

Abstract
Since its discovery as a lysosomal hydrolase, Cathepsin D (CatD) has been the subject of intensive scrutiny by numerous scientists. Those accumulated efforts have defined its biosynthetic pathway, structure, and companion proteins in the context of its perceived “house keeping” function. However, in the past two decades CatD has emerged as a multifunctional enzyme, involved in myriad biological processes beyond its original “housekeeping” role. CatD is responsible for selective and limited cleavage (quite distinct from non-specific protein degradation) of particular substrates vital to proper cellular function. These proteolytic events are critical in the control of biological processes, including cell cycle progression, differentiation and migration, morphogenesis and tissue remodeling, immunological processes, ovulation, fertilization, neuronal outgrowth, angiogenesis, and apoptosis. Consistent with the biological relevance of CatD, its deficiency, altered regulation or post-translational modification underlie important pathological conditions such as cancer, atherosclerosis, neurological and skin disorders. Specifically, deregulated synthesis, post-translational modifications and hyper-secretion of CatD, along with its mitogenic effects, are established hallmarks of cancer. More importantly, but less studied, is its significance in regulating the sensitivity to anticancer drugs.
This review outlines CatD’s post-translational modifications, cellular trafficking, secretion and protein binding partners in normal mammary gland, and restates the “site-specific” function of CatD which is most probably dictated by its post-translational modifications and binding partners. Noteworthy, CatD’s association with one of its binding partners in the context of drug sensitivity is highlighted, with the optimism that it could contribute to the development of more effective chemotherapeutic agent(s) tailored for individual patients.
Keywords: Cathepsin D, Binding partners, Post-translational modification, Cancer, Mammary gland
Historical Overview
The term “Cathepsin”, was first introduced in 1929, and described the proteolytic activity of acidified tissue extracts towards hemoglobin [1]. Subsequently, this proteolytic activity proved to contain multiple forms of Cathepsin, termed A, B and C [2]. Cathepsin(s) were purified from spleen in 1940, and their lysosomal association was established in 1955 [3,4]. By late 1959, the fourth member of the family was identified as “Cathepsin D” (CatD) [5]. Ensuing studies characterized its glycoprotein nature, conversion to two chains mature enzyme, and revealed its homology with other acid proteases [6].
The discovery of CatD’s elevated levels in muscular dystrophy and arthritis underscored its pathological significance [7,8]. It’s possible involvement in breast cancer was first noted in rat model of breast carcinoma induced by 3-methylcholanthrene (or 7,12-dimethylbenz[a] anthracene). In this model, tumors undergo repeated growth and regression following successive pregnancies [9]. Changes in Cathepsin (with no reference to Cathepsin subtypes) were noted in regressing mammary tumors [10], however, the significance of these findings was overlooked, and it was not until the 1980s that Cat D’s association with human breast cancer was established [11,12].
In 1995, CatD’s involvement in apoptotic cell death emphasized its functional significance in embryonic development [13], and the generation of CatD knockout mice further established its vital role in proper organ development after birth [14].
CatD Gene Transcription and Regulation
The 5′ upstream region of CatD promoter contains several GC boxes and a TATAA sequence [15]. This mixed promoter directs two types of transcription: TATA-independent transcription starting at several sites upstream from the TATA box (directed by GC boxes and Sp1 factor), and TATA-dependent transcription initiating about 28 bp downstream from the TATA box (Figure 1A).
Figure 1.
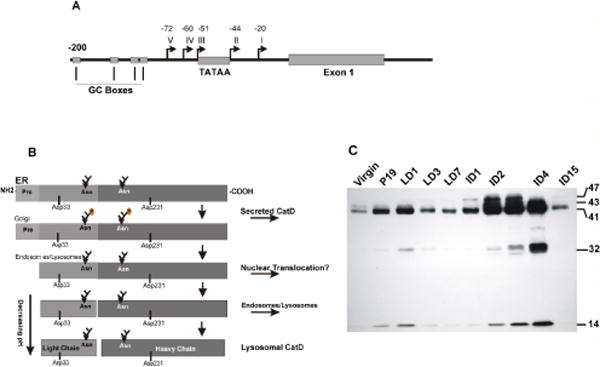
(A) Schematic presentation of CatD promoter region. The TATA and GC sequences are represented by square boxes, five transcription start sites are indicated by arrows and their distance from the +1 nucleotide are indicated. (B) Pictorial presentation of proteolytic processing of pre-pro-CatD. CatD is synthesized in the ER as a pre-pro enzyme containing a signal peptide at its amino terminus. As the enzyme traverses the ER, it loses its signal peptide and is glycosylated at two N-glycosylation sites. The pro-enzyme is transported to the Golgi, tagged with Man-6-P for binding to Man-6-PR. The complex is transported across the Golgi and reaches the endosomal compartment. The acidic environment causes the release of the receptor and the pro-peptide is cleaved generating the single chain active enzyme. Further removal of seven amino acids generates the light chain and the heavy chain mature enzyme (please see the text). (C) Developmental regulation of CatD level and proteolytic processing in mouse mammary tissue. Cytosolic extracts prepared from the mammary gland at different stages of development were subjected to SDS-PAGE and Western blot analysis for the presence of CatD cleavage products. V: virgin, P: pregnant, LD: lactation days 1, 3 and 7, IND: involution days 1–15. Molecular mass of CatD proteolytic products are indicated on the right.
The former confers house keeping gene properties to CatD, while the latter is a feature of regulated genes and could be induced under specific physiological conditions (i.e. during development and tissue remodeling). Transcription from different start sites leads to mRNA variants of different sizes which might affect stability, initiation of translation and/or subcellular localization of the protein product [15]. In this context, TATA-dependent transcription of CatD is greatly induced by estrogen and heavily exploited in breast cancer [15,16], the response to estrogen is tissue (or cell) specific, as endometrial derived Ishiwaka cancer cell line is non-responsive to estrogen [17].
Adding to the complexity, growth factors (insulin, insulin-like growth factor I, TGF-α, basic fibroblast growth factor and epidermal growth factor) induce CatD mRNA [17,18]. However, these effects are mostly mediated via GC-rich sites and imperfect estrogen response element in CatD promoter [17]. EGF rapidly induces CatD mRNA by 2- to 4-fold, and its effect is dependent upon de novo protein synthesis [19]. The mixed promoter, tissue-specific responses of CatD mRNA to hormones, the direct and indirect responses to growth factors are all indicative of a very complex regulatory mechanism(s) governing CatD’s gene transcription [19].
CatD Synthesis, Intercellular Localization and Secretion
The early era of CatD research provided fundamental insight into its synthesis, posttranslational modifications, and transport route to its lysosomal destination. CatD is synthesized as a single chain pre-pro-enzyme (412 amino acids), is glycosylated at two N-linked glycosylation sites (Asn residues 134 and 263 (pre-pro-CatD numbering, UniProtKB/Swiss-Prot)], and is transported to the Golgi. The N-glycan structures acquire mannose-6phosphate (Man-6P) residues for binding to Man-6P receptor(s) (Man-6PR)[20]. The complex is transported to the lysosomal compartment, the acidic environment dissociates the complex, the receptor recycles back, and pro-CatD is processed to the 48 kDa active single chain, and finally to the mature two chain (34 and 14 kDa, respectively) enzyme [21–23].
The more contemporary CatD research has uncovered functions beyond and above its original housekeeping role. In this context, CatD also functions in the cytosol and the extracellular environment. The cytosolic traverse of CatD occurs via limited release from the lysosome and is a key signaling event initiating the apoptotic cascade [24–26]. The apoptotic process can occur via multiple pathways depending on the cell type and the apoptotic stimuli, leading to limited cleavage of effector molecules (i.e. Bid, caspases, Bax). The mechanism of this translocation is not fully defined; a sphingosine-based lipid ceramide is believed to be the mediator [27]. What regulates the limited release of CatD from the lysosomal membrane, and the signaling mechanism(s) remains to be fully elucidated.
Under normal conditions, a minor fraction of pro-CatD is secreted and is detected in biological fluids [28,29]. In the mammary glands, CatD is secreted mostly at the apical surface, and this apical release varies considerably at different stages of development, with maximal secretion noted at early lactation [30]. However, basal release of single chain active CatD is also noted in lactating gland and is prompted by prolactin (PRL) [31]. The 23 kDa plasma-borne PRL binds prolactin receptors (PRLR) on the basolateral membrane of the mammary epithelial cells, promotes the basal transport of CatD-containing vesicles (which lack endocytic markers), and release of mature active CatD. This basolaterally secreted CatD (also catalytically active at pH greater than that of the lysosome), cleaves the 23 kDa PRL, generating 16 kDa fragment [32]. Notably, in the lactating mammary gland, the basal release of CatD has to be tightly regulated to circumvent the adverse catalytic effect(s) of the enzyme on the basement membrane proteins and the extracellular matrix (ECM). Indeed the expression of ECM-degrading proteases are repressed at lactation [33], but dramatically induced at involution to remodel the gland [34,35]. The bilateral release of CatD signifies distinct signaling mechanisms which are yet to be elucidated. More importantly, what distinguishes CatD for basolateral versus apical release remains unclear. The extent of phosphorylation of N-linked oligosaccharides, or the composition of the glycan structures on CatD (i.e. high mannose versus complex or hybrid), and the receptor chaperoning the enzyme, are believed to play a role, and will be discussed later in the text. It is noteworthy that modulators of lysosomal pH (i.e. chloroquine, or inhibitors of vacuolar ATPase) alter targeting and processing of CatD and increase its secretion [36,37].
In breast cancer, CatD is overexpressed, it’s processing and cellular location(s) are altered, and its secretion is highly elevated [11,12,38,39]. Consequently, it has become a marker of poor prognosis correlating with the prevalence of clinical metastasis [40]. However, direct involvement of this protease in the invasive and metastatic potential of breast cancer has not been demonstrated. The clue(s) to transformation of CatD from a “physiological guardian angle” to a “pathological demon” must lie in the signals which regulate its proper functions in development and under normal conditions. Alas, the complexity of CatD gene regulation, transcription, post-translational modifications, location(s) and specifically binding partners, render this a colossal task.
Functional Significance of CatD Post-translational Modifications
To date, the most studied post-translational modifications of CatD include proteolytic cleavages, glycosylation, phosphorylation, and nitration.
Proteolytic Cleavages
Conversion of the pre-pro-CatD to the active two-chain enzyme is a non-reversible process encompassing consecutive and highly regulated proteolytic steps [41,42]. Initially, the pre-peptide (20 amino acids), and the pro-peptide (44 amino acids) are sequentially removed to form the 48 kDa single-chain molecule [21,43]. Next, seven amino acids of the single chain’s NH2-terminus are removed by unidentified endo- and amino-peptidases, followed by removal of the sequence Ser-Ala-Ser-Ser-Ala-Ser-Ala-Leu (position 97–105). This modified single chain enzyme is cleaved by cysteine endopeptidase(s) to two chains, which undergo further processing to yield the active light and heavy chain CatD (Figure 1B). The significance of these precise proteolytic cleavages has eluded scientists, and with the increasing list of CatD functions, specifically during development and in embryonic stages, it is tempting to speculate that “functional specificity” tightly regulates the generation of CatD cleavage products. In this context, our laboratory has reported the differential processing of CatD during mammary gland development and remodeling (Figure 1C) [30]. These studies were first to demonstrate the plasticity of mammary tissue with respect to CatD production, proteolytic cleavage and activity. Notably, quiescent, non-lactating mammary epithelial cells have low constitutive levels of CatD in the pro-, single chain and two chain active enzyme format. At lactation, CatD’s cleavage profile remains comparable to non-lactating gland, while a considerable level of pro-CatD is apically released into the lumen. The level and apical release of pro-CatD diminishes considerably as lactation progresses. At the onset of involution, CatD is tyrosine nitrated [43], its processing is halted at the single chain active enzyme form [30] (Figure 1C). The generation of the mature two-chain active enzyme is resumed after 48h and peaks at days 3–4 of involution. In depth analysis of signals directing these proteolytic processes could unravel the specific regulatory checkpoints that have gone awry in cancer. Comparison of CatD production and processing profiles of normal mammary epithelial cells and breast cancer cell lines MCF-7 and MDA-MB231 indicates dramatic increase in the production, differences in the apparent molecular mass and level of the single chain form in the latter two cell lines (Figure 2A, Khalkhali-Ellis, unpublished observations). Specifically, pro-CatD secretion in cancer cell lines is ~30–40 folds higher than that of normal mammary epithelial cells, which further confirms the reported studies of delayed processing, accumulation of the 52 and 48 kDa forms, and secretion of over 50% of the pro-CatD in majority of cancer cell lines [12, 44].
Figure 2.
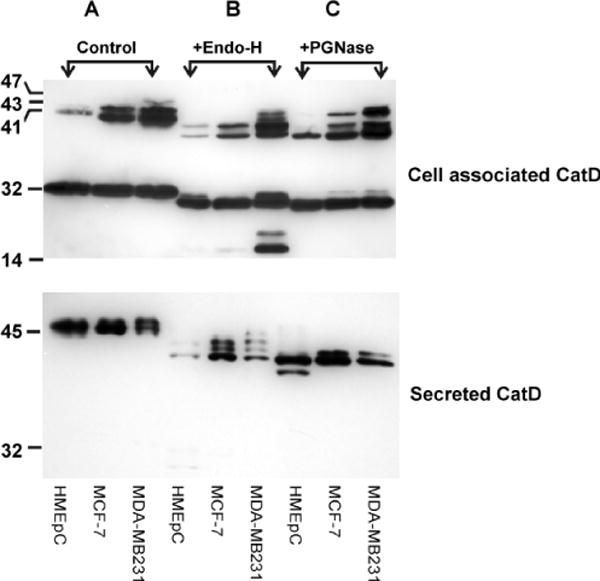
CatD cleavage products and N-glycan structures in normal mammary epithelial cells (HMEpCs) compared to breast cancer cell lines MCF-7 and MDA-MB231. Cytosolic fractions (25 μg total protein), and conditioned media (CM) from HMEpCs and breast cancer cell lines MCF-7 and MDA-MB231 were treated with or without endoglycosidase H (Endo-H, to remove the chitobiose core of high mannose and some hybrid oligosaccharides), and peptide-N-glycosidase F (PGNase, to remove high mannose, hybrid and complex glycans), then subjected to SDS-PAGE (10% acrylamide gel) and Western blot analysis. CM from the three cell lines was concentrated prior to treatment (HMEpC: 35×, MCF-7 and MDA-MB231: 2×). Differences in the abundance of the processed forms (A), and the N-glycan moieties of CatD (B&C), between normal mammary epithelial cells and breast cancer cell lines are evident in these Western blots. Note the preferential presence of multiple high mannose N-glycan structures (indicated by the appearance of multiple bands following Endo-H treatment) of the CM from the cancer cell lines.
Glycosylation
CatD is glycosylated on two asparagine residues 134 and 263 in the rough endoplasmic reticulum (ER). These glycan chains are later modified by phosphorylation of their mannose residues in the Golgi compartment. This post-translational modification of CatD is most extensively studied and elegantly described in several review articles [45], and will not be addressed here. However, glycosylation of CatD has no effect on protein folding and enzymatic activity, but it plays a significant role in the proper targeting to the lysosomal compartment, and most probably other sites of CatD function [36,41]. Disparate glycosylation of CatD was first demonstrated in MCF-7 breast cancer cells [12], and further confirmed in other breast cancer cell lines [46]. Studies from our laboratory also revealed the majority of breast cancer cell lines harbor differential glycosylation patterns (specifically in the secreted pro-CatD), with prevalence of endoglycosidase H (EndoH) sensitive N-glycan structures (Figure 2 B and 2C, Khalkhali-Ellis, unpublished observations) [47]. Indeed, the N-glycan structures in the tumor and serum of breast cancer patients (and other types of cancer) are distinctly different from that of normal tissue [47,48], and probably results from altered expression of glycosl transferases in cancer patients [49]. The effect of glycan alterations on lysosomal targeting and secretion of CatD remains to be investigated.
Nitration
Nitration of tyrosine residues is a common post-translational modification of proteins, and depending on the nature of the protein and the position of the tyrosine residues could lead to both loss or gain of protein activity [50–52]. CatD has a highly conserved nitration motif spanning residues 165–173. In rat mammary gland, the onset of involution prompts nitration of CatD on tyrosine residue 168, and the process could be a signaling pathway in mammary gland involution [43]. Interestingly, tyrosine nitration of CatD at involution increases CatD activity [43].
CatD Protein Binding Partners
The complex nature of CatD traffic (“inside→out”, or “outside→in”), its partitioning into different subcellular compartments, and bilateral secretion are indicative of different functions and command the presence of site-specific binding partners. To date, only a handful of CatD binding proteins have been identified, and are discussed here.
Man-6PR
The cation-independent Man-6PR has been the first and the best studied binding partner for CatD [53]. This receptor is concentrated in the cis Golgi, where it binds the newly translated CatD for lysosomal targeting [54]. A small fraction (~3–10%) of Man-6PR is present at the plasma membrane, and functions in endocytosis of Man-6P-containing ligands [53]. In addition, Man-6PR binds other ligands such as insulin-like growth factor II and retinoic acid, in a Man-6P-independent manner, and both ligands regulate Man-6-PR mediated trafficking of CatD [55,56]. In polarized Caco-2 human intestinal epithelial cell line, Man-6PR distributes CatD to apical and basolateral areas [57]. The basolateral sorting of Man-6-PR depends on the recognition of a sequence located in its cytoplasmic region and distinct from that of endosomal trafficking motifs [57]. A similar bilateral distribution of pro-CatD is noted in lactating mammary gland [37], however, the signaling mechanism, and the involvement of Man-6PR have not been determined. Importantly, increased lysosomal pH results in selective increase in CatD’s basolateral secretion [57], and defective acidification of lysosomes in breast cancer cell lines contributes to increased CatD secretion [58].
Prosaposin
This highly conserved, heavily glycosylated protein (Pro-Sap) is the precursor of sphingolipid activator proteins, saposins A, B, C and D [59]. In the ER, Pro-Sap binds pro-CatD, and the complex is transported to the Golgi and the lysosomes independent of Man-6PR [60]. In the acidic lysosomes, Pro-Sap activates pro-CatD, and the active enzyme cleaves Pro-Sap to generate saposins A-D [61]. The pro-CatD:Pro-Sap complex is also secreted and is detected in the body fluids (i.e. milk, serum, seminal fluid, and conditioned media of cultured cells) [62–64]. Contrary to their common intercellular and secretory pathways, their endocytic re-entry into the cell is mediated by different receptors and is cell–specific, as cancer cells don’t endocytose Pro-Sap.
Pro-Sap’s functional diversity is highlighted in its ability to sustain the stemness feature(s) of human embryonic neural progenitor cells, its neurotrophic and anti-apoptotic functions, and its involvement in the prostate gland development [65–67]. The latter has led to Pro-Sap’s critical function in prostate cancer [68–70], and underscores its utility as a therapeutic agent [70].
Pro-Sap’s function in the mammary gland remains mostly unexplored. However, immortalized mammary epithelial cells and majority of breast cancer cell lines secret significant levels of Pro-Sap, and estrogen stimulates the process [63]. In xenograft experiments using MCF-7 breast cancer cells, exogenous addition of recombinant Pro-Sap induced estrogen receptor alpha expression (via MAPK-signaling pathway), stimulated proliferation of MCF-7 cells, and tumor growth [71]. Analogous to CatD, Pro-Sap is heavily glycosylated, and changes in its glycan structure (as noted in cancer) could significantly affect its targeting, secretion and binding to CatD.
Ceramide
The sphingosine-based lipid ceramide regulates cellular proliferation, differentiation and apoptosis. However, its function depends on the subcellular topology of its production and is cell-type specific [72,73]. Ceramide binds CatD, resulting in the autocatalytic proteolysis of the pre-pro CatD to the active 48/32 kDa isoforms [74]. Indeed, the deficiency of the lysosomal enzyme acid sphingomyelinase, which is involved in the generation of ceramide, is associated with decreased CatD activity [74]. The ceramide:CatD interaction connects lipid rafts and lysosomal compartments, causes a limited permeablization of the lysosomal membrane and the release of CatD in the cytosol, a process intrinsic to the generation of the apoptotic cascade. When in the cytosol, CatD cleaves Bid, resulting in the release of cytochrome c from the mitochondria and activation of caspase-9 and -3 [27].
Notably, treatment of breast cancer cell lines with chemotherapeutic agents such as Taxol and Reservatol increases cellular levels of ceramide and induces apoptosis. In MDA-MB-468 breast cancer cells, ceramide is generated within 6h of exposure to Taxol, apoptosis is detectable 12 h post treatment, and by 24 h the apoptotic index reaches six times that of untreated cells [75,76]. Use of cell-permeable C6-ceramide as a medium supplement also renders the cells sensitive to the drugs [75,76]. However, prolonged drug treatment could also activate the glycosyl ceramide synthase gene expression, a positive feedback loop which is anti-apoptotic and drives cellular resistance to ceramide-generating chemotherapy approaches [77,78].
Low-density lipoprotein receptor-related protein-1 (LRP1)
LRP1 is a widely expressed type I integral membrane protein with diverse biological functions. It is composed of an extracellular α-chain in a non-covalent association with the membrane-spanning β-chain [79]. Deletion of the LRP1 gene leads to lethality in mice, revealing a critical, but yet undefined role in development [80]. In pathological conditions such as atherosclerosis, cancer, and nervous system injury, LRP1 recruits inflammatory cells, and regulates their survival [81].
Beaujouin and colleagues have recently identified LRP1 as CatD binding partner. Their findings indicate pro-CatD secreted by MCF-7 cancer cells binds to extracellular domain of the LRP1 β chain present on the surface of fibroblasts and triggers its growth. This binding might play a critical role in tumor growth via a paracrine loop [82]. The receptor is highly expressed by tumor fibroblasts and at low levels by breast cancer cells. However, its expression is induced in hypoxic conditions and could affect cancer growth via an autocrine loop.
Maspin
Our laboratory has uncovered partnership between a mammary non-inhibitory serpin (Maspin) and CatD [83]. Our studies indicate that in mammary epithelial cells Maspin is deposited in the extracellular matrix, and functions as a positional cue in directing CatD-mediated ECM degradation [83]. Notably, in breast cancer Maspin is among the very early genes silenced by promoter methylation [84], while CatD is excessively produced and aberrantly secreted. In the absence of Maspin, CatD-mediated matrix degradation proceeds unrestricted, thus facilitating the progression to metastasis. Maspin-CatD interaction is also a limiting factor to CatD’s secretion. This is based on the observation that transfection of breast cancer cell lines (which are often devoid of Maspin and secrete ~50% of their CatD) with Maspin gene results in considerable reduction in CatD secretion (Khalkhali-Ellis, unpublished observation). More importantly, Maspin transfection sensitizes these cells to apoptotic cell death by some chemotherapeutic agents and will be discussed later in the text [37].
Sortilin
Sortilin, also known as neurotensin receptor 3 [85], is a multifunctional type I transmembrane glycoprotein with striking structural similarities to Man-6-PR (and other transport proteins such as vacuolar sorting protein Vps10p). A major portion of sortilin resides in the Golgi (co-localized with Man-6PR300), cycles to the endosomal compartment [85,86], delivering Pro-Sap, acid sphingomyelinase, CatD and cysteine proteinase CatH to the lysosome [87,88]. Based on knock out approaches, Sortilin is the sole receptor for lysosomal delivery of CatH, while both Man-6PR and Sortilin are required for the efficient transport of CatD [89]. Interestingly, lysosomal transport of Pro-Sap is also affected, and as Pro-Sap’s traffic to lysosomes is independent of Man-6-PR, the alternative receptor for Pro-Sap’s lysosomal trafficking remains to be identified. The presence of sortilin in human cancer cell lines derived from colon, pancreas, prostate and ovarian carcinoma has been reported [90], but its expression in normal mammary tissue or breast cancer cell lines remains unexplored.
Cystatin C
Cystatin C, is a small ubiquitously expressed protein found in nearly all body fluids, and is the most potent endogenous extracellular inhibitor of cysteine cathepsins [91]. It blocks cystein cathepsin-mediated invasion, inhibits and antagonizes TGF-β signalling in normal and cancer cells by physically interacting with the TGF-β receptor II, thereby preventing TGF-β binding [92]. Laurent-Matha and colleagues [93] have employed yeast-2-hybrid screening approach and identified cystatin C as a binding partner for secreted pro-CatD. Their studies indicate that pro-CatD secreted by breast cancer cells binds to and degrades cystatin C in the extracellular environment. The reduced level (or absence) of this inhibitor leads to increased proteolytic activity of cysteine cathepsins (specifically Cathepsin B), and indirectly promotes tumor progression and metastasis. Whether cystatin C is involved in mammary gland development and remodeling is not known. However, it is quite likely that it’s functional inhibition of TGF-β signalling, and cystein cathepsins degradative effect would be crucial in mammary gland development and remodelling. Collectively, Man-6-PR, sortilin and LRP1 bind CatD extracellularly, and are involved in its outside→in transport, while Man-6-PR, Pro-Sap and ceramide bind CatD intercellularly. Maspin, Pro-Sap and cystatin C also function in the extracellular environment, but are not involved in the outside→in transport of CatD. The majority of these binding partners have overlapping functions, and the hierarchy of their binding precedence is presently undefined.
CatD and Its Binding Partners in Drug Sensitivity
The prediction of patient response to drug therapy is the ultimate goal of pharmacogenomics research. This is specifically important in cancer treatment, and instrumental in selecting effective chemotherapeutic agent tailored for individual patients. Unfortunately to date, the administration of ineffective chemotherapeutic agents often diminishes the quality of life for many cancer patients. Expression profile analyses have identified genes (specifically CatD) associated with sensitivity to anticancer drugs, however [94,95], the very complex nature of the drug response in a multi-organ system compared to single cells renders the clinical application of such findings quite challenging. The critical role of CatD in apoptosis [24–27], highlights a possible function for its binding partners in these processes. Our laboratory has exploited CatD partnership with Maspin to investigate such a possibility. IFN-γ is a widely used chemotherapeutic agent in many types of cancer; however, the majority of breast cancer cell lines are refractory to this cytokine [96, Z-Khalkhali-Ellis, unpublished observation]. This non-conformity was determined to be (at least in part) due to silencing of Maspin and deregulated expression and secretion of CatD [37]. Notably, IFN-γ reduces proliferation, changes vacuolar pH, alters CatD processing, and disrupts cell polarity, ultimately resulting in cell death. While, breast cancer cell lines devoid of Maspin are refractory to this cytokine. Maspin transfection of these cell lines reduces their pro-CatD secretion, and renders them responsive to IFN-γ(Figure 3).
Figure 3.
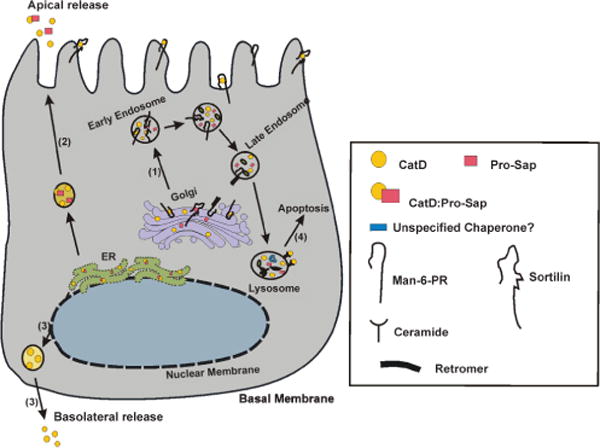
Model depicting different routes of CatD trafficking in association with its binding partners in polarized normal mammary epithelial cells, and their relevance to cancer: 1). When in the Golgi, the Man-6-P tagged CatD binds Man-6-PR (and/or sortilin) and is transported to the endosomal compartment. In the acidic environment, the complex is dissociated and Man-6-PR returns to the membrane with the retromer complex, while CatD is cleaved and processed in the late endosome and lysosomes. 2). Under normal conditions ≤5% of pro-CatD (either alone or in a complex with Pro-Sap) is secreted from the ER. 3). In polarized epithelial cells, the basolateral release of CatD is also observed, the binding partner in this case is unknown. However, in Caco-2 colon epithelial cell line, Man-6PR binds CatD and transports it basolateraly. 4). Generation of ceramide by acid sphingomyelinase results in the limited permeability of lysosomal membrane and release of CatD, leading to the induction of apoptosis. The majority of these pathways are altered in breast cancer. In pathway 1, the reduced acidification of endosomal/lysosomal compartment noted in cancer cells affects proper processing of CatD, resulting in increased secretion of pro-CatD. Routes 2 and 3 are greatly elevated and could lead to excessive ECM degradation. Route 4 could be equally affected by changes in vacuolar acidification, which alters CatD processing, its lysosomal release and participation in apoptosis.
Exciting as these findings might be, detailed mechanistic analyses are required to determine the crosstalk between CatD and its binding partners (in response to specific drug) in a cell-specific context under physiological conditions. The complex overlapping interactions of distinct binding partners with CatD requires further molecular dissection and could prove to be quite illuminating. We propose that this selection process is cell- (or tissue-) specific, and dictated by the precise function assigned for CatD in that particular environment. In addition, the topological presence of the binding partner(s) and avidity of their binding related to the particular form of CatD could significantly affect the selection process and are presently unidentified. These binding partners could be involved in directing signals which regulate the generation of CatD’s proteolytic fragments and post translational modifications.
In conclusion, CatD traverses boundaries in terms of form, function and location. The range and complexity of biological processes reliant on this enzyme, and the diversity of its degradomes are testament to its fundamental role in mammary gland development and tissue homeostasis. As such, it’s deregulated syntheses, processing, targeting and activity (noted specifically in breast cancer) are the key to transition from a normal tissue microenvironment to one which favors tumor growth and progression to metastasis. In brief, deciphering CatD as “physiological guardian angle” could direct us to what has unleashed the “pathological demon”.
Acknowledgments
Supported by: NIH/CA 75681
Contributor Information
Zhila Khalkhali-Ellis, Stanley Manne Children’s Research Institute, Northwestern University Feinberg School of Medicine, 2300 Children’s Plaza, Box 222, Chicago, Illinois, 60614-3394, USA.
Mary J.C. Hendrix, Robert H. Lurie Comprehensive Cancer Center, Northwestern University Feinberg School of Medicine, 2300 Children’s Plaza, Box 222, Chicago, Illinois, 60614-3394, USA
References
- 1.Willstätter R, Bamann E. Über die Proteasen der Magenschleimhaut. Erste Abhandlung über die Enzyme der Leukozyten. Hoppe-Seylers Z Physiol Chemie. 1929;180:127–143. [Google Scholar]
- 2.Fruton JS, Irving GW, Bergmann M. On the proteolytic enzymes of animal tissues. II. The composite nature of beef spleen cathepsin. J Biol Chem. 1941;138:249–262. [Google Scholar]
- 3.Anson ML. The Purification of Cathepsin. J Gen Physiol. 1940;23:695–704. doi: 10.1085/jgp.23.6.695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.De Duve C, Pressman Bc, Gianetto R, Wattiaux R, Appelmans F. Tissue fractionation studies. 6. Intracellular distribution patterns of enzymes in rat-liver tissue. Biochem J. 1955;60:604–617. doi: 10.1042/bj0600604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Press EM, Porter RR, Cebra J. The isolation and properties of a proteolytic enzyme, cathepsin D, from bovine spleen. Biochem J. 1960;74:501–514. doi: 10.1042/bj0740501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Huang JS, Huang SS, Tang J. Cathepsin D isozymes from porcine spleens. Large scale purification and polypeptide chain arrangements. J Biol Chem. 1979;254:11405–11417. [PubMed] [Google Scholar]
- 7.Tappel A, Zalkin H, Caldwell Ka, Desai ID, Shibko S. Increased lysosomal enzymes in genetic muscular dystrophy. Arch Biochem Biophys. 1962;96:340–346. doi: 10.1016/0003-9861(62)90418-6. [DOI] [PubMed] [Google Scholar]
- 8.Fell HB, Dingle JT. Studies on the mode of action of excess of vitamin A. 6. Lysosomal protease and the degradation of cartilage matrix. Biochem J. 1963;87:403–408. doi: 10.1042/bj0870403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dao T, Bock FG, Greiner MJ. Mammary carcinogenesis by 3-methylcholanthrene. II. Inhibitory effect of pregnancy and lactation on tumor induction. J Natl Cancer Inst. 1960;25:991–1003. [PubMed] [Google Scholar]
- 10.Shamberger RJ. Lysosomal enzyme changes in growing and regressing mammary tumours. Biochem J. 1969;111:375–383. doi: 10.1042/bj1110375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Reid WA, Valler MJ, Kay J. Immunolocalization of cathepsin D in normal and neoplastic human tissues. J Clin Pathol. 1986;39:1323–1330. doi: 10.1136/jcp.39.12.1323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Capony F, Rougeot C, Montcourrier P, Cavailles V, Salazar G, et al. Increased secretion, altered processing, and glycosylation of procathepsin D in human mammary cancer cells. Cancer Res. 1989;49:3904–3909. [PubMed] [Google Scholar]
- 13.Moallem SA, Hales BF. Induction of apoptosis and cathepsin D in limbs exposed in vitro to an activated analog of cyclophosphamide. Teratology. 1995;52:3–14. doi: 10.1002/tera.1420520103. [DOI] [PubMed] [Google Scholar]
- 14.Saftig P, Hetman M, Schmahl W, Weber K, Heine L, et al. Mice deficient for the lysosomal proteinase cathepsin D exhibit progressive atrophy of the intestinal mucosa and profound destruction of lymphoid cells. EMBO J. 1995;14:3599–3608. doi: 10.1002/j.1460-2075.1995.tb00029.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cavaillès V, Augereau P, Rochefort H. Cathepsin D gene is controlled by a mixed promoter, and estrogens stimulate only TATA-dependent transcription in breast cancer cells. Proc Natl Acad Sci U S A. 1993;90:203–207. doi: 10.1073/pnas.90.1.203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Westley B, Rochefort H. A secreted glycoprotein induced by estrogen in human breast cancer cell lines. Cell. 1980;20:353–362. doi: 10.1016/0092-8674(80)90621-2. [DOI] [PubMed] [Google Scholar]
- 17.Touitou I, Cavaillès V, Garcia M, Defrenne A, Rochefort H. Differential regulation of cathepsin D by sex steroids in mammary cancer and uterine cells. Mol Cell Endocrinol. 1989;66:231–238. doi: 10.1016/0303-7207(89)90035-x. [DOI] [PubMed] [Google Scholar]
- 18.Wang F1, Duan R, Chirgwin J, Safe SH. Transcriptional activation of cathepsin D gene expression by growth factors. J Mol Endocrinol. 2000;24:193–202. doi: 10.1677/jme.0.0240193. [DOI] [PubMed] [Google Scholar]
- 19.Cavailles V, Garcia M, Rochefort H. Regulation of cathepsin-D and pS2 gene expression by growth factors in MCF7 human breast cancer cells. Mol Endocrinol. 1989;3:552–558. doi: 10.1210/mend-3-3-552. [DOI] [PubMed] [Google Scholar]
- 20.Baranski TJ, Koelsch G, Hartsuck JA, Kornfeld S. Mapping and molecular modeling of a recognition domain for lysosomal enzyme targeting. J Biol Chem. 1991;266:23365–23372. [PubMed] [Google Scholar]
- 21.Gieselmann V, Pohlmann R, Hasilik A, Von Figura K. Biosynthesis and transport of cathepsin D in cultured human fibroblasts. J Cell Biol. 1983;97:1–5. doi: 10.1083/jcb.97.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kornfeld S, Mellman I. The biogenesis of lysosomes. Annu Rev Cell Biol. 1989;5:483–525. doi: 10.1146/annurev.cb.05.110189.002411. [DOI] [PubMed] [Google Scholar]
- 23.Cantor AB, Baranski TJ, Kornfeld S. Lysosomal enzyme phosphorylation. II. Protein recognition determinants in either lobe of procathepsin D are sufficient for phosphorylation of both the amino and carboxyl lobe oligosaccharides. J Biol Chem. 1992;267:23349–23356. [PubMed] [Google Scholar]
- 24.Biddere N, Lorenzo HK, Carmona S, Laforge M, Harper F, et al. Cathepsin D triggers Bax activation, resulting in selective apoptosis-inducing factor relocation in T lymphocytes entering the early commitment phase to apoptosis. J Biol Chem. 2003;278:31401–11. doi: 10.1074/jbc.M301911200. [DOI] [PubMed] [Google Scholar]
- 25.Minarowska A, Minarowski L, Karwowska A, Gacko M. Regulatory role of cathepsin D in apoptosis. Folia Histochem Cytobiol. 2007;45:159–163. [PubMed] [Google Scholar]
- 26.Appelqvist H, Johansson AC, Linderoth E, Johansson U, Antonsson B, et al. Lysosome-mediated apoptosis is associated with cathepsin D-specific processing of bid at Phe24, Trp48, and Phe183. Ann Clin Lab Sci. 2012;42:231–242. [PubMed] [Google Scholar]
- 27.Heinrich M, Neumeyer J, Jakob M, Hallas C, Tchikov V, et al. Cathepsin D links TNF-induced acid sphingomyelinase to Bid-mediated caspase-9 and -3 activation. Cell Death Differ. 2004;11:550–563. doi: 10.1038/sj.cdd.4401382. [DOI] [PubMed] [Google Scholar]
- 28.Zühlsdorf M, Imort M, Hasilik A, von Figura K. Molecular forms of beta-hexosaminidase and cathepsin D in serum and urine of healthy subjects and patients with elevated activity of lysosomal enzymes. Biochem J. 1983;213:733–740. doi: 10.1042/bj2130733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Davies PH, Messer M. Intestinal cathepsin B and D activities of suckling rats. Biol Neonate. 1984;45:197–202. doi: 10.1159/000242004. [DOI] [PubMed] [Google Scholar]
- 30.Margaryan NV, Kirschmann DA, Lipavsky A, Bailey CM, Hendrix MJ, et al. New insights into cathepsin D in mammary tissue development and remodeling. Cancer Biol Ther. 2010;10:457–466. doi: 10.4161/cbt.10.5.12534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lkhider M, Castino R, Bouguyon E, Isidoro C, Ollivier-Bousquet M. Cathepsin D released by lactating rat mammary epithelial cells is involved in prolactin cleavage under physiological conditions. J Cell Sci. 2004;117:5155–64. doi: 10.1242/jcs.01396. [DOI] [PubMed] [Google Scholar]
- 32.Castino R, Delpal S, Bouguyon E, Demoz M, Isidoro C, et al. Prolactin promotes the secretion of active cathepsin D at the basal side of rat mammary acini. Endocrinology. 2008;149:4095–4105. doi: 10.1210/en.2008-0249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sorrell DA, Szymanowska M, Boutinaud M, Robinson C, Clarkson RW, et al. Regulation of genes encoding proteolytic enzymes during mammary gland development. J Dairy Res. 2005;72:433–441. doi: 10.1017/S0022029905001202. [DOI] [PubMed] [Google Scholar]
- 34.Radisky DC, Hartmann LC. Mammary involution and breast cancer risk: transgenic models and clinical studies. J Mammary Gland Biol Neoplasia. 2009;14:181–191. doi: 10.1007/s10911-009-9123-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Khokha R, Werb Z. Mammary gland reprogramming: metalloproteinases couple form with function. Cold Spring Harb Perspect Biol. 2011;3 doi: 10.1101/cshperspect.a004333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Rosenfeld MG, Kreibich G, Povo D, Kato K, Sabatini DD. Biosynthesis of lysosomal hydrolases: Their synthesis in bound polysomes and the role of co- and post-translational processing in determining their subcellular distribution. J Biol Chem. 1982;93:135–143. doi: 10.1083/jcb.93.1.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Khalkhali-Ellis Z, Abbott DE, Bailey CM, Goossens W, Margaryan NV, et al. IFN-gamma regulation of vacuolar pH, cathepsin D processing and autophagy in mammary epithelial cells. J Cell Biochem. 2008;105:208–218. doi: 10.1002/jcb.21814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Johnson MD, Torri JA, Lippman ME, Dickson RB. The role of cathepsin D in the invasiveness of human breast cancer cells. Cancer Res. 1993;53:873–877. [PubMed] [Google Scholar]
- 39.Benes P, Vetvicka V, Fusek M. Cathepsin D–many functions of one aspartic protease. Crit Rev Oncol Hematol. 2008;68:12–28. doi: 10.1016/j.critrevonc.2008.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Liaudet-Coopman E, Beaujouin M, Derocq D, Garcia M, Glondu-Lassis M, et al. Cathepsin D: newly discovered functions of a longstanding aspartic protease in cancer and apoptosis. Cancer Lett. 2006;237:167–179. doi: 10.1016/j.canlet.2005.06.007. [DOI] [PubMed] [Google Scholar]
- 41.Zaidi N, Maurer A, Nieke S, Kalbacher H. Cathepsin D: a cellular roadmap. Biochem Biophys Res Commun. 2008;376:5–9. doi: 10.1016/j.bbrc.2008.08.099. [DOI] [PubMed] [Google Scholar]
- 42.Minarowska A, Gacko M, Karwowska A, Minarowski Å. Human cathepsin D. Folia Histochem Cytobiol. 2008;46:23–38. doi: 10.2478/v10042-008-0003-x. [DOI] [PubMed] [Google Scholar]
- 43.Zaragozá R, Torres L, García C, Eroles P, Corrales F, et al. Nitration of cathepsin D enhances its proteolytic activity during mammary gland remodelling after lactation. Biochem J. 2009;419:279–288. doi: 10.1042/BJ20081746. [DOI] [PubMed] [Google Scholar]
- 44.Hasilik A, von Figura K, Conzelmann E, Nehrkorn H, Sandhoff K. Lysosomal enzyme precursors in human fibroblasts. Activation of cathepsin D precursor in vitro and activity of beta-hexosaminidase A precursor towards ganglioside GM2. Eur J Biochem. 1982;125:317–321. doi: 10.1111/j.1432-1033.1982.tb06685.x. [DOI] [PubMed] [Google Scholar]
- 45.Kornfeld R, Kornfeld S. Comparative aspects of glycoprotein structure. Annu Rev Biochem. 1976;45:217–237. doi: 10.1146/annurev.bi.45.070176.001245. [DOI] [PubMed] [Google Scholar]
- 46.Fortenberry SC, Schorey JS, Chirgwin JM. Role of glycosylation in the expression of human procathepsin D. J Cell Sci. 1995;108:2001–2006. doi: 10.1242/jcs.108.5.2001. [DOI] [PubMed] [Google Scholar]
- 47.Sleat DE, Chen TL, Raska K, Jr, Lobel P. Increased levels of glycoproteins containing mannose 6-phosphate in human breast carcinomas. Cancer Res. 1995;55:3424–3430. [PubMed] [Google Scholar]
- 48.Kyselova Z, Mechref Y, Kang P, Goetz JA, Dobrolecki LE, et al. Breast cancer diagnosis and prognosis through quantitative measurements of serum glycan profiles. Clin Chem. 2008;54:1166–1175. doi: 10.1373/clinchem.2007.087148. [DOI] [PubMed] [Google Scholar]
- 49.Patani N1, Jiang W, Mokbel K. Prognostic utility of glycosyltransferase expression in breast cancer. Cancer Genomics Proteomics. 2008;5:333–340. [PubMed] [Google Scholar]
- 50.Radi R. Protein tyrosine nitration: biochemical mechanisms and structural basis of functional effects. Acc Chem Res. 2013;46:550–559. doi: 10.1021/ar300234c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Abello N, Kerstjens HAM, Postma DS, Bischoff R. Protein tyrosine nitration: Selectivity, physicochemical and biological consequences, denitration, and proteomics methods for the identification of tyrosine-nitrated proteins. J Proteome Res. 2009;8:3222–3238. doi: 10.1021/pr900039c. [DOI] [PubMed] [Google Scholar]
- 52.García-Heredia JM1, Díaz-Moreno I, Nieto PM, Orzáez M, Kocanis S, et al. Nitration of tyrosine 74 prevents human cytochrome c to play a key role in apoptosis signaling by blocking caspase-9 activation. Biochim Biophys Acta. 2010;1797:981–993. doi: 10.1016/j.bbabio.2010.03.009. [DOI] [PubMed] [Google Scholar]
- 53.Ghosh P, Dahms NM, Kornfeld S. Mannose 6-phosphate receptors: new twists in the tale. Nat Rev Mol Cell Biol. 2003;4:202–212. doi: 10.1038/nrm1050. [DOI] [PubMed] [Google Scholar]
- 54.Brown WJ, Farquhar MG. The mannose-6-phosphate receptor for lysosomal enzymes is concentrated in cis Golgi cisternae. Cell. 1984;36:295–307. doi: 10.1016/0092-8674(84)90223-x. [DOI] [PubMed] [Google Scholar]
- 55.Kang JX, Bell J, Leaf A, Beard RL, Chandraratna RA. Retinoic acid alters the intracellular trafficking of the mannose-6-phosphate/insulin-like growth factor II receptor and lysosomal enzymes. Proc Natl Acad Sci U S A. 1998;95:13687–13691. doi: 10.1073/pnas.95.23.13687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Braulke T. Type-2 IGF receptor: a multi-ligand binding protein. Horm Metab Res. 1999;31:242–246. doi: 10.1055/s-2007-978725. [DOI] [PubMed] [Google Scholar]
- 57.Wick DA, Seetharam B, Dahms NM. Basolateral sorting signal of the 300-kDa mannose 6-phosphate receptor. Am J Physiol Gastrointest Liver Physiol. 2002;282:G51–60. doi: 10.1152/ajpgi.00028.2001. [DOI] [PubMed] [Google Scholar]
- 58.Kokkonen N, Rivinoja A, Kauppila A, Suokas M, Kellokumpu I, et al. Defective acidification of intracellular organelles results in aberrant secretion of cathepsin D in cancer cells. J Biol Chem. 2004;279:39982–39988. doi: 10.1074/jbc.M406698200. [DOI] [PubMed] [Google Scholar]
- 59.Kishimoto Y, Hiraiwa M, O’Brien JS. Saposins: structure, function, distribution, and molecular genetics. J Lipid Res. 1992;33:1255–1267. [PubMed] [Google Scholar]
- 60.Igdoura SA, Rasky A, Morales CR. Trafficking of sulfated glycoprotein-1 (prosaposin) to lysosomes or to the extracellular space in rat Sertoli cells. Cell Tissue Res. 1996;283:385–394. doi: 10.1007/s004410050549. [DOI] [PubMed] [Google Scholar]
- 61.Gopalakrishnan MM, Grosch HW, Locatelli-Hoops S, Werth N, Smolenová E, et al. Purified recombinant human prosaposin forms oligomers that bind procathepsin D and affect its autoactivation. Biochem J. 2004;383((Pt) doi: 10.1042/BJ20040175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Hineno T, Sano A, Kondoh K, Ueno S, Kakimoto Y, et al. Secretion of sphingolipid hydrolase activator precursor, prosaposin. Biochem Biophys Res Commun. 1991;176:668–674. doi: 10.1016/s0006-291x(05)80236-0. [DOI] [PubMed] [Google Scholar]
- 63.Campana WM, O’Brien JS, Hiraiwa M, Patton S. Secretion of prosaposin, a multifunctional protein, by breast cancer cells. Biochim Biophys Acta. 1999;1427:392–400. doi: 10.1016/s0304-4165(99)00036-7. [DOI] [PubMed] [Google Scholar]
- 64.Patton S, Carson GS, Hiraiwa M, O’Brien JS, Sano A. Prosaposin, a neurotrophic factor: presence and properties in milk. J Dairy Sci. 1997;80:264–272. doi: 10.3168/jds.S0022-0302(97)75934-4. [DOI] [PubMed] [Google Scholar]
- 65.Salvioli R, Ricci-Vitiani L, Tatti M, Scarpa S, De Maria R, et al. The secretion and maturation of prosaposin and procathepsin D are blocked in embryonic neural progenitor cells. Biochim Biophys Acta. 2008;1783:1480–1489. doi: 10.1016/j.bbamcr.2008.01.033. [DOI] [PubMed] [Google Scholar]
- 66.Sikora J, Harzer K, Elleder M. Neurolysosomal pathology in human prosaposin deficiency suggests essential neurotrophic function of prosaposin. Acta Neuropathol. 2007;113:163–175. doi: 10.1007/s00401-006-0148-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Morales CR, Zhao Q, El-Alfy M, Suzuki K. Targeted disruption of the mouse prosaposin gene affects the development of the prostate gland and other male reproductive organs. J Androl. 2000;21:765–775. [PubMed] [Google Scholar]
- 68.Koochekpour S, Lee TJ, Wang R, Culig Z, Delorme N, et al. Prosaposin upregulates AR and PSA expression and activity in prostate cancer cells (LNCaP) Prostate. 2007;67:178–189. doi: 10.1002/pros.20513. [DOI] [PubMed] [Google Scholar]
- 69.Koochekpour S, Hu S, Vellasco-Gonzalez C, Bernardo R, Azabdaftari G, et al. Serum prosaposin levels are increased in patients with advanced prostate cancer. Prostate. 2012;72:253–269. doi: 10.1002/pros.21427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Hu S, Delorme N, Liu Z, Liu T, Velasco-Gonzalez C, et al. Prosaposin down-modulation decreases metastatic prostate cancer cell adhesion, migration, and invasion. Mol Cancer. 2010;9:30. doi: 10.1186/1476-4598-9-30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Wu Y, Sun L, Zou W, Xu J, Liu H, et al. Prosaposin, a regulator of estrogen receptor alpha, promotes breast cancer growth. Cancer Sci. 2012;103:1820–1825. doi: 10.1111/j.1349-7006.2012.02374.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Kolesnick RN, Krönke M. Regulation of ceramide production and apoptosis. Annu Rev Physiol. 1998;60:643–665. doi: 10.1146/annurev.physiol.60.1.643. [DOI] [PubMed] [Google Scholar]
- 73.Green DR. Apoptosis and sphingomyelin hydrolysis. The flip side. J Cell Biol. 2000;150:F5–7. doi: 10.1083/jcb.150.1.f5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Heinrich M, Wickel M, Schneider-Brachert W, Sandberg C, Gahr J, et al. Cathepsin D targeted by acid sphingomyelinase-derived ceramide. EMBO J. 1999;18:5252–5263. doi: 10.1093/emboj/18.19.5252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Charles AG, Han TY, Liu YY, Hansen N, Giuliano AE, et al. Taxol-induced ceramide generation and apoptosis in human breast cancer cells. Cancer Chemother Pharmacol. 2001;47:444–450. doi: 10.1007/s002800000265. [DOI] [PubMed] [Google Scholar]
- 76.Scarlatti F, Sala G, Somenzi G, Signorelli P, Sacchi N, et al. Resveratrol induces growth inhibition and apoptosis in metastatic breast cancer cells via de novo ceramide signaling. FASEB J. 2003;17:2339–2341. doi: 10.1096/fj.03-0292fje. [DOI] [PubMed] [Google Scholar]
- 77.Liu YY, Yu JY, Yin D, Patwardhan GA, Gupta V, et al. A role for ceramide in driving cancer cell resistance to doxorubicin. FASEB J. 2008;22:2541–2551. doi: 10.1096/fj.07-092981. [DOI] [PubMed] [Google Scholar]
- 78.Morad SA, Cabot MC. Ceramide-orchestrated signalling in cancer cells. Nat Rev Cancer. 2013;13:51–65. doi: 10.1038/nrc3398. [DOI] [PubMed] [Google Scholar]
- 79.Herz J, Strickland DK. LRP: a multifunctional scavenger and signaling receptor. J Clin Invest. 2001;108:779–784. doi: 10.1172/JCI13992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lillis AP, Van Duyn LB, Murphy-Ullrich JE, Strickland DK. LDL receptor-related protein 1: unique tissue-specific functions revealed by selective gene knockout studies. Physiol Rev. 2008;88:887–918. doi: 10.1152/physrev.00033.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Gonias SL, Campana WM2. LDL receptor-related protein-1: a regulator of inflammation in atherosclerosis, cancer, and injury to the nervous system. Am J Pathol. 2014;184:18–27. doi: 10.1016/j.ajpath.2013.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Beaujouin M, Prébois C, Derocq D, Laurent-Matha V, Masson O, et al. Pro-cathepsin D interacts with the extracellular domain of the beta chain of LRP1 and promotes LRP1-dependent fibroblast outgrowth. J Cell Sci. 2010;123:3336–3346. doi: 10.1242/jcs.070938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Khalkhali-Ellis Z, Hendrix MJC. Elucidating the function of secreted Maspin: inhibiting Cathepsin D-mediated matrix degradation. Cancer Res. 2007;67:3535–3539. doi: 10.1158/0008-5472.CAN-06-4767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Futscher BW, Oshiro MM, Wozniak RJ, Holtan N, Hanigan CL, et al. Role for DNA methylation in the control of cell type specific maspin expression. Nat Genet. 2002;31:175–179. doi: 10.1038/ng886. [DOI] [PubMed] [Google Scholar]
- 85.Petersen CM, Nielsen MS, Nykjaer A, Jacobsen L, Tommerup N, et al. Molecular identification of a novel candidate sorting receptor purified from human brain by receptor-associated protein affinity chromatography. J Biol Chem. 1997;272:3599–3605. doi: 10.1074/jbc.272.6.3599. [DOI] [PubMed] [Google Scholar]
- 86.Carlo AS, Gustafsen C, Mastrobuoni G, Nielsen MS, Burgert T, et al. The pro-neurotrophin receptor sortilin is a major neuronal apolipoprotein E receptor for catabolism of amyloid-β peptide in the brain. J Neurosci. 2013;33:358–370. doi: 10.1523/JNEUROSCI.2425-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Lefrancois S, Zeng J, Hassan AJ, Canuel M, Morales CR. The lysosomal trafficking of sphingolipid activator proteins (SAPs) is mediated by sortilin. EMBO J. 2003;22:6430–6437. doi: 10.1093/emboj/cdg629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ni X, Morales CR. The lysosomal trafficking of acid sphingomyelinase is mediated by sortilin and mannose 6-phosphate receptor. Traffic. 2006;7:889–902. doi: 10.1111/j.1600-0854.2006.00429.x. [DOI] [PubMed] [Google Scholar]
- 89.Canuel M, Korkidakis A, Konnyu K, Morales CR. Sortilin mediates the lysosomal targeting of cathepsins D and H. Biochem Biophys Res Commun. 2008;373:292–297. doi: 10.1016/j.bbrc.2008.06.021. [DOI] [PubMed] [Google Scholar]
- 90.Hemmati S, Zarnani AH, Mahmoudi AR, Sadeghi MR, Soltanghoraee H, et al. Ectopic Expression of Sortilin 1 (NTR-3) in Patients with Ovarian Carcinoma. Avicenna J Med Biotechnol. 2009;1:125–131. [PMC free article] [PubMed] [Google Scholar]
- 91.Vray B, Hartmann S, Hoebeke J. Immunomodulatory properties of cystatins. Cell Mol Life Sci. 2002;59:1503–1512. doi: 10.1007/s00018-002-8525-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Sokol JP, Schiemann WP. Cystatin C antagonizes transforming growth factor beta signaling in normal and cancer cells. Mol Cancer Res. 2004;2:183–195. [PubMed] [Google Scholar]
- 93.Laurent-Matha V, Huesgen PF, Masson O, Derocq D, Prébois C, et al. Proteolysis of cystatin C by cathepsin D in the breast cancer microenvironment. FASEB J. 2012;26:5172–5181. doi: 10.1096/fj.12-205229. [DOI] [PubMed] [Google Scholar]
- 94.Nakatsu N, Yoshida Y, Yamazaki K, Nakamura T, Dan S, et al. Chemosensitivity profile of cancer cell lines and identification of genes determining chemosensitivity by an integrated bioinformatical approach using cDNA arrays. Mol Cancer Ther. 2005;4:399–412. doi: 10.1158/1535-7163.MCT-04-0234. [DOI] [PubMed] [Google Scholar]
- 95.Salter KH, Acharya CR, Walters KS, Redman R, Anguiano A, et al. An integrated approach to the prediction of chemotherapeutic response in patients with breast cancer. PLoS One. 2008;3:e1908. doi: 10.1371/journal.pone.0001908. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 96.Harvat BL, Jetten AM. Gamma-interferon induces an irreversible growth arrest in mid-G1 in mammary epithelial cells which correlates with a block in hyperphosphorylation of retinoblastoma. Cell Growth Differ. 1996;7:289–300. [PubMed] [Google Scholar]