

Abstract
We consider the evolution of cognition and the emergence of creative behaviour, in relation to vocal communication. We address two key questions: (i) what cognitive and/or social mechanisms have evolved that afford aspects of creativity?; (ii) has natural and/or sexual selection favoured human behaviours considered ‘creative’? This entails analysis of ‘creativity’, an imprecise construct: comparable properties in non-humans differ in magnitude and teleology from generally agreed human creativity. We then address two apparent problems: (i) the difference between merely novel productions and ‘creative’ ones; (ii) the emergence of creative behaviour in spite of high cost: does it fit the idea that females choose a male who succeeds in spite of a handicap (costly ornament); or that creative males capable of producing a large and complex song repertoire grew up under favourable conditions; or a demonstration of generally beneficial heightened reasoning capacity; or an opportunity to continually reinforce social bonding through changing communication tropes; or something else? We illustrate and support our argument by reference to whale and bird song; these independently evolved biological signal mechanisms objectively share surface properties with human behaviours generally called ‘creative’. Studying them may elucidate mechanisms underlying human creativity; we outline a research programme to do so.
Keywords: creativity, vocal communication, music, computational modelling, information theory
1. Introduction
One of the defining features of humanity is the ability to be creative. This ability is exhibited throughout human society and is a fundamental force in the development of humankind. However, the concept of creativity itself is shrouded in imprecision and subjectivity, making it difficult to address from a scientific perspective. One approach to the rational study of creativity in humans is to consider it from an evolutionary perspective, aiming to identify related behaviours in other species that can be studied without the cloud of human subjectivity that the word creativity entails.
Evolution, for the purpose of our argument, refers to the process of the gradual change of form and behaviour, as a result of differential advantages of some forms or behaviour over others. In the case of biological evolution, we talk of fitness and mean the numbers of offspring produced and surviving.
In the next section, we decompose the idea of creativity into tractable components, to allow us to examine whether music and other forms of vocal communication (including language in humans) share similar functional roots and may have evolved out of similar cognitive precursors. Whether a society values or eschews creativity, whether we agree on what constitutes good or bad music, or where one stands in the balance between humans as cognitive individuals and humans as cultural components is secondary to understanding the essence of the concepts.
2. Components of creativity
(a). Valuing creativity and creating value
We begin our decomposition of creativity with the relationship between perceived creativity and attributed value.
In Western society, ‘creativity’ is most commonly used to refer to the embodied cognitive process that gives rise to pieces of music, sculptures, paintings, poems and other things that are taken or presented as art. We, less conventionally, include science and engineering in our list of creative endeavours. Creativity is intensely context-dependent: reproducing the style of Monteverdi in the twentieth century would be regarded negatively as pastiche or plagiarism or an exercise of style replication. Creativity is heavily dependent on the nature of the creator: for example, Harold Cohen's AARON painter program [1] has made paintings that have hung in galleries and sold for thousands of dollars; his daughter was (in 1999) also a keen artist, producing (then) the kind of drawings one might expect from a 3-year old, for which most people would not be inclined to pay. Cohen, however, rates his daughter's creativity as much greater than that of his program [2]. Accordingly, he makes a distinction between Big-C creativity and Little-C creativity, also seen elsewhere in the literature [2], where Big-C is Picasso level, and Little-C is what AARON can manage. Margaret Boden [3] makes another perhaps more tractable distinction between psychological creativity—the act of generating an artefact that is novel and of value to an individual—and historical creativity—that of generating an artefact that is novel and valued in historical terms. However, this notion must be generalized: rather than two discrete kinds of creativity, value and novelty should not be thought of as simple quantities, but as relations between observers and the created artefact. Thus, for example, we can account for cycles of fashion: retro styles may be valued by both teenagers and their parents, the former enjoying their (relative) novelty and the latter doing exactly the opposite. We return to the matter of novelty below.
Value is dependent not only on the observer but also on the context in which the observation is made. It is present in many more pursuits than the artistic ones mentioned above, and in manifold ways. A prime example is mathematics, where the creation of the proof of a theorem is more highly valued if it is ‘elegant’, according to the principles of the particular branch of mathematics to which it applies; mathematics has its own aesthetics, as does engineering. Often, the aesthetic of one context is utterly incomprehensible, and even offensive, to observers comfortable in another: consider, for example, the riot that followed the première of Stravinsky's The Rite of Spring in the 1920s. Thus, the value relation is between not just the observer and the artefact, but between the observer and the artefact in a given context. Finally, value is also a function of the creator. Expectations are based on past experience. We are disappointed when our favourite author, admired musician or best-loved car company turns out a product that underperforms.
In summary, we treat value as a relation between an artefact, its creator and its observers and the context in which creation and observation takes place.
(b). Exploration, transformation and the paradigm shift
Boden [3] also introduces an important philosophical distinction, between exploratory creativity, where the conceptual space being explored is fixed (though possibly not all visible, and possibly infinite) and exploration occurs within that space (for example, different songs in a particular style), and transformational creativity in which the space itself is subject to change (developing from one style to another). Coupled with successful persuasion, transformational creativity is what leads to a paradigm shift in Kuhn's philosophy of science [4]. Boden proposes that Little-C creativity is exploratory, and Big-C creativity is transformational, but history is littered with exceptions to this: Mozart, for example, perfected a style that Haydn introduced, but Mozart is universally regarded as the greater creator. Wiggins [5] shows that, in any case, transformational creativity is formally exploratory creativity at the meta-level, where the conceptual space of artefacts is replaced by the conceptual space of conceptual spaces. This way of thinking, where the conceptual space can be taken to define the class of artefacts at which a creator is aiming, yields some elegant ways of discussing what happens when a creator pushes the boundaries of the expected, in a process taxonomized as different kinds of aberration by Wiggins [5]. This concept allows further objective, mechanistic, description and prediction of creative behaviour [5].
Humpback whale song has a nearly invariant pattern of theme transitions, so much so that Frumhoff [6] called the few backwards transitions therein ‘aberrant’ (though the common terminology here is coincidental). There also are three different kinds of theme in humpback song. The variation in structure is somewhat like a theme and variations; but they recur, so while they fit Boden's exploratory framework, their generation is not as free as that term might suggest. However, a restricted exploration of a (notionally) larger language can be modelled in this context as a conceptual space accompanied by a value measure, which filters out unvalued artefacts; we return to this below.
(c). Creativity: process or property?
Boden's approach raises some interesting questions concerning the conceptual space, and the attribution of value to artefacts in it: these things are separable, and the conceptual space is neutral with respect to both value and novelty: it inherently captures cognitive generation, not the subsequent value or novelty of that which is generated. Thus, the paintings of Harold Cohen (for he was a successful human-only artist before AARON), and of AARON, and of Cohen's daughter, all coexist, equally, in the conceptual space of paintings: it is only when they are evaluated by an observer (possibly the artist) that issues of novelty and value arise. In a less Western-centric perspective, we might conflate these two and argue that novelty is a kind of value, since in some cultures it does not have the high status accorded in the West, and in some it is actively eschewed in favour of the strict maintenance of tradition. This feature of creativity in the social context does not decrease the importance of novelty in the evolutionary context, as we shall see below.
Thus, we see that the production of the painting per se is not what guarantees its value: while, of course, the artefact must exist to be valued, it is interaction between production and (probably, at least initially, introspective) evaluation by an artist, and then by a social community, that identifies relative value and relative novelty, of both the artefact and the way it was made. Thus, we can decompose creativity into a series of steps and tests within a process, of which a ‘creative’ agent is capable, and can begin to study it. This is altogether more scientifically tractable than the philosophical debate about the ineffable nature of creativity itself.
(d). Size does not matter
Given the nature of the conceptual space as distinct from novelty and value of the concepts in the space, a natural question to ask is: need there be a difference in kind between big-C and little-C creativity? For some authors, the answer is clearly ‘no’: Plotkin [7] describes creativity as the sine qua non of everyday language generation; for others, the word should only be applied to the great creators of great historical import.
From the perspective of the current paper, this latter view is destructively problematic. We aim here to understand what evolutionary advantage may have been given to humans and/or animals by the ability to be creative. At the extreme level, it is hard to argue for evolutionary advantage in the authorship of very large scale created constructs such as symphonies. However, it has been argued that sexual selection may be a factor in smaller creativity [8,9]. Thus, if we were to restrict our definition to great human creators, ruling out minor creative acts, we would also rule out a priori the possibility of incremental development of creative faculties over evolutionary time. Instead, it is necessary to look for the roots of that ability both in humans and non-humans, with a view to understanding how the extreme (‘great creativity’ in the terms of the relevant culture) emerged from the ordinary (everyday creative activity). One unbiased way of approaching the question of how creativity evolved is thus to deconstruct the components and explore which ones exist in non-human animals and to what degree.
Equally, there is no scientific evidence to support the position that the ability to create did not evolve, step by step, as opposed to merely appearing fully formed in humans, and there is evidence of creativity or proto-creativity in other species, both in animals belonging to the same direct evolutionary lineage [10] and those more distantly related [11]. Therefore, when studying the development of creativity in our own and other species, it is necessary to admit and value the creation of less-than-amazing artefacts (as we do in our children) in order to encompass the overall development of the faculty, over evolutionary and ontogenetic time.
(e). Novelty and its perception
We now consider another key dimension of creativity, novelty, and the ability to perceive it. In Western culture, as we argue above, the attribution of creativity entails the attribution of novelty—various authors have argued that the human creative drive is the search for novelty [12], or, differently termed, curiosity [13]. While this is not the case in all cultures, the fact remains that novelty detection is a feature of creative behaviour—whether it is a feature to be valued or (in some contexts) suppressed. Regardless of one's response to novelty, the fact that one can respond to it means that it can be detected, and we propose that this is a fundamental component of creative behaviour. It is to be noted, however, that too much novelty prevents recognition, a fact embodied in the famous Wundt curve of hedonic response to novelty [14,15]: the inverted-U shape captures the notion that not enough variation is boring, while too much is unpleasantly incomprehensible, yielding a sweet spot in between. This is illustrated in figure 1.
Figure 1.
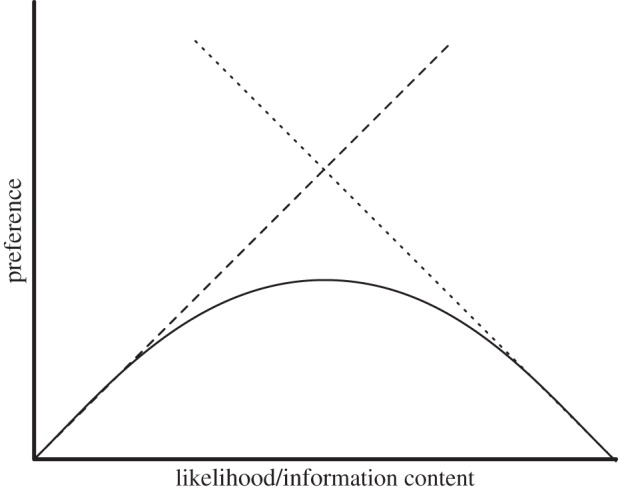
The Wundt curve illustrates the rise and fall of preference (y-axis) in perceivers for complexity of stimulus (x-axis). Very simple stimuli are uninteresting, while extremely complex ones are inaccessible, either case producing dissatisfaction. Intermediate levels of complexity, however, are preferred.
Novelty detection is a requirement for noticing changes in the environment, a feature all animals need for survival. When a pattern deviates from the known, it is novel and can signal good things (a new food source) or bad things (a new type of predator). Thus, animals need carefully to balance exploration of novelty, because it can open up new niches that enhance evolutionary fitness or are detrimental to it. The ability to detect novelty in the environment likewise allows animals to detect novelty in behaviour of conspecifics. Famous examples are the cultural transmission of novel behaviour through a population, as observed for sweet potato washing in Japanese macaques and opening the aluminium foil covers of milk bottles by chickadees [16].
Huron [17] extends this argument to affective response, exapted to music. Because the outcome of a novel experience is sometimes dangerous, it is appropriate for an animal to be alert and prepared for fight or flight in the face of novel circumstances. Thus, there is evolutionary incentive to perceive not just danger, but uncertainty and/or novelty in their own right. In humans, this situation is experienced as tension, leading to arousal and, in extremis, to fear, and simple observation suggests that other species share the same affective response. The experience of tension entails its subsequent release, which seems to be accompanied by positive affective states. Huron, following Meyer [18], suggests that tension thus stimulated by expectation, and its denial or fulfilment, is in large part responsible for affect stimulated by Western music, whose emotive content is frequently theoretically conceived as an ebb and flow of tension of various kinds. This affective experience is highly valued and is altogether more subtle and dynamic than the common labelling of emotional analysis of music as ‘tender’, ‘sad’, etc. [19,20].
It is possible to mathematically model expectations over a well-defined symbol system (musical melodies constructed from a known range of pitches and durations, or bird- or whale song categorized into appropriate symbol sets) using uncomplicated statistical techniques [21, ch. 9]. From these models, human melodic expectations can be estimated [22,23] and bird song can be modelled [24]. Pearce's model of Western tonal musical melody, IDyOM (Information Dynamics of Music) [23,25], predicts human expectations very well (r = 0.91 in four studies) [26,27]. Expectations are expressed as probability distributions over the set of symbols allowed (musical pitches, here). Given such a probability distribution, we can estimate the unexpectedness of an event drawn from it, using Shannon's information theory [28]. It is important to understand that this property is relative: it is computed in terms of the statistical model, so unexpectedness is relative to the information that the model contains about the set of sequences being modelled, and to the immediately precedent sequence. Thus, we can model an individual's memory and predict the unexpectedness of perceived events. Two quantities, entropy and information content, model uncertainty and unexpectedness, respectively [23]. More recent work on physiological and behavioural measures of human response to live music suggests that the unexpectedness value of pitch, calculated as above, explains a significant part of the variance in physiological measures (heart rate, skin conductivity) that correspond with arousal [29]. This constitutes evidence that unexpectedness in music correlates with arousal in listeners, and that both correlate with the predictions of the model. These model-driven empirical methods can be applied to any form of vocal communication, given enough examples.
Ikebuchi et al. [30] showed that female Bengalese finch hearts respond with tachycardia to more complex male song (that is, song with higher information content). This is a result comparable with the human musical response outlined above [29]. Further investigation of these phenomena via the models introduced in §3 may yield understanding of the relationships between the birds' reaction to song and the humans' reaction to music.
Weiss et al. [31] found that when nightingales heard a playback consisting of song types with branch transition patterns, they responded with song types with bottleneck transition patterns. Conversely, when they heard song types with bottleneck transition patterns, they responded with song types that tended to be branching transitions in their population—that is, they responded with the unexpected. While it remains unclear why this behaviour would arise, the fact that it does so entails the ability to detect high and low-entropy distributions, and/or high and low information content, as in Pearce et al.'s human studies and Huron's evolutionary argument.
Here, then, is a scientific question which the study of creativity can ask: do species other than humans exhibit similar responses to novelty and/or complexity, and, if so, how does their behaviour inform our understanding of our own? Given a sufficient amount of song produced by a particular species, or even individual, we can construct a model of the sequences, using the above techniques, and generate new sequences from it, with particular information-theoretic properties (e.g. surprising, neutral or very obvious). We can monitor the response of the relevant animal to the constructed sequence, by means of judicious audio editing, and thus test hypotheses regarding the value of novelty and complexity in vocal display. This view of song construction raises the possibility that it is valued by its own species for some of the same reasons that humans value music: the effect of rising and falling tension caused by unexpectedness/information content/complexity. This is a testable hypothesis to which we return below.
3. Modelling the process of creativity
In order to study creativity effectively, we need a rigorous frame of reference, including the ability to simulate perception and creative generation. Historically, there are not many scientific theories of creativity, and those that do exist are fundamentally qualitative. We now survey them, in contrast to a newer, quantitative approach.
Wallas [32] focuses on the cognitive process of creativity. He identifies four parts of a sequence: Preparation, in which the creative goal is identified and considered; Incubation, during which conscious attempts at creativity are not made; Illumination, the moment of enlightenment when an idea appears in conscious awareness, sometimes called the ‘Aha!’ moment; and Verification, in which the new idea is applied. These ideas highlight a further distinction that is useful in focusing on creativity: that between conscious, or deliberate, creativity and non-conscious, or spontaneous, creativity [33]. The former of these is the creativity where, for example, a professional composer must produce a TV theme in too short a time to wait for inspiration: she consciously applies rules of her craft to create what is necessary. The latter is the creativity where an idea or concept appears in one's awareness, apparently without bidding, effort or intention, in the way described by Mozart as the beginning of his mode of creativity (Holmes [34]). Most human creativity processes, including Mozart's overall description, are probably a cyclic combination of the two. Wallas, however, is considering spontaneous creativity resulting from earlier conscious consideration, and he considers the illumination point to be the arrival of a spontaneously produced concept in consciousness: the ‘Aha!’ moment. Wallas' theory requires created artefacts to undergo Validation, where they are examined to make sure they are fit for purpose. This may suggest that the theory is meant to account for larger-scale acts of creativity than, for example, spontaneous sentence production; or maybe successful communication of meaning would fulfil the definition in this example. In any case, the theory does not propose an underlying mechanism, but rather describes a series of stages. As such, it at most provides an overarching framework for the study of creativity.
Guilford's model [35] is more qualitative, but does not contradict Wallas. Guilford proposes a phase of divergent thinking, where possibilities are opened, followed by one of convergent thinking, in which the creator homes in on her idea. Both phases could happen either consciously or non-consciously, and one can also imagine repeating cycles of the two phases. The model has less predictive power than Wallas', however, and we will not refer to it further. More recently Csikszentmihalyi [36] described the subjective experience of creativity, involving the state of flow; again, this lacks quantitative analysis and predictive power. A final theory worthy of mention is that of Koestler [37]: the cognitive operation of bisociation is proposed, enabling cognitive structures representing two or more ideas to be combined to produce new concepts. This theory, though convincing, is not specified with mathematical precision.
None of the four frameworks outlined above affords a quantitative means to examine creative processes in detail. A more recent hypothetical mechanism for a cognitive creative process is provided by the Information Dynamics Of Thinking (IDyOT) cognitive architecture [33,38], based on Baars' Global Workspace Theory [39] and using the same information-theoretic notions as the IDyOM model cited above [23]. The key idea is that cognitive creativity is a result of prediction, which itself is a means for managing information and action in the world. Statistical generators continually predict outcomes from sensory inputs, based on statistical models trained by unsupervised observation. They compete in terms of the information content of their predictions (quantified in terms of Shannon Information Theory [28]) for access to the Global Workspace (GW), which equates with conscious awareness. When an item enters the GW, it may be novel, or it may be a predictable part of an on-going experience; in the former case, creativity has happened, and passage into the GW corresponds with Wallas' moment of Illumination, the preceding activity being Incubation. What enters the GW is recorded in memory and becomes available for future prediction, and thus the cycle repeats. This theory gives a concrete mechanism for creative production, and is applicable directly to discrete and continuous symbolic data represented on a computer. Thus it can be applied to transcriptions of bird and whale song, with a view to comparing their information-theoretic properties. This approach, then, can be used directly on real data to make testable predictions about animal behaviour, as it has done for humans.
4. Affording creative behaviour
Charles Darwin described two primary mechanisms of selection as driving biological evolution: natural selection and sexual selection. The critical elements for evolution by natural selection are variation in traits within a population, differential reproduction of animals with the differing traits, and inheritance of the trait from one generation to the next. Sexual selection can be viewed as a special case of natural selection which acts on an individual's ability to mate. Some traits, for example ones that increase fighting ability, may improve an individual's ability to compete with members of the same sex for mating, while others, such as ornaments or song, may make a member of the other sex more likely to select an individual for mating.
The topic of mate choice is important for our discussion of selection for creative behaviour, especially for creativity in communication. Biologists have investigated a variety of modes of sexual selection for mate choice. The simplest selection would be for a character that provides a direct benefit, such as if a female bird chooses a male whose genes produced a tail of the optimal size for flight. But suppose males also use the tail in a display to impress females. Females might have a sensory bias to choose males with even larger tails than optimal for flight, because the display is more visible [40]. Here sexual selection might drive the evolution of tails that are longer than optimal under natural selection. And if a population of females have a preference for longer tails, then this could lead to a runaway process of evolution of longer and longer tails until the benefit from sexual selection is outweighed by other natural selection pressures [41]. The evolution of large complex ornaments in males raises the question of why a female should choose a male with a trait that may make it more visible to predators and less able to escape. Zahavi [42] argued that males with such a handicap might have to be better quality, thus suggesting that handicaps help a female choose a better quality male.
How does creative behaviour fit onto these categories? Creative behaviour could result in biological selective advantage in all the above cases. Perhaps the most celebrated case of animal innovation involves a young female Japanese macaque who invented the idea of washing the sand off potatoes in the ocean and then three years later, the idea of separating grain from sand by throwing the mixture in water and scooping out the floating grain. Both of these innovations would be selected because they improve foraging. This kind of innovation is particularly important in species capable of social learning so that beneficial innovations diffuse through the population. Creative behaviour may also be the substrate for sexual selection: mimicking the sounds of other birds and adding them to his own repertoire of song may signal to the female lyre bird that her mate has particularly good cognitive skills that will also help to raise their young and pass on his intelligence as well. This logic has been applied to a more specific issue for the songs of birds. Nowicki et al. [43] pointed out that the nuclei in the brain that control song develop during critical periods of development. If a young bird does not have adequate nutrition at this time, it may suffer broader developmental problems. They reasoned that large and complex repertoires of song may indicate a history of good nutrition, and they suggest that females might select males with large and complex song repertoires for this reason [43]. Or creative behaviour could evolve as a by-product of something else: the need to explore to find new food sources or new territories might have selected animals that are less neophobic, and more curious, leading to more novel behaviours—not all of them necessarily beneficial to survival and reproduction. Thus, the expense of creative behaviour in terms of time, energy, and risk, which might at first seem problematic, can be motivated in biological terms, either in terms of introducing beneficial behaviours, creating a particularly attractive display, or as a demonstration of a valuable capacity that underlies creativity itself.
However, while the substrate of the variability required for biological evolution, as exemplified above, is genetic, the behaviours we are considering are complex, learned and cultural, involving not just generation of short sequences grounded in action, but substantial long-term abstract sequence production. The larger question is therefore: why and when is there selection for innovation—forming new combinations of behaviours, versus reliance on unlearned behaviours or social learning of successful behaviours? Laland [44] discusses strategies animals might use for selecting when to rely on unlearned behaviours, when and who to copy in social learning, and when to innovate. However, he is primarily considering instrumental behaviours for solving non-social problems rather than learning about signalling for communication. We now consider cases of animal communication that appear to be examples of creative behaviour in the terms proposed here, and then discuss how the dynamics of the communication might be quantitatively studied using a computational framework such that we propose.
5. Creativity in animal communication
The very attribution of the word ‘song’ to the vocal communication behaviours of birds and whales is based on the problematic romanticization of that phenomenon, akin to the romanticization of creativity, mentioned above. First, then, we must dissociate ourselves from the metaphorical notion of theatrical or concert-hall performance, and focus instead on the functional, communicative aspects of the behaviours.
A comparable danger is the naïve assumption that the behaviours described below are due to the same mechanisms as superficially similar behaviours in humans. Indeed, this claim is one we would like to test. One means of doing so might be through the observation-based model of Wallas [32]. However, it is hard to know whether the Preparation and Incubation phases exist in animals: they cannot be asked, and current lack of understanding of the human mechanisms at the neural level is not detailed enough to make search for comparable effects in animals possible: we are currently limited to measures such as EEG frequency band power, which do not explain mechanism (e.g. [45]). Illumination and Verification may be more accessible because they may manifest behaviourally, e.g. when an animal immediately repeats material once it has been internalized. More work is needed in this area.
However, there is evidence, cited above, of the effect of information content and entropy with respect to a context on humans, measurable directly from physiological responses [29], and of information content on birds [46]. A more direct comparison of these two phenomena can be made, using the tripartite empirical approach used by Pearce et al. [27]: a computer program is used to embody the proposed mechanism, and its predictions are then tested empirically with both behavioural responses and electrophysicological measures; here, the idea is extended to a comparison between species.
The setting in animal communication where the concept of creativity seems most relevant concerns reproductive advertisement displays called ‘songs', which are a product of sexual selection. In some animal species, the songs of each individual singer are learned through listening to the songs of other individuals. When one individual learns the song of another, it will probably not be a perfect copy, as there may be errors in the stored memory, and differences in the vocal production apparatus between individuals. This process of vocal copying within a community of animal singers leads to vocal traditions—which may be formalized as conceptual spaces—that often map onto habitats as geographical dialects in song; similar effects of vocal tradition, coupled with migratory patterns, arise in human folk music [47]. However, there are also situations when an animal actively appears to innovate, producing sounds that are more novel than would be expected to arise from copy errors alone, in a step akin to transformational creativity. Existing work in this area focuses on what is copied, at the expense of studying the ‘unrecognizable’ new material: the corollary, invention of new song types, seems not to have been studied formally.
Kroodsma [48] suggests a relationship between site fidelity and mode of vocal learning in birds, high site fidelity being correlated with imitation and low site fidelity correlated with improvisation. One possible explanation for this would be the need to associate a clear signal with territory, so as to mark it, and also for the signal to vary as fledglings leave the nest and lay out their own territorial boundaries: it is necessary first to innovate and then to fix, so as to identify a difference between the territories, while maintaining recognizability to members of one's own species for the purposes of sexual attraction. Similarly, in animals such as killer whales that form groups bonded by call repertoires, innovation is concomitant with the need to form new social groups as new individuals mature; otherwise, groups could not distinguish themselves. In both these cases, the ability to recognize and value ‘just enough’ variation is paramount. This notion of ‘just enough’ corresponds with the maximum of the Wundt curve, described above; it also corresponds with a middling, moderate value of information content, as measured by the models outlined in §3.
The development of song through vocal learning is common among songbirds but very rare among non-human mammals [49]. Some of the best evidence comes from the songs of bats [50] and humpback whales. At any one time, the songs of different individuals within a population of whales are quite similar [51], but many acoustic features of the songs change rapidly enough that they can be tracked from month to month [52]. The changes are progressive over time in the sense that if a sound is increasing in frequency, or decreasing in duration, that trend is likely to continue for some time rather than vary randomly. The rapidity of the song change coupled with the similarity between whales at one time makes it difficult to identify whether some individuals are innovators who are copied, especially as it is so difficult to make repeated recordings from the same individual at different times given such a large and mobile population. However, it is clear that a strong pressure for conformity must drive each whale to copy the song of the moment, while at the same time there must be a selection for specific innovations that are picked up by the population to change the vocal tradition. Until now, there has been no way of studying this process or identifying the benefits and process of innovation. Computational modelling based on creativity theory may help.
Following the Boden analysis of creativity, and our subsequent suggestion that value and novelty should be thought of as relations between observers and the created artefact, we can evaluate differences in the value of novel sounds produced by different singers in terms of whether they are copied by others or not. We know little about the psychological process by which an individual animal generates a novel sound nor about what ‘value’ the sound may offer to that individual. But in parallel with our explicitly relativistic version of Boden's ‘historical’ creativity, we can study what novel sounds are incorporated into the vocal tradition of the population, modelling the whale song as a conceptual space.
A striking case of adoption of novel songs involves the song of the humpback whale. There are two populations of humpback whales that winter off the coast of Australia: one on the east coast and one on the west. Males sing on their winter breeding grounds and as they migrate to and from the breeding grounds. These two populations are separated by thousands of kilometres. With little interchange, each population is characterized by one song at any one time, and the songs of the two populations are usually very different. However, Noad et al. [11] noticed an unusual pattern in 1996 when 2 of 82 singers recorded off the east coast produced a song that was completely different from the rest of this population but that matched the 1996 song of the west coast population.
During 1997, some songs mixed features of both west and east coast, but by the end of the year nearly all of the east coast whales had switched to singing west coast song. By 1998, no whales were left singing the old east coast song and all had switched to the west coast song. The rarity of west coast songs recorded in the east during 1996, coupled with the following independent evolution of the west coast song on both coasts, led Noad et al. [11] to conclude that only a few singers transferred from west to east during 1996, bringing the new vocal tradition with them. This rapid and complete replacement of one vocal tradition with another suggests recognition of a value for very specific kinds of novelty is what drives the change in the song, even when this is usually a less radical process driven from within the population.
Analysis of songs recorded during 1998–2008 from eastern Australia and the other populations of the South Pacific show a remarkable pattern. Garland et al. [53] report that over this time period, eight different song types originated in the eastern Australia population and spread over several years across six humpback populations from west to east, all the way to French Polynesia, 5000 km away. They suggest that as with the uptake of a new song as reported by Noad et al. [11], diffusion of a vocal tradition occurs when individual males from adjacent populations spend enough time together for one to learn the others' song [53]. However, this does not explain the directionality of information transfer. Available data on movement of individuals from one population to adjacent ones suggest that this is bidirectional with no bias to the east. The suggestion of Garland et al. for the remarkable directionality of the change is that the eastern Australia population is much larger than the others. While this may account for a more likely flow of animals from eastern Australia to the adjacent population to the east, it fails to account for the broader eastward pattern of information flow.
One way to think about this pattern from the current perspective would be to consider the value of particular innovations within the context of a particular vocal tradition at a particular time. The 1998 song in eastern Australia was the song originally from western Australia that was valued so highly that it swept through the population in 1997. At this point, this song started to evolve within the eastern Australia population, and at the same time, its high value made it likely to spread to populations to the east. Given the time this took and the speed at which song evolves within a population, the large eastern Australia population had an advantage in being more likely to offer high value changes within the shared vocal tradition, and these high value changes would maintain the directionality as they spread to other populations to the eastward. Once this dynamic was set up, if the easternmost populations were several years behind in the process of innovation and selection for value, then it was less likely for any innovations in this setting to spread west.
There has been growing interest in studying the strategies animals might adopt when they learn from others. Laland [44] points out that the costs and benefits of social learning depend on the context, and he suggests more attention be paid to strategies of when to copy and whom to copy. One when strategy suggests copying another singer when the copier's current behaviour is unproductive. From the perspective of a singer, this would suggest copying if you are not attracting females or if you are failing in competition with males. A whom strategy might be ‘copy the majority’, which would lead to conformist behaviour. Another whom strategy is ‘copy the most successful’: if singing whales can monitor the success of others, and if successful whales have variations in their song, this could drive a process of change, although it is difficult to see how it would lead to the progressive evolution observed most of the time in humpback song. We still do not understand what drives the conformity in humpback song, what drives the selection of specific novelties, and how or whether this is driven by sexual selection.
The movement and variation of whale song bears comparison with the movement of human music during migration. Pamjav et al. [47] conducted a large study of musical melody styles for 31 Eurasian nations. They found that close musical relations indicate close genetic relations (FST < 0.05 [54]) with probability 82%. This is one of the largest studies ever done of folk music, and almost certainly the largest computational study: they used databases of 1000–2500 melodies for each of the 31 cultures. The notion of musical similarity here revolves round a Euclidean distance metric derived from a Self-Organizing Map (SOM) [55], and this is an area requiring further validation: musical similarity is strongly context-dependent, and the workings of a SOM are somewhat inscrutable. Nevertheless, this work presents an interesting opportunity, given the models of music that we propose, to compare the whale song behaviour with the human musical behaviour in detailed and explicable ways.
6. A research programme on creativity in vocal communication in humans and non-humans
In this paper, we have identified parallels between human and animal vocal communication behaviours, at the immediate phenomenological level, and suggested that they are worthy of further investigation in the context of creativity research.
We decomposed the notion of creativity into an objective process of generation, coupled with a combination of relative value judgements, some of which, notably novelty, can be objectively modelled. This added objectivity allows us to ask questions that were not previously scientifically formulable, regarding the nature of vocal communication, its effect on humans and other species, and the mechanisms that underlie it.
We have deployed Boden's philosophical approach to human creativity [3] to hypothesize a possible explanation for new song construction in migrating whales, and identified evidence of music migration in humans. We have presented evidence, from normally separate research fields, of comparable physiological responses to aural sequence perception in birds and humans, which might suggest similar processes at deeper levels, suggesting a computational method by which these empirical studies can be implemented.
We propose, therefore, that when we examine the evolution of vocal communication in animals and humans from the perspective of creativity, we can shed new light on processes which seem to be common (though probably not commonly derived) between very distantly related species. Therefore, we suggest that the philosophical framework outlined here is a potentially fruitful means of addressing the communicative behaviour of animals that improvise (individually or collectively), and perhaps thence of understanding better the mechanisms that underlie human communication and human creativity.
More specifically, we can propose
— comparative studies on heart rate and other physiological and electrophysiological measures in birds and in humans in response to complex aural stimulation, relative to a known vocal communication form; subsequent neural studies to seek neural correlates of information content [27];
— comparative studies on the dynamics of whale migration and song variation as compared with the dynamics of human migration and song variation; subsequent modelling to compare the processes, novelty and complexity involved;
— the development of new measurement techniques to allow physiological and neural analysis of birds, whales and other improvising animals to be compared with human analysis, and thence modelled as we have described above.
We believe that these approaches and others entailed by questioning the relationship between creative behaviour in humans and the superficially similar behaviours in other species offer a new and exciting approach to understanding the cognitive mechanisms involved both in vocal communication and in creativity.
Acknowledgements
We gratefully acknowledge the advice and support of Björn Merker and two anonymous reviewers in assembling this paper. We gratefully acknowledge the support of the Lorentz Centre, Leiden, The Netherlands, whose workshop on Music and Evolution made this paper possible.
Funding statement
G.A.W. is funded by the Lrn2Cre8 and ConCreTe projects, which acknowledge the financial support of the Future and Emerging Technologies (FET) programme within the Seventh Framework Programme for Research of the European Commission, under FET grant numbers 610859 and 611733. P.L.T. acknowledges the support of the MASTS pooling initiative (The Marine Alliance for Science and Technology for Scotland). MASTS is funded by the Scottish Funding Council (grant reference HR09011) and contributing institutions. M.R. has been supported by the MIT Department of Linguistics and Philosophy as well as the Zukunftskonzept at TU Dresden funded by the Exzellenzinitiative of the Deutsche Forschungsgemeinschaft.
References
- 1.McCorduck P. 1991. AARON'S CODE: meta-art, artificial intelligence and the work of Harold Cohen. New York, NY: Freeman. [Google Scholar]
- 2.Cohen H. 1999. Colouring without seeing: a problem in machine creativity. AISB Q. 102, 26–35. See http://crca.ucsd.edu/hcohen/cohenpdf/colouringwithoutseeing.pdf. [Google Scholar]
- 3.Boden MA. 1990. The creative mind: myths and mechanisms. London, UK: Weidenfield and Nicholson. [Google Scholar]
- 4.Kuhn TS. 1962. The structure of scientific revolutions. Chicago, IL: University of Chicago Press. [Google Scholar]
- 5.Wiggins GA. 2006. A preliminary framework for description, analysis and comparison of creative systems. J. Knowledge Based Syst. 19, 449–458. ( 10.1016/j.knosys.2006.04.009) [DOI] [Google Scholar]
- 6.Frumhoff P. 1983. Aberrant songs of humpback whales (Megaptera novaeangliae): clues to the structure of humpback songs. In Communication and behavior of whales. AAAS Selected Symposia Series (ed. R Payne), pp. 81–127. Boulder, CO: Westview Press. [Google Scholar]
- 7.Plotkin H. 1998. Evolution in mind. Cambridge, MA: Harvard University Press. [Google Scholar]
- 8.Bown O, Wiggins GA. 2009. From maladaptation to competition to cooperation in the evolution of musical behaviour. Music. Sci. 13, 387–411. Special issue on Evolution of Music ( 10.1177/1029864909013002171) [DOI] [Google Scholar]
- 9.van den Broek EMF, Todd PM. 2009. Evolution of rhythm as an indicator of mate quality. Music. Sci. 13, 369–386. ( 10.1177/1029864909013002161) [DOI] [Google Scholar]
- 10.Miller GF. 1997. Protean primates: the evolution of adaptive unpredictability in competition and courtship. In Machiavellian intelligence II: extensions and evaluations, vol. xii (eds Whiten A, Byrne RW.), pp. 312–340. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 11.Noad MJ, Cato DH, Bryden MM, Jenner MN, Jenner KCS. 2000. Cultural revolution in whale songs. Nature 408, 537 ( 10.1038/35046199) [DOI] [PubMed] [Google Scholar]
- 12.Martindale C. 1990. The clockwork muse: the predictability of artistic change. New York, NY: Basic Books. [Google Scholar]
- 13.Saunders R. 2001. Curious design agents and artificial creativity. Sydney, Australia: The University of Sydney. [Google Scholar]
- 14.Wundt WM. 1874. Grundzüge der physiologischen Psychologie. Leipzig, Germany: W. Engelman. [Google Scholar]
- 15.Margulis EH, Beatty AP. 2008. Musical style, psychoaesthetics, and prospects for entropy as an analytic tool. Comput. Music J. 32, 64–78. ( 10.1162/comj.2008.32.4.64) [DOI] [Google Scholar]
- 16.Shettleworth SJ. 2010. Cognition, evolution, and behavior. New York, NY: Oxford University Press. [Google Scholar]
- 17.Huron D. 2006. Sweet anticipation: music and the psychology of expectation. Bradford books. Cambridge, MA: MIT Press. [Google Scholar]
- 18.Meyer LB. 1956. Emotion and meaning in music. Chicago, IL: University of Chicago Press. [Google Scholar]
- 19.Skowronek J, McKinney MF, van de Par S. 2006. Ground truth for automatic music mood classification. In Proceedings of ISMIR 2006 (eds Dannenberg R, Lemström K.). See http://ismir2006.ismir.net/PAPERS/ISMIR06105_Paper.pdf. [Google Scholar]
- 20.Juslin PN, Sloboda JA. 2010. Handbook of music and emotion: theory, research, applications. Affective science. Oxford, UK: Oxford University Press. [Google Scholar]
- 21.Manning CD, Schütze H. 1999. Foundations of statistical natural language processing. Cambridge, MA: MIT Press. [Google Scholar]
- 22.Conklin D, Witten IH. 1995. Multiple viewpoint systems for music prediction. J. New Music Res. 24, 51–73. ( 10.1080/09298219508570672) [DOI] [Google Scholar]
- 23.Pearce MT, Wiggins GA. 2012. Auditory expectation: the information dynamics of music perception and cognition. Top. Cogn. Sci. 4, 625–652. ( 10.1111/j.1756-8765.2012.01214.x) [DOI] [PubMed] [Google Scholar]
- 24.Cate CT, Okanoya K. 2012. Revisiting the syntactic abilities of non-human animals: natural vocalizations and artificial grammar learning. Phil. Trans. R. Soc. B 367, 1984–1994. ( 10.1098/rstb.2012.0055) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pearce MT. 2005. The construction and evaluation of statistical models of melodic structure in music perception and composition. London, UK: Department of Computing, City University, London. [Google Scholar]
- 26.Pearce MT, Wiggins GA. 2006. Expectation in melody: the influence of context and learning. Music Percept. 23, 377–405. ( 10.1525/mp.2006.23.5.377) [DOI] [Google Scholar]
- 27.Pearce MT, Herrojo Ruiz M, Kapasi S, Wiggins GA, Bhattacharya J. 2010. Unsupervised statistical learning underpins computational, behavioural and neural manifestations of musical expectation. NeuroImage 50, 303–314. ( 10.1016/j.neuroimage.2009.12.019) [DOI] [PubMed] [Google Scholar]
- 28.Shannon CE. 1948. A mathematical theory of communication. Bell Syst. Tech. J. 27, 379–423. ( 10.1002/j.1538-7305.1948.tb01338.x) [DOI] [Google Scholar]
- 29.Egermann H, Pearce M, Wiggins G, McAdams S. 2013. Probabilistic models of expectation violation predict psychophysiological emotional responses to live concert music. Cogn. Affect. Behav. Neurosci. 13, 533–553. ( 10.3758/s13415-013-0161-y) [DOI] [PubMed] [Google Scholar]
- 30.Ikebuchi M, Futamatsu M, Okanoya K. 2003. Sex differences in song perception by Bengalese finches as measured by cardiac response. Anim. Behav. 65, 123–130. ( 10.1006/anbe.2002.2012) [DOI] [Google Scholar]
- 31.Weiss M, Hultsch H, Adam I, Scharff C, Kipper S. 2014. The use of network analysis to study complex animal communication systems: a study on nightingale song. Proc. R. Soc. B 281, 20140460 ( 10.1098/rspb.2014.0460) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Wallas G. 1926. The art of thought. New York, NY: Harcourt Brace. [Google Scholar]
- 33.Wiggins GA. 2012. The mind's chorus: creativity before consciousness. Cogn. Comput. 4, 306–319. ( 10.1007/s12559-012-9151-6) [DOI] [Google Scholar]
- 34.Holmes E. 2009. The life of Mozart: including his correspondence. Cambridge Library Collection Cambridge, UK: Cambridge University Press. [Google Scholar]
- 35.Guilford JP. 1967. The nature of human intelligence. New York, NY: McGraw-Hill. [Google Scholar]
- 36.Csikszentmihalyi M. 1996. Creativity: flow and the psychology of discovery and invention. New York, NY: HarperCollins. [Google Scholar]
- 37.Koestler A. 1964. The act of creation. London, UK: Hutchinson & Co. [Google Scholar]
- 38.Wiggins GA, Forth JC. 2015. IDyOT: a computational theory of creativity as everyday reasoning from learned information. In Computational creativity research: towards creative machines. Atlantis thinking machines. Paris, France: Springer. [Google Scholar]
- 39.Baars BJ. 1988. A cognitive theory of consciousness. Cambridge, UK: Cambridge University Press. [Google Scholar]
- 40.Endler JA, Basolo AL. 1998. Sensory ecology, receiver biases and sexual selection. TREE 13, 415–420. ( 10.1016/S0169-5347(98)01471-2) [DOI] [PubMed] [Google Scholar]
- 41.Fisher RA. 1930. The genetical theory of natural selection. Oxford, UK: Clarendon Press. [Google Scholar]
- 42.Zahavi A. 1975. Mate selection—a selection for a handicap. J. Theor. Biol. 53, 205–213. ( 10.1016/0022-5193(75)90111-3) [DOI] [PubMed] [Google Scholar]
- 43.Nowicki S, Peters S, Podos J. 1998. Song learning, early nutrition, and sexual selection in songbirds. Am. Zool. 38, 179–190. [Google Scholar]
- 44.Laland KN. 2004. Social learning strategies. Learn. Behav. 32, 4–14. ( 10.3758/BF03196002) [DOI] [PubMed] [Google Scholar]
- 45.Schwab D, Benedek M, Papousek I, Weiss EM, Fink A. 2014. The time-course of EEG alpha power changes in creative ideation. Front. Hum. Neurosci. 8, 310 ( 10.3389/fnhum.2014.00310) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Okanoya K. 2012. Behavioural factors governing song complexity in Bengalese finches. Int. J. Comp. Psychol. 25, 44–59. [Google Scholar]
- 47.Pamjav H, Juhász Z, Zalán A, Németh E, Damdin B. 2012. A comparative phylogenetic study of genetics and folk music. Mol. Genet. Genom. 287, 337–349. ( 10.1007/s00438-012-0683-y) [DOI] [PubMed] [Google Scholar]
- 48.Kroodsma DE. 1983. Acoustic communication in birds. New York, NY: Academic Press. [Google Scholar]
- 49.Janik V, Slater PJB. 1997. Vocal learning in mammals. Adv. Study Behav. 26, 59–99. ( 10.1016/S0065-3454(08)60377-0) [DOI] [Google Scholar]
- 50.Morell V. 2014. When the bat sings. Science 344, 1334–1337. ( 10.1126/science.344.6190.1334) [DOI] [PubMed] [Google Scholar]
- 51.Guinee LN, Chu K, Dorsey EM. 1983. Changes over time in the songs of known individual humpback whales (Megaptera novaeangliae). In Communication and behavior of whales. AAAS selected symposia series (ed. Payne R.), pp. 59–80. Boulder, CO: Westview Press. [Google Scholar]
- 52.Payne K, Tyack P, Payne R. 1983. Progressive changes in the songs of humpback whales (Megaptera novaeangliae): a detailed analysis of two seasons in Hawaii. In Communication and behavior of whales. AAAS selected symposia series (ed. Payne R.), pp. 9–57. Boulder, CO: Westview Press. [Google Scholar]
- 53.Garland EC, et al. 2011. Dynamic horizontal cultural transmission of humpback whale song at the ocean basin scale. Curr. Biol. 21, 687–691. ( 10.1016/j.cub.2011.03.019) [DOI] [PubMed] [Google Scholar]
- 54.Wright S. 1969. Evolution and the genetics of populations, vol. 2 Chicago, IL: University of Chicago Press. [Google Scholar]
- 55.Kohonen T. 1995. Self-organizing maps, vol. 30. Information Sciences, 2nd edn Heidelberg, Germany: Springer. [Google Scholar]