

Abstract
A 56-day feeding trial was conducted to evaluate the effects of supplemented diets with heat-killed Lactobacillus plantarum (HK-LP) with graded levels of soybean meal (SBM) on growth, digestibility, blood parameters, and immune response of Seriola dumerili (initial weight, 25.05 ± 0.1 g). Seven isonitrogenous and isolipidic practical diets were formulated to contain 0%, 15%, 30%, and 45% SBM, and each SBM level was supplemented with HK-LP at 0.0 and 0.1%. Fish fed diet which contains 30% SBM with HK-LP grew significantly faster than the other groups with notable feed intake and protein retention. Further, protein gain, whole body protein content, protease activity, protein, and lipid digestibility were significantly increased for all fish groups except for fish fed diet which contains 45% SBM with or without HK-LP. Interestingly, lysozyme activity was significantly enhanced in fish fed diets that contain 15% and 30% SBM with HK-LP. Hematocrit, peroxidase, and bactericidal activities revealed a significant increase in 30% SBM with HK-LP group. In addition, fish fed diets which contain 0% and 30% SBM with HK-LP showed higher tolerance against low-salinity stress compared with other groups. In conclusion, the addition of HK-LP to amberjack diets appeared to improve SBM utilization, immune response, and stress resistance.
1. Introduction
Fish meal (FM) represents an ideal nutritional source of dietary protein for fish. Increasing demand, unstable supply, and high prices of FM along with the continuous expansion of aquaculture are reasons for many nutritionists to realize that soon they will no longer be able to afford it as a major protein source in aquafeeds. Currently one of the challenges that fish nutritionists face is the need to partially or totally replace FM with less expensive, non-traditional animal or plant protein sources [1, 2].
Soybean proteins have been recognized as one of the most appropriate alternative protein sources for FM in aquafeed because of their consistent nutritional composition, comparatively balanced amino acid profile, availability, and reasonable price [3]. Soybean meal (SBM) has proven to be well accepted by yellowtail [4–6]. Tomás et al. [7] investigated the possible use of SBM as a substitute for FM in the diets of yellowtail by progressively increasing its inclusion level. The authors found a decrease in final weights as the SBM content increased starting from 30% protein substitution rate.
Methods for increasing SBM's inclusion rates in soy-sensitive species such as amberjack are required, and one of the methods is to apply dietary supplementation of functional compounds. Non-viable microbes exhibit beneficial effects due to their function as immunostimulants. Using live bacteria may cause a potential risk to wild aquatic organisms considering the fact the bacteria may escape into the environment. Therefore, the use of inactivated bacteria clearly solves such safety-related issues since they can no longer interact with other aquatic organisms [8]. Besides that, inactivated bacteria are considered one of the most practical candidates of feed additives. This is due to its high tolerance against temperatures which are produced during preparation of fish diets especially in the course of mincing and pelleting, without affecting its functional activity. This makes it more efficient than other candidates to achieve success in fish farming [8–10].
Heat-killed Lactobacillus plantarum (HK-LP) is a potential candidate as one of the functional additives for fish. Recently, effects of HK-LP have been investigated as immunostimulants [11–14]. Khonyoung and Yamauchi [14] reported that the diet supplemented with HK-LP (L-137) might activate intestinal function by increasing segmented filamentous bacteria, while inducing a better body weight gain in broilers. Oral administration of HK-LP has enhanced growth performance and immune responses of larval and postlarval Kuruma shrimp, Marsupenaeus japonicus bate [13, 15]. Oral administration of inactivated Lactobacillus delbrueckii subsp. lactis and Bacillus subtilis appears to cause good immune stimulatory properties of gilthead seabream (Sparus aurata L.) [8, 9]. Oral administration of heat-killed Enterococcus faecalis enhanced growth performance and immune responses of rainbow trout [10]. Furthermore, heat-killed bacteria were also compared with live form in tilapia [16]. According to these observations, it was hypothesized that HK-LP may also be effective in responses of growth and non-specific immune systems of amberjack, Seriola dumerili.
The amberjack is one of the most important cultured species in Japan because of its delicacy and comparatively higher market value. It is distributed throughout the tropical and subtropical seas except the Pacific Ocean [17, 18]. There have been no studies about dietary SBM and the effect of HK-LP have been undertaken on amberjack to date; the trial reported here was conducted to determine the effects of the partial substitution of FM by SBM with or without HK-LP on growth, digestibility, blood chemistry, immune responses, and stress resistance of amberjack juveniles.
2. Materials and Methods
2.1. Test Diets
Tables 1 and 2 show the composition and chemical analysis of the experimental diets. All the dietary components were obtained commercially, except for HK-LP preparation which was provided by House Wellness Foods Corp. (Itami, Japan) and it contains 20% HK-LP and 80% dextrin in dried-weight basis. HK-LP Prep (LP20) was prepared based on the method previously described by Murosaki et al. [19]. The product was stored at −20°C until use. Using brown fish meal and soybean meal as main protein sources and Pollack liver oil and soybean lecithin as main lipid source, seven isonitrogenous (50.5% crude protein) and isolipidic (12.3% crude lipid) practical diets were formulated to contain 0%, 15%, 30%, and 45% soybean meal and two levels of HK-LP (0.0 and 0.1%) (SBM0, SBM15, SBM15(0.1), SBM30, SBM30(0.1), SBM45, and SBM45(0.1)). Moreover, crystalline amino acid (CAA) mixture of lysine, methionine, betaine, glycine, and alanine were supplemented to meet essential amino acid (EAA) requirements of juvenile amberjack. Wheat flour was supplied as the carbohydrate or nitrogen-free extract source, activated gluten was used as a binder to produce pellet diet, and cellulose powder was used to adjust to 100% total proportion. The diets were prepared by thoroughly mixing all the dry ingredients in a food mixer for 15 minutes. Pollack liver oil, soybean lecithin, and HK-LP Prep were premixed with a sonicator (CA-4488Z, Kaijo Corporation, Tokyo, Japan), added to the dry ingredients, and mixed for another 15 min. Water (35–40% of the dry ingredients) was then added to the premixed ingredients and mixed for an additional 15 min. The pH of the diets was adjusted to the range of 7.0–7.5 with 4 N sodium hydroxide. The mixture was then passed through a meat grinder with an appropriate diameter (2.2–3.1 mm) to prepare pellets, which were then dried in a dry-air mechanical convection oven (DK 400, Yamato Scientific, Tokyo, Japan) at 50°C for about 120 min to approximately 10-11% moisture. The test diets were stored in a freezer at −20°C until use.
Table 1.
Formulation of the experimental diets (% dry matter).
| Ingredient | Soybean meal (SBM) replacement, % (HK-LP Prep., %) | ||||||
|---|---|---|---|---|---|---|---|
| SBM0 | SBM15 | SBM15(0.1) | SBM30 | SBM30(0.1) | SBM45 | SBM45(0.1) | |
| Brown fish meal1 | 61 | 51 | 51 | 40 | 40 | 30.5 | 30.5 |
| Soybean meal2 | 0 | 15 | 15 | 30 | 30 | 45 | 45 |
| Wheat flour | 10 | 8 | 8 | 5 | 5 | 1 | 1 |
| Soybean lecithin3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Pollack liver oil4 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| Vitamin mixture5 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Mineral mixture6 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Stay-C7 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Activated gluten8 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| α-Cellulose | 9 | 5.63 | 5.53 | 4.17 | 4.07 | 2.2 | 2.1 |
| Amino acid premix9 | 0.9 | 1.27 | 1.27 | 1.73 | 1.73 | 2.2 | 2.2 |
| HK-LP Prep10 | 0 | 0 | 0.1 | 0 | 0.1 | 0 | 0.1 |
|
| |||||||
| Total | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
1Nihon Suisan Co. Ltd (Tokyo, Japan), 2J. Oil Mills, Japan, 3,4Riken Vitamins, Tokyo, Japan.
5Vitamin mixture (g kg−1 diet): β-carotene, 0.10; Vitamin D3, 0.01; Menadione NaHSO3·3H2O (K3), 0.05; DL-α-tochopheryl acetate (E), 0.38; thiamine-nitrate (B1), 0.06; riboflavin (B2), 0.19; pyridoxine-HCl (B6), 0.05; cyanocobalamin (B12), 0.0001; biotin, 0.01; inositol, 3.85; niacine (Nicotic acid), 0.77; Ca pantothenate, 0.27; folic acid, 0.01; choline chloride, 7.87; ρ-aminobenzoic acid, 0.38; cellulose, 1.92.
6Mineral mixture (g kg−1 diet): MgSO4, 5.07; Na2HPO4, 3.23; K2HPO4, 8.87; Fe citrate, 1.10; Ca lactate, 12.09; Al(OH)3, 0.01; ZnSO4, 0.13; CuSO4, 0.004; MnSO4, 0.03; Ca(IO3)2, 0.01; CoSO4, 0.04.
7L-ascrobil-2-phosphate-Mg.
8Glico Nutrition Company Ltd. Osaka, Japan. Commercial name: “A-glu SS”.
9Amino acid premix (g 100 g−1 diet) at soybean meal replacement level of 15%; the mixed amino acids just as follows: lysine, 0.20; methionine, 0.17; alanine, 0.30; betaine, 0.30; glycine, 0.30. Amino acid premix (g 100 g−1diet) at fish meal replacement level of 30%; the mixed amino acids just as follows: lysine, 0.46; methionine, 0.38; alanine, 0.30; betaine, 0.30; glycine, 0.30. Amino acid premix (g 100 g−1diet) at fish meal replacement level of 45%; the mixed amino acids just as follows: lysine, 0.72; methionine, 0.58; alanine, 0.30; betaine, 0.30; glycine, 0.30.
10HK-LP Prep: preparation of heat-killed Lactobacillus plantarum made by House Wellness Foods Corp. (Itami, Japan).
Table 2.
Chemical analysis of the experimental diets.
| Ingredient | Soybean meal (SBM) replacement, % (HK-LP Prep., %) | ||||||
|---|---|---|---|---|---|---|---|
| SBM0 | SBM15 | SBM15(0.1) | SBM30 | SBM30(0.1) | SBM45 | SBM45(0.1) | |
| Proximate composition (%, dry matter basis) | |||||||
| Crude protein | 50.81 | 50.47 | 50.34 | 50.15 | 50.74 | 51.38 | 50.82 |
| Total lipid | 11.85 | 12.29 | 11.93 | 12.74 | 12.71 | 12.66 | 12.30 |
| Ash | 11.10 | 10.78 | 11.41 | 11.50 | 11.54 | 11.37 | 11.82 |
| Gross energy (KJ g−1)1 | 19.75 | 19.92 | 19.63 | 19.84 | 19.91 | 19.95 | 19.76 |
|
| |||||||
| Amino acid profiles (AA g 100 g−1 diet, dry matter basis) | |||||||
| Arginine | 2.81 | 2.94 | 2.86 | 3.02 | 2.93 | 3.17 | 3.01 |
| Histidine | 1.42 | 1.23 | 1.35 | 1.36 | 1.47 | 1.54 | 1.39 |
| Isoleucine | 2.26 | 2.24 | 2.04 | 2.38 | 2.18 | 2.27 | 2.47 |
| Leucine | 3.97 | 3.74 | 3.94 | 3.64 | 3.84 | 3.45 | 3.85 |
| Lysine | 3.91 | 3.81 | 3.61 | 3.75 | 3.66 | 3.89 | 3.59 |
| Methionine | 1.77 | 1.60 | 1.54 | 1.39 | 1.59 | 1.54 | 1.24 |
| Phenylalanine | 2.01 | 2.07 | 2.17 | 2.10 | 2.21 | 2.18 | 2.18 |
| Threonine | 2.18 | 2.26 | 2.26 | 2.11 | 2.11 | 2.01 | 2.29 |
| Valine | 2.55 | 2.76 | 2.66 | 2.52 | 2.62 | 2.46 | 2.36 |
| ΣIDAA2 | 22.86 | 22.64 | 22.42 | 22.27 | 22.61 | 22.50 | 22.37 |
1Calculated using combustion values for protein, lipid, and carbohydrate of 23.6, 39.5, and 17.2 kJ g−1, respectively. Carbohydrate was calculated by the difference: 100 − (protein + lipid + ash + moisture).
2ΣIDAA: total indispensable amino acid contents.
2.2. Experimental Fish and Feeding Protocol
Juvenile amberjack (Seriola dumerili), with mean initial body weight of 25.05 ± 0.1 g (mean ± S.E.), were purchased from Kagoshima prefecture seed production center, Kagoshima Prefecture, Japan, and transferred to the Kamoike Marine Production Laboratory, Faculty of Fisheries, Kagoshima University, Japan. The fish were acclimatized for two weeks in laboratory conditions and reared in a 500 L tank with flow-through system. During this period, a commercial diet (50% crude protein; Higashimaru, Japan) was supplied to the fish. Stocking was done at twenty fish per tank with the triplicate tanks per treatment in 200 L polycarbonate tanks (filled with 180 L of water) in a flow-through sea water system where each tank was equipped with an inlet, outlet, and continuous aeration. The tanks were maintained under natural light/dark regime. All fish were fed the respective test diets to satiation level by hand twice a day at 9.00 and 16.00 h., 7 days per week for 56 days. Any uneaten feed left was removed after feeding and dried using a freeze drier then subtracted from the total feed intake. The seawater was pumped from the deep basin of Kagoshima Bay, Japan. It was gravel-filtered and supplied into the system. A flow rate of 1.5 L min−1 was maintained throughout the experimental period. During the experimental period, the monitored water quality parameters (mean ± S.D.) were as follows, water temperature 25.2 ± 1.3°C, pH 8 ± 0.5, salinity 33.3 ± 0.5 ppt, and dissolved oxygen 6.1 ± 0.5 mg L−1. These ranges were considered within optimal values for juvenile amberjack.
2.3. Sample Collection and Biochemical Analysis
At the beginning, a pooled sample of 10 fish was stored at −20°C for initial whole body analysis. While at the end of the feeding trial, all fish were fasted for 24 h prior to final sampling. All the fish were anaesthetized with Eugenol (4-allylmethoxyphenol, Wako Pure Chemical Ind., Osaka, Japan) at 50 mg L−1. Then the total number, individual body weight, and length of fish from each tank were measured. Three fish from each replicate tank were randomly collected and stored at −20°C for final whole body analysis. Blood was taken from the caudal vein of five fish in each replicate tank using heparinized disposable syringes. A small fraction of the heparinized blood was used to analyze the hematocrit and hemoglobin levels. Hematocrit was determined using the microhematocrit technique. Plasma samples were obtained by centrifugation at 3000 ×g for 15 min at 4°C using a high-speed refrigerated microcentrifuge (MX-160; Tomy Tech USA Inc., Tokyo, Japan) and kept at −80°C. In addition, non-heparinized disposable syringes were used to collect blood for serum analysis. Serum samples were obtained by centrifugation at 3000 ×g for 15 min at 4°C to collect serum. Three other fish were randomly sampled from each dietary tank and used for collection of liver and viscera. Viscera and liver were removed then weighed to get viscerosomatic index (VSI) and hepatosomatic index (HSI), respectively. Digestive tracts were separated, cut into small pieces, washed with pure water, pooled together, and stored at −80°C.
Hemoglobin, plasma chemical parameters, and total serum protein (TSP) were measured spectrophotometrically with an automated analyzer (SPOTCHEM EZ model SP-4430, Arkray, Inc., Kyoto, Japan) [20]. Biological antioxidant potential (BAP) and reactive oxygen metabolites (d-ROMs) were also measured spectrophotometrically from blood plasma with an automated analyzer (FRAS4, Diacron International s.r.l., Grosseto, Italy) by following [21, 22]. Plasma cortisol was measured using commercial kits (Cortisol EIA Kit, product number EA65, Oxford Biomedical Research Inc., Oxford, MI) according to the procedure outlined by the manufacturer. Protease activity (PA) was analyzed using digestive organ samples according to Kader et al. [23].
The ingredients, diets, and fish whole body were analyzed for moisture, crude protein, total lipid, and ash, in triplicate, using standard methods [24]. This entailed moisture analysis by oven-drying at 110°C to constant weight, crude protein analysis by the Kjeldahl method, crude lipid analysis by the Soxhlet extraction method, and ash content analysis by combustion in Muffle furnace at 550°C for 4 h. The amino acid profiles of the experimental diets were analyzed by high performance liquid chromatography (HPLC, Shimadzu Corp. Kyoto, Japan) according to the previous studies [22, 25].
2.4. Low-Salinity Stress Test
Tolerance against exposure to low-salinity seawater was examined. After the feeding trial, five fish from each rearing tank (total of 15 fish per treatment) were randomly selected and transferred into a 100 L black tank containing low-salinity water (0.2%). The city tap water was dechlorinated by strongly aerating for at least 24 h and mixed with seawater, and then used as low-salinity water. The tanks for stress test were equipped with continuous aeration and kept under ambient temperature during the stress test. The number of dead fish in each test tank was recorded every 20 min. The passing of time to reach 50% death was calculated using the method previously described by [26, 27].
2.5. Evaluation of Non-Specific Immune Responses
Lysozyme activity of serum was determined with turbidimetric assays [28] at 450 nm with ImmunoMini NJ-2300 (System Instruments, Tokyo, Japan). A unit of enzyme activity was defined as the amount of enzyme that causes a decrease in absorbance of 0.001/min.
The serum bactericidal activity was measured according to Iida et al. [29]. Serum was diluted 3, 4, and 5 times with a Tris buffer (pH 7.5). The dilutions were mixed with a bacterial suspension (0.001 g/mL, Escherichia coli, IAM1239 cell line, Kagoshima, Japan) and incubated at 25°C for 24 h by microtube rotator (MTR-103, AS ONE, Osaka, Japan). The solutions were incubated on TSA (Trypto-Soya agar, Nissui Phatmaceutical Co. Ltd., Japan) at 25°C for 24 h. Colony forming unit (CFU) was counted by the plate counting method as described by Ren et al. [27].
The total peroxidase content in serum was measured according to Salinas et al. [9], with some modifications. Briefly, 15 μL of serum was diluted with 35 μL of Hank's buffered salt solution (HBSS) without Ca+2 or Mg+2 in flat-bottomed 96-well plates. Then, 50 μL of peroxidase substrate (3, 30, 5, 50-tetramethylbenzidine hydrochloride) (TMB; Thermo Scientific Inc., USA) was added. The serum mixture was incubated for 15 min. The colour-developing reaction in serum samples was stopped by adding 50 μL of 2 M sulphuric acid and the OD (450 nm) was measured in a plate reader. PBS was used as a blank instead of serum.
2.6. Digestibility Assessment
Digestibility of each diet was measured after the growth trial. For the digestibility measurement, remaining fish from the same treatments were distributed randomly into duplicate tanks. The fish were fed a diet containing chromium oxide (Wako Pure Chemical Industries, Ltd) as the inert marker at a level of 0.5% (Cr2O3, 5 g/kg) was added. Fish were acclimated to the diet containing chromic oxide for five days. In the morning of the 6th day, fish were fed each diet to apparent satiation twice daily. Six hours after feeding, feces were collected by putting pressure from belly to anus. Feces collection continued for ten days until a sufficient amount of feces had been collected for analysis. Pooled fecal samples were immediately ground after freeze-drying and kept at −20°C until analysis. Concentration of chromium oxide in diets and feces was determined according to Furukawa and Tsukahara [30].
2.7. Evaluation of Growth Performance Parameters
The following variables were evaluated:
-
weight gain (%) = (final weight − initial weight) × 100/initial weight;
-
specific growth rate (SGR %, day−1) = {(Ln(final weight) − Ln(initial weight))/duration (56 days)} × 100;
-
survival (%) = 100 × (final no. of fish/initial no. of fish);
-
feed intake (FI, g fish−1 56 days−1) = (dry diet given − dry remaining diet recovered)/no. of fish;
-
feed efficiency ratio (FER) = live weight gain (g)/dry feed intake (g);
-
protein efficiency ratio (PER) = live weight gain (g)/dry protein intake (g);
-
protein gain (PG, g kg weight gain−1) = {(final weight (g) × final whole body protein content (%)/100) − (initial weight (g) × initial whole body protein content (%)/100)}/(weight gain (g)) × 1000;
-
protein retention (PR, % of intake) = [protein gain (g kg weight gain−1) × 100]/protein intake (g kg weight gain−1);
-
condition factor (CF) = weight of fish (g)/(length of fish)3 (cm)3 × 100;
-
hepatosomatic index (HSI, %) = weight of liver/weight of fish × 100;
-
viscerosomatic index (VSI, %) = weight of viscera/weight of fish × 100;
-
apparent digestibility coefficient (ADC, %) = 100 − [(% Cr2O3 in diet/% Cr2O3 in feces) × (% nutrient in feces/% nutrient in diet)].
2.8. Statistical Analysis
All data were subjected to statistical verification using package super ANOVA 1.11, Abacus Concepts, Berkeley, California, USA. Probabilities of P < 0.05 were considered significant. Differences in significance between means were evaluated using the Turkey Kramer test.
3. Results
3.1. Growth Parameters and Nutrient Utilization
Growth performance and feed utilization of the fish are given in Table 3. Survival (%) of fish did not differ significantly (P > 0.05) among treatments. Final weight, weight gain (%), specific growth rate (SGR), and protein gain (PG) of fish fed SBM30(0.1) diet was significantly higher than those fed the other diets. On the other hand, the growth parameters of fish fed SBM15, SBM15(0.1), and SBM30 were not significantly different from those of fish fed FM (SBM0). The poorest growth performance was found in fish fed SBM45 and SBM45(0.1). However, SGR was significantly (P < 0.05) higher in fish fed SBM45(0.1) than SBM45. Similarly, PG and protein retention (PR) were also significantly decreased in fish fed SBM45 and SBM45(0.1) while no difference was detected between FM (SBM0) and the remaining treatments. However, no difference was detected in feed efficiency ratio (FER) and protein efficiency ratio (PER) between FM (SBM0) and other dietary groups.
Table 3.
Growth parameters and nutrient utilization in amberjack fed test diets for 56 days*.
| Parameters | Soybean meal (SBM) replacement, % (HK-LP Prep., %) | ||||||
|---|---|---|---|---|---|---|---|
| SBM0 | SBM15 | SBM15(0.1) | SBM30 | SBM30(0.1) | SBM45 | SBM45(0.1) | |
| Fn wt1 | 136.6 ± 2.8b | 136 ± 3.06b | 136.7 ± 2.78b | 136.9 ± 3.11b | 149.2 ± 2.36c | 101.7 ± 1.31a | 105.8 ± 1.29a |
| WG2 | 445.1 ± 11.5b | 443.8 ± 11.94b | 446.4 ± 11.21b | 446.8 ± 12.72b | 494.1 ± 8.52c | 304.8 ± 5.78a | 320.95 ± 6.06a |
| SGR3 | 3.03 ± 0.04c | 3.02 ± 0.04c | 3.03 ± 0.04c | 3.04 ± 0.04c | 3.18 ± 0.03d | 2.49 ± 0.03a | 2.57 ± 0.02b |
| FI4 | 120.62 ± 3.06b | 118.68 ± 3.19b | 120.63 ± 1.56b | 119.58 ± 2.39b | 130.27 ± 2.6c | 88.2 ± 4.24a | 90.33 ± 2.68a |
| FER5 | 0.93 ± 0.03 | 0.94 ± 0.04 | 0.93 ± 0.02 | 0.94 ± 0.02 | 0.95 ± 0.03 | 0.87 ± 0.03 | 0.89 ± 0.03 |
| PER6 | 1.82 ± 0.06 | 1.86 ± 0.08 | 1.83 ± 0.05 | 1.85 ± 0.04 | 1.87 ± 0.06 | 1.7 ± 0.06 | 1.76 ± 0.05 |
| PG7 | 203.65 ± 1.88b | 204.38 ± 5.08b | 207.05 ± 3.69b | 200.92 ± 1.22b | 206.85 ± 2.01b | 179.9 ± 1.64a | 183.17 ± 0.2a |
| PR8 | 124.76 ± 2.57b | 122.67 ± 6.1b | 126.49 ± 2.19b | 121.12 ± 3.55b | 137.17 ± 1.93c | 81.32 ± 3.26a | 84.07 ± 2.29a |
| Sur9 | 100 | 95 | 100 | 100 | 100 | 90 | 93.33 |
*Values are means of triplicate groups ± S.E.M. Within a row, means with different letters are significantly different (P < 0.05); means with the same letters are not significantly different (P > 0.05). Absence of letters indicates no significant difference between treatments.
Average initial body weight; means ± S.E.M., 25.05 ± 0.1 g.
1Fn wt: final weight (g), 2WG: percent weight gain (%), 3SGR: specific growth rate (% day−1), 4FI: feed intake (g dry diet fish−1 56 days−1), 5FER: feed efficiency ratio, 6PER: protein efficiency ratio, 7PG: protein gain (g kg body weight gain−1), 8PR: protein retention (% of intake), and 9Sur: survival (%).
Dietary treatments significantly affected (P < 0.05) feed intake (FI) of fish. FI was markedly improved by supplementing CAA and HK-LP Prep. Significantly higher FI was found in fish fed SBM30(0.1) compared to other diet groups. However, there were no significant differences in FI of fish fed SBM15, SBM15(0.1), SBM30, and FM (SBM0). On the other hand, SBM45 and SBM45(0.1) diets were not well accepted by the fish, and the value was significantly lower (P < 0.05) than other test diets.
3.2. Whole Body Proximate Analysis
The proximate composition of the whole body of juvenile amberjack is shown in Table 4. In comparison with the control, dietary treatments had no significant influence on the total lipid and crude ash contents at the end of the feeding trial. However, whole body crude protein contents in all experimental groups were significantly higher than SBM45 and SBM45(0.1) groups. Moreover, moisture content was significantly (P < 0.05) decreased in fish fed SBM0 and SBM15(0.1) groups. No difference (P > 0.05) was also detected in CF, HSI, and VSI of fish among treatments (Table 4).
Table 4.
Whole body proximate analysis (%) and somatic parameters in juvenile amberjack fed test diets for 56 days*.
| Parameters | Initial1 | Soybean meal (SBM) replacement, % (HK-LP Prep., %) | ||||||
|---|---|---|---|---|---|---|---|---|
| SBM0 | SBM15 | SBM15(0.1) | SBM30 | SBM30(0.1) | SBM45 | SBM45(0.1) | ||
| Moisture | 72.84 | 71.07 ± 0.2a | 71.17 ± 0.31ab | 71.02 ± 0.09a | 71.18 ± 0.55ab | 71.2 ± 0.2ab | 72.44 ± 0.05b | 72.44 ± 0.05b |
| Crude protein | 19.58 | 20.22 ± 0.15b | 20.27 ± 0.41b | 20.5 ± 0.3b | 20 ± 0.1b | 20.5 ± 0.16b | 18.38 ± 0.13a | 18.62 ± 0.01a |
| Total lipid | 3.42 | 4.48 ± 0.08 | 4.17 ± 0.14 | 4.35 ± 0.15 | 4.43 ± 0.15 | 4.26 ± 0.05 | 4.49 ± 0.01 | 4.33 ± 0.14 |
| Crude ash | 4.05 | 4.1 ± 0.09 | 3.95 ± 0.09 | 3.78 ± 0.02 | 3.96 ± 0.01 | 3.99 ± 0.05 | 3.75 ± 0.36 | 3.72 ± 0.17 |
| CF2 | — | 1.41 ± 0.02 | 1.39 ± 0.04 | 1.37 ± 0.03 | 1.45 ± 0.03 | 1.4 ± 0.05 | 1.53 ± 0.04 | 1.39 ± 0.04 |
| HSI3 | — | 1.1 ± 0.06 | 1.13 ± 0.12 | 1.08 ± 0.1 | 1.13 ± 0.06 | 1.04 ± 0.09 | 1.35 ± 0.01 | 1.13 ± 0.06 |
| VSI4 | — | 3.75 ± 0.21 | 3.67 ± 0.12 | 3.65 ± 0.27 | 3.84 ± 0.07 | 4.04 ± 0.28 | 3.84 ± 0.05 | 3.78 ± 0.1 |
*Values are means of triplicate groups ± S.E.M. Within a row, means with different letters are significantly different (P < 0.05); means with the same letters are not significantly different (P > 0.05). Absence of letters indicates no significant difference between treatments. Crude protein, crude lipid, and ash are expressed on a wet weight basis.
1Initial values are not included in the statistical analysis.
2CF: condition factor (%), 3HSI: hepatosomatic index (%), and 4VSI: viscerosomatic index.
3.3. Blood Parameters and Responses against Stress
Table 5 represents the blood parameters of amberjack after 56 days of feeding trial. Overall, dietary treatments had no effect on blood chemical parameters of fish except for the case of hematocrit, hemoglobin, glucose, and triglyceride (TG). Hematocrit level was significantly (P < 0.05) higher in fish fed SBM30(0.1) than those fed SBM30 diet while no significant (P > 0.05) differences were detected among other groups. Similarly, hemoglobin was significantly increased in fish fed SBM0 when compared with the SBM45 group while no significant differences were detected among other groups. Plasma glucose content was significantly (P < 0.05) more decreased in the SBM0 group than in other groups. On the other hand, TG was significantly (P < 0.05) higher in SBM0 than SBM30(0.1) group while no significant (P > 0.05) differences were detected among other groups. Experimental diets had no significant effect on the relative value (%) of plasma cortisol levels among all treatments.
Table 5.
Blood parameters in juvenile amberjack fed test diets for 56 days*.
| Parameters | Soybean meal (SBM) replacement, % (HK-LP Prep., %) | ||||||
|---|---|---|---|---|---|---|---|
| SBM0 | SBM15 | SBM15(0.1) | SBM30 | SBM30(0.1) | SBM45 | SBM45(0.1) | |
| Hematocrit (%) | 48.7 ± 1.2ab | 46.7 ± 0.7ab | 47.3 ± 0.3ab | 44 ± 1a | 49.7 ± 1.7b | 44.7 ± 0.9ab | 46.7 ± 1.2ab |
| Hemoglobin (g/dL) | 12.5 ± 0.00b | 12 ± 0.3ab | 12.3 ± 0.2ab | 11.2 ± 0.8ab | 12.2 ± 0.2ab | 10.7 ± 0.2a | 11.8 ± 0.1ab |
| Total protein (g/dL) | 4 ± 0.1 | 3.6 ± 0.3 | 3.9 ± 0.03 | 4.1 ± 0.2 | 4.1 ± 0.3 | 3.6 ± 0.1 | 3.6 ± 0.1 |
| Total bilirubin (mg/dL) | 0.5 ± 0.1 | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.7 ± 0.1 | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.6 ± 0.2 |
| Glucose (mg/dL) | 68 ± 3.1a | 93.3 ± 3.3bc | 97.3 ± 1.2c | 90.7 ± 3.2bc | 89 ± 3.2bc | 79 ± 4.6ab | 85.7 ± 4.4bc |
| GOT (IU/l)1 | 35 ± 12.3 | 44 ± 8.7 | 39.7 ± 3.8 | 55.3 ± 7.8 | 40.3 ± 5.5 | 31.3 ± 3.7 | 53 ± 4.9 |
| GPT (IU/l)2 | <1.00 | <1.00 | <1.00 | <1.00 | <1.00 | <1.00 | <1.00 |
| BUN (mg/dL)3 | 9 ± 0.6 | 8.7 ± 0.9 | 11.7 ± 0.9 | 9 ± 1.5 | 13.3 ± 1.5 | 11 ± 2 | 10.3 ± 0.3 |
| TG (mg/dL)4 | 158 ± 9b | 108 ± 5ab | 145.3 ± 10.7ab | 143.7 ± 22.9ab | 93 ± 4.4a | 123.3 ± 17.6ab | 121 ± 6.5ab |
| T-Cho (mg/dL)5 | 238 ± 3.6 | 242.3 ± 25.8 | 290 ± 4 | 298.3 ± 29.7 | 255.3 ± 15.4 | 251.7 ± 20.5 | 229.3 ± 14.3 |
| CORT (%)6 | 103.1 ± 3.1 | 101.3 ± 2.4 | 100.6 ± 1.4 | 102.2 ± 1.1 | 100 ± 2 | 110 ± 2.5 | 106.9 ± 4.9 |
| d-ROMs7 | 32.7 ± 5.2a | 33.7 ± 4.8a | 51.7 ± 8.8b | 38.3 ± 3.8ab | 32 ± 2.3a | 42.7 ± 3ab | 33.7 ± 5.4ab |
| BAP8 | 3369.3 ± 84ab | 3267.7 ± 169.9ab | 3429.3 ± 335.6ab | 3570.7 ± 16.9ab | 3848 ± 92.5b | 2803 ± 186.6a | 3202.3 ± 108.3ab |
1GOT: glutamyl oxaloacetic transaminase, 2GPT: glutamic-pyruvate transaminase, 3BUN: blood urea nitrogen, 4TG: triglyceride, 5T-Cho: total cholesterol, 6CORT (%): relative value of cortisol, 7d-ROMs: reactive oxygen metabolites, and 8BAP: biological antioxidant potential.
*Values are means of triplicate groups ± S.E.M. Within a row, means with different letters are significantly different (P < 0.05); means with the same letters are not significantly different (P > 0.05). Absence of letters indicates no significant difference between treatments.
Oxidative status of fish was analyzed from plasma (Table 5). The lowest values of reactive oxygen molecules (d-ROMs) were detected in fish fed SBM0, SBM15, and SBM30(0.1) diets. On the other hand, biological antioxidant potential (BAP) was found highest in the SBM30(0.1) group. Figure 2 shows the pattern of combined effects of d-ROMs and BAP. The SBM0, SBM30, and SBM30(0.1) groups were located in zone (A), SBM15(0.1) group in zone (B), SBM15 and SBM45(0.1) in zone (C), and SBM45 group in zone (D), respectively.
Figure 2.
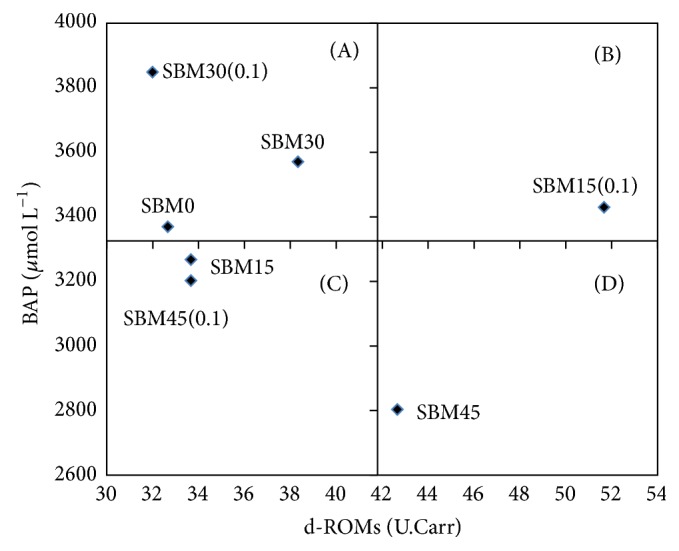
Oxidative stress parameters in amberjack fed test diets for 56 days. Values are expressed as mean ± SE (n = 3). Central axis based on mean values of d-ROMs and BAP from each treatment. Zone (A): high antioxidant potential and low reactive oxygen metabolites (good condition); Zone (B): high antioxidant potential and high reactive oxygen metabolites (acceptable condition); Zone (C): low antioxidant potential and low reactive oxygen metabolites (acceptable condition); Zone (D): low antioxidant potential and high reactive oxygen metabolites (stressed condition). Abbreviation used: SBM0, SBM15, SBM15(0.1), SBM30, SBM30(0.1), SBM 45, and SBM 45(0.1), respectively, refer to soybean replacement/HK-LP, %/%.
Figure 1 shows the results of the low-salinity stress test. The fish that received SBM0 and SBM30(0.1) diets clearly showed significantly (P < 0.05) higher tolerance against low-salinity stress than those of other groups. However, time to 50% mortality was found significantly the lowest in SBM45 group.
Figure 1.
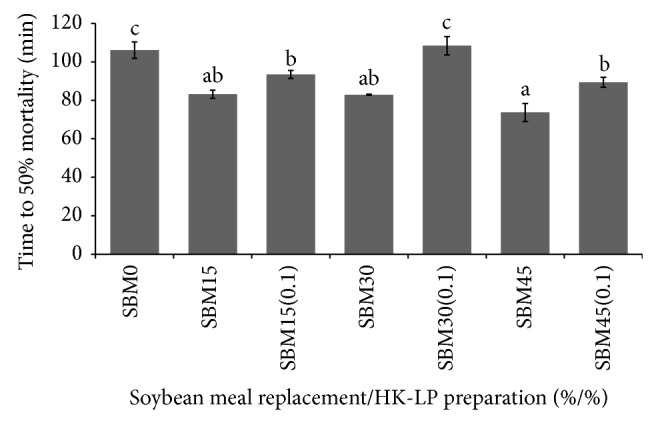
Time to 50% mortality (min) after low salinity stress test when amberjack fed increasing levels of SBM with or without the inclusion of HK-LP for 56 days. Values are means ± SE from triplicate groups. Means with different alphabet are significantly different (P < 0.05).
3.4. Non-Specific Immune Responses
Figure 3 shows immune parameters after 56 days feeding trial. Serum lysozyme activity was significantly (P < 0.05) increased in the SBM15(0.1) group when compared with other groups while no significant difference was detected between the SBM15(0.1) and SBM30(0.1) groups (Figure 3(a)). Fish fed SBM30 diet supplemented with 1 g kg−1 HK-LP showed significantly higher serum bactericidal activity than the other groups (Figure 3(b)). Similarly, serum peroxidase activity recorded the highest significant values (P < 0.05) in the SBM15 and SBM30(0.1) groups (Figure 3(c)) while no significant differences were detected among other groups. Although not statistically significant, the comparatively higher total serum protein values were found in SBM0, SBM15, and SBM15(0.1) groups (Figure 3(d)).
Figure 3.

Immune parameters of amberjack juveniles fed diets containing increasing levels of SBM with or without the inclusion of HK-LP for 56 days. (a) Serum lysozyme activity (unit/mL, n = 3); (b) serum bactericidal activity (108 cfu/mL, n = 3); (c) serum peroxidase activity (n = 9); (d) total serum protein (g/dL, n = 3). Data represent means ± SE. Values with the same letter are not significantly different (P > 0.05). Means with different alphabet are significantly different (P < 0.05). Absence of letters indicates no significant difference between treatments.
3.5. Protease Activity and Digestibility Coefficients
Protease activity (PA, unit mg−1 protein) in the digestive tract of amberjack recorded no significant differences between SBM0, SBM15, SBM15(0.1), SBM30, and SBM30(0.1) groups. However, the SBM45 group recorded the poorest PA value among other experimental groups; moreover, the PA was significantly higher in the SBM45(0.1) group than the SBM45 group. The apparent digestibility coefficient (ADC) of protein was significantly (P < 0.05) higher in fish fed SBM0, SBM15, SBM15(0.1), SBM30, and SBM30(0.1) groups than SBM45 and SBM45(0.1) groups. ADC of lipid was found to be significantly different (P < 0.05) with being higher in the SBM15 and SBM30(0.1) groups than the other experimental groups (Table 6).
Table 6.
Protease activity (PA, unit mg−1 protein) in the digestive tract and apparent digestibility coefficients (ADC) in amberjack fed test diets*.
| Parameters | Soybean meal (SBM) replacement, % (HK-LP Prep., %) | ||||||
|---|---|---|---|---|---|---|---|
| SBM0 | SBM15 | SBM15(0.1) | SBM30 | SBM30(0.1) | SBM45 | SBM45(0.1) | |
| PA (unit mg−1 protein)1 | 0.039 ± 0.001c | 0.04 ± 0.002c | 0.041 ± 0.001c | 0.04 ± 0.001c | 0.043 ± 0.003c | 0.029 ± 0.001a | 0.035 ± 0.001b |
| ADCProtein 2 | 90.01 ± 0.17b | 90.77 ± 0.48b | 89.68 ± 0.6b | 90.11 ± 0.42b | 91.34 ± 0.94b | 86.93 ± 0.69a | 87.59 ± 0.69a |
| ADCLipid 3 | 85.39 ± 0.63b | 89.51 ± 0.23c | 85.78 ± 0.63b | 86.76 ± 0.85b | 88.78 ± 0.5c | 82.32 ± 0.49a | 83.26 ± 0.55a |
*Values are means of triplicate groups ± S.E.M. Within a row, means with different letters are significantly different (P < 0.05); means with the same letters are not significantly different (P > 0.05).
1Protease activity (PA, unit mg−1 protein) in the digestive tract; apparent digestibility coefficients (ADC %). 2For crude protein; 3for lipid, respectively.
4. Discussion
Usually, lower feed intake could be the main reason for reduced growth performance when fish meal was replaced by soybean meal (SBM) [31]. However, the practical application of heat-killed Lactobacillus plantarum (HK-LP) to improve SBM utilization in amberjack diets represents a novel HK-LP application in the present study. Up to 30% SBM replacement level with amino acid mixture did not significantly reduce growth and feed utilization of amberjack. This was consistent with the findings of other previous studies in yellowtail [4–7]. Moreover, SBM30 diet supplemented with 1 g kg−1 HK-LP recorded the highest growth performance compared with other experimental diets. The beneficial effects of HK-LP supplementation on final body weight, weight gain, and specific growth rate of fish fed SBM30(0.1) diet were also found in the study of Tung et al. [13], who reported improved growth performance of kuruma shrimp Marsupenaeus japonicus fed with HK-LP. Growth promoting activity has been noted also in rainbow trout fed diet supplemented with heat-killed Enterococcus faecalis [10]. Significantly higher protein gain and protein retention in fish fed diet SBM30(0.1) would be a possible reason for the higher performances of fish in this group. These results suggest that the tested fish utilized experimental diets effectively by HK-LP supplementation resulting in increased feed intake in SBM30(0.1) group.
Several authors have reported that the dietary administration of different bacterial forms enhanced the secretion of intestinal enzymes and characterization of these enzymes provides some information regarding the digestive capacity of fish to hydrolyze carbohydrate, protein, and lipid of feed ingredients, leading to better growth performance and feed efficiency [32–35]. Khonyoung and Yamauchi [14] reported that the intestine is the direct organ for digestion, absorption, and immunity, as the gut microflora is continuously exposed to other strain of HK-LP (L-137). The latter was also thought to affect the production of extracellular enzymes by the microflora within the gastrointestinal (GI) tract of fish. The bacterial flora in the GI tract of fish shows very broad and variable enzymatic potential, and these enzymatic masses may positively affect the digestive process of fish [34, 36]. All together, the relatively enhanced growth performance and feed efficiency in the amberjack fingerlings fed the HK-LP supplemented diets could be related to the improved intestinal microbiota.
The protease activity (PA) of the digestive tract could provide further insight into the possible effects of different diets on fish performance [23]. In this study, PA was significantly enhanced in the SBM0, SBM15, SBM15(0.1), SBM30, and SBM30(0.1) groups compared to other experimental groups, while PA was significantly higher in fish fed SBM45(0.1) diet than that in fish fed SBM45 without HK-LP diet, indicating the positive effect of HK-LP. Similarly, it has been reported that the secretion of proteases was enhanced by supplementing immunostimulants in yellowtail, Seriola quinqueradiata diet [37]. Watanabe et al. [6] and Tomás et al. [7] reported that ADC of dry matter, protein, and lipid was high due to the process used for preparing diets in which pellets heating might have inactivated the trypsin inhibitor. Previous studies have also demonstrated that growth-promoting additives resulted in the improved digestibility of nutrients [10, 38]. Comparatively low digestibility values recorded here were likely due to the quality of raw material or due to the method of feces collection [39].
Blood parameters are important tools for indication of physiological stress response, general health conditions, and welfare of fish towards nutritional and environmental changes [40]. Blood parameters obtained in the present experiment are considered to be within the normal range for juvenile amberjack, compared to those of the previous findings [18, 41]. Results of the present study showed that the hematocrit values increased in the case of SBM30(0.1) group, implying improved health status. High hematocrit values indicate HK-LP efficiency, wherein iron is evenly distributed without any reduction in the synthesis of hemoglobin. Similarly, Rodriguez-Estrada et al. [10] reported that hematocrit level was enhanced by the supplementation of inactivated Enterococcus faecalis in rainbow trout diets. Moreover, lower triglyceride and cholesterol contents in fish fed SBM30(0.1) diet showed that the optimum availability of HK-LP in fish diets maintains low level of plasma triglycerides and cholesterol in fish.
Oxidative stress was measured using the free radical analytical system assessing the derivatives of oxidative stress by measuring reactive oxygen metabolites (d-ROMs test) and biological antioxidant potential (BAP test) in plasma samples. It is the consequence of an imbalance between oxidants and antioxidants in which oxidant activity exceeds the neutralizing capacity of antioxidants [42]. Recently, d-ROMs and BAP were reported to be reliable parameters for determining the oxidative stress conditions of fish [43]. It would be concluded that fish fed diets SBM0, SBM30, and SBM30(0.1) were in less oxidative stress conditions compared to the SBM45 group in this study.
The lethal stress test is used to assess the healthy status by measuring the lethal time of 50% mortality (LT50) in fresh water of the fish [25]. It is well known that stress affects the survival and growth of fish, since stress responses tend to increase the energy demand at the expense of anabolic processes [44]. The higher value of LT50 in the SBM0 and SBM30(0.1) groups indicated a higher tolerance of the amberjack against low-salinity stress. Fish antioxidant status is strongly related to immune system, contributing to enhance resistance towards different stressors [45]. In the light of the previous findings, results of the current study confirmed a higher tolerance against low-salinity stress in fish in less oxidative stress conditions.
Lysozyme is an important defense molecule of fish innate immune system [46]. Lysozyme activity has been used to evaluate the non-specific defense ability in many fish species, such as Japanese eel, Anguilla japonica [27], yellowtail kingfish, Seriola lalandi [47], and here Japanese flounder, Paralichthys olivaceus [48]. The lowest lysozyme activity was found in SBM45 group, which together with other parameters (bactericidal activity, peroxidase activity, and total serum protein) implied a less-healthy condition of the fish fed with this diet. The increasing trends in serum lysozyme activity in this study might have contributed to the enhancement in the non-specific defense mechanisms [49]. Serum bactericidal activity is one of the most important factors in host resistance against pathogenic bacteria [50]. In this study, the highest serum bactericidal activity was found in SBM30(0.1) group. Similarly, the highest levels of peroxidase were observed in the case of SBM15 and SBM30(0.1) confirming other results obtained by Salinas et al. [9]. From the mentioned results, it could be concluded that the non-specific immune response was enhanced by HK-LP supplementation. Similarly, Irianto and Austin [51] illustrated that dietary supplementation of inactivated bacteria also stimulated the innate immune parameters of rainbow trout, Oncorhynchus mykiss.
The potentials for reducing stress and enhancement of immunity and stress resistance by manipulation of nutritional factors and use of feed additives (such as HK-LP) were demonstrated in this study. However, very little work in this area has been conducted in fish. Thus, the effects of dietary functional feed additives and their interactions need to be assessed to develop economically viable feeds and feeding practices to optimize growth, improve stress resistance, immune response, and disease resistance and improve product quality of aquaculture species.
5. Conclusions
In conclusion, the present study shows that up to 30% SBM substitution level with essential amino acid supplementation did not significantly reduce growth, feed utilization, and immune response of amberjack. Furthermore, the addition of HK-LP to diets appeared to improve SBM utilization by amberjack. However, further studies are needed in order to evaluate the effects of HK-LP on amberjack health with attention to the intestinal microbiota and histology.
Acknowledgments
The first author would like to thank the Egyptian government for financial support. We express our sincere gratitude to Mrs. Amina Moss for reading the paper. This research was partially funded by the Management Expenses Grants of the United Graduate School of Agriculture Sciences, Kagoshima University, provided to Dr. Shunsuke Koshio.
Conflict of Interests
The authors declare that they have no conflict of interests.
References
- 1.Lunger A. N., McLean E., Craig S. R. The effects of organic protein supplementation upon growth, feed conversion and texture quality parameters of juvenile cobia (Rachycentron canadum) Aquaculture. 2007;264(1–4):342–352. doi: 10.1016/j.aquaculture.2006.12.012. [DOI] [Google Scholar]
- 2.Peng M., Xu W., Ai Q., Mai K., Liufu Z., Zhang K. Effects of nucleotide supplementation on growth, immune responses and intestinal morphology in juvenile turbot fed diets with graded levels of soybean meal (Scophthalmus maximus L.) Aquaculture. 2013;392–395:51–58. doi: 10.1016/j.aquaculture.2013.02.002. [DOI] [Google Scholar]
- 3.Storebakken T., Refsite S., Ruyter B. Soy products as fat and protein sources in fish feeds for intensive aquaculture. In: Darckly J. K., editor. Soy in Animal Nutrition. Savoy, Ill, USA: Federation of Animal Science Societies; 2000. pp. 127–170. [Google Scholar]
- 4.Shimeno S., Kumon M., Ando H., Ukawa M. The growth performance and body composition of young yellowtail fed with diets containing defatted soybean meal for a long period. Bulletin of the Japanese Society of Scientific Fisheries. 1993;59:821–825. [Google Scholar]
- 5.Viyakarn V., Watanabe T., Aoki H., et al. Use of soybean meal as a substitute for fish meal in a newly developed soft-dry pellet for yellowtail. Bulletin of the Japanese Society of Scientific Fisheries. 1992;58:1991–2000. [Google Scholar]
- 6.Watanabe T., Viyakarn V., Kimura H., Ogawa K., Okamoto N., Iso N. Utilization of soybean meal as a protein source in a newly developed soft-dry pellet for yellowtail. Bulletin of the Japanese Society of Scientific Fisheries. 1992;58:1761–1773. [Google Scholar]
- 7.Tomás A., de la Gándara F., García-Gomez A., Pérez L., Jover M. Utilization of soybean meal as an alternative protein source in the Mediterranean yellowtail, Seriola dumerili . Aquaculture Nutrition. 2005;11(5):333–340. doi: 10.1111/j.1365-2095.2005.00365.x. [DOI] [Google Scholar]
- 8.Díaz-Rosales P., Salinas I., Rodríguez A., et al. Gilthead seabream (Sparus aurata L.) innate immune response after dietary administration of heat-inactivated potential probiotics. Fish and Shellfish Immunology. 2006;20(4):482–492. doi: 10.1016/j.fsi.2005.06.007. [DOI] [PubMed] [Google Scholar]
- 9.Salinas I., Abelli L., Bertoni F., et al. Monospecies and multispecies probiotic formulations produce different systemic and local immunostimulatory effects in the gilthead seabream (Sparus aurata L.) Fish and Shellfish Immunology. 2008;25(1-2):114–123. doi: 10.1016/j.fsi.2008.03.011. [DOI] [PubMed] [Google Scholar]
- 10.Rodriguez-Estrada U., Satoh S., Haga Y., Fushimi H., Sweetman J. Effects of inactivated Enterococcus faecalis and mannan oligosaccharide and their combination on growth, immunity, and disease protection in rainbow trout. North American Journal of Aquaculture. 2013;75(3):416–428. doi: 10.1080/15222055.2013.799620. [DOI] [Google Scholar]
- 11.Hirose Y., Murosaki S., Yamamoto Y., Yoshikai Y., Tsuru T. Daily intake of heat-killed Lactobacillus plantarum L-137 augments acquired immunity in healthy adults. The Journal of Nutrition. 2006;136(12):3069–3073. doi: 10.1093/jn/136.12.3069. [DOI] [PubMed] [Google Scholar]
- 12.Hirose Y., Murosaki S., Yamamoto Y., et al. Safety studies of LP20 powder produced from heat-killed Lactobacillus plantarum L-137. Regulatory Toxicology and Pharmacology. 2009;54(3):214–220. doi: 10.1016/j.yrtph.2009.03.007. [DOI] [PubMed] [Google Scholar]
- 13.Tung H. T., Koshio S., Traifalgar R. F., Ishikawa M., Yokoyama S. Effects of dietary heat-killed Lactobacillus plantarum on larval and post-larval kuruma shrimp, Marsupenaeus japonicus bate. Journal of the World Aquaculture Society. 2010;41(supplement 1):16–27. doi: 10.1111/j.1749-7345.2009.00329.x. [DOI] [Google Scholar]
- 14.Khonyoung D., Yamauchi K. Effects of heat-killed Lactobacillus plantarum L-137 on morphology of intestinal villi and epithelial cells in broiler chickens. Journal of Applied Animal Research. 2012;40(2):140–147. doi: 10.1080/09712119.2011.640208. [DOI] [Google Scholar]
- 15.Tung H. T., Koshio S., Teshima S., et al. Effects of heat-killed Lactobacillus plantarum supplemental diets on growth performance, stress resistance and immune response of juvenile Kuruma shrimp Marsupenaeus japonicus bate. Aquaculture Science. 2009;57(2):175–184. [Google Scholar]
- 16.Taoka Y., Maeda H., Jo J.-Y., et al. Use of live and dead probiotic cells in tilapia Oreochromis niloticus . Fisheries Science. 2006;72(4):755–766. doi: 10.1111/j.1444-2906.2006.01215.x. [DOI] [Google Scholar]
- 17.Laroche W. A., Smith-Vaniz W. F., Richardson S. L. Carangidae: development. In: Moser H. G., editor. Ontogeny and Systematics of Fishes. Special publication no. 1. Lawrence, Kan, USA: American Society of Ichthyologists and Herpetologists, Allen Press; 1984. pp. 510–522. [Google Scholar]
- 18.Kader M. A., Bulbul M., Hossain M. S., Yokoyama S., Ishikawa M., Koshio S. Improved utilization of a plant by-products mixture by supplementing dietary bamboo charcoal for Juvenile Amberjack Seriola dumerili . South Pacific Studies. 2013;34(1):1–12. [Google Scholar]
- 19.Murosaki S., Yamamoto Y., Ito K., et al. Heat-killed Lactobacillus plantarum L-137 suppresses naturally fed antigen-specific IgE production by stimulation of IL-12 production in mice. Journal of Allergy and Clinical Immunology. 1998;102(1):57–64. doi: 10.1016/S0091-6749(98)70055-7. [DOI] [PubMed] [Google Scholar]
- 20.Tatsumi N., Tsuji R., Yamada T., Kubo K., Matsuda T. Spot chem. EZ SP- 4430 no kiso teki kento. Journal of Clinical Laboratory Instruments and Reagents. 2000;23(6):427–433. [Google Scholar]
- 21.Morganti P., Bruno C., Guarneri F., Cardillo A., Del Ciotto P., Valenzano F. Role of topical and nutritional supplement to modify the oxidative stress. International Journal of Cosmetic Science. 2002;24(6):331–339. doi: 10.1046/j.1467-2494.2002.00159.x. [DOI] [PubMed] [Google Scholar]
- 22.Kader M. A., Bulbul M., Koshio S., et al. Effect of complete replacement of fishmeal by dehulled soybean meal with crude attractants supplementation in diets for red sea bream, Pagrus major . Aquaculture. 2012;350–353:109–116. doi: 10.1016/j.aquaculture.2012.04.009. [DOI] [Google Scholar]
- 23.Kader M. A., Koshio S., Ishikawa M., Yokoyama S., Bulbul M. Supplemental effects of some crude ingredients in improving nutritive values of low fishmeal diets for red sea bream, Pagrus major . Aquaculture. 2010;308(3-4):136–144. doi: 10.1016/j.aquaculture.2010.07.037. [DOI] [Google Scholar]
- 24.AOAC. Official Methods of Analysis. 16th. Washington, DC, USA: Association of Official Analytical Chemists; 1998. [Google Scholar]
- 25.Han Y., Koshio S., Ishikawa M., Yokoyama S. Interactive effects of dietary arginine and histidine on the performances of Japanese flounder Paralichthys olivaceus juveniles. Aquaculture. 2013;414-415:173–182. doi: 10.1016/j.aquaculture.2013.07.001. [DOI] [Google Scholar]
- 26.Moe Y. Y., Koshio S., Teshima S.-I., Ishikawa M., Matsunaga Y., Panganiban A., Jr. Effect of vitamin C derivatives on the performance of larval kuruma shrimp, Marsupenaeus japonicus . Aquaculture. 2004;242(1–4):501–512. doi: 10.1016/j.aquaculture.2004.08.028. [DOI] [Google Scholar]
- 27.Ren T., Koshio S., Ishikawa M., et al. Influence of dietary vitamin C and bovine lactoferrin on blood chemistry and non-specific immune responses of Japanese eel, Anguilla japonica . Aquaculture. 2007;267(1–4):31–37. doi: 10.1016/j.aquaculture.2007.03.033. [DOI] [Google Scholar]
- 28.Lygren B., Sveier H., Hjeltness B., Waagbø R. Examination of the immunomodulatory properties and the effect on disease resistance of dietary bovine lactoferrin and vitamin C fed to atlantic salmon (Salmo salar) for a short-term period. Fish and Shellfish Immunology. 1999;9(2):95–107. doi: 10.1006/fsim.1998.0179. [DOI] [Google Scholar]
- 29.Iida T., Takahashi T., Wakabayashi H. Decrease in the bactericidal activity of normal serum during the spawning period of rainbow trout. Bulletin of the Japanese Society of Scientific Fisheries. 1989;55:463–465. [Google Scholar]
- 30.Furukawa A., Tsukahara H. On the acid digestion method for the determination of chromic oxides as an index substance in the study of digestion of fish feed. Bulletin of the Japanese Society of Scientific Fisheries. 1966;32:502–506. [Google Scholar]
- 31.Chen W., Ai Q., Mai K., et al. Effects of dietary soybean saponins on feed intake, growth performance, digestibility and intestinal structure in juvenile Japanese flounder (Paralichthys olivaceus) Aquaculture. 2011;318(1-2):95–100. doi: 10.1016/j.aquaculture.2011.04.050. [DOI] [Google Scholar]
- 32.Tovar D., Zambonino J., Cahu C., Gatesoupe F. J., Vázquez-Juárez R., Lésel R. Effect of live yeast incorporation in compound diet on digestive enzyme activity in sea bass (Dicentrarchus labrax) larvae. Aquaculture. 2002;204(1-2):113–123. doi: 10.1016/S0044-8486(01)00650-0. [DOI] [Google Scholar]
- 33.Mohapatra S., Chakraborty T., Prusty A. K., Das P., Paniprasad K., Mohanta K. N. Use of different microbial probiotics in the diet of rohu, Labeo rohita fingerlings: effects on growth, nutrient digestibility and retention, digestive enzyme activities and intestinal microflora. Aquaculture Nutrition. 2012;18(1):1–11. doi: 10.1111/j.1365-2095.2011.00866.x. [DOI] [Google Scholar]
- 34.Ray A. K., Ghosh K., Ringø E. Enzyme-producing bacteria isolated from fish gut: a review. Aquaculture Nutrition. 2012;18(5):465–492. doi: 10.1111/j.1365-2095.2012.00943.x. [DOI] [Google Scholar]
- 35.Lemieux H., Blier P., Dutil J.-D. Do digestive enzymes set a physiological limit on growth rate and food conversion efficiency in the Atlantic cod (Gadus morhua)? Fish Physiology and Biochemistry. 1999;20(4):293–303. doi: 10.1023/A:1007791019523. [DOI] [Google Scholar]
- 36.Das P., Mandal S., Khan A., Manna S. K., Ghosh K. Distribution of extracellular enzyme-producing bacteria in the digestive tracts of 4 brackish water fish species. Turkish Journal of Zoology. 2014;38(1):79–88. doi: 10.3906/zoo-1205-3. [DOI] [Google Scholar]
- 37.Morimoto Kofuji P. Y., Hosokawa H., Masumoto T. Effects of dietary supplementation with feeding stimulants on yellowtail Seriola quinqueradiata (Temminck & Schlegel; Carangidae) protein digestion at low water temperatures. Aquaculture Research. 2006;37(4):366–373. doi: 10.1111/j.1365-2109.2005.01435.x. [DOI] [Google Scholar]
- 38.Ringø E., Gatesoupe F.-J. Lactic acid bacteria in fish: a review. Aquaculture. 1998;160(3-4):177–203. doi: 10.1016/S0044-8486(97)00299-8. [DOI] [Google Scholar]
- 39.Regost C., Arzel J., Kaushik S. J. Partial or total replacement of fish meal by corn gluten meal in diet for turbot (Psetta maxima) Aquaculture. 1999;180(1-2):99–117. doi: 10.1016/S0044-8486(99)00026-5. [DOI] [Google Scholar]
- 40.Maita M., Satoh K., Satoh S., Kiron V., Watanabe T. Effects of Non-Fish Meal Diet on Hematological Parameters, Disease Resistance and Lipid Profiles of Liver and Erythrocytes in Yellow Tail. Beijing, China: World Aquaculture Society Book of Abstracts; 2002. [Google Scholar]
- 41.Ruchimat T., Masumoto T., Hosokawa H., Shimeno S. Quantitative methionine requirement of yellowtail (Seriola quinqueradiata) Aquaculture. 1997;150(1-2):113–122. doi: 10.1016/S0044-8486(96)01465-2. [DOI] [Google Scholar]
- 42.Celi P., Sullivan M., Evans D. The stability of the reactive oxygen metabolites (d-ROMs) and biological antioxidant potential (BAP) tests on stored horse blood. The Veterinary Journal. 2010;183(2):217–218. doi: 10.1016/j.tvjl.2008.09.018. [DOI] [PubMed] [Google Scholar]
- 43.Gao J., Koshio S., Ishikawa M., Yokoyama S., Mamauag R. E. P., Han Y. Effects of dietary oxidized fish oil with vitamin E supplementation on growth performance and reduction of lipid peroxidation in tissues and blood of red sea bream Pagrus major . Aquaculture. 2012;356-357:73–79. doi: 10.1016/j.aquaculture.2012.05.034. [DOI] [Google Scholar]
- 44.Kubilay A., Ulukoy G. The effects of acute stress on rainbow trout (Oncorhynchus mykiss) Turkish Journal of Zoology. 2002;26(2):249–254. [Google Scholar]
- 45.Tovar-Ramírez D., Mazurais D., Gatesoupe J. F., Quazuguel P., Cahu C. L., Zambonino-Infante J. L. Dietary probiotic live yeast modulates antioxidant enzyme activities and gene expression of sea bass (Dicentrarchus labrax) larvae. Aquaculture. 2010;300(1–4):142–147. doi: 10.1016/j.aquaculture.2009.12.015. [DOI] [Google Scholar]
- 46.Saurabh S., Sahoo P. K. Lysozyme: an important defence molecule of fish innate immune system. Aquaculture Research. 2008;39(3):223–239. doi: 10.1111/j.1365-2109.2007.01883.x. [DOI] [Google Scholar]
- 47.Le K. T., Fotedar R., Partridge G. Selenium and vitamin E interaction in the nutrition of yellowtail kingfish (Seriola lalandi): physiological and immune responses. Aquaculture Nutrition. 2014;20(3):303–313. doi: 10.1111/anu.12079. [DOI] [Google Scholar]
- 48.Zhou J., Song X. L., Huang J., Wang X. H. Effects of dietary supplementation of A3α-peptidoglycan on innate immune responses and defense activity of Japanese flounder (Paralichthys olivaceus) Aquaculture. 2006;251(2–4):172–181. doi: 10.1016/j.aquaculture.2005.06.015. [DOI] [Google Scholar]
- 49.Engstad R. E., Robertsen B., Frivold E. Yeast glucan induces increase in lysozyme and complement-mediated haemolytic activity in Atlantic salmon blood. Fish & Shellfish Immunology. 1992;2(4):287–297. doi: 10.1016/S1050-4648(06)80033-1. [DOI] [Google Scholar]
- 50.Kawakami H., Yamashita H., Sakai M. Comparative sensitivity of yellowtail Seriola quinqueradiata and goldstriped amberjack S. aureovittata to Photobacterium damsela subsp. piscicida . Journal of the World Aquaculture Society. 2000;31(2):213–217. doi: 10.1111/j.1749-7345.2000.tb00355.x. [DOI] [Google Scholar]
- 51.Irianto A., Austin B. Use of dead probiotic cells to control furunculosis in rainbow trout, Oncorhynchus mykiss (Walbaum) Journal of Fish Diseases. 2003;26(1):59–62. doi: 10.1046/j.1365-2761.2003.00414.x. [DOI] [PubMed] [Google Scholar]