

Abstract
AIM: To investigate the effects of troglitazone (TGZ), an anti-diabetic drug which activates peroxisome proliferator-activated receptor-γ (PPAR-γ), for liver tissue repair, and the development of ductular reaction, following common bile duct ligation (BDL) in rats.
METHODS: Rats were supplemented with TGZ (0.2% w/w in the pelleted food) for 1 wk before BDL or sham operation. Animals were killed at 1, 2, or 4 wk after surgery.
RESULTS: The development of liver fibrosis was reduced in rats receiving TGZ, as indicated by significant decreases of procollagen type I gene expression and liver hydroxy-proline levels. Accumulation of α-smooth-muscle actin (SMA)-expressing cells surrounding newly formed bile ducts following BDL, as well as total hepatic levels of SMA were partially inhibited by TGZ treatment, indicating the presence of a reduced number and/or activation of hepatic stellate cells (HSC) and myofibroblasts. Development of the ductular reaction was inhibited by TGZ, as indicated by histochemical evaluation and hepatic activity of γ-glutamyl-transferase (GGT).
CONCLUSION: Treatment with thiazolidinedione reduces ductular proliferation and fibrosis in a model of chronic cholestasis, and suggests that limiting cholangiocyte proliferation may contribute to the lower development of scarring in this system.
Keywords: Cholangiocytes, Ductular reaction, PPAR-γ, Hepatic stellate cells, Myofibroblasts, Troglitazone
INTRODUCTION
A number of conditions characterized by chronic liver damage are associated with the development of liver fibrosis, ultimately leading to cirrhosis and functional alterations[1]. Although fibrotic progression represents the final outcome of the liver ‘wound healing’ response in several conditions of chronic injury, different types of fibrosis show relatively unique features. Biliary fibroses, such as the one observed in conditions of chronic obstruction of the biliary tree, are characterized by the accumulation of extracellular matrix at portal tracts, surrounding newly formed bile ducts emerging during the course of the so-called ‘typical’ ductular reaction and originated from proliferation of pre-existing bile duct epithelial cells[2,3]. If the obstruction persists, fibrosis bridging different portal tracts develops, ultimately leading to biliary cirrhosis. Common bile duct ligation (BDL) and scission in rats represents a classical experimental model for the analysis of the events leading the initial injury of the bile ducts to the organization of a fibrotic reaction, which involves different matrix-producing cells of mesenchymal origin, including α-smooth muscle actin (SMA)-positive portal myofibroblasts and activated hepatic stellate cells (HSC)[2,4-6]. This model has been widely used to investigate the molecular and cellular mechanisms of liver fibrosis and to assess potential treatments for this disorder. In addition, due to the close anatomical and functional contacts between cholangiocytes and matrix-producing cells, the BDL model represents an optimal tool for the study of epithelial-mesenchymal interactions.
Peroxisome proliferator-activated receptors (PPARs) belong to the nuclear hormone receptor superfamily[7]. All these molecules share the ability to function as transcription factors by binding to target DNA elements, and change their transcriptional activity after binding to specific ligands. Three different PPAR isoforms have been cloned and characterized. PPAR-α binds to fatty acids and fibrates and is known to induce transcription of genes involved in ω- and β-oxidation of fatty acids[7]. PPAR-β (or -δ) is expressed ubiquitously, and could be involved in fatty acid and lipid metabolism[8]. Finally, the ligands for PPAR-γ include anti-diabetic drugs of the thiazolidinedione group and arachidonic acid metabolites, such as prostaglandins of the D series and its derivatives such as 15deoxy-△12,14PGJ2. PPAR-γ is expressed at high levels in the adipose tissue, where it regulates lipid storage and modulates the action of insulin[9-11].
Agonists of PPAR-γ have been shown to regulate multiple and critical functions in diverse pathophysiologic conditions, including type 2 diabetes and the metabolic syndrome, cancer, and inflammation[12]. In the liver, considerable attention has been directed to the modulation of the development of fibrosis through actions on HSC and liver myofibroblasts[13-15]. In this study, we investigated the effects of treatment with troglitazone (TGZ), the first thiazolidinedione introduced in clinical practice, on the development of ductular reaction and fibrosis in the BDL model. The results indicate that TGZ not only limits extracellular matrix deposition and HSC activation, but also inhibits the formation of newly formed bile ducts, thus acting on both epithelial and mesenchymal cells involved in the response to this type of chronic liver injury.
MATERIALS AND METHODS
In vivo model
Male Wistar rats (Harlan-Nossan, Correnzana, Italy), weighing 180-200 g, were used according to the national and local ethical guidelines. For experimental purposes, animals were divided into two experimental groups and fed for 2 wk either with a standard pelleted diet (rat diet No. 48, Piccioni, Gessate Milanese, Italy) and water ad libitum (control diet) or with the same pelleted diet containing 0.2% (w/w) troglitazone (TGZ diet), according to previously described procedures[16,17]. In the animals from both experimental groups, permanent extrahepatic cholestasis was induced by common BDL and scission under light ether anesthesia as previously described[18,19]. Animals receiving sham operation were subjected only to the manipulation of the common bile duct, and served as controls. After BDL or sham operation, the animals continued to receive the same diet according to the following experimental groups: (1) Sham (control, receiving normal diet), (2) TGZ (sham, receiving TGZ-supplemented diet), (3) BDL (operated, receiving normal diet), and (4) BDL/TGZ (operated, receiving TGZ-supplemented diet). The amount of consumed food in each experimental group was recorded throughout the study. The animals were then killed after 1, 2, and 4 wk of surgery (n = 6-8 animals for each experimental group at any time point). The liver was promptly removed, and portions of the organ were either immediately used or frozen in liquid nitrogen and stored at -80 °C for further determinations or processed for histological and histochemical examination. Blood samples were also collected from individual animals before they were killed.
Serum and liver biochemistry
Serum activities of glutamic oxalacetic transaminase (GOT) and alkaline phosphatase (AP) were determined using a commercial kit (Sigma Diagnostics, Sigma, Milan, Italy). L-Hydroxyproline concentration in liver samples was determined according to standard procedures[20]. γ-Glutamyltransferase (GGT) activity was evaluated on diluted liver homogenate samples using a modified standard procedure[21] as previously described[22].
RNase protection assay
Total RNA was isolated from frozen liver tissues using Nucleospin columns (Mackerey-Nagel, Dürer, Germany). Integrity of RNA was checked by agarose electrophoresis. 32P-labeled cRNA probes were transcribed from templates encoding for rat α1(I) procollagen (kindly provided by Dr. Jackie J. Maher, University of California at San Francisco) and housekeeping gene 36B4. After hybridization, protected fragments were separated on a sequencing gel and autoradiographed. All procedures for probe labeling, hybridization, and digestion have been described in detail elsewhere[23].
Western blotting
Liver tissue was homogenized in an Ultra-Turrax homogenizer in RIPA buffer containing 1% Nonidet P40 and protease inhibitors. Insoluble proteins were discarded by high-speed centrifugation at 4 °C. The protein concentration in the supernatant was measured using a commercially available kit (Pierce, Rockford, IL, USA). Fifty micrograms of protein was separated by 10% SDS-PAGE, and electroblotted on a PVDF membrane. After transfer, the membrane was stained with a Ponceau red solution (Sigma) to ensure an equal protein loading. The staining was then removed by washing in PBS-Tween and the membrane was blocked with 3% BSA in PBS-Tween overnight. After an additional washing, the blots were sequentially incubated with monoclonal anti-α-SMA antibodies and anti-mouse horseradish peroxidase-conjugated antibodies. Detection was performed by chemiluminescence, according to the manufacturer’s protocol (Amersham, Arlington Heights, IL, USA).
Immunohistochemistry
Experiments were conducted on frozen sections as described in detail elsewhere[24]. Dried sections were sequentially incubated with the primary antibody followed by affinity-purified rabbit anti-mouse antibodies after washing. At the end of incubation, sections were washed twice in TBS, incubated with APAAP and developed. Negative controls were treated with omission of the primary antibody or its substitution with non-immune rabbit immunoglobulins.
Histology and histochemistry
Frozen liver specimens or specimens fixed in 4% formaldehyde in phosphate buffer (pH 7.2) and embedded in paraffin were used for morphological analysis. Standard liver sections (4-6-μm thick) embedded in paraffin were stained with hematoxylin and eosin. Biliary epithelial cells of the newly formed ductular structures were identified by histochemical staining for GGT activity as previously described[18].
RESULTS
In vivo administration of TGZ-supplemented diet to control animals (TGZ group) did not significantly affect either the liver morphology or the serum chemistry (Table 1 and Figures 1A and B). Following BDL, an increase in serum transaminase and AP was observed in all animals (BDL group, Table 1).
Table 1.
Serum biochemical parameters, body, and liver weight in the different experimental groups (mean±SD)
| Sham | TGZ | BDL | BDL/TGZ | |
| ALT (U/L) | ||||
| 1 wk | 35±5 | 22±2 | 90±19b | 81±7b |
| 4 wk | 23±7 | 21±5 | 102±34b | 75±12b |
| ALP (U/L) | ||||
| 1 wk | 83±5 | 76±12 | 176±19b | 168±22b |
| 4 wk | 88±7 | 82±9 | 137±14b | 142±27b |
| Body weight (g) | 332±7 | 319±9 | 293±5b | 337±12a |
| Liver weight (g) | 9.7±0.6 | 9.9±0.7 | 24.4±2.2b | 20.2±1.9b |
P<0.05 vs BDL,
P<0.01 vs Sham.
Figure 1.
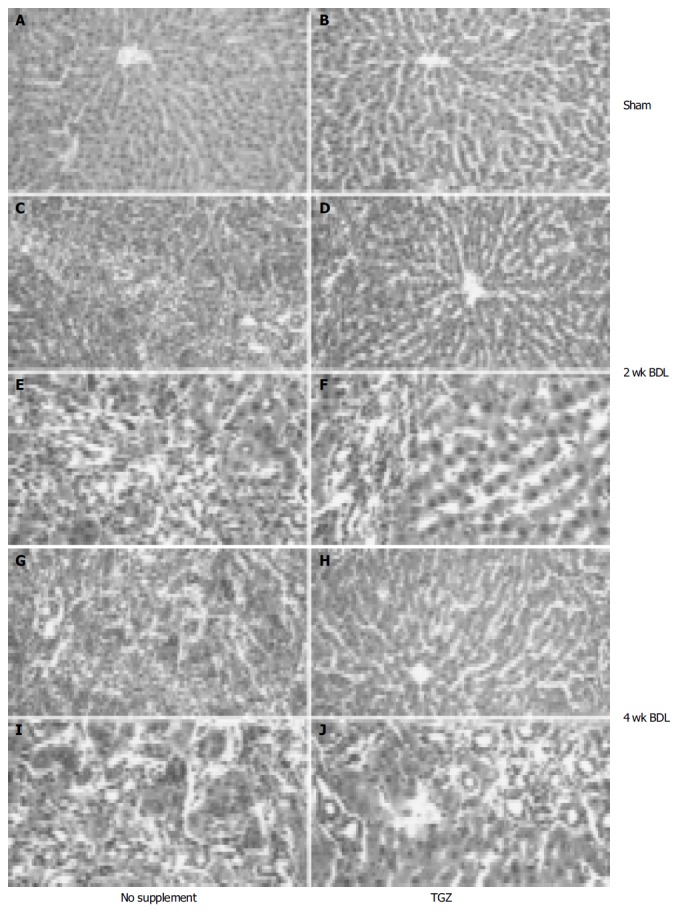
Effects of oral supplementation with TGZ on liver pathology in control rats (A and B) and bile duct-ligated rats (C-J). Animals received sham operation (A and B) or were subjected to BDL for 2 wk (C-F) or for 4 wk (G-J).
Comparison of the BDL and BDL/TG groups showed no significant differences in serum enzyme levels or in liver weight, while body weight was significantly lower in BDL animals (Table 1). No difference in food consumption was found between the two groups undergoing BDL. Histological examination of rats undergoing BDL revealed bile duct proliferation in the portal tract (Figure 1), already evident after 1 wk and progressively worsening thereafter. This ‘typical’ ductular reaction was associated with progressive deposition of extracellular matrix surrounding the newly formed bile ducts. After 4 wk, alteration of liver structure was evident, with bridging fibrosis encircling most of the newly formed bile ducts. These histological alterations were considerably less evident in the group of BDL/TGZ rats (Figure 1). Thus, at 2 wk, only moderate fibrosis was present, and the area occupied by proliferating bile ducts was more limited than in rats not undergoing TGZ supplementation (Figures 1C-F). Similarly, after 4 wk, less fibrosis and a more limited disarray of liver structure were observed in BDL/TGZ animals (Figures 1G-J).
To establish whether these morphologic changes were associated with reduced fibrogenesis, gene expression levels of procollagen I were measured at different time points. In all animals undergoing BDL, a marked increase in procollagen I gene expression was observed, as indicated by RNase protection assay (Figure 2). In BDL/TGZ rats, the increase in procollagen expression was less marked at 1st and 2nd wk after surgery (Figure 2). Interestingly, the inhibitory effects on procollagen type I expression were no longer evident 4 wk after BDL. We next evaluated whether reduced fibrosis and procollagen expression were associated with modified collagen levels, by measuring hepatic hydroxyproline levels. As expected, a progressive increase in hepatic hydroxyproline was evident in animals undergoing BDL, with a peak at 4 wk, when mean values were approximately five times higher than in controls (Figure 3). Administration of TGZ resulted in a marked inhibition of hydroxyproline accumulation after BDL. At 4 wk, hepatic hydroxyproline levels were inhibited more than 50% in BDL/TGZ rats, being comparable to those measured after 2 wk in BDL animals not receiving the drug. These data indicate that TGZ could effectively reduce hepatic fibrosis caused by BDL.
Figure 2.
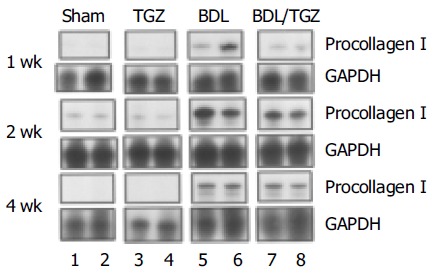
Effects of TGZ on type I procollagen expression in bile duct-ligated rats.
Figure 3.
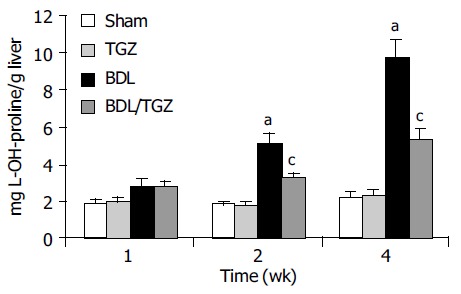
TGZ inhibits collagen accumulation in bile duct-ligated rats in different experimental groups. aP<0.05 vs Sham, cP<0.05 vs BDL.
In vitro studies indicated that thiazolidinediones could inhibit HSC activation and related phenotypic responses, including proliferation and migration. We analyzed the expression of SMA as an index of the number and activation state of HSC and other fibrogenic mesenchymal cells in the different groups of animals.
Immunohistochemical analysis showed that there was a progressive accumulation of SMA-expressing cells around the expanded portal tracts, in close proximity to bile duct epithelial cells of the ductular reaction. At 4 wk after BDL, SMA-positive cells occupied a significant portion of the hepatic lobule, being a part of the porto-portal septae (Figure 4). This picture was clearly modified in BDL/TGZ rats, where a less marked abundance of cells expressing SMA was present. To substantiate and quantify the inhibitory effect of TGZ on activated matrix-producing cells, SMA expression in the whole liver was analyzed by Western blotting. The progressive increase in SMA expression observed in the BDL group (up to fivefold at 4 wk) was markedly reduced by TGZ treatment (Figure 5). Taken together, these data indicate that TGZ treatment was associated with a reduction in the number of matrix-producing cells accumulated in close contact with cells of the ductular reaction.
Figure 4.
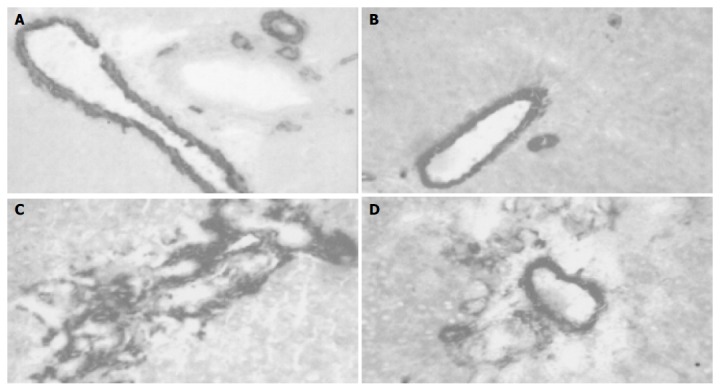
TGZ reduces accumulation of SMA-positive cells after BDL in different experimental groups. A: Sham; B: TGZ; C: BDL; D: BDL/TGZ.
Figure 5.
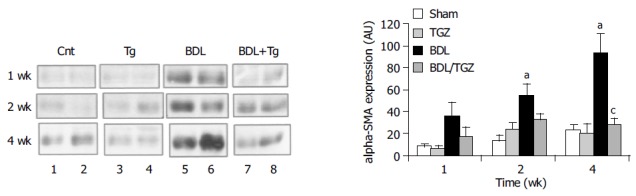
Effects of TGZ on SMA expression in rats undergoing BDL. aP<0.05 vs Sham, cP<0.05 vs BDL.
Morphological analysis of tissue samples from BDL/TGZ animals suggested a reduction of the extent of the ductular reaction in this group. We estimated the number of bile duct epithelial cells in the different groups of animals by performing a histochemical staining for GGT, which allowed to identify this cell type (Figure 6). In BDL rats, a progressive accumulation of newly formed bile ducts was evident, reaching a maximum after 4 wk. At all time points after BDL, TGZ treatment reduced the number of GGT-expressing cells, indicating a marked inhibition on the development of the ductular reaction (Figure 6). To quantify this effect, the enzymatic GGT activity was measured in total liver tissue lysates obtained from control and BDL rats (Figure 7). We observed a progressive increase in GGT activity in the BDL group, which was in parallel to the accumulation of newly formed bile ducts. A significant reduction of GGT activity was present in rats treated with TGZ at 2nd and 4th wk after BDL, confirming the inhibitory action shown by histochemical analysis. Thus, treatment with TGZ was also associated with a reduction in the severity of the ductular reaction caused by BDL.
Figure 6.
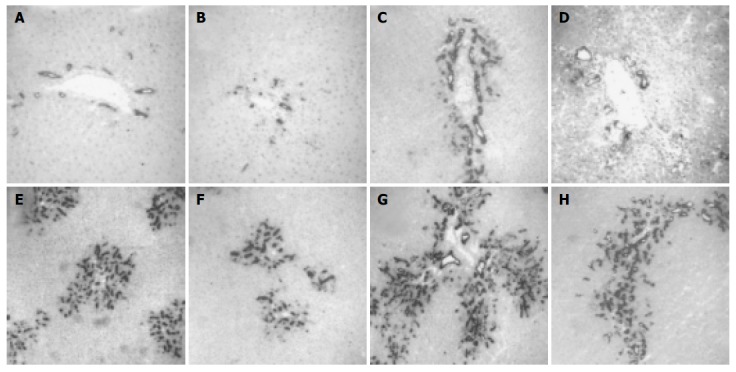
Development of ductular reaction is inhibited in bile duct ligated and TGZ-treated rats in different experimental groups. (A: Sham; B: TGZ; C, E and G: BDL; D, F, and H: BDL/TGZ) for different periods of time (C and D: 1 wk; E and F: 2 wk; A, B, G, and H: 4 wk).
Figure 7.
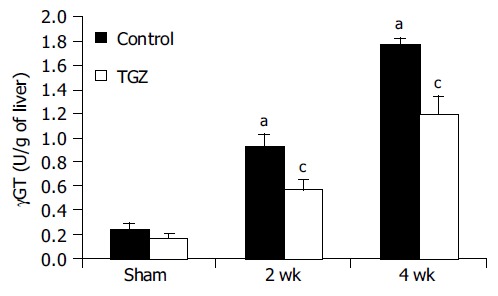
Effects of TGZ on liver GGT activity in bile duct-ligated rats. aP<0.05 vs Sham, cP<0.05 vs BDL.
DISCUSSION
Biliary fibrosis is a reproducible and reliable model for the study of experimental liver fibrogenesis[25]. Long-standing cholestasis leads to an ordered bile duct proliferation, or typical ductular reaction[3], associated with progressive accumulation of extracellular matrix, starting from the portal tracts and extending to the other areas of the lobule. This model is characterized by low-grade inflammation, activation of HSC and other mesenchymal cells, including portal myofibroblasts participating in the deposition of extracellular matrix components and contributing to the development of fibrosis[5,6,26]. In this study, we have provided evidence that TGZ, which activates the nuclear hormone receptor PPAR-γ, reduces the development of fibrosis and the extent of bile duct proliferation following BDL in rats. In the past 3 years, the potential role of PPAR-γ agonists as negative modulators of liver fibrosis has received considerable attention. We and others have shown that thiazolidinediones and 15deoxy-△12,14PGJ2 inhibit HSC activation and downregulate several biologic actions of activated HSC, which are important for liver wound healing and fibrogenesis, including cell proliferation, migration, extracellular matrix synthesis, and expression of chemokines[13-15]. 15deoxy-△12,14PGJ2 also induces apoptosis and reduces matrix expression in cultured liver myofibroblasts, which contribute to extracellular matrix production during injury, although this effect seems to be mediated by PPAR-γ-independent mechanisms[27]. These mechanisms highlighted in in vitro studies are likely to contribute to the observed inhibition of the development of liver fibrosis in this in vivo model. Moreover, data from the present study are in agreement with recently reported findings, demonstrating that other thiazolidinediones inhibit collagen deposition in several models of chronic liver damage, including chronic intoxication with carbon tetrachloride or dimethylnitrosamine, or BDL[28,29]. In our study, collagen accumulation was evaluated using liver hydroxyproline content, which is considered as the ‘gold standard’ for measuring matrix deposition. We used TGZ, the prototype of thiazolidinediones, as a PPAR-γ agonist. Although TGZ has been withdrawn from the marked increase because of its severe hepatotoxicity[30], it is still currently used in animal models or in cell culture systems, to establish a ‘proof of concept’ that may be expanded to other drugs of the same class, such as rosiglitazone or pioglitazone[31,32]. Thiazolidinediones also exert a protective role in other conditions of liver injury, as indicated by the recent findings, showing amelioration of ethanol- or diet-induced liver injury by pioglitazone[33-35].
The most novel and relevant finding of the present work is the observation that treatment with TGZ inhibits bile duct proliferation or typical ductular reaction in this model and in some cases of chronic cholestasis in humans. At the level of portal tract, newly formed bile ducts progressively invade the other parts of the lobule and are surrounded by fibrous tissue. TGZ reduces formation of bile ducts as indicated by histochemical analysis of GGT that is known to be selectively expressed by bile duct cells[36]. In addition, analysis of the enzymatic activity of GGT in total tissue homogenates has confirmed that this observation is not due to focal changes, but is the result of a general inhibitory action exerted by thiazolidinedione. The reported inhibitory effect on bile duct proliferation is indirectly in agreement with the report of Han et al[37]. Although these data may not be directly extrapolated to normal cells, they are consistent with the involvement of the PPAR-γ system in growth control of cholangiocytes. Further studies are needed to establish if the molecular mechanisms mediating the effects of TGZ on cholangiocarcinoma cells, such as elevated expression of p53, also operate in non-transformed cholangiocytes.
The intriguing findings on bile duct proliferation provide further insight on the mechanisms of reduced fibrogenesis and epithelial-mesenchymal interactions in this model. Bile duct epithelial cells express soluble mediators affecting the biology of matrix-producing cells. We have previously reported that in the BDL model, cholangiocytes express platelet-derived growth factor (PDGF)[19], which may contribute to local accumulation of matrix-producing cells in proximity to newly formed bile ducts, as the result of chemotaxis and proliferation[26,38-40]. There is evidence that PDGF stimulates the conversion of peri-biliary cells to matrix-producing myofibroblasts, and that an inhibitor of PDGF receptor’s tyrosine kinase activity reduces fibrosis in BDL rats[6]. In addition, bile duct segments isolated from cholestatic rats exert a potent chemotactic action on HSC[38]. Along these lines, secretion of endothelin-1 by cholangiocytes may induce proliferation and contraction of HSC, and inhibition of endothelin’s activity is associated with reversal of fibrosis following BDL[41-43]. Chemokines produced by cholangiocytes, such as monocyte chemoattractant protein-1 and interleukin-8[44] are responsible for the recruitment of inflammatory cells that may amplify the fibrogenic stimulus through secretion of additional mediators. Moreover, monocyte chemoattractant protein-1 is chemotactic for activated HSC, providing an additional mechanism leading to local accumulation of fibrogenic cells[45]. On the other hand, it is also possible that the inhibitory effects of TGZ on HSC and myofibroblasts lead to reduced secretion of mitogenic factors for cholangiocytes. Indeed, liver pro-fibrogenic cells have been shown to secrete cytokines such as interleukin-6 and hepatocyte growth factor modulating cholangiocyte survival and growth[46,47]. Thus, a bi-directional modulation of the biology of epithelial and mesenchymal cells by soluble factors is likely to occur in this model system.
In conclusion, TGZ can affect not only the development of fibrosis through direct action on matrix-producing cells, but also may modulate the epithelial-mesenchymal interactions characterizing chronic obstructive cholestasis.
ACKNOWLEDGMENTS
The authors thank Wanda Delogu and Nadia Navari for excellent technical assistance, Dr. Jackie J. Maher for kindly providing the plasmid for RNase protection assay of rat procollagen, Dr. Hidekuni Takahagi (Sankyo Co., Tokyo, Japan) for the kind gift of TGZ, and Dr. Stefano Milani for his help with immunohistology.
Footnotes
Supported by the Italian MIUR Grant, No. MM_06315722, by the University of Florence and by the Italian Liver Foundation. Eva Efsen was Supported in Part by the Tode Travel Grant, the Direktør Madsen’s Grant and Fhv. Direktør Nielsen’s Grant (Denmark)
Science Editor Wang XL and Guo SY Language Editor Elsevier HK
References
- 1.Friedman SL. Liver fibrosis -- from bench to bedside. J Hepatol. 2003;38 Suppl 1:S38–S53. doi: 10.1016/s0168-8278(02)00429-4. [DOI] [PubMed] [Google Scholar]
- 2.Cassiman D, Roskams T. Beauty is in the eye of the beholder: emerging concepts and pitfalls in hepatic stellate cell research. J Hepatol. 2002;37:527–535. doi: 10.1016/s0168-8278(02)00263-5. [DOI] [PubMed] [Google Scholar]
- 3.Desmet V, Roskams T, Van Eyken P. Ductular reaction in the liver. Pathol Res Pract. 1995;191:513–524. doi: 10.1016/s0344-0338(11)80870-8. [DOI] [PubMed] [Google Scholar]
- 4.Knittel T, Kobold D, Piscaglia F, Saile B, Neubauer K, Mehde M, Timpl R, Ramadori G. Localization of liver myofibroblasts and hepatic stellate cells in normal and diseased rat livers: distinct roles of (myo-)fibroblast subpopulations in hepatic tissue repair. Histochem Cell Biol. 1999;112:387–401. doi: 10.1007/s004180050421. [DOI] [PubMed] [Google Scholar]
- 5.Cassiman D, Libbrecht L, Desmet V, Denef C, Roskams T. Hepatic stellate cell/myofibroblast subpopulations in fibrotic human and rat livers. J Hepatol. 2002;36:200–209. doi: 10.1016/s0168-8278(01)00260-4. [DOI] [PubMed] [Google Scholar]
- 6.Kinnman N, Francoz C, Barbu V, Wendum D, Rey C, Hultcrantz R, Poupon R, Housset C. The myofibroblastic conversion of peribiliary fibrogenic cells distinct from hepatic stellate cells is stimulated by platelet-derived growth factor during liver fibrogenesis. Lab Invest. 2003;83:163–173. doi: 10.1097/01.lab.0000054178.01162.e4. [DOI] [PubMed] [Google Scholar]
- 7.Shearer BG, Hoekstra WJ. Recent advances in peroxisome proliferator-activated receptor science. Curr Med Chem. 2003;10:267–280. doi: 10.2174/0929867033368295. [DOI] [PubMed] [Google Scholar]
- 8.Gilde AJ, Van Bilsen M. Peroxisome proliferator-activated receptors (PPARS): regulators of gene expression in heart and skeletal muscle. Acta Physiol Scand. 2003;178:425–434. doi: 10.1046/j.1365-201X.2003.01161.x. [DOI] [PubMed] [Google Scholar]
- 9.Kliewer SA, Lenhard JM, Willson TM, Patel I, Morris DC, Lehmann JM. A prostaglandin J2 metabolite binds peroxisome proliferator-activated receptor gamma and promotes adipocyte differentiation. Cell. 1995;83:813–819. doi: 10.1016/0092-8674(95)90194-9. [DOI] [PubMed] [Google Scholar]
- 10.Forman BM, Tontonoz P, Chen J, Brun RP, Spiegelman BM, Evans RM. 15-Deoxy-delta 12, 14-prostaglandin J2 is a ligand for the adipocyte determination factor PPAR gamma. Cell. 1995;83:803–812. doi: 10.1016/0092-8674(95)90193-0. [DOI] [PubMed] [Google Scholar]
- 11.Nagy L, Tontonoz P, Alvarez JG, Chen H, Evans RM. Oxidized LDL regulates macrophage gene expression through ligand activation of PPARgamma. Cell. 1998;93:229–240. doi: 10.1016/s0092-8674(00)81574-3. [DOI] [PubMed] [Google Scholar]
- 12.Murphy GJ, Holder JC. PPAR-gamma agonists: therapeutic role in diabetes, inflammation and cancer. Trends Pharmacol Sci. 2000;21:469–474. doi: 10.1016/s0165-6147(00)01559-5. [DOI] [PubMed] [Google Scholar]
- 13.Galli A, Crabb D, Price D, Ceni E, Salzano R, Surrenti C, Casini A. Peroxisome proliferator-activated receptor gamma transcriptional regulation is involved in platelet-derived growth factor-induced proliferation of human hepatic stellate cells. Hepatology. 2000;31:101–108. doi: 10.1002/hep.510310117. [DOI] [PubMed] [Google Scholar]
- 14.Marra F, Efsen E, Romanelli RG, Caligiuri A, Pastacaldi S, Batignani G, Bonacchi A, Caporale R, Laffi G, Pinzani M, et al. Ligands of peroxisome proliferator-activated receptor gamma modulate profibrogenic and proinflammatory actions in hepatic stellate cells. Gastroenterology. 2000;119:466–478. doi: 10.1053/gast.2000.9365. [DOI] [PubMed] [Google Scholar]
- 15.Miyahara T, Schrum L, Rippe R, Xiong S, Yee HF, Motomura K, Anania FA, Willson TM, Tsukamoto H. Peroxisome proliferator-activated receptors and hepatic stellate cell activation. J Biol Chem. 2000;275:35715–35722. doi: 10.1074/jbc.M006577200. [DOI] [PubMed] [Google Scholar]
- 16.Miles PD, Higo K, Romeo OM, Lee MK, Rafaat K, Olefsky JM. Troglitazone prevents hyperglycemia-induced but not glucosamine-induced insulin resistance. Diabetes. 1998;47:395–400. doi: 10.2337/diabetes.47.3.395. [DOI] [PubMed] [Google Scholar]
- 17.Mizushige K, Noma T, Yao L, Yu Y, Kiyomoto H, Hosomi N, Fukui T, Kimura S, Abe Y, Matsuo H. Effects of troglitazone on collagen accumulation and distensibility of aortic wall in prestage of non-insulin-dependent diabetes mellitus of Otsuka Long-Evans Tokushima Fatty rats. J Cardiovasc Pharmacol. 2000;35:150–155. doi: 10.1097/00005344-200001000-00020. [DOI] [PubMed] [Google Scholar]
- 18.Parola M, Leonarduzzi G, Robino G, Albano E, Poli G, Dianzani MU. On the role of lipid peroxidation in the pathogenesis of liver damage induced by long-standing cholestasis. Free Radic Biol Med. 1996;20:351–359. doi: 10.1016/0891-5849(96)02055-2. [DOI] [PubMed] [Google Scholar]
- 19.Grappone C, Pinzani M, Parola M, Pellegrini G, Caligiuri A, DeFranco R, Marra F, Herbst H, Alpini G, Milani S. Expression of platelet-derived growth factor in newly formed cholangiocytes during experimental biliary fibrosis in rats. J Hepatol. 1999;31:100–109. doi: 10.1016/s0168-8278(99)80169-x. [DOI] [PubMed] [Google Scholar]
- 20.Boigk G, Stroedter L, Herbst H, Waldschmidt J, Riecken EO, Schuppan D. Silymarin retards collagen accumulation in early and advanced biliary fibrosis secondary to complete bile duct obliteration in rats. Hepatology. 1997;26:643–649. doi: 10.1002/hep.510260316. [DOI] [PubMed] [Google Scholar]
- 21.Persijn JP, van der Slik W. A new method for the determination of gamma-glutamyltransferase in serum. J Clin Chem Clin Biochem. 1976;14:421–427. doi: 10.1515/cclm.1976.14.1-12.421. [DOI] [PubMed] [Google Scholar]
- 22.Parola M, Cheeseman KH, Biocca ME, Dianzani MU, Slater TF. Biochemical studies on bile duct epithelial cells isolated from rat liver. J Hepatol. 1990;10:341–345. doi: 10.1016/0168-8278(90)90143-f. [DOI] [PubMed] [Google Scholar]
- 23.Marra F, Choudhury GG, Pinzani M, Abboud HE. Regulation of platelet-derived growth factor secretion and gene expression in human liver fat-storing cells. Gastroenterology. 1994;107:1110–1117. doi: 10.1016/0016-5085(94)90236-4. [DOI] [PubMed] [Google Scholar]
- 24.Marra F, DeFranco R, Grappone C, Milani S, Pastacaldi S, Pinzani M, Romanelli RG, Laffi G, Gentilini P. Increased expression of monocyte chemotactic protein-1 during active hepatic fibrogenesis: correlation with monocyte infiltration. Am J Pathol. 1998;152:423–430. [PMC free article] [PubMed] [Google Scholar]
- 25.Kountouras J, Billing BH, Scheuer PJ. Prolonged bile duct obstruction: a new experimental model for cirrhosis in the rat. Br J Exp Pathol. 1984;65:305–311. [PMC free article] [PubMed] [Google Scholar]
- 26.Kinnman N, Goria O, Wendum D, Gendron MC, Rey C, Poupon R, Housset C. Hepatic stellate cell proliferation is an early platelet-derived growth factor-mediated cellular event in rat cholestatic liver injury. Lab Invest. 2001;81:1709–1716. doi: 10.1038/labinvest.3780384. [DOI] [PubMed] [Google Scholar]
- 27.Li L, Tao J, Davaille J, Feral C, Mallat A, Rieusset J, Vidal H, Lotersztajn S. 15-deoxy-Delta 12,14-prostaglandin J2 induces apoptosis of human hepatic myofibroblasts. A pathway involving oxidative stress independently of peroxisome-proliferator-activated receptors. J Biol Chem. 2001;276:38152–38158. doi: 10.1074/jbc.M101980200. [DOI] [PubMed] [Google Scholar]
- 28.Kon K, Ikejima K, Hirose M, Yoshikawa M, Enomoto N, Kitamura T, Takei Y, Sato N. Pioglitazone prevents early-phase hepatic fibrogenesis caused by carbon tetrachloride. Biochem Biophys Res Commun. 2002;291:55–61. doi: 10.1006/bbrc.2002.6385. [DOI] [PubMed] [Google Scholar]
- 29.Galli A, Crabb DW, Ceni E, Salzano R, Mello T, Svegliati-Baroni G, Ridolfi F, Trozzi L, Surrenti C, Casini A. Antidiabetic thiazolidinediones inhibit collagen synthesis and hepatic stellate cell activation in vivo and in vitro. Gastroenterology. 2002;122:1924–1940. doi: 10.1053/gast.2002.33666. [DOI] [PubMed] [Google Scholar]
- 30.Chitturi S, George J. Hepatotoxicity of commonly used drugs: nonsteroidal anti-inflammatory drugs, antihypertensives, antidiabetic agents, anticonvulsants, lipid-lowering agents, psychotropic drugs. Semin Liver Dis. 2002;22:169–183. doi: 10.1055/s-2002-30102. [DOI] [PubMed] [Google Scholar]
- 31.Wang MY, Unger RH. Role of PP2C in cardiac lipid accumulation in obese rodents and its prevention by troglitazone. Am J Physiol Endocrinol Metab. 2005;288:E216–E221. doi: 10.1152/ajpendo.00004.2004. [DOI] [PubMed] [Google Scholar]
- 32.Wakino S, Hayashi K, Kanda T, Tatematsu S, Homma K, Yoshioka K, Takamatsu I, Saruta T. Peroxisome proliferator-activated receptor gamma ligands inhibit Rho/Rho kinase pathway by inducing protein tyrosine phosphatase SHP-2. Circ Res. 2004;95:e45–55. doi: 10.1161/01.RES.0000142313.68389.92. [DOI] [PubMed] [Google Scholar]
- 33.Enomoto N, Takei Y, Hirose M, Konno A, Shibuya T, Matsuyama S, Suzuki S, Kitamura KI, Sato N. Prevention of ethanol-induced liver injury in rats by an agonist of peroxisome proliferator-activated receptor-gamma, pioglitazone. J Pharmacol Exp Ther. 2003;306:846–854. doi: 10.1124/jpet.102.047217. [DOI] [PubMed] [Google Scholar]
- 34.Kawaguchi K, Sakaida I, Tsuchiya M, Omori K, Takami T, Okita K. Pioglitazone prevents hepatic steatosis, fibrosis, and enzyme-altered lesions in rat liver cirrhosis induced by a choline-deficient L-amino acid-defined diet. Biochem Biophys Res Commun. 2004;315:187–195. doi: 10.1016/j.bbrc.2004.01.038. [DOI] [PubMed] [Google Scholar]
- 35.Tomita K, Azuma T, Kitamura N, Nishida J, Tamiya G, Oka A, Inokuchi S, Nishimura T, Suematsu M, Ishii H. Pioglitazone prevents alcohol-induced fatty liver in rats through up-regulation of c-Met. Gastroenterology. 2004;126:873–885. doi: 10.1053/j.gastro.2003.12.008. [DOI] [PubMed] [Google Scholar]
- 36.Rutenburg AM, Kim H, Fischbein JW, Hanker JS, Wasserkrug HL, Seligman AM. Histochemical and ultrastructural demonstration of gamma-glutamyl transpeptidase activity. J Histochem Cytochem. 1969;17:517–526. doi: 10.1177/17.8.517. [DOI] [PubMed] [Google Scholar]
- 37.Han C, Demetris AJ, Michalopoulos GK, Zhan Q, Shelhamer JH, Wu T. PPARgamma ligands inhibit cholangiocarcinoma cell growth through p53-dependent GADD45 and p21 pathway. Hepatology. 2003;38:167–177. doi: 10.1053/jhep.2003.50296. [DOI] [PubMed] [Google Scholar]
- 38.Pinzani M, Gesualdo L, Sabbah GM, Abboud HE. Effects of platelet-derived growth factor and other polypeptide mitogens on DNA synthesis and growth of cultured rat liver fat-storing cells. J Clin Invest. 1989;84:1786–1793. doi: 10.1172/JCI114363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Marra F, Gentilini A, Pinzani M, Choudhury GG, Parola M, Herbst H, Dianzani MU, Laffi G, Abboud HE, Gentilini P. Phosphatidylinositol 3-kinase is required for platelet-derived growth factor's actions on hepatic stellate cells. Gastroenterology. 1997;112:1297–1306. doi: 10.1016/s0016-5085(97)70144-6. [DOI] [PubMed] [Google Scholar]
- 40.Kinnman N, Hultcrantz R, Barbu V, Rey C, Wendum D, Poupon R, Housset C. PDGF-mediated chemoattraction of hepatic stellate cells by bile duct segments in cholestatic liver injury. Lab Invest. 2000;80:697–707. doi: 10.1038/labinvest.3780073. [DOI] [PubMed] [Google Scholar]
- 41.Pinzani M, Milani S, De Franco R, Grappone C, Caligiuri A, Gentilini A, Tosti-Guerra C, Maggi M, Failli P, Ruocco C, et al. Endothelin 1 is overexpressed in human cirrhotic liver and exerts multiple effects on activated hepatic stellate cells. Gastroenterology. 1996;110:534–548. doi: 10.1053/gast.1996.v110.pm8566602. [DOI] [PubMed] [Google Scholar]
- 42.Pinzani M, Failli P, Ruocco C, Casini A, Milani S, Baldi E, Giotti A, Gentilini P. Fat-storing cells as liver-specific pericytes. Spatial dynamics of agonist-stimulated intracellular calcium transients. J Clin Invest. 1992;90:642–646. doi: 10.1172/JCI115905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Cho JJ, Hocher B, Herbst H, Jia JD, Ruehl M, Hahn EG, Riecken EO, Schuppan D. An oral endothelin-A receptor antagonist blocks collagen synthesis and deposition in advanced rat liver fibrosis. Gastroenterology. 2000;118:1169–1178. doi: 10.1016/s0016-5085(00)70370-2. [DOI] [PubMed] [Google Scholar]
- 44.Morland CM, Fear J, McNab G, Joplin R, Adams DH. Promotion of leukocyte transendothelial cell migration by chemokines derived from human biliary epithelial cells in vitro. Proc Assoc Am Physicians. 1997;109:372–382. [PubMed] [Google Scholar]
- 45.Marra F, Romanelli RG, Giannini C, Failli P, Pastacaldi S, Arrighi MC, Pinzani M, Laffi G, Montalto P, Gentilini P. Monocyte chemotactic protein-1 as a chemoattractant for human hepatic stellate cells. Hepatology. 1999;29:140–148. doi: 10.1002/hep.510290107. [DOI] [PubMed] [Google Scholar]
- 46.Park J, Gores GJ, Patel T. Lipopolysaccharide induces cholangiocyte proliferation via an interleukin-6-mediated activation of p44/p42 mitogen-activated protein kinase. Hepatology. 1999;29:1037–1043. doi: 10.1002/hep.510290423. [DOI] [PubMed] [Google Scholar]
- 47.Ishida Y, Smith S, Wallace L, Sadamoto T, Okamoto M, Auth M, Strazzabosco M, Fabris L, Medina J, Prieto J, et al. Ductular morphogenesis and functional polarization of normal human biliary epithelial cells in three-dimensional culture. J Hepatol. 2001;35:2–9. doi: 10.1016/s0168-8278(01)00078-2. [DOI] [PubMed] [Google Scholar]