscle and expressed as the rate constant (in s−1) before (solid bars) and after training (open bars). A high rate constant reflects high in vivo mitochondrial function. Pre- and posttraining leg extension exercise was performed at 0.5 Hz to an acoustic cue on a magnetic resonance-compatible ergometer and a weight corresponding to 60% of the predetermined maximum. Postexercise phosphocreatine (PCr) resynthesis is driven almost purely oxidatively, and the resynthesis rate reflects in vivo mitochondrial function. Data are expressed as means ± SE. #Data for T2D subjects were significantly different from those of the control (C) group. *Posttraining was significantly different from pretraining. [Adapted from Ref. 52.]
Effects of Chronic Exercise on Skeletal Muscle GLUT4 Expression
It is well established that endurance exercise training increases skeletal muscle GLUT4 (22, 63). The increase in skeletal muscle GLUT4 causes an increase in skeletal muscle glucose uptake (24, 26, 47, 63). Most studies that have examined the therapeutic effects of exercise in people with type 2 diabetes have focused on low- to moderate-intensity aerobic exercise, which has been established as an effective strategy to improve the metabolic health of people with type 2 diabetes (9). The recommended amount of exercise for people with type 2 diabetes is 150 min of moderate intensity exercise per week (9). However, the amount of exercise required for improvements in whole body and skeletal muscle metabolic health is debatable. Recent studies have examined if the effects of low-volume high-intensity interval training (HIT), a time-efficient strategy to improve health and fitness, could be applied to people with type 2 diabetes as an intervention to improve glucose regulation and skeletal muscle metabolism (82). This study involved 14 people with type 2 diabetes (45). Eight of these subjects underwent HIT for 2 wk, where they trained on a cycle ergometer a total of six times over the 2-wk period. Each session consisted of a 5-min warmup, 10 × 60 s of cycling at 90% maximal effort with 60 s of recovery, and a 5-min cooldown. After 2 wk of HIT, people with type 2 diabetes had a marked increase in skeletal muscle GLUT4 expression and a significant decrease in blood glucose concentrations. Markers of mitochondrial activity, such as citrate synthase activity and expression of complex I, II, and III, were also significantly increased after 2 wk of HIT (Fig. 3). These data indicate that only 2 wk of HIT, where the weekly training time was 50% lower than the recommended guidelines for exercise in type 2 diabetic patients (9), increased glucose uptake and improved mitochondrial function in people with type 2 diabetes and improved whole body metabolism. This study indicated that HIT is a potential time-efficient exercise strategy for the treatment of type 2 diabetes.
Fig. 3.
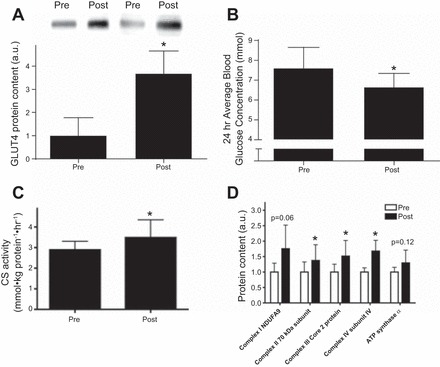
GLUT4 and mitochondrial markers increased after 2 wk of low-volume high-intensity interval training. A–D: 2 wk of high-intensity training increased GLUT4 protein content in skeletal muscle (A), glycemic control (average blood glucose) as measured by continuous glucose monitoring (B), citrate synthase (CS) activity (C), and mitochondrial markers in skeletal muscle (D). Pre, before training; Post, after training; NDUFA9, NADH dehydrogenase (ubiquinone) 1α subcomplex subunit 9. Values are means ± SD; n = 7. *P < 0.05. [Adapted from Ref. 46.]
Summary
In summary, exercise is critical in the treatment and prevention of type 2 diabetes. Acute exercise activates alternative molecular signals that can bypass defects in insulin signaling in skeletal muscle, resulting in an insulin-independent increase in glucose uptake. Exercise training results in increased skeletal muscle mitochondria and GLUT4 protein expression, which are associated with improved skeletal muscle insulin sensitivity and whole body metabolic health. Exercise-induced adaptations to skeletal muscle are essential to prevent and combat type 2 diabetes.
GRANTS
This work was supported by an American College of Sports Medicine Research Endowment Grant (to K. I. Stanford), Mary K. Iacocca Fellowship (to K. I. Stanford), and National Institutes of Health Grants R01-AR-42238 (to L. J. Goodyear), R01-DK-101043 (to L. J. Goodyear), and 5-P30-DK-36836 (Diabetes Research Center, Joslin Diabetes Center).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: K.I.S. drafted manuscript; K.I.S. and L.J.G. edited and revised manuscript; K.I.S. and L.J.G. approved final version of manuscript.
REFERENCES
- 1.Abbott MJ, Bogachus LD, Turcotte L. AMPKα2 deficiency uncovers time-dependency in the regulation of contraction-induced palmitate and glucose uptake in mouse muscle. J Appl Physiol 111: 125–134, 2011. [DOI] [PubMed] [Google Scholar]
- 2.An D, Toyoda T, Taylor EB, Yu H, Fujii N, Hirshman MF, Goodyear LJ. TBC1D1 regulates insulin- and contraction-induced glucose transport in mouse skeletal muscle. Diabetes 59: 1358–1365, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bain J, Plater L, Elliott M, Shpiro N, Hastie CJ, McLauchlan H, Klevernic I, Arthur JS, Alessi DR, Cohen P. The selectivity of protein kinase inhibitors: a further update. Biochem J 408: 297–315, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Bonadonna RC, Del Prato S, Saccomani MP, Bonora E, Gulli G, Ferrannini E, Bier D, Cobelli C, DeFronzo RA. Transmembrane glucose transport in skeletal muscle of patients with non-insulindependent diabetes. J Clin Invest 92: 486–494, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bruss MD, Arias EB, Lienhard GE, Cartee GD. Increased phosphorylation of Akt substrate of 160 kDa (AS160) in rat skeletal muscle in response to insulin or contractile activity. Diabetes 54: 41–50, 2005. [DOI] [PubMed] [Google Scholar]
- 6.Capaldo B, Gastaldelli A, Antoniello S, Auletta M, Pardo F, Ciociaro D, Guida R, Ferrannini E, Sacca L. Splanchnic and leg substrate exchange after ingestion of a natural mixed meal in humans. Diabetes 48: 958–966, 1999. [DOI] [PubMed] [Google Scholar]
- 7.Cartee GD, Funai K. Exercise and insulin: convergence or divergence at AS160 and TBC1D1? Exerc Sport Sci Rev 37: 188–195, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7a.Centers for Disease Control and Prevention. Prevalence of overweight and obesity among adults with diagnosed diabetes–United States, 1988–1994 and 1999–2000. Morbid Mortal Week Rep 53: 1066–1068, 2004. [PubMed] [Google Scholar]
- 8.Chavez JA, Roach WG, Keller SR, Lane WS, Lienhard GE. Inhibition of GLUT4 translocation by Tbc1d1, a Rab GTPase-activating protein abundant in skeletal muscle, is partially relieved by AMP-activated protein kinase activation. J Biol Chem 283: 9187–9195, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Colberg SR, Sigal RJ, Fernahll B, Regensteiner JG, Blissmer BJ, Rubin RR, Chasan-Taber L, Albright AL, Braun B. Exercise and type 2 diabetes: the American College of Sports Medicine and the American Diabetes Association: joint position statement executive summary. Diabetes Care 33: 2692–2696, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Daugaard JR, Richter EA. Muscle- and fibre type-specific expression of glucose transporter 4, glycogen synthase and glycogen phosphorylase proteins in human skeletal muscle. Pflügers Arch 447: 452–456, 2004. [DOI] [PubMed] [Google Scholar]
- 11.DeFronzo RA, Tripathy D. Skeletal muscle insulin resistance is the primary defect in type 2 diabetes. Diabetes Care 32: S157–S163, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Deshmukh A, Coffey VG, Zhong Z, Chibalin AV, Hawley JA, Zierath JR. Exercise-induced phosphorylation of the novel Akt substrates AS160 and filamin A in human skeletal muscle. Diabetes 55: 1776–1782, 2006. [DOI] [PubMed] [Google Scholar]
- 13.Espeland MA, Glick HA, Bertoni A, Brancati FL, Bray GA, Clark JM, Curtis JM, Egan C, Evans M, Foreyt JP, Ghazarian S, Gregg EW, Hazuda HP, Hill JO, Hire D, Horton ES, Hubbard VS, Jakicic JM, Jeffery RW, Johnson KC, Kahn SE, Killean T, Kitabchi AE, Knowler WC, Kriska A, Lewis CE, Miller M, Montez MG, Murillo A, Nathan DM, Nyenwe E, Patricio J, Peters AL, Pi-Sunyer X, Pownall H, Redmon JB, Rushing J, Ryan DH, Safford M, Tsai AG, Wadden TA, Wing RR, Yanovski SZ, Zhang P; Look AHEAD. Research Group. Impact of an intensive lifestyle intervention on use and cost of medical services among overweight and obese adults with type 2 diabetes: the action for health in diabetes. Diabetes Care 37: 2548–56, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Folli F, Saad MJA, Backer JM, Kahn CR, Saad MJ. Insulin stimulation of phosphatidylinositol 3-kinase activity and association with insulin receptor substrate 1 in liver and muscle of the intact rat. J Biol Chem 267: 22171–22177, 1992. [PubMed] [Google Scholar]
- 15.Fujii N, Hirshman MF, Kane EM, Ho RC, Peter LE, Seifert MM, Goodyear LJ. AMP-activated protein kinase α2 activity is not essential for contraction- and hyperosmolarity-induced glucose transport in skeletal muscle. J Biol Chem 280: 39033–39041, 2005. [DOI] [PubMed] [Google Scholar]
- 16.Funai K, Cartee GD. Inhibition of contraction-stimulated AMP-activated protein kinase inhibits contraction-stimulated increases in PAS-TBC1D1 and glucose transport without altering PAS-AS160 in rat skeletal muscle. Diabetes 58: 1096–1104, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Goodyear LJ, Giorgino F, Balon TW, Condorelli G, Smith RJ. Effects of contractile activity on tyrosine phosphoproteins and phosphatidylinositol 3-kinase activity in rat skeletal muscle. Am J Physiol Endocrinol Metab 268: E987–E995, 1995. [DOI] [PubMed] [Google Scholar]
- 18.Goodyear LJ, Kahn BB. Exercise, glucose transport, and insulin sensitivity. Annu Rev Med 49: 235–261, 1998. [DOI] [PubMed] [Google Scholar]
- 19.Gregg EW, Chen H, Wagenknecht LE, Clark JM, Delahanty LM, Bantle J, Pownall HJ, Johnson KC, Safford MM, Kitabchi AE, Pi-Sunyer FX, Wing RR, Bertoni AG; Look AHEAD. Research Group. Association of an intensive lifestyle intervention with remission of type 2 diabetes. JAMA 308: 2489–96, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hardie DG, Carling D, Carlson M. The AMP-activated/SNF1 protein kinase subfamily: metabolic sensors of the eukaryotic cell? Annu Rev Biochem 67: 821–855, 1998. [DOI] [PubMed] [Google Scholar]
- 21.Harris MI, Flegal KM, Cowie CC, Eberhardt MS, Goldstein DE, Little RR, Wiedmeyer HM, Byrd-Holt DD. Prevalence of diabetes, impaired fasting glucose, and impaired glucose tolerance in U.S. adults. The Third National Health and Nutrition Examination Survey, 1988–1994. Diabetes Care 21: 518–524, 1998. [DOI] [PubMed] [Google Scholar]
- 22.Higashida K, Kim SH, Higuchi M, Holloszy JO, Han DH. Normal adaptations to exercise despite protection against oxidative stress. Am J Physiol Endocrinol Metab 301: E779–E784, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hirshman MF, Koh HJ, Goodyear LJ. LKB1 in muscle is critical for exercise capacity and partially regulates glucose transport. Diabetes 55, Suppl 1: A13, 2006. [Google Scholar]
- 24.Howlett KF, Andrikopoulos S, Proietto J, Hargreaves M. Exercise-induced muscle glucose uptake in mice with graded, muscle-specific GLUT4 deletion. Physiol Rep 1: e00065, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hu FB, Manson JE, Stampfer MJ, Colditz G, Liu S, Solomon CG, Willett WC. Diet, lifestyle, and the risk of type 2 diabetes mellitus in women. N Engl J Med 345: 790–797, 2001. [DOI] [PubMed] [Google Scholar]
- 26.Hussey SE, McGee SL, Garnham A, McConell GK, Hargreaves M. Exercise increases skeletal muscle GLUT4 gene expression in patients with type 2 diabetes. Diabetes Obes Metab 14: 768–771, 2012. [DOI] [PubMed] [Google Scholar]
- 27.Jebailey L, Rudich A, Huang X, DiCiano-Oliveira C, Kapus A, Klip A. Skeletal muscle cells and adipocytes differ in their reliance on TC10 and Rac for insulin-induced actin remodeling. Mol Endocrinol 18: 359–372, 2004. [DOI] [PubMed] [Google Scholar]
- 28.Jensen TE, Rose AJ, Jorgensen SB, Brandt N, Schjerling P, Wojtaszewski JF, Richter EA. Possible CaMKK-dependent regulation of AMPK phosphorylation and glucose uptake at the onset of mild tetanic skeletal muscle contraction. Am J Physiol Endocrinol Metab 292: E1308–E1317, 2007. [DOI] [PubMed] [Google Scholar]
- 29.Jensen TE, Sylow L, Rose AJ, Madsen AB, Angin YM, Stine J, Richter EA. Contraction-stimulated glucose transport in muscle is controlled by AMPK and mechanical stress but not sarcoplasmic reticulum Ca2+ release. Molecular Metabolism. In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jensen TE, Schjerling P, Viollet B, Wojtaszewski JF, Richter EA. AMPK α1 activation is required for stimulation of glucose uptake by twitch contraction, but not by H2O2, in mouse skeletal muscle. PLOS ONE 3: e2102, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jeppesen J, Maarbjerg SJ, Jordy AB, Fritzen AM, Pehmoller C, Sylow L, Serup AK, Jessen N, Thorsen K, Prats C, Qvortrup K, Dyck DJR, Hunter RW, Sakamoto K, Thomson D, Schjerling P, Wojtaszewski JF, Richter EA, Kiens B. LKB-1 regulates lipid oxidation during exercise independently of AMPK. Diabetes 62: 1490–1499, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jonker JT, de Mol P, de Vries ST, Widya RL, Hammer S, van Schinkel LD, van der Meer RW, Gans RO, Webb AG, Kan HE, de Koning EJ, Bilo HJ, Lamb HJ. Exercise and type 2 diabetes mellitus: changes in tissue-specific fat distribution and cardiac function. Radiology 269: 434–442, 2013. [DOI] [PubMed] [Google Scholar]
- 33.Jorgensen SB, Viollet B, Andreelli F, Frosig C, Birk JB, Schjerling P, Vaulont S, Richter EA, Wojtaszewski JF. Knockout of the α2 but not α1 5′-AMP-activated protein kinase isoform abolishes 5-aminoimidazole-4-carboxamide-1-β-4-ribofuranosidebut not contraction-induced glucose uptake in skeletal muscle. J Biol Chem 279: 1070–1079, 2004. [DOI] [PubMed] [Google Scholar]
- 34.Kaddai V, Le Marchand-Brustel Y, Cormont M. Rab proteins in endocytosis and Glut4 trafficking. Acta Physiol 192: 75–88, 2008. [DOI] [PubMed] [Google Scholar]
- 35.Kahn CR. Banting Lecture. Insulin action, diabetogenes, and the cause of type II diabetes. Diabetes 43: 1066–1084, 1994. [DOI] [PubMed] [Google Scholar]
- 36.Kane S, Sano H, Liu SC, Asara JM, Lane WS, Garner CC, Lienhard GE. A method to identify serine kinase substrates. Akt phosphorylates a novel adipocyte protein with a Rab GTPase-activating protein (GAP) domain. J Biol Chem 277: 22115–22118, 2002. [DOI] [PubMed] [Google Scholar]
- 37.Karlsson HK, Zierath JR, Kane S, Krook A, Lienhard GE, Wallberg-Henriksson H. Insulin-stimulated phosphorylation of the Akt substrate AS160 is impaired in skeletal muscle of type 2 diabetic subjects. Diabetes 54: 1692–1697, 2005. [DOI] [PubMed] [Google Scholar]
- 38.Kelley DE, He J, Menshikova EV, Ritov VB. Dysfunction of mitochondria in human skeletal muscle in type 2 diabetes. Diabetes 51: 2944–2950, 2002. [DOI] [PubMed] [Google Scholar]
- 39.Kemp BE, Mitchelhill KI, Stapleton D, Michell BJ, Chen ZP, Witters LA. Dealing with energy demand: the AMP-activated protein kinase. Trends Biochem Sci 24: 22–25, 1999. [DOI] [PubMed] [Google Scholar]
- 40.Kennedy JW, Hirshman MF, Gervino EV, Ocel JV, Forse RA, Hoenig SJ, Aronson D, Goodyear LJ, Horton ES. Acute exercise induces GLUT4 translocation in skeletal muscle of normal human subjects and subjects with type 2 diabetes. Diabetes 48: 1192–1197, 1999. [DOI] [PubMed] [Google Scholar]
- 41.Koh HJ, Arnolds DE, Fujii N, Tran TT, Rogers MJ, Jessen N, Li Y, Liew CW, Ho RC, Hirshman MF, Kulkarni RN, Kahn CR, Goodyear LJ. Skeletal muscle-selective knockout of LKB1 increases insulin sensitivity, improves glucose homeostasis, and decreases TRB3. Mol Cell Biol 26: 8217–8227, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Koh HJ, Toyoda T, Fujii N, Jung MM, Rathod A, Middelbeek RJ, Lessard SJ, Treebak JT, Tsuchihara K, Esumi H, Richter EA, Wojtaszewski JF, Hirshman MF, Goodyear LJ. Sucrose nonfermenting AMPK-related kinase (SNARK) mediates contraction-stimulated glucose transport in mouse skeletal muscle. Proc Natl Acad Sci USA 107: 15541–15546, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kramer HF, Witczak CA, Fujii N, Jessen N, Taylor EB, Arnolds DE, Sakamoto K, Hirshman MF, Goodyear LJ. Distinct signals regulate AS160 phosphorylation in response to insulin, AICAR, and contraction in mouse skeletal muscle. Diabetes 55: 2067–2076, 2006. [DOI] [PubMed] [Google Scholar]
- 44.Kramer HF, Witczak CA, Taylor EB, Fujii N, Hirshman MF, Goodyear LJ. AS160 regulates insulin- and contraction-stimulated glucose uptake in mouse skeletal muscle. J Biol Chem 281: 31478–31485, 2006. [DOI] [PubMed] [Google Scholar]
- 45.Kramer HF, Taylor EB, Witczak CA, Fujii N, Hirshman MF, Goodyear LJ. The calmodulin-binding domain of AS160 regulates contraction- but not insulin-stimluated glucose uptake in skeletal muscle. Diabetes 56: 2854–2058, 2006. [DOI] [PubMed] [Google Scholar]
- 46.Little JP, Gillen JB, Percival ME, Safdar A, Tarnopolsky MA, Punthakee Z, Jung ME, Gibala MJ. Low-volume high-intensity interval training reduces hyperglycemia and increases skeletal muscle mitochondrial capacity in patients with type 2 diabetes. J Appl Physiol 111: 1554–1560, 2011. [DOI] [PubMed] [Google Scholar]
- 47.Look AHEAD Research Group. Effect of a long-term behavioral weight loss intervention on nephropathy in overweight or obese adults with type 2 diabetes: a secondary analysis of the Look AHEAD randomized clinical trial. Lancet Diabetes Endocrinol 2: 801–809, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Look AHEAD Research Group. Cardiovascular effects of intensive lifestyle intervention in type 2 diabetes. N Engl J Med 369: 145–154, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lowell BB, Shulman GI. Mitochondrial dysfunction and type 2 diabetes. Science 307: 384–387, 2005. [DOI] [PubMed] [Google Scholar]
- 50.Maarbjerg SJ, Jørgensen SB, Rose AJ, Jeppesen J, Jensen TE, Treebak JT, Birk JB, Schjerling P, Wojtaszewski JF, Richter EA. Genetic impairment of AMPKα2 signaling does not reduce muscle glucose uptake during treadmill exercise in mice. Am J Physiol Endocrinol Metab 297: E924–E934, 2009. [DOI] [PubMed] [Google Scholar]
- 51.Martin IK, Katz A, Wahren J. Splanchnic and muscle metabolism during exercise in NIDDM patients. Am J Physiol Endocrinol Metab 269: E583–E590, 1995. [DOI] [PubMed] [Google Scholar]
- 52.Meex RC, Schrauwen-Hinderling VB, Moonen-Kornips E, Schaart G, Mensink M, Phielix E, van de Weijer T, Sels JP, Schrauwen P, Hesselink MK. Restoration of muscle mitochondrial function and metabolic flexibility in type 2 diabetes by exercise training is paralleled by increased myocellular fat storage and improved insulin sensitivity. Diabetes 59: 572–579, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Merrill GF, Kurth EJ, Hardie DG, Winder WW. AICA riboside increases AMP-activated protein kinase, fatty acid oxidation, and glucose uptake in rat muscle. Am J Physiol Endocrinol Metab 273: E1107–E1112, 1997. [DOI] [PubMed] [Google Scholar]
- 54.Mootha VK, Lindgren CM, Eriksson KF, Subramanian A, Sihaq S, Lehar J, Puigserver P, Carlsson E, Ridderstrale M, Laurila E, Houstis N, Daly MJ, Patterson N, Mesirov JP, Golub TR, Tamayo P, Spiegelman B, Lander ES, Hirschhorn JN, Altshuler D, Groop LC. PCG-1α-responsive genes involved in oxidative phosphorylation are coordinately down regulated in human diabetes. Nat Genet 34: 267–273, 2003. [DOI] [PubMed] [Google Scholar]
- 55.Mu J, Brozinick JT, Jr, Valladares O, Bucan M, Birnbaum MJ. A role for AMP-activated protein kinase in contraction- and hypoxia-regulated glucose transport in skeletal muscle. Mol Cell 7: 1085–1094, 2001. [DOI] [PubMed] [Google Scholar]
- 56.National Task Force on the Prevention and Treatment of Obesity. Overweight, obesity, and health risk. National Task Force on the Prevention and Treatment of Obesity. Arch Intern Med 160: 898–904, 2000. [DOI] [PubMed] [Google Scholar]
- 57.O'Neill HM, Maarbjerg SJ, Crane JD, Jeppesen J, Jorgensen SB, Schertzer JD, Shyroka O, Kiens B, Van Denderen BJ, Tarnopolsky MA, Kemp BE, Richter EA, Steinberg GR. AMP-activated protein kinase (AMPK) β1β2 muscle null mice reveal an essential role for AMPK in maintaining mitochondrial content and glucose uptake during exercise. Proc Natl Acad Sci USA 108: 16092–16097, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Pan XR, Li GW, Hu YH, Wang JX, Yang WY, An ZX, Hu ZX, Lin J, Xiao JZ, Cao HB, Liu PA, Jiang XG, Jiang YY, Wang JP, Zheng H, Zhang H, Bennet PH, Howard PV. Effects of diet and exercise in preventing NIDDM in people with impaired glucose tolerance. The Da Qing IGT and Diabetes Study. Diabetes Care 20: 537–544, 1997. [DOI] [PubMed] [Google Scholar]
- 59.Patti ME, Butte AJ, Crunkhorn S, Cusi K, Berria R, Kashyap S, Miyazaki Y, Kohane I, Costello M, Saccone R, Landaker EJ, Goldfine AB, Mun E, DeFronzo R, Finlayson J, Kahn CR, Mandarino LJ. Coordinated reduction of genes of oxidative metabolism in humans with insulin resistance and diabetes: potential role of PGC1 and NRF1. Proc Natl Acad Sci USA 100: 8466–8471, 2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Pette D, Staron RS. Transitions of muscle fiber phenotypic profiles. Histochem Cell Biol 115: 359–372, 2001. [DOI] [PubMed] [Google Scholar]
- 62.Rejeski WJ, Bray GA, Chen SH, Clark JM, Evans M, Hill JO, Jakicic JM, Johnson KC, Neiberg R, Ip EH; Look AHEAD. Research Group. Aging and physical function in type 2 diabetes: 8 years of an intensive lifestyle intervention. J Gerontol A Biol Sci Med Sci; 10.1093/gerona/glu083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Richter EA, Hargreaves M. Exercise, GLUT4, and skeletal muscle glucose uptake. Physiol Rev 93: 993–1017, 2013. [DOI] [PubMed] [Google Scholar]
- 64.Roach WG, Chavez JA, Mîinea CP, Lienhard GE: Substrate specificity and effect on GLUT4 translocation of the Rab GTPase-activating protein Tbc1d1. Biochem J 403: 353–358, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Robertson RP. Antagonist: diabetes and insulin resistance-philosophy, science, and the multiplier hypothesis. J Lab Clin Med 125: 560–564, 1995. [PubMed] [Google Scholar]
- 66.Rockl KS, Witczak CA, Goodyear LJ. Signaling mechanisms in skeletal muscle: acute responses and chronic adaptations to exercise. IUBMB Life 60: 145–53, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Sakamoto K, McCarthy A, Smith D, Green KA, Grahame HD, Ashworth A, Alessi DR. Deficiency of LKB1 in skeletal muscle prevents AMPK activation and glucose uptake during contraction. EMBO J 24: 1810–1820, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Sano H, Kane S, Sano E, Miinea CP, Asara JM, Lane WS, Garner CW, Lienhard GE. Insulin-stimulated phosphorylation of a Rab GTPase-activating protein regulates GLUT4 translocation. J Biol Chem 278: 14599–14602, 2003. [DOI] [PubMed] [Google Scholar]
- 69.Simard-Lefort N, St-Amand E, Morasse S, Cote CH, Marette A. The α subunit of AMPK is essential for submaximal contraction-mediated glucose transport in skeletal muscle in vitro. Am J Physiol Endocrinol Metab 295: E1447–E1454, 2008. [DOI] [PubMed] [Google Scholar]
- 70.Sriwijitkamol A, Coletta DK, Wajcberg E, Balbontin GB, Reyna SM, Barrientes J, Eagan PA, Jenkinson CP, Cersosimo E, DeFronzo RA, Sakamoto K, Musi N. Effect of acute exercise on AMPK signaling in skeletal muscle of subjects with type 2 diabetes: a time-course and dose-response study. Diabetes 56: 836–848, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Sylow L, Jensen TE, Kleinert M, Hojlund K, Kiens B, Wojtaszewski J, Prats C, Schjerling P, Richter EA. Rac1 signaling is required for insulin-stimulated glucose uptake and is dysregulated in insulin-resistant murine and human skeletal muscle. Diabetes 62: 1865–1875, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sylow L, Jensen TE, Kleinert M, Mouatt JR, Maarbjerg SJ, Jeppesen J, Prats C, Chiu TT, Boguslavsky S, Klip A, Schjerling P, Richter EA. Rac1 is a novel regulator of contraction-stimulated glucose uptake in skeletal muscle. Diabetes 62: 1139–1151, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Taylor EB, An D, Kramer HF, Yu H, Fujii NL, Roeckl KS, Bowles N, Hirshman MF, Xie J, Feener EP, Goodyear LJ. Discovery of TBC1D1 as an insulin-, AICAR-, and contraction-stimulated signaling nexus in mouse skeletal muscle. J Biol Chem 283: 9787–9796, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Thong FS, Dugani CB, Klip A. Turning signals on and off: GLUT4 traffic in the insulin-signaling highway. Physiology (Bethesda) 20: 271–284, 2005. [DOI] [PubMed] [Google Scholar]
- 75.Treadway JL, James DE, Burcel E, Ruderman NB. Effect of exercise on insulin receptor binding and kinase activity in skeletal muscle. Am J Physiol Endocrinol Metab 256: E138–E144, 1989. [DOI] [PubMed] [Google Scholar]
- 76.Treebak JT, Birk JB, Rose AJ, Kiens B, Richter EA, Wojtaszewski JF. AS160 phosphorylation is associated with activation of α2β2γ1- but not α2β2γ3-AMPK trimeric complex in skeletal muscle during exercise in humans. Am J Physiol Endocrinol Metab 292: E715–E722, 2007. [DOI] [PubMed] [Google Scholar]
- 77.Treebak JT, Glund S, Deshmukh A, Klein DK, Long YC, Jensen TE, Jorgensen SB, Viollet B, Andersson L, Neumann D, Wallimann T, Richter EA, Chibalin AV, Zierath JR, Wojtaszewski JF. AMPK-mediated AS160 phosphorylation in skeletal muscle is dependent on AMPK catalytic and regulatory subunits. Diabetes 55: 2051–2058, 2006. [DOI] [PubMed] [Google Scholar]
- 78.Tuomilehto j Lindstrom J, Eriksson JG, Valle TT, Hamalainen H, Ilanne-Parikka P, Keinanen-Kiukaanniemi S, Laakso M, Louheranta A, Rastas M, Salminen V, Uusitupa M. Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. N Engl J Med 344: 1343–1350, 2001. [DOI] [PubMed] [Google Scholar]
- 79.Ueda S, Kitazaw S, Ishida K, Nishikawa Y, Matsui M, Matsumoto H, Aoki T, Nozaki S, Takeda T, Tamori Y, Aiba A, Kahn CR, Kataoka T, Satoh T. Crucial role of the small GTPase Rac1 in insulin-stimulated translocation of glucose transporter 4 to the mouse skeletal muscle sarcolemma. FASEB J 24: 2254–2261, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Vichaiwong K, Purohit S, An D, Toyoda T, Jessen N, Hirshman MF, Goodyear LJ. Contraction regulates site-specific phosphorylation of TBC1D1 in skeletal muscle. Biochem J 431: 311–320, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Wennerberg K, Rossman KL, Der CJ. The Ras superfamily at a glance. J Cell Sci 118: 843–846, 2005. [DOI] [PubMed] [Google Scholar]
- 82.Wing RR, Lang W, Wadden TA, Safford M, Knowler WC, Bertoni AG, Hill JO, Brancati FL, Peters A, Wagenknecht L; Look AHEAD Research Group. Benefits of modest weight loss in improving cardiovascular risk factors in overweight and obese individuals with type 2 diabetes. Diabetes Care 34: 1481–1486, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Witczak CA, Fujii N, Hirshman MF, Goodyear LJ. Ca2+/calmodulin-dependent protein kinase kinase-alpha regulates skeletal muscle glucose uptake independent of AMP-activated protein kinase and Akt activation. Diabetes 56: 1403–1409, 2007. [DOI] [PubMed] [Google Scholar]
- 84.Witczak CA, Jessen N, Warro DM, Toyoda T, Fujii N, Anderson ME, Hirshman MF, Goodyear LJ. CaMKII regulates contraction- but not insulin-induced glucose uptake in mouse skeletal muscle. Am J Physiol Endocrinol Metab 298: E1150–E1160, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Wojtaszewski JF, Higaki Y, Hirshman MF, Michael MD, Dufresne SD, Kahn CR, Goodyear LJ. Exercise modulates postreceptor insulin signaling and glucose transport in muscle-specific insulin receptor knockout mice. J Clin Invest 104: 1257–1264, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Wright DC, Hucker KA, Holloszy JO, Han DH. Ca2+ and AMPK both mediate stimulation of glucose transport by muscle contractions. Diabetes 53: 330–335, 2004. [DOI] [PubMed] [Google Scholar]
- 87.Zierath JR, Krook A, Wallberg-Henriksson H. Insulin action in skeletal muscle from patients with NIDDM. Mol Cell Biochem 182: 153–160, 1998. [PubMed] [Google Scholar]