

Abstract
Different functional roles for the hands have been demonstrated, however leg control is not as well understood. The purpose of the present study was to evaluate bilateral knee neuromuscular control to determine if the limb receiving greater attention would have more well-tuned control compared to an unattended limb. Surface electrodes were placed on seven muscles of each limb, before standing on two force platforms. Visual feedback was given of the forces and moments of the “focus limb,” but not the “unattended limb.” Static isometric forces were matched with their focus limb, requiring their unattended limb to push in the opposite direction, using a combination of forward-backward-medial-lateral shear forces while muscle activity was collected bilaterally. There was a significant main effect for limb task (p = 0.02), with the medial hamstrings being more specific (p = 0.001) while performing the unattended limb and the lateral hamstring being more well-tuned (p = 0.007) while performing the focus limb task. The focus limb's medial and lateral gastrocnemius were principally active in the forwards direction, but only the unattended limb's lateral gastrocnemius was active in the backwards direction. Findings suggest unique neuromuscular control strategies are used for the legs depending on limb task.
Keywords: Neuromechanics, Specificity, Synergy, Bidirectional, Intermuscular coordination
1. Introduction
Human interlimb, or between leg, coordination is critical for upright stable function during activities of daily living (e.g. walking). While functional limb symmetry has been assumed, experimental evidence is equivocal (Arsenault et al., 1986; Carpes et al., 2010; Gundersen et al., 1989; Reisman et al., 2005). Interlimb coordination is complex and the difference in findings may be related to specialization of lower extremities, or the complexity of the task (Reisman et al., 2005) used within the study.
Specialization of lower extremity function has been examined widely (Sadeghi et al., 2000). The legs may have functional specialization based upon the task, suggested by asymmetrical injury rates (Brophy et al., 2010; Ruedl et al., 2012), joint stability (Rein et al., 2010), and strength (Ostenberg et al., 1998). Specialization of limb task may be important to consider during rehabilitating an injured limb. For instance, if the preferred kicking leg of a soccer player was injured while performing the support limb task, as is the most common mechanism for male soccer players (Brophy et al., 2010), then perhaps the rehabilitation of this leg would benefit maximally for return to sport by retraining it in support limb tasks rather than tasks similar to kicking. However, interlimb inter-muscular coordination is not well understood with regard to task (Sadeghi et al., 2000).
Activities of daily living often require the lower extremities to coordinate bidirectional forces while both feet are on the ground. During the initiation of stepping, for instance, the two feet are required to produce forces in opposite directions on the floor by activating muscles of the legs in different patterns to support the swing and stance limb tasks (Elble et al., 1994). Further, interlimb coordination is required to produce bidirectional forces in other activities such as hitting a baseball (Welch et al., 1995), opening a door, or lifting an object and twisting at the trunk. Studying inter-limb coordination during a task that requires static and isometric voluntary neuromuscular coordination of bidirectional forces, allows for experimental control of limb task and simplifies the movement as joint angle and load magnitude may be constrained (Buchanan and Lloyd, 1995).
Lower extremity interlimb coordination has not been widely studied. Coordination of muscle activity has been studied using a specificity index in an unloaded seated position (Krishnan et al., 2008; Williams et al., 2003, 2004, 2005a,b) and while standing (Flaxman et al., 2012; Smith et al., 2012), but neither examined the effects of interlimb coordination. Further, interlimb coordination has been examined with regards to symmetry during locomotion (Carpes et al., 2010; Gundersen et al., 1989; Hunt et al., 2004; Sadeghi et al., 2000) and a reaction to external perturbations (Dietz et al., 1989; Torres-Oviedo et al., 2006). However, there is still a gap in the literature regarding standing interlimb muscle coordination with regard to limb task. Findings have the potential to refine our understanding of muscle recruitment that would be applicable clinically.
Therefore, the purpose of this study was to evaluate neuromuscular control differences based upon leg task. Specifically, we evaluated individual muscle activation patterns for legs receiving the focus of attention (“focus limb”) as well as the limb left unattended (“unattended limb”) across a variety of static isometric force postures. Comparisons were made between the two limbs while under nearly identical external forces. We hypothesized that the muscles of the focus limb would be more well-tuned to a principal direction of force in comparison to the unattended limb.
2. Methods
2.1. Subjects
Twenty healthy subjects (11 males and 9 females) were recruited with an average age of 23.2 ± 2.8 years, based upon a priori power analyses from piloting. Inclusion criteria were being a regular participant (>50 h/year) of high-level sport requiring running, jumping, and cutting activities. Subjects were screened by a physical therapist for impairments that would be expected to affect the result so for this study the exclusion criteria were; having a history of any significant lower extremity injury (i.e. cruciate ligament tear); any lower limb ligament sprain, muscle or tendon sprain, or fracture within 6 months of participation; any central or peripheral nerve impairment; or the presence of knee joint effusion. Subjects gave written informed consent and the study was approved by the institutional human subjects review board.
2.2. Electromyography
Standardized electrode sites (Hermens et al., 2000) were cleaned of hair and dirt for the following muscles: rectus femoris (RF), vastus medialis (VM) and lateralis (VL), medial (MH) and lateral hamstrings (LH), and medial (MG) and lateral gastrocnemius (LG). Disk shaped adhesive Ag/AgCl surface electrodes (Neurotrode 120, Myotronics-Neuromed, Inc., Tukwila, WA) with a 2 cm inter-electrode distance were applied and then connected to a differential preamplifier with a gain of 20 and a two-pole (10–2000 Hz) bandpass filter. Each EMG preamplifier consisted of two standard snap sensor contacts, and a reference contact (connected to a monotrode). Input impedance was >100,000,000 ohms, common mode rejection ratio was >100 dB at 65 Hz, signal-to-noise ratio was <1.2 uV RMS. Electrode location was confirmed and optimized using multiple isometric contractions. Surface electrodes and preamplifiers were further secured with tape (Hypafix™, Smith and Nephew, London, UK) and then wrapped with 10 cm wide elastic bands (Superwrap™, Fabrifoam, Inc., Exton, PA, USA).
The analog signal was anti-aliased at 500 Hz through a backpack unit (MA-300-28, Motion Laboratory Systems, Baton Rouge, Louisiana, USA), sampled at 1000 Hz, and these raw data were stored in the computer for digital post-processing. All data were collected through a 16-bit analog-to-digital board (Measurement Computing, Norton, MA; PCI-DAS4020/16).
A resting trial was recorded prior to the experimental protocol while the subject stood on the force platforms, so that the baseline resting activity could be removed during EMG data reduction. Resting EMG trials were performed in a standing, rather than seated or supine, position so that we could later remove the postural activity and any background noise of the muscle in post-processing and thus characterize the muscle activity required while coordinating external force postures.
The EMG of each muscle were full-wave rectified and normalized to the peak EMG magnitudes recorded prior to testing during maximal voluntary isometric contractions (MVICs) at standardized joint positions. EMG data were not filtered during the experiment. The static isometric nature of the task required subjects to hold relatively still, so that motion artifact that normally requires filtering was not present.
MVICs were performed separately for each leg in a seated position, with the exception of the gastrocnemii, in standardized joint positions: RF at 20° hip flexion, the vasti and hamstring muscles at 90° of hip and knee flexion, and the gastrocnemii in single-leg stance in maximal plantar flexion. All subjects regularly performed exercises of these types, and were encouraged vocally to produce maximal contractions.
2.3. Standing target-matching experimental protocol
Subjects stood with bare feet approximately hip-width apart on two force-platforms, one for each foot (Fig. 1; OR-6, AMTI, Water-town, MA, USA). Outlines of the subject's feet were made using a pencil on the force-platforms so that if they wished to move for any reason they were able to easily place their feet in the original location. While performing the task, subjects were required to keep both feet firmly in contact with the force-platforms and distribute their mass equally between their feet.
Fig. 1.

The experimental set-up for the standing target matching.
One limb was randomly selected to perform the target-matching task first, to prevent a test-order affect, and this limb was operationally defined as the “focus limb” (making the other leg the “unattended limb”). Visual feedback was given of the forces and moments created by the focus limb on the force-platform via a circular cursor (Fig. 2) projected on a screen in front of them (Fig. 1).
Fig. 2.
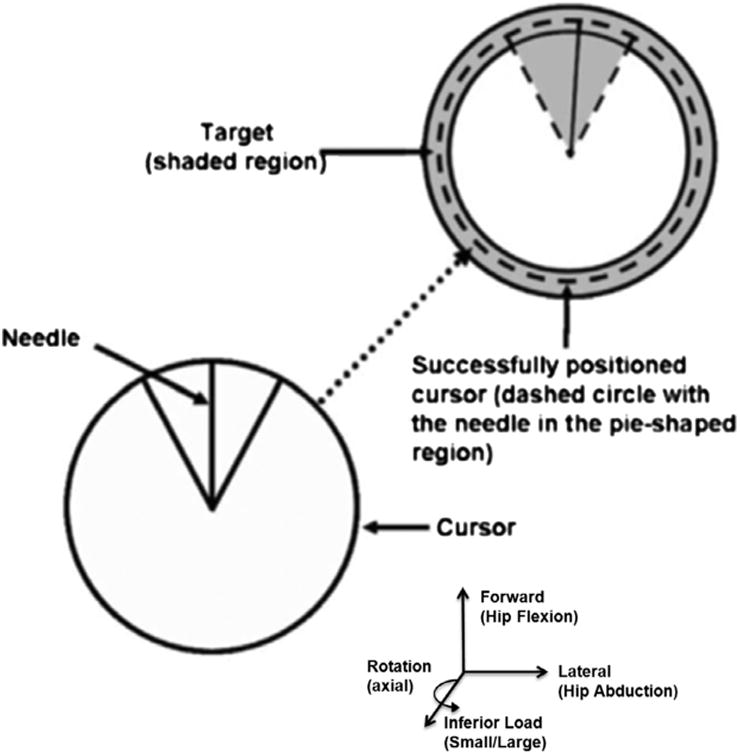
Depiction of the cursor and target used in the target-matching protocol.
They did not receive any visual feedback of the forces and moments produced by their unattended limb. Subjects used their focus limb to position the cursor within the randomly produced target locations. Subsequently, the limb tasks were reversed so that both legs were tested while performing the focus and unattended limb tasks.
Subjects were instructed to position the cursor using their focus limb over a narrow target that consists of two concentric circles (Fig. 2). The cursor projected on the screen moved in response to focus limb forces and moments with four degrees of freedom: (1) anterior–posterior foot shear force (i.e., hip extension/flexion) moved the cursor up and down, (2) medial/lateral foot shear forces (i.e., varus/hip adduction and valgus/hip abduction) moved the cursor right and left, (3) vertical (i.e., inferior/superior) loads made the cursor circumference smaller or larger, and (4) rotational foot shear moments (i.e., internal/external rotation) of the shank moved the “needle” within the cursor clockwise or counter-clockwise, respectively. Targets appeared one at a time and in random order at one of 18 positions (located every 20° around the circumference of a circle in the anterior–posterior–varus–valgus plane; Fig. 3). Each target location was matched four times for a total of 72 trials, and these four repetitions were averaged for analysis.
Fig. 3.
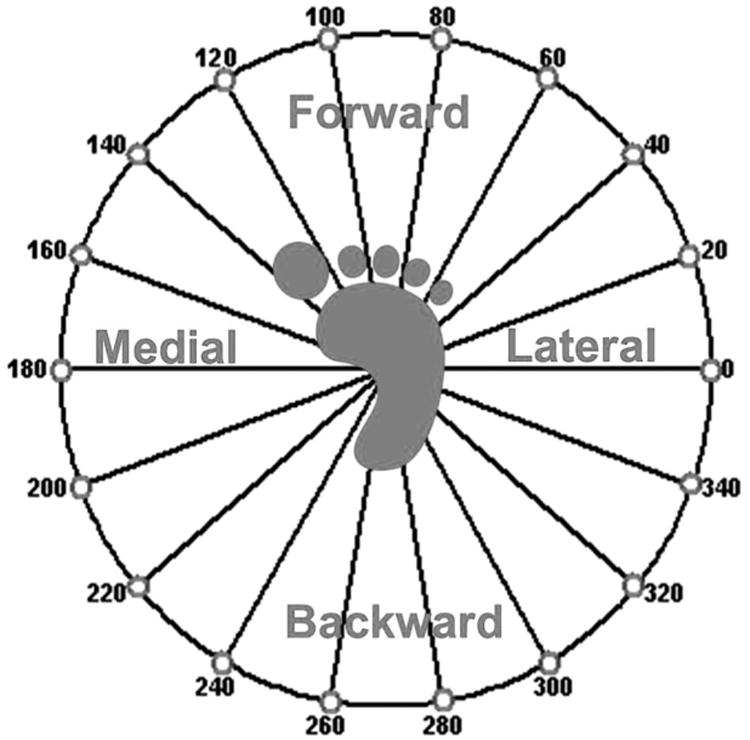
Polar plot of the static isometric force posture locations from the perspective of the focus limb.
To standardize the forces and moments required to match targets across subjects, subjects performed MVICs in the four degrees of freedom used to manipulate the cursor, to determine the peak forces and moments. A shear force of 30% of the peak magnitude of the anterior–posterior (whichever was lower) directed forces recorded during the standing MVICs was required to successfully position the cursor within the target. This load was chosen out of safety considerations, as it created sagittal plane joint moments that were found to be similar to slow walking (1.0 m/s) during initial piloting. Body mass, or the vertical force, was required to be balanced between the two feet. Subjects were required to hold the cursor within the target area for 500 ms before the EMG data were recorded and the subject was allowed to move onto the next target location.
Prior to collecting data for each limb subjects performed all 18 targets in a random order to familiarize themselves with the task and to minimize the effects of task novelty and learning. There was no time limit for the target-matching task and subjects were encouraged to rest whenever they felt the need to prevent fatigue.
The shear forces on the focus limb foot were equal and opposite to those on the foot of the unattended limb, so the muscle activity patterns could be compared between the two limbs. For instance, if the right foot (as in Fig. 3) were pushing to the 0° target, then the left foot must push towards the 180° target with equal and opposite shear force. By transposing the left onto the coordinate system of the right, the two limbs may be compared while under the same external shear forces (in the example case while under a medially directed external force). After performing all 18-target locations the muscle activations patterns were plotted and compared between the focus and unattended limb tasks while in the coordinate system of the right foot. All comparisons were made between the two limb tasks while the limbs were under the same external forces. For instance, the forward direction of the focus and unattended limbs depicts when the subject pushed in the forwards direction during both tasks.
2.4. Data reduction
Circular statistics were used to analyze muscle specificity and principal direction of action in the anterior–posterior–medial–lateral foot shear force plane in a manner that has been described previously (Dewald et al., 1995; Williams et al., 2003, 2004, 2005a), as a method of describing how specific the muscle activation pattern of a single muscle was to a single shear force direction, or in this case a target. The principal direction of muscle activation was determined as an EMG vector (EMGi), which has the magnitude as the percent MVIC and orientation representing the shear force direction (direction of the target). The specificity index was calculated as the sum of the vectors across all 18 targets divided by the scalar magnitude. The specificity index ranged between 0.0 and 1.0, where 1.0 indicated that a muscle was only active while performing one target and totally “specific” only to that direction(muscle activity depicted by a single radius), whereas a specificity index of 0.0 would indicate that the muscle was equally active at all targets and thus, not “specific” to any force posture (muscle activity depicted by a circle).
2.5. Data analysis
Descriptive statistics were performed for each dependent variable. Limb task (focus versus unattended) was the independent variable. The specificity index, magnitude of EMG activity as a percentage of MVIC, and the principal direction of action of the seven muscles were the dependent variables. The left and right limbs performed both the focus and unattended limb tasks, and the mean of the left and right limbs for the two tasks was used for analysis.
Multivariate analysis of variance (MANOVA) was used to evaluate potential differences between limb tasks for each of the dependent variables. The significance level for all statistical procedures was set at α < 0.05. Post hoc testing was performed, when appropriate, with univariate tests using Bonferroni adjustments for multiple comparisons.
3. Results
3.1. Limb task
There was a significant main effect for limb task with regards to specificity index (p < 0.001). Mean specificity index values ranged from 0.16 to 0.50 across the seven muscles tested for the two tasks, with the two vasti muscles being the least specific muscles and the two hamstring muscles being the most specific (Fig. 4). Post hoc analyses revealed that the LH had a significantly (p = 0.001) greater specificity index value while performing the focus limb task (0.477 ± 0.13) than the unattended limb task (0.337 ± 0.16). Further, the MH had a significantly (p = 0.007) greater specificity index value while performing the unattended limb task (0.497 ± 0.12) than as a focus limb (0.420 ± 0.14). The LG of the unattended limb (0.271 ± 0.156) had on average a greater specificity index value than the focus limb (0.204 ± 0.126; p = 0.081).
Fig. 4.
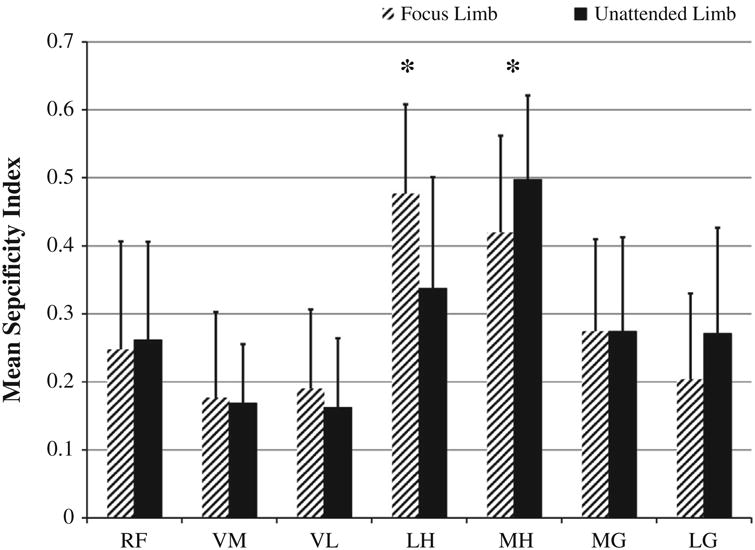
Specificity of muscle action results for the muscles of the focus and unattended limbs. Asterisks denote significant differences at the p < 0.05 level. Abbreviations: RF, rectus femoris; VM, vastus medialis; VL, vastus lateralis; LH, lateral hamstrings; MH, medial hamstrings; MG, medial gastrocnemius; LG, lateral gastrocnemius.
3.2. Muscle recruitment
There was also a significant main effect for limb task with regards to muscle recruitment as a percentage of MVIC (p = 0.020). Muscle activation percentage ranged from 7% to 14% of MVIC (Fig. 5). On average, the LH was recruited with a greater percentage of MVIC as the focus limb (13.40 ± 8.87%) than as the unattended limb (10.4 ± 9.58%; p = 0.150). Also, on average the MH was recruited with a greater percentage of MVIC as the unattended limb (14.26 ± 11.64%) than as a focus limb (11.64 ± 8.53%; p = 0.254). Further, on average the LG was recruited with a greater percentage of MVIC while performing the unattended limb task (11.1 ± 7.30%) in comparison to the focus limb (8.12 ± 6.50%; p = 0.057).
Fig. 5.
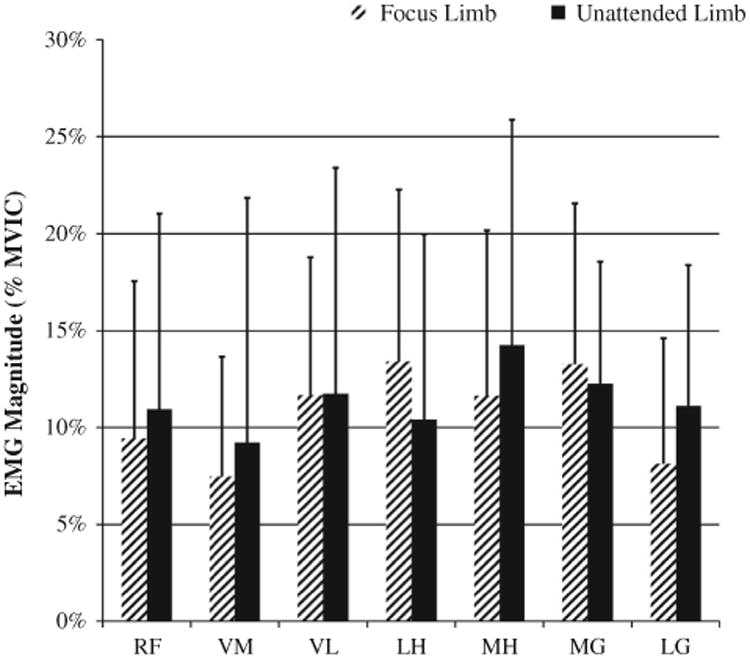
Relative percentage of muscle activation results for the muscles of the focus and unattended limbs as a percentage of MVIC. Abbreviations: RF, rectus femoris; VM, vastus medialis; VL, vastus lateralis; LH, lateral hamstrings; MH, medial hamstrings; MG, medial gastrocnemius; LG, lateral gastrocnemius.
3.3. Patterns of muscle activation
Different patterns of muscle activation were found between limb tasks and across muscles when plotted in polar coordinates (Fig. 6). Both the direction of muscle activation as well as the magnitude of muscle activation varied across muscles and tasks. VM was less active than VL across target locations, and the vasti were activated in both the forwards and backwards direction, co-contracting with both the RF and hamstring muscles while performing both the focus and unattended limb tasks. RF had a similar magnitude of activation between the two limb tasks, and was primarily active in the anterior direction for both limb tasks. The two hamstring muscles are represented by patterns of activation that are primarily in the posterior direction. MG and LG had different patterns of activation, both in magnitude and in pattern, with MG being active at a higher magnitude of MVIC and in the forwards direction for both the focus and unattended limb tasks; LG was active in the forwards direction for the focus limb task and in the backwards direction for the unattended limb task.
Fig. 6.
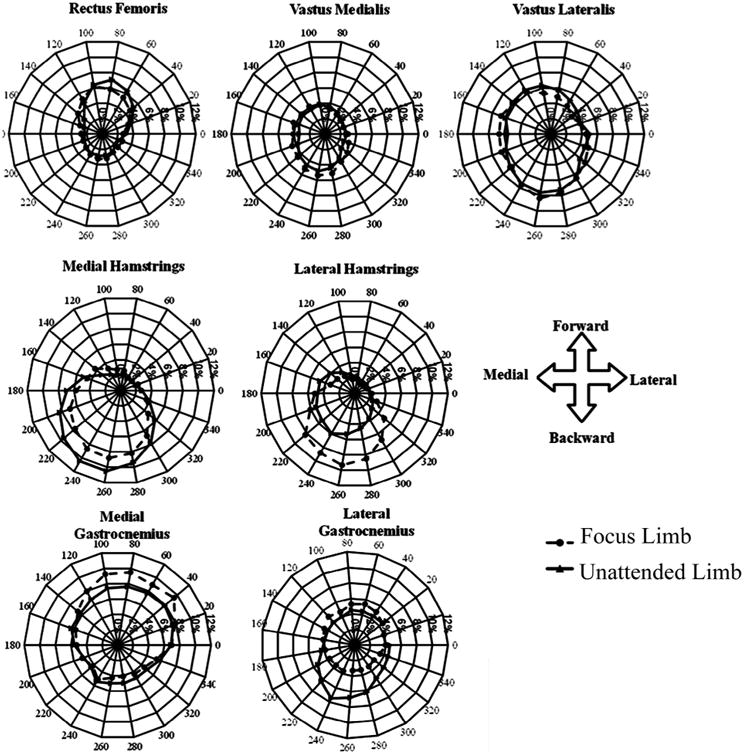
Polar plots of the subject's focus and unattended limb knee muscle activity patterns. Muscle activity of the left limb was transposed, and polar plots are represented in the coordinate system of the right limb.
3.4. Principal direction of muscle action
The principal direction of muscle action was different across muscles and limb tasks (Fig. 7). During the focus limb task the principal direction of muscle action for both the LG and MG was in the forward direction. However, during the unattended limb task the LG was principally directed in the backwards direction, while the MG remained principally active in the forward direction. RF was principally active in the forwards direction, while the vasti and hamstring muscles were principally active in the backwards direction.
Fig. 7.
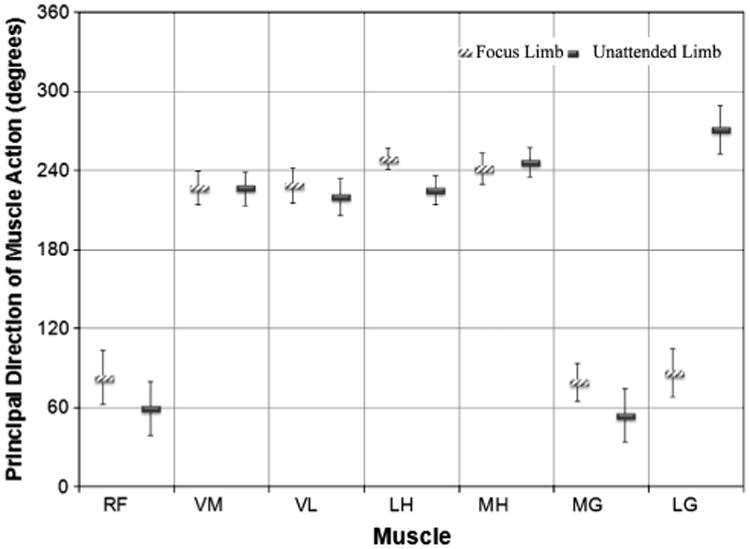
Bar graph of the principal direction of action of the knee muscles while performing the mobilizer and stabilizer tasks. Whiskers represent the standard deviation of the mean. Abbreviations: RF, rectus femoris; VM, vastus medialis; VL, vastus lateralis; LH, lateral hamstrings; MH, medial hamstrings; MG, medial gastrocnemius; LG, lateral gastrocnemius.
4. Discussion
The aim of this study was to examine the effect that leg task has on neuromuscular coordination of leg muscles in a standing and weight bearing posture. Our results indicate that neuromuscular coordination differences exist between the focus and unattended limbs. In part, findings support our initial hypotheses that muscle activation patterns of the focus limb would be more well-tuned to a principal direction of action. Findings suggest that hamstring muscles are tuned differently for either the focus or unattended limb tasks, even though the external forces are nearly identical. The implications of our findings suggest that specialization of muscle function should be considered when the performance of limb task is being evaluated. For instance, a physical therapist may facilitate the lateral hamstrings with manual tapping or electrical stimulation to enhance that muscle's activation while the client's attention is on that limb.
One of the most interesting findings of this study was that the hamstring muscles and the LG had functions that were specialized for the focus and unattended limb tasks. Perhaps the focus and unattended limbs require different knee joint stiffness. The MG and LH are external rotators of the tibia on the femur, and the LG and MH are internal rotators. The tibiofemoral joint becomes rigid with external rotation, due to the “screw home mechanism.” We found that the LH was activated with a higher percentage of MVIC while performing the focus limb task, in comparison to the unattended limb task, perhaps allowing greatest control of force at the foot. Further, the LG and the MH were activated with a higher percentage of their MVIC, more specifically, and in the backwards direction while performing the unattended limb task, in comparison to the focus limb task. LG and MH muscles internally rotate the tibia on the femur (Besier et al., 2003; Buford et al., 2001; Wretenberg et al., 1996), thus unscrewing the tibiofemoral joint (Levens et al., 1948; Pappas et al., 2013), so perhaps these muscles were coordinated differently for the unattended limb task for a less rigid limb segment.
In this group, we did not find significant differences between limb tasks with regards to the relative muscle activation as a percentage of MVIC. This may in part be explained by our inclusion of both male and females subjects, for others have found males contract the muscles about the knee with a greater percentage of MVIC than females during a seated target-matching task (Krishnan et al., 2008). However, on average the hamstring muscles were recruited with a greater percentage of MVIC while performing the focus limb task, and the lateral gastrocnemius was recruited with a greater percentage of MVIC while performing the unattended limb task. With this study we have laid the groundwork for future examination of the effect of sex on relative muscle activation in this standing posture. While the sample size was based upon a priori comparisons, perhaps with a larger sample there would be significant differences observed in a pairwise comparison by task.
Interestingly, the vasti (VL and VM) were found to have synergic patterns of contraction with both the RF and the hamstrings. We believe that the uniarticulate vasti were acting to brace the knee, while the biarticulate rectus femoris, medial hamstrings, and lateral hamstrings transferred work from the hip down to the force-platforms. The biarticulate hamstring muscles were supported by their antagonistic uniarticulate vasti muscles while the foot pushed in the backwards direction on the force-platform. A similar pattern of activation has been described in cycling, where the hamstring and vasti muscles coactivate during the downstroke (van Ingen Schenau et al., 1992). There is a similar pattern of coactivation between the hamstrings and vasti during the push-off phase of sprinting (Jacobs and van Ingen Schenau, 1992b). During a seated single leg multi-joint task where individuals were tasked with directional force production on a force-platform the biarticulate hamstrings and rectus femoris were found to be responsible to force direction control, while the uniarticulate vasti were active during force postures that required both knee flexion and extension (Jacobs and van Ingen Schenau, 1992a). In the current study, activation of the vasti was more pronounced in the backwards direction likely because the task was weight bearing, bipedal, and our subjects chose to position their knees with less flexion than during the previously mentioned study where subjects were seated.
The experimental protocol used in this study was specifically designed to evaluate the voluntary muscle control of the two limbs under nearly identical standing force postures. One potential limitation of this study is that we currently do not know the degree to which the muscle control strategies observed in this study relate to control strategies used in functional activities like running and jumping. Future studies should build upon this one by evaluating static muscle specificity and dynamic motion analysis that includes electromyography.
In summary, neuromuscular control differences exist in the muscle activity patterns of the subject's focus and unattended legs, especially in the hamstrings and lateral gastrocnemius. The vasti were found to act synergistically with both the rectus femoris and the hamstrings, indicating that they are recruited to brace the knee to allow energy to be transmitted from the hip to the force-plate. The medial hamstrings and lateral gastrocnemius displayed more specific muscle activity patterns while performing the unattended limb task, which may represent a strategy for knee stabilization control. The lateral hamstrings pattern of muscle activation was more specific while performing the focus limb task, which may represent a preferential use of the muscle typically used for power generation during gait. Muscle activity differences between the focus and unattended legs indicate that the CNS uses different neuromuscular control for the two limb tasks.
Acknowledgments
We wish to thank the subjects for their time and effort while completing this experiment. This work was supported by the National Institutes of Health AR 46386.
Biographies

Toran D. MacLeod is a post-doctoral in the Musculo-skeletal Quantitative Imaging Research Group at UC San Francisco. Dr. MacLeod has a PhD in Biomechanics & Movement Sciences from the University of Delaware and is a licensed Physical Therapist. He research is on gait analysis and running mechanics, ACL rehabilitation research, neuromuscular control and magnetic resonance imaging.

Kurt Manal is a research assistant professor in the Department of Mechanical Engineering at the University of Delaware with appointments in Biomedical Engineering and the Biomechanics and Movement Science Program. Dr. Manal received his Ph.D. in Biomechanics & Movement Science from the University of Delaware in 1998. His current research interests include the prediction of muscle forces and joint loading in patients with ACL injury and in people with knee osteoarthritis.

Karin Grävare Silbernagel is an Assistant Professor of Physical Therapy at the University of the Sciences in Philadelphia. She has a PhD in Medicine from the University of Gothenburg and has been a clinician, researcher, and educator in the U.S. and Sweden. She has been a practicing physical therapist for 20 years, mainly in orthopaedics and sports medicine. Her research areas are on tendon injuries especially the clinical aspects of prevention, treatment, and evaluation.

Lynn Snyder-Mackler is the Alumni Distinguished Professor of Physical Therapy at the University of Delaware. She has a BA from Johns Hopkins University, an Sc.D. in Applied Anatomy and Physiology from Boston University, and Physical Therapy certification from The University of Pennsylvania. She is a world expert in sports medicine, especially ACL injuries. She currently holds a prestigious NIH MERIT Award for her work on ACL reconstruction.

Thomas S. Buchanan is the George W. Laird Professor of Mechanical Engineering & Biomedical Engineering at the University of Delaware and Director of the Delaware Rehabilitation Institute. He received a BS from UCSD in Bioengineering and a PhD in Theoretical & Applied Mechanics from Northwestern University. He has been President of the American Society of Biomechanics and editor-in-chief of the Journal of Applied Biomechanics. His current grants include “Center of Biomedical Research Excellence on Osteoarthritis Prevention & Treatment” and “ACL Injured Knee: MRI and Biomechanical Modeling.”
Footnotes
Conflict of interest: None declared.
References
- Arsenault AB, Winter DA, Marteniuk RG. Bilateralism of EMG profiles in human locomotion. Am J Phys Med. 1986;65(1):1–16. [PubMed] [Google Scholar]
- Besier TF, Lloyd DG, Ackland TR. Muscle activation strategies at the knee during running and cutting maneuvers. Med Sci Sports Exer. 2003;35(1):119–27. doi: 10.1097/00005768-200301000-00019. [DOI] [PubMed] [Google Scholar]
- Brophy R, Silvers HJ, Gonzales T, Mandelbaum BR. Gender influences: the role of leg dominance in ACL injury among soccer players. Brit J Sports Med. 2010;44(10):694–7. doi: 10.1136/bjsm.2008.051243. [DOI] [PubMed] [Google Scholar]
- Buchanan TS, Lloyd DG. Muscle activity is different for humans performing static tasks which require force control and position control. Neurosci Lett. 1995;194(1–2):61–4. doi: 10.1016/0304-3940(95)11727-e. [DOI] [PubMed] [Google Scholar]
- Buford WL, Jr, Ivey FM, Jr, Nakamura T, Patterson RM, Nguyen DK. Internal/external rotation moment arms of muscles at the knee: moment arms for the normal knee and the ACL-deficient knee. The knee. 2001;8(4):293–303. doi: 10.1016/s0968-0160(01)00106-5. [DOI] [PubMed] [Google Scholar]
- Carpes FP, Mota CB, Faria IE. On the bilateral asymmetry during running and cycling- a review considering leg preference. Phys Ther Sport: Official J Assoc Chart Physiother Sports Med. 2010;11(4):136–42. doi: 10.1016/j.ptsp.2010.06.005. [DOI] [PubMed] [Google Scholar]
- Dewald JP, Pope PS, Given JD, Buchanan TS, Rymer WZ. Abnormal muscle coactivation patterns during isometric torque generation at the elbow and shoulder in hemiparetic subjects. Brain. 1995;118(Pt 2):495–510. doi: 10.1093/brain/118.2.495. [DOI] [PubMed] [Google Scholar]
- Dietz V, Horstmann GA, Berger W. Interlimb coordination of leg-muscle activation during perturbation of stance in humans. J Neurophysiol. 1989;62(3):680–93. doi: 10.1152/jn.1989.62.3.680. [DOI] [PubMed] [Google Scholar]
- Elble RJ, Moody C, Leffler K, Sinha R. The initiation of normal walking. Mov Disord. 1994;9(2):139–46. doi: 10.1002/mds.870090203. [DOI] [PubMed] [Google Scholar]
- Flaxman TE, Speirs AD, Benoit DL. Joint stabilisers or moment actuators: the role of knee joint muscles while weight-bearing. J Biomech. 2012;45(15):2570–6. doi: 10.1016/j.jbiomech.2012.07.026. [DOI] [PubMed] [Google Scholar]
- Gundersen LA, Valle DR, Barr AE, Danoff JV, Stanhope SJ, Snyder-Mackler L. Bilateral analysis of the knee and ankle during gait: an examination of the relationship between lateral dominance and symmetry. Phys Ther. 1989;69(8):640–50. doi: 10.1093/ptj/69.8.640. [DOI] [PubMed] [Google Scholar]
- Hermens HJ, Freriks B, Disselhorst-Klug C, Rau G. Development of recommendations for SEMG sensors and sensor placement procedures. J Electromyogr Kinesiol. 2000;10(5):361–74. doi: 10.1016/s1050-6411(00)00027-4. [DOI] [PubMed] [Google Scholar]
- Hunt MA, Sanderson DJ, Moffet H, Inglis JT. Interlimb asymmetry in persons with and without an anterior cruciate ligament deficiency during stationary cycling. Arch Phys Med Rehabil. 2004;85(9):1475–8. doi: 10.1016/j.apmr.2003.10.017. [DOI] [PubMed] [Google Scholar]
- Jacobs R, van Ingen Schenau GJ. Control of an external force in leg extensions in humans. J Physiol. 1992a;457:611–26. doi: 10.1113/jphysiol.1992.sp019397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobs R, van Ingen Schenau GJ. Intermuscular coordination in a sprint push-off. J Biomech. 1992b;25(9):953–65. doi: 10.1016/0021-9290(92)90031-u. [DOI] [PubMed] [Google Scholar]
- Krishnan C, Huston K, Amendola A, Williams GN. Quadriceps and hamstrings muscle control in athletic males and females. J Orthop Res. 2008;26(6):800–8. doi: 10.1002/jor.20592. [DOI] [PubMed] [Google Scholar]
- Levens AS, Inman VT, Blosser JA. Transverse rotation of the segments of the lower extremity in locomotion. J Bone Joint Surg Am. 1948;30A(4):859–72. [PubMed] [Google Scholar]
- Ostenberg A, Roos E, Ekdahl C, Roos H. Isokinetic knee extensor strength and functional performance in healthy female soccer players. Scand J Med Sci Sports. 1998;8(5 Pt 1):257–64. doi: 10.1111/j.1600-0838.1998.tb00480.x. [DOI] [PubMed] [Google Scholar]
- Pappas E, Zampeli F, Xergia SA, Georgoulis AD. Lessons learned from the last 20 years of ACL-related in vivo-biomechanics research of the knee joint. Knee Surg, Sports Traumatol, Arthrosc: Official J ESSKA. 2013;21(4):755–66. doi: 10.1007/s00167-012-1955-0. [DOI] [PubMed] [Google Scholar]
- Rein S, Fabian T, Zwipp H, Mittag-Bonsch M, Weindel S. Influence of age, body mass index and leg dominance on functional ankle stability. Foot Ankle Int/Am Orthopaed Foot Ankle Soc Swiss Foot Ankle Soc. 2010;31(5):423–32. doi: 10.3113/FAI.2010.0423. [DOI] [PubMed] [Google Scholar]
- Reisman DS, Block HJ, Bastian AJ. Interlimb coordination during locomotion: what can be adapted and stored? J Neurophysiol. 2005;94(4):2403–15. doi: 10.1152/jn.00089.2005. [DOI] [PubMed] [Google Scholar]
- Ruedl G, Webhofer M, Helle K, Strobl M, Schranz A, Fink C, et al. Leg dominance is a risk factor for noncontact anterior cruciate ligament injuries in female recreational skiers. Am J Sports Med. 2012;40(6):1269–73. doi: 10.1177/0363546512439027. [DOI] [PubMed] [Google Scholar]
- Sadeghi H, Allard P, Prince F, Labelle H. Symmetry and limb dominance in able-bodied gait: a review. Gait Posture. 2000;12(1):34–45. doi: 10.1016/s0966-6362(00)00070-9. [DOI] [PubMed] [Google Scholar]
- Smith AJ, Flaxman TE, Speirs AD, Benoit DL. Reliability of knee joint muscle activity during weight bearing force control. J Electromyogr Kinesiol: Official J Int Soc Electrophysiol Kinesiol. 2012;22(6):914–22. doi: 10.1016/j.jelekin.2012.03.010. [DOI] [PubMed] [Google Scholar]
- Torres-Oviedo G, Macpherson JM, Ting LH. Muscle synergy organization is robust across a variety of postural perturbations. J Neurophysiol. 2006;96(3):1530–46. doi: 10.1152/jn.00810.2005. [DOI] [PubMed] [Google Scholar]
- van Ingen Schenau GJ, Boots PJ, de Groot G, Snackers RJ, van Woensel WW. The constrained control of force and position in multi-joint movements. Neuroscience. 1992;46(1):197–207. doi: 10.1016/0306-4522(92)90019-x. [DOI] [PubMed] [Google Scholar]
- Welch CM, Banks SA, Cook FF, Draovitch P. Hitting a baseball: a biomechanical description. J Orthopaed Sports Phys Ther. 1995;22(5):193–201. doi: 10.2519/jospt.1995.22.5.193. [DOI] [PubMed] [Google Scholar]
- Williams GN, Barrance PJ, Snyder-Mackler L, Axe MJ, Buchanan TS. Specificity of muscle action after anterior cruciate ligament injury. J Orthop Res. 2003;21(6):1131–7. doi: 10.1016/S0736-0266(03)00106-2. [DOI] [PubMed] [Google Scholar]
- Williams GN, Barrance PJ, Snyder-Mackler L, Buchanan TS. Altered quadriceps control in people with anterior cruciate ligament deficiency. Med Sci Sports Exer. 2004;36(7):1089–97. doi: 10.1249/01.mss.0000131959.20666.11. [DOI] [PubMed] [Google Scholar]
- Williams GN, Snyder-Mackler L, Barrance PJ, Axe MJ, Buchanan TS. Neuromuscular function after anterior cruciate ligament reconstruction with autologous semitendinosus-gracilis graft. J Electromyogr Kinesiol. 2005a;15(2):170–80. doi: 10.1016/j.jelekin.2004.08.005. [DOI] [PubMed] [Google Scholar]
- Williams GN, Snyder-Mackler L, Barrance PJ, Buchanan TS. Quadriceps femoris muscle morphology and function after ACL injury: a differential response in copers versus non-copers. J Biomech. 2005b;38(4):685–93. doi: 10.1016/j.jbiomech.2004.04.004. [DOI] [PubMed] [Google Scholar]
- Wretenberg P, Nemeth G, Lamontagne M, Lundin B. Passive knee muscle moment arms measured in vivo with MRI. Clin Biomech. 1996;11(8):439–46. doi: 10.1016/s0268-0033(96)00030-7. [DOI] [PubMed] [Google Scholar]