

Abstract
Epigenetic mechanisms play a crucial role in cellular proliferation, migration and differentiation in both normal and neoplastic development. One of the key signaling pathways whose components are altered through the epigenetic mechanisms is the Wnt signaling pathway. In this review, we briefly discuss the key concepts of epigenetics and focus on the recent advances in the Wnt signaling pathway research and its potential diagnostic and therapeutic implications.
KEY WORDS: cancer, epigenetics, wnt signaling pathway
INTRODUCTION
When we think of epigenetics, we mean those changes in the DNA molecule that alter the expression of genes, but keep intact the DNA nucleotide sequence. The term was introduced by Conrad Waddington when describing inherited changes in cellular phenotype independent of DNA bases alterations [1]. Three epigenetic mechanisms whereby a cell regulates its basic processes of transcription, translation, replication and DNA repair include DNA methylation, histone methylation and acetylation (known as histone code) and the action of small interfering RNA molecules that disrupt protein synthesis [2]. Epigenetic changes may be inherited and can occur during embryonal development or after birth. Once the change in DNA methylation takes place, following cell division the altered pattern is transferred into daughter cells by the action DNA methyltransferase enzyme, which recognizes hemimethylated sites and methylates newly synthesized DNA formed during replication [3].
Since these mechanisms are utilized by all our cells to conduct their predetermined functions, it is likely that they become altered in malignant cells, and possibly even initiate malignant transformation [4]. In the case of DNA methylation, the most extensively studied mechanism of epigenetic control of gene expression it is global hypomethylation in tumor cells that leads to chromosomal (genome) instability, which is central to carcinogenesis [5]. At the same time, hypermethylation of promoter regions has been detected in a vast majority of tumor suppressor genes, which are strongly associated with tumor development as well [6]. Besides DNA methylation, tumor cells are also characterized by modification in their chromatin structure, termed “histone code” by Jenuwein and Allis in 2001, which was established after years of comparing histone acetylation and methylation between normal and neoplastic tissues [6,7].
One of the main benefits of epigenetic regulation discoveries is that epigenetic changes are reversible and that they can be chemically controlled by drugs. Such “epigenetic drugs” fall largely into two broad groups: DNA methyltransferase inhibitors and histone deacetylase inhibitors [8]. Some of them have already been approved for clinical use in the treatment of certain malignant diseases (e.g. azacitidine, decitabine, vorinostat, romidepsine and ruxolitinib), whereas other drugs are still under investigation, in different phases of preclinical and clinical studies [e.g. histone lysine acetyltransferase, histone lysine methyltransferase, histone arginine methyltransferase, poly(ADP-ribose) polymerase inhibitors] [9]. In addition to therapeutic potential, epigenetic changes can also be employed in early tumor detection, prognosis and evaluation of ongoing anti-cancer therapy. For example, detection of septin 9 (SEPT9) gene hypermethylation in the blood has been used in colorectal carcinoma screening whereas detection of glutathione S-transferase pi (GSTP1) gene hypermethylation in urine has been utilized as a prostate cancer biomarker [10-12]. Most of the signaling pathways that affect cellular proliferation, migration and differentiation play important roles in both embryonal development and in tumor initiation and progression. One of the most important such signaling pathways is the evolutionary conserved Wnt pathway whose member components are epigenetically controlled [13].
Epigenetic modification of the Wnt signaling pathway members
The evolutionarily conserved Wnt-signaling pathway has pivotal roles during the development of many organ systems. Central to this pathway is a multiprotein scaffold composed of adenomatous polyposis coli (APC), glycogen synthase kinase (GSK)–3β, axin and the transcriptional cofactor β-catenin (encoded by CTNNB1 gene). In the absence of Wnt ligands, β-catenin levels retain low through constitutive phosphorylation by GSK-3β, which leads to the ubiquitination and degradation of β-catenin [14]. Binding of the Wnt ligands to their receptors, Frizzled (Fz), leads to activation of the adaptor protein, Disheveled (Dvl), and the inhibition of GSK-3 activity, reducing phosphorylation and subsequent degradation of β-catenin (Figure 1). Thus, β -catenin is stabilized and translocates to the nucleus where it binds members of the T-cell factor (Tcf)/lymphoid-enhancing factor (Lef) family of transcription factors and induces the expression of target genes such as CMYC and CCND1 [15].
FIGURE 1.
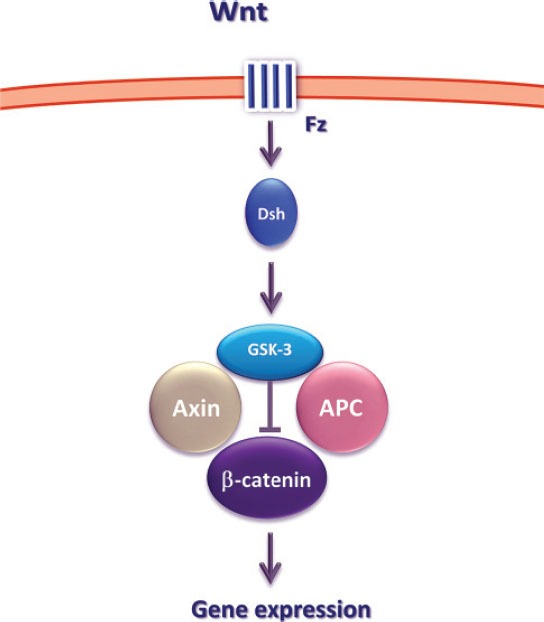
Wnt signaling pathway members: Central to this pathway is a multiprotein scaffold composed of adenomatous polyposis coli (APC), glycogen synthase kinase (GSK)–3β, axin and the transcriptional cofactor β-catenin. Fz – Frizzled receptor.
Notably, a range of human cancers shows elevated levels of nuclear β-catenin, a hallmark of active WNT/β-catenin signaling while mutations of APC, AXIN or CTNNB1 genes are substantially less frequent. Aberrant activation of WNT/β-catenin signaling can also be achieved by overexpression of pathway components including some oncogenic WNT ligands (WNT1, WNT2, WNT2B2 and WNT3A), Fzd receptors (Fzd7 and Fzd10) and Dvl family members [16-18].
For a variety of human malignancies, there is growing evidence that an epigenetic inactivation of the WNT/β-catenin pathway inhibitors is associated with a tumor-favorable phenotypic outcome. Abnormal Wnt signaling has become a hallmark of some types of solid tumors, most notably colorectal and hepatocellular carcinomas [19]. Functional loss of negative Wnt regulators by epigenetic gene silencing, through both DNA methylation and histone modification of the tumor suppressor gene-associated promoters, has been frequently reported to contribute to the activation of aberrant WNT/β-catenin signaling in tumors [20]. Recent studies have shown that impaired regulation of Wnt-antagonists such as SFRP, WIF1, HDPR1 and DKK3 by promoter hypermethylation is present in several human malignancies [21-23]. For examples, some Wnt proteins like Wnt1, Wnt2, Wnt3A and Wnt5A have been found to be overexpressed in cancers of breast, colon, lung and prostate, acting as oncogenic activators for canonical Wnt signaling [24-26]. In some other solid tumors and hematological malignancies, WNT5A acts as a tumor suppressor that inhibits tumor cell proliferation through antagonizing the WNT/β-catenin signaling and is frequently silenced by tumor-specific methylation [27,28]. In parallel, epigenetic inactivation of WNT7A and WNT9A, through promoter methylation, has recently been reported in pancreatic and colorectal cancer and acute lymphoid leukemia [29,30].
As epigenetic dysregulation of WNT/β-catenin signaling frequently contributes to tumor pathogenesis, identification of aberrant epigenetic events that activate WNT/β-catenin signaling may provide useful biomarkers for cancer detection and prognosis prediction. Veeck et al. reported that aberrant promoter methylation of SFRP1 was associated with an overall shorter survival of patients with breast cancer [31]. SFRP2 methylation was also shown to be a tumor-specific biomarker for early breast cancer, while WIF1 methylation was a poor prognostic factor for acute promyelocytic leukemia [32,33]. Yu et al. also reported that Dickkopf-related protein (DKK3) methylation was an independent prognostic marker for gastric cancer and strongly associated with poor survival of patients [34].
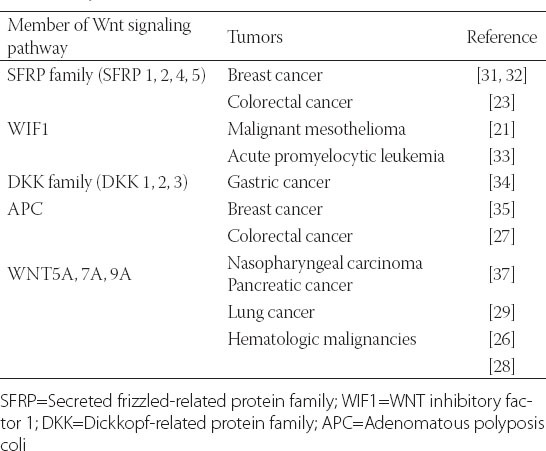
Hypermethylation of the gene promoters of Wnt inhibitors was observed in various cell lines and tissues and was associated with down-regulation of gene expression. The epithelial adhesion molecule E-cadherin (encoded by CDH1) also acts as a negative regulator of WNT/β-catenin signaling by affecting the intracellular localization of β-catenin. Epigenetic silencing of CDH1, the gene encoding E-cadherin, by promoter methylation has been observed in various cancers, leading to aberrant activation of WNT/β-catenin signaling [35]. The growing list of epigenetically silenced WNT antagonists involved in human cancers indicates an important role for epigenetic inactivation events in tumor initiation and progression (see Table 1). Epigenetic changes (promoter methylation or histone methylation/deacetylation) are pharmacologically reversible, using epigenetic agents including DNA methyltransferase inhibitors (5-aza-2’-deoxycytidine, Zebularine) and histone deacetylase inhibitors (TSA, SAHA and PXD101) [36,37].
TABLE 1.
Members of the WNT signaling pathway with aberrant DNA methylation in human tumors.
CONCLUSIONS
Epigenetic mechanisms play a crucial role in normal and neoplastic development. Growing evidence indicates that the Wnt signaling pathway components are epigenetically regulated and are actively involved in the pathogenesis of various human malignancies. Further investigations are necessary to translate the current epigenetic discoveries into useful diagnostic and therapeutic tools.
DECLARATION OF INTEREST
The authors declare no conflict of interest.
REFERENCES
- [1].Goldberg AD, Allis CD, Bernstein E. Epigenetics: a landscape takes shape. Epigenetics. 2007;128:635–638. doi: 10.1016/j.cell.2007.02.006. [DOI] [PubMed] [Google Scholar]
- [2].Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell. 2012;150:12–27. doi: 10.1016/j.cell.2012.06.013. http://dx.doi.org/10.1016/j.cell.2012.06.013 . [DOI] [PubMed] [Google Scholar]
- [3].Serman A, Vlahovic M, Serman L, Bulic-Jakus F. DNA methylation as regulatory mechanism for gene expression in mammals. Coll Antropol. 2006;30:665–671. [PubMed] [Google Scholar]
- [4].Baylin SB, Jones PA. A decade of exploring the cancer epigenome –biological and translational implications. Nat Rev Cancer. 2011;11:726–734. doi: 10.1038/nrc3130. http://dx.doi.org/10.1038/nrc3130 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Brookes E, Shi Y. Diverse epigenetic mechanisms of human disease. Annu Rev Genet. 2014. doi:10.1146/annurev-genet-120213-092518. http://dx.doi.org/10.1146/annurev-genet-120213-092518 . [DOI] [PubMed]
- [6].Hattori N, Ushijima T. Compentium of aberrant DNA methylation and histone modification in cancer. Biochem Biophys Res Commun. 2014. doi:10.1016/j.bbrc.2014.08.140. http://dx.doi.org/10.1016/j.bbrc.2014.08.140 . [DOI] [PubMed]
- [7].Jenuwein T, Allis CD. Translating the histone code. Science. 2001;293:1074–1080. doi: 10.1126/science.1063127. http://dx.doi.org/10.1126/science.1063127 . [DOI] [PubMed] [Google Scholar]
- [8].Miceli M, Bontempo P, Nebbioso A, Altucci L. Natural compounds in epigenetics: A current view. Food Chem Toxicol. 2014;73C:71–83. doi: 10.1016/j.fct.2014.08.005. http://dx.doi.org/10.1016/j.fct.2014.08.005 . [DOI] [PubMed] [Google Scholar]
- [9].Geutjes EJ, Bajpe PK, Bernards RB. Targeting the epigenome for treatment of cancer. Oncogene. 2012;2012:3827–3844. doi: 10.1038/onc.2011.552. http://dx.doi.org/10.1038/onc.2011.552 . [DOI] [PubMed] [Google Scholar]
- [10].Lofton-Day C, Model F, Devos T, Tetzner R, Distler J, et al. DNA methylation biomarkers for blood-based colorectal cancer screening. Clin Chem. 2008;54:414–423. doi: 10.1373/clinchem.2007.095992. http://dx.doi.org/10.1373/clinchem.2007.095992 . [DOI] [PubMed] [Google Scholar]
- [11].Millar DS, Ow KK, Paul CL, Russell PJ, Molloy PL, Clark SJ. Detailed methylation analysis of the glutathione S-transferase pi (GSTP1) gene in prostate cancer. Oncogene. 1999;18:1313–1324. doi: 10.1038/sj.onc.1202415. http://dx.doi.org/10.1038/sj.onc.1202415 . [DOI] [PubMed] [Google Scholar]
- [12].Jimenez RE, Fischer AH, Petros JA, Amin MB. Glutathione S-transferase pi gene methylation: the search for a molecular marker of prostatic adenocarcinoma. Adv Anat Pathol. 2000;7:382–389. doi: 10.1097/00125480-200007060-00005. http://dx.doi.org/10.1097/00125480-200007060-00005 . [DOI] [PubMed] [Google Scholar]
- [13].Surana R, Sikka S, Cai W, Shin EM, Warrier SR, Tan HJ, et al. Secreted frizzled related proteins: Implications in cancers. Biochim Biophys Acta. 2014;1845:53–65. doi: 10.1016/j.bbcan.2013.11.004. [DOI] [PubMed] [Google Scholar]
- [14].Widelitz R. Wnt signaling through canonical and non-canonical pathways: recent progress. Growth Factors. 2005;23:111–116. doi: 10.1080/08977190500125746. http://dx.doi.org/10.1080/08977190500125746 . [DOI] [PubMed] [Google Scholar]
- [15].Staal FJ, Clevers HC. WNT signalling and haematopoiesis: a WNT-WNT situation. Nat Rev Immunol. 2005;5:21–30. doi: 10.1038/nri1529. http://dx.doi.org/10.1038/nri1529 . [DOI] [PubMed] [Google Scholar]
- [16].Bonci D, Coppola V, Musumeci M, Addario A, Giuffrida R, Memeo L, et al. The miR-15a-miR-16-1 cluster controls prostate cancer by targeting multiple oncogenic activities. Nat Med. 2008;14:1271–1277. doi: 10.1038/nm.1880. http://dx.doi.org/10.1038/nm.1880 . [DOI] [PubMed] [Google Scholar]
- [17].Uematsu K, Kanazawa S, You L, He B, Xu Z, Li K, et al. Wnt pathway activation in mesothelioma: evidence of Dishevelled overexpression and transcriptional activity of beta-catenin. Cancer Res. 2003;63:4547–4551. [PubMed] [Google Scholar]
- [18].Terasaki H, Saitoh T, Shiokawa K, Katoh M. Frizzled-10, upregulated in primary colorectal cancer, is a positive regulator of the WNT—beta-catenin—TCF signaling pathway. Int J Mol Med. 2002;9:107–112. [PubMed] [Google Scholar]
- [19].Gregorieff A, Clevers H. Wnt signaling in the intestinal epithelium: from endoderm to cancer. Genes Dev. 2005;19:877–890. doi: 10.1101/gad.1295405. http://dx.doi.org/10.1101/gad.1295405 . [DOI] [PubMed] [Google Scholar]
- [20].Jones PA, Baylin SB. The fundamental role of epigenetic events in cancer. Nat Rev Genet. 2002;3:415–428. doi: 10.1038/nrg816. [DOI] [PubMed] [Google Scholar]
- [21].Batra S, Shi Y, Kuchenbecker KM, He B, Reguart N, Mikami I, et al. Wnt inhibitory factor-1, a Wnt antagonist, is silenced by promoter hypermethylation in malignant pleural mesothelioma. Biochem Biophys Res Commun. 2006;342:1228–1232. doi: 10.1016/j.bbrc.2006.02.084. http://dx.doi.org/10.1016/j.bbrc.2006.02.084 . [DOI] [PubMed] [Google Scholar]
- [22].Liu TH, Raval A, Chen SS, Matkovic JJ, Byrd JC, Plass C. CpG island methylation and expression of the secreted frizzled-related protein gene familyin chronic lymphocytic leukemia. Cancer Res. 2006;66:653–658. doi: 10.1158/0008-5472.CAN-05-3712. http://dx.doi.org/10.1158/0008-5472.CAN-05-3712 . [DOI] [PubMed] [Google Scholar]
- [23].Suzuki H, Watkins DN, Jair KW, Schuebel KE, Markowitz SD, Chen WD, et al. Epigenetic inactivation of SFRP genes allows constitutive WNT signaling in colorectal cancer. Nat Genet. 2004;36:417–422. doi: 10.1038/ng1330. http://dx.doi.org/10.1038/ng1330 . [DOI] [PubMed] [Google Scholar]
- [24].Katoh M. Expression and regulation of WNT1 in human cancer: upregulation of WNT1 by beta-estradiol in MCF-7 cells. Int J Oncol. 2003;22:209–212. [PubMed] [Google Scholar]
- [25].Bonci D, Coppola V, Musumeci M, Addario A, Giuffrida R, Memeo L, et al. The miR-15a-miR-16-1 cluster controls prostate cancer by targeting multiple oncogenic activities. Nat Med. 2008;14:1271–1277. doi: 10.1038/nm.1880. http://dx.doi.org/10.1038/nm.1880 . [DOI] [PubMed] [Google Scholar]
- [26].Huang CL, Liu D, Nakano J, Ishikawa S, Kontani K, Yokomise H, et al. Wnt5a expression is associated with the tumor proliferation and the stromal vascular endothe¬lial growth factor—an expression in non-small-cell lung cancer. J Clin Oncol. 2005;23:8765–8773. doi: 10.1200/JCO.2005.02.2871. http://dx.doi.org/10.1200/JCO.2005.02.2871 . [DOI] [PubMed] [Google Scholar]
- [27].Ying J, Li H, Yu J, Ng KM, Poon FF, Wong SC, et al. WNT5A exhibits tumor-sup-pressive activity through antagonizing the Wnt/beta-catenin signaling, and is frequently methylated in colorectal cancer. Clin Cancer Res. 2008;14:55–61. doi: 10.1158/1078-0432.CCR-07-1644. http://dx.doi.org/10.1158/1078-0432.CCR-07-1644 . [DOI] [PubMed] [Google Scholar]
- [28].Ying J, Li H, Chen YW, Srivastava G, Gao Z, Tao Q. WNT5A is epigenetically silenced in hematologic malignancies and inhibits leukemia cell growth as a tumor suppressor. Blood. 2007;110:4130–4132. doi: 10.1182/blood-2007-06-094870. http://dx.doi.org/10.1182/blood-2007-06-094870 . [DOI] [PubMed] [Google Scholar]
- [29].Sato N, Fukushima N, Maitra A, Matsubayashi H, Yeo CJ, Cameron JL, et al. Discovery of novel targets for aberrant methylation in pancreatic carcinoma using high-throughput microarrays. Cancer Res. 2003;63:3735–3742. [PubMed] [Google Scholar]
- [30].Shu J, Jelinek J, Chang H, Shen L, Qin T, Chung W, et al. Silencing of bidirectional promoters by DNA methylation in tumorigenesis. Cancer Res. 2006;66:5077–5084. doi: 10.1158/0008-5472.CAN-05-2629. http://dx.doi.org/10.1158/0008-5472.CAN-05-2629 . [DOI] [PubMed] [Google Scholar]
- [31].Veeck J, Niederacher D, An H, Klopocki E, Wiesmann F, Betz B, et al. Aberrant methylation of the Wnt antagonist SFRP1 in breast cancer is associated with unfavourable prognosis. Oncogene. 2006;25:3479–3488. doi: 10.1038/sj.onc.1209386. http://dx.doi.org/10.1038/sj.onc.1209386 . [DOI] [PubMed] [Google Scholar]
- [32].Veeck J, Noetzel E, Bektas N, Jost E, Hartmann A, Knuchel R, et al. Promoter hypermethylation of the SFRP2 gene is a high-frequent alteration and tumor-specific epigenetic marker in human breast cancer. Mol Cancer. 2008;7:83. doi: 10.1186/1476-4598-7-83. http://dx.doi.org/10.1186/1476-4598-7-83 . [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Chim CS, Chan WW, Pang A, Kwong YL. Preferential methylation of Wnt inhibitory factor-1 in acute promyelocytic leukemia: an independent poor prognostic factor. Leukemia. 2006;20:907–909. doi: 10.1038/sj.leu.2404176. http://dx.doi.org/10.1038/sj.leu.2404176 . [DOI] [PubMed] [Google Scholar]
- [34].Yu J, Tao Q, Cheng YY, Lee KY, Ng SS, Cheung KF, et al. Promoter methylation of the Wnt/beta-catenin signaling antagonist Dkk-3 is associated with poor survival in gastric cancer. Cancer. 2009;115:49–60. doi: 10.1002/cncr.23989. http://dx.doi.org/10.1002/cncr.23989 . [DOI] [PubMed] [Google Scholar]
- [35].Prasad CP, Mirza S, Sharma G, Prashad R, DattaGupta S, Rath G, et al. Epigenetic alterations of CDH1 and APC genes: relationship with activation of Wnt/beta-catenin pathway in invasive ductal carcinoma of breast. Life Sci. 2008;83:318–325. doi: 10.1016/j.lfs.2008.06.019. http://dx.doi.org/10.1016/j.lfs.2008.06.019 . [DOI] [PubMed] [Google Scholar]
- [36].Cheng JC, Yoo CB, Weisenberger DJ, Chuang J, Wozniak C, Liang G, et al. Preferential response of cancer cells to zebularine. Cancer Cell. 2004;6:151–158. doi: 10.1016/j.ccr.2004.06.023. http://dx.doi.org/10.1016/j.ccr.2004.06.023 . [DOI] [PubMed] [Google Scholar]
- [37].Tao Q, Chan AT. Nasopharyngeal carcinoma: molecular pathogenesis and therapeutic developments. Expert Rev Mol Med. 2007;9:1–24. doi: 10.1017/S1462399407000312. http://dx.doi.org/10.1017/S1462399407000312 . [DOI] [PubMed] [Google Scholar]