

Abstract
Planctomyces brasiliensis Schlesner 1990 belongs to the order Planctomycetales, which differs from other bacterial taxa by several distinctive features such as internal cell compartmentalization, multiplication by forming buds directly from the spherical, ovoid or pear-shaped mother cell and a cell wall consisting of a proteinaceous layer rather than a peptidoglycan layer. The first strains of P. brasiliensis, including the type strain IFAM 1448T, were isolated from a water sample of Lagoa Vermelha, a salt pit near Rio de Janeiro, Brasil. This is the second completed genome sequence of a type strain of the genus Planctomyces to be published and the sixth type strain genome sequence from the family Planctomycetaceae. The 6,006,602 bp long genome with its 4,811 protein-coding and 54 RNA genes is a part of the G enomic E ncyclopedia of Bacteria and Archaea project. Phylogenomic analyses indicate that the classification within the Planctomycetaceae is partially in conflict with its evolutionary history, as the positioning of Schlesneria renders the genus Planctomyces paraphyletic. A re-analysis of published fatty-acid measurements also does not support the current arrangement of the two genera. A quantitative comparison of phylogenetic and phenotypic aspects indicates that the three Planctomyces species with type strains available in public culture collections should be placed in separate genera. Thus the genera Gimesia, Planctopirus and Rubinisphaera are proposed to accommodate P. maris, P. limnophilus and P. brasiliensis, respectively. Pronounced differences between the reported G + C content of Gemmata obscuriglobus, Singulisphaera acidiphila and Zavarzinella formosa and G + C content calculated from their genome sequences call for emendation of their species descriptions. In addition to other features, the range of G + C values reported for the genera within the Planctomycetaceae indicates that the descriptions of the family and the order should be emended.
Keywords: Non-peptidoglycan bacteria, Stalked bacteria, Halotolerant, Gram-negative, Taxonomic descriptions, Planctomycetales, Planctomycetes, GEBA
Introduction
Strain IFAM 1448T (=DSM 5305 = ATCC 49424 = JCM 21570) is the type strain of Planctomyces brasiliensis. Although the genus currently consists of six species with validly published names, only three of them, P. brasiliensis, P. limnophilus, and P. maris contain cultured type strains [1-3]. The other three species, including the type species P. bekefii[4] as well as P. stranskae and P. guttaeformis[5] are to date not represented by cultured strains and have been described solely on the basis of their morphological properties [6], with the descriptions and illustrations serving as the type material. P. bekefii was initially described as a fungus under the International Code of Botanical Nomenclature [4,7]. The genus name derives from the Greek words ‘planktos’, wandering, floating, and ‘mukês’ meaning ‘fungus’ to indicate a floating fungus [4], reflecting their initial descriptions as members of the fungi. The species epithet ‘brasiliensis’ is a Latin masculine adjective that means “of or belonging to Brazil” [1]. Strain IFAM 1448T together with other strains designated IFAM 1450, IFAM 1454 and IFAM 1456 were isolated from water samples collected in November 1982 from Lagoa Vermelha, a salt pit at the Atlantic coast north of Rio de Janeiro, Brazil [1]. Another strain (DSM 11908) potentially belonging to P. brasiliensis was isolated from postlarvae of the Giant Tiger Prawn, Penaeus monodon, infected with the monodon baculovirus [8].
The similarity of genes of both cultured and uncultured Planctomyces species has been studied not only based on 16S rRNA gene sequences, but also using the RNase P RNA genes [9]. The unique compartmentalized cell structure shared by all Planctomycetes investigated so far is remarkable [10], as is the relatively large size of Planctomycetes genomes, which could be attributed to their free-living lifestyles [6]. The membrane organization of organisms belonging to the Planctomycetes, Verrucomicrobia and Chlamydiae superphylum is currently the subject of intense discussion in the literature, e.g. as an exception to [11] or as a variation of [12] the classical Gram-negative cell plan. P. limnophilus has been established as model organism for the phylum, as it is the first and only species of Planctomycetes that has been genetically modified [13,14]. Here we present a summary classification and a set of features for P. brasiliensis IFAM 1448T, together with the description of the complete genomic sequencing and annotation, as well as a re-assessment of the taxonomy of the group based on phylogenomic as well as traditionally sampled characters.
Organism information
Classification and features
16S rRNA gene analysis
A representative genomic 16S rRNA gene sequence of strain DSM 5305T was compared with the Greengenes database for determining the weighted relative frequencies of taxa and (truncated) keywords as previously described [15]. The most frequently occurring genus was Planctomyces (100.0%) (18 hits in total). Regarding the two hits to sequences from members of the species, the average identity within high-scoring segment pairs (HSPs) was 100.0%, whereas the average coverage by HSPs was 97.2%. Regarding the five hits to sequences from other members of the genus, the average identity within HSPs was 90.1%, whereas the average coverage by HSPs was 58.5%. Among all other species, the one yielding the highest score was P. maris, which corresponded to an identity of 90.3% and an HSP coverage of 63.5%. The highest-scoring environmental sequence was DQ015774 (‘Antarctic lake water clone ELB25-062’) [16], which showed an identity of 96.2% and an HSP coverage of 98.3%. The most frequently occurring keywords within the labels of environmental samples that yielded hits were ‘treatment’ (2.7%), ‘microbi’ (2.1%), ‘sediment’ (1.8%), ‘gut’ (1.8%) and ‘microbiom’ (1.7%) (232 hits in total). Environmental samples which yielded hits of a higher score than the highest scoring species were not found, which might indicate that P. brasiliensis is rarely found in the environment. But overall Planctomyces species have been isolated from an ecologically large variety of environments [2,3], which is also reflected by the above most frequently occurring keywords.
Figure 1 shows the phylogenetic neighborhood of P. brasiliensis DSM 5305T in a 16S rDNA based tree. The sequences of the two identical 16S rRNA gene copies in the genome do not differ from the previously published 16S rDNA sequence (AJ231190).
Figure 1.
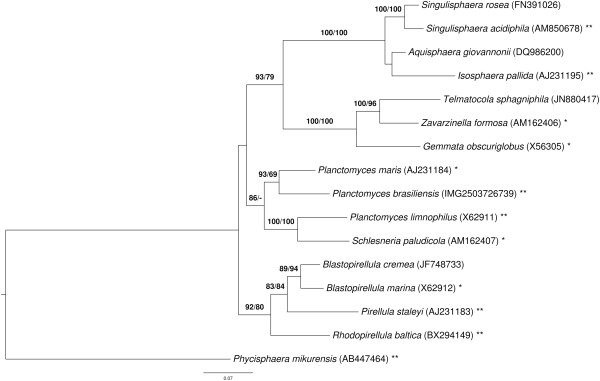
Phylogenetic tree highlighting the position of P. brasiliensis relative to the other species within the family Planctomycetaceae . The tree was inferred from 1,343 aligned characters of the 16S rRNA gene sequence under the maximum likelihood (ML) criterion as previously described [15]. Rooting was done initially using the midpoint method [17] and then checked for its agreement with the current classification (Table 1). The branches are scaled in terms of the expected number of substitutions per site. Numbers above the branches are support values from 400 ML bootstrap replicates (left) and from 1,000 Maximum-Parsimony bootstrap replicates (right) if larger than 60% [15]. Lineages with type strain genome sequencing projects registered in GOLD [18] as unpublished are marked with one star, those listed as published (as well as the target organism) with two stars [19-22].
Morphology and physiology
The morphology and life cycle of the organism resembles those of P. maris and P. limnophilus rather than P. guttaeformis or P. stranskae[1]. Adult cells have an excreted stalk consisting of many loosely twisted fibrils at the distal end where the holdfast is located, enabling the cells to attach to surfaces or to attach to one another, aggregating to rosettes (Figure 2) [1]. The reproductive pole is located opposite the site of the stalk. At the free cell pole, the bud has a polar to sub-polar, thick flagellum with a diameter of about 20 nm. Both the mother cell and bud have crateriform structures scattered over the whole of the cell surface [1]. The cells are spherical to ovoid with a diameter of 0.7 to 1.8 μm, hence, the cells are larger than those of P. limnophilus or P. maris[1]. The colonies have a dry, rough surface and a yellow to ochre pigmentation [1]. P. brasiliensis requires NaCl for growth and has a broad tolerance to salt (0.1 to 1.7 mol Na+/l) [1]. The strain grows chemoorganotrophically (Table 1) on the following carbon sources: D-cellobiose, D(+)-glucose, D(+)-galactose, maltose, D(+)-mannose, melibiose, rhamnose, ribose, trehalose, N-acetyl glucosamine and glucuronate. Carbon sources not utilized are D(−)-fructose, D-fucose, D(−)-lyxose, α-D-melezitose, raffinose, L(−)-sorbose, D(+)-xylose, methanol, ethanol, glycerol, D(−)-mannitol, D(−)-sorbitol, acetate, fumarate, lactate, malate, pyruvate and succinate [1]. Strain IFAM 1448T can utilize ammonia, nitrate and N-acetyl glucosamine as a nitrogen source and can hydrolyze gelatin and tween 80. It does not utilize L-glutamic acid, L-glutamine, L-serine amygdalin, gluconate or creatinine and does not require vitamins for growth. The organism is able to reduce nitrate to nitrite under anaerobic conditions, a trait that is also predicted from the genome sequence (phenotype prediction by the IMG Pathway browser) [1].
Figure 2.

Scanning-electron micrograph of P. brasiliensis DSM 5305 T highlighting stalks and crateriform structures on the cell surface.
Table 1.
Classification and general features of P. brasiliensis DSM 5305 T in accordance with the MIGS recommendations[23] as published by the Genome Standards Consortium [24]
| MIGS ID | Property | Term | Evidence code |
|---|---|---|---|
| |
Current classification |
Domain Bacteria |
TAS [25] |
| Phylum Planctomycetes |
TAS [26] |
||
| Class "Planctomycetacia" |
TAS [26] |
||
| Order "Planctomycetales" |
TAS [27-29] |
||
| Family Planctomycetaceae |
TAS [27,29] |
||
| Genus Planctomyces |
TAS [4,28,30,31] |
||
| Species Planctomyces brasiliensis |
TAS [32] |
||
| Type strain DSM 5305 |
TAS [1] |
||
| |
Gram stain |
negative |
TAS [1] |
| |
Cell shape |
sphere shaped |
TAS [1] |
| |
Motility |
motile |
TAS [1] |
| |
Sporulation |
none |
TAS [1] |
| |
Temperature range |
mesophile |
TAS [1] |
| |
Optimum temperature |
27°C-35°C |
TAS [1] |
| |
Salinity |
halotolerant |
TAS [1] |
| MIGS-22 |
Oxygen requirement |
aerobe |
TAS [1] |
| |
Carbon source |
several sugars, such as D-cellobiose, maltose, trehalose |
TAS [1] |
| |
Energy metabolism |
chemoorganotroph |
TAS [1] |
| MIGS-6 |
Habitat |
marine fresh water |
TAS [1] |
| MIGS-15 |
Biotic relationship |
free-living |
NAS |
| MIGS-14 |
Pathogenicity |
not reported |
NAS |
| |
Biosafety level |
1 |
NAS [33] |
| |
Isolation |
water from salt pit |
TAS [1] |
| MIGS-4 |
Geographic location |
Lagoa Vermelha, Brazil |
TAS [1] |
| MIGS-5 |
Sample collection time |
November 1982 |
TAS [1] |
| MIGS-4.1 |
Latitude |
−22.929 |
NAS |
| MIGS-4.2 |
Longitude |
−42.390 |
NAS |
| MIGS-4.3 |
Depth |
surface water |
TAS [1] |
| MIGS-4.4 | Altitude | 0 m, sea level | NAS |
Evidence codes - TAS: Traceable Author Statement (i.e., a direct report exists in the literature); NAS: Non-traceable Author Statement (i.e., not directly observed for the living, isolated sample, but based on a generally accepted property for the species, or anecdotal evidence). These evidence codes are from the Gene Ontology project [34].
Chemotaxonomy
Chemical analysis of the cell wall of P. brasiliensis showed that proteins are the principal constituents with a value of 79.9% of the dry weight [35]. Detailed analysis of the protein composition indicated that the major amino acids were asparagine, threonine, serine, glutamine, proline, glycine and alanine [36]. Total extractable lipids represent 10% of the cell dry weight of strain IFAM 1448T[23]. The major fatty acids (>1%) are C16:0 (30.7%), C16:1 (25%), C18:1 (13.1%), C20:1 (9.9%), C15:0 (8.4%), C18:0 (5.6%), C14:0 (4.0%), C17:0 (1.3%) and C17:1 (1.8%) [36]. A comparison with other representatives of the Planctomycetes is given below.
Genome sequencing and annotation
Genome project history
This organism was selected for sequencing on the basis of its phylogenetic position [37,38], and is part of the G enomic E ncyclopedia of Bacteria and Archaea project [39]. The genome project is deposited in the Genome On Line Database [18] and the complete genome sequence is deposited in GenBank. Sequencing, finishing and annotation were performed by the DOE Joint Genome Institute (JGI) using state-of-the-art sequencing technology [40]. A summary of the project information is shown in Table 2.
Table 2.
Genome sequencing project information
| MIGS ID | Property | Term |
|---|---|---|
| MIGS-31 |
Finishing quality |
Finished |
| MIGS-28 |
Libraries used |
Three genomic libraries: one 454 pyrosequence standard library, one 454 PE library (14 kb insert size), one Illumina library |
| MIGS-29 |
Sequencing platforms |
Illumina GAii, 454 GS FLX Titanium |
| MIGS-31.2 |
Sequencing coverage |
124.8 × Illumina; 91.0 × pyrosequence |
| MIGS-30 |
Assemblers |
Newbler version 2.0.00.20-PostRelease-10-28-2008-g-3.4.6, phrap |
| MIGS-32 |
Gene calling method |
Prodigal 1.4, GenePRIMP |
| |
INSDC ID |
CP002546 |
| |
Genbank Date of Release |
March 2, 2011 |
| |
GOLD ID |
Gc01674 |
| |
NCBI project ID |
47863 |
| |
Database: IMG-GEBA |
2503707005 |
| MIGS-13 |
Source material identifier |
DSM 5305 |
| Project relevance | Tree of Life, GEBA |
Growth conditions and DNA isolation
A culture of DSM 5305T was grown in DSMZ medium 607 (M13 Verrucomicrobium medium) [41] at 30°C. DNA was isolated from 0.5-1 g of cell paste using Jetflex Genomic DNA Purification Kit (GENOMED 600100) following the standard protocol as recommended by the manufacturer: Cell lysis was enhanced by adding 20 μl proteinase K for two hours at 58°C. DNA is available through the DNA Bank Network [42].
Genome sequencing and assembly
The genome was sequenced using a combination of Illumina and 454 sequencing platforms. All general aspects of library construction and sequencing can be found at the JGI website [43]. Pyrosequencing reads were assembled using the Newbler assembler (Roche). The initial Newbler assembly consisting of 53 contigs in one scaffold was converted into a Phrap assembly (http://www.phrap.com) making fake reads from the consensus, to collect the read pairs in the 454 paired-end library. Illumina GAii sequencing data (3,029.9 Mb) were assembled with Velvet [44] and the consensus sequences were shredded into 1.5 kb overlapped fake reads and assembled together with the 454 data. 454 draft assembly was based on 164.2 Mb 454 draft data and all of the 454 paired end data. Newbler parameters were -consed -a 50 -l 350 -g -m -ml 20. The Phred/Phrap/Consed software package (http://www.phrap.com/) was used for sequence assembly and quality assessment in the subsequent finishing process. After the shotgun stage, reads were assembled with parallel phrap (High Performance Software, LLC). Possible mis-assemblies were corrected with gapResolution (http://www.jgi.doe.gov/), Dupfinisher, or sequencing cloned bridging PCR fragments with subcloning or transposon bombing (Epicentre Biotechnologies, Madison, WI) [45]. Gaps between contigs were closed by editing in Consed, by PCR and by Bubble PCR primer walks (J.-F. Chang, unpublished). A total of 156 additional reactions were necessary to close gaps and to raise the quality of the finished sequence. Illumina reads were also used to correct potential base errors and increase consensus quality using the Polisher software developed at JGI [46]. The error rate of the completed genome sequence is less than one in 100,000. Together, the combination of the Illumina and 454 sequencing platforms provided 285.0 × coverage of the genome. The final assembly contained 281,884 pyrosequence and 39,867,623 Illumina reads.
Genome annotation
Genes were identified using Prodigal [47] as part of the DOE-JGI [48] genome annotation pipeline, followed by a round of manual curation using the JGI GenePRIMP pipeline [49]. The predicted CDSs were translated and used to search the National Center for Biotechnology Information (NCBI) nonredundant database, UniProt, TIGR-Fam, Pfam, PRIAM, KEGG, COG, and InterPro databases. Additional gene prediction analysis and functional annotation was performed within the Integrated Microbial Genomes - Expert Review (IMG-ER) platform [50].
Genome properties
The genome consists of a 6,006,602 bp long chromosome with a G + C content of 56.5% (Table 3 and Figure 3). Of the 4,865 genes predicted, 4,811 were protein-coding genes, and 54 RNAs; 61 pseudogenes were also identified. The majority of the protein-coding genes (57.6%) were assigned with a putative function while the remaining ones were annotated as hypothetical proteins. The distribution of genes into COGs functional categories is presented in Table 4.
Table 3.
Genome statistics
| Attribute | Value | % of total |
|---|---|---|
| Genome size (bp) |
6,006,602 |
100.00% |
| DNA coding region (bp) |
5,145,779 |
85.67% |
| DNA G + C content (bp) |
3,390,645 |
56.45% |
| Number of replicons |
1 |
|
| Extrachromosomal elements |
0 |
|
| Total genes |
4,865 |
100.00% |
| RNA genes |
54 |
1.11% |
| rRNA operons |
2 |
|
| Protein-coding genes |
4,811 |
98.89% |
| Pseudo genes |
61 |
1.25% |
| Genes with function prediction |
2,800 |
57.55% |
| Genes in paralog clusters |
2,351 |
48.32% |
| Genes assigned to COGs |
3,220 |
66.19% |
| Genes assigned Pfam domains |
3,439 |
70.69% |
| Genes with signal peptides |
645 |
13.26% |
| Genes with transmembrane helices |
1,210 |
24.87% |
| CRISPR repeats | 2 |
Figure 3.

Graphical circular map of chromosome. From outside to the center: Genes on forward strand (colored by COG categories), Genes on reverse strand (colored by COG categories), RNA genes (tRNAs green, rRNAs red, other RNAs black), GC content, GC skew (purple/olive).
Table 4.
Number of genes associated with the general COG functional categories
| Code | Value | % age | Description |
|---|---|---|---|
| J |
149 |
4.0 |
Translation, ribosomal structure and biogenesis |
| A |
3 |
0.0 |
RNA processing and modification |
| K |
216 |
5.7 |
Transcription |
| L |
223 |
5.9 |
Replication, recombination and repair |
| B |
4 |
0.1 |
Chromatin structure and dynamics |
| D |
32 |
0.9 |
Cell cycle control, cell division, chromosome partitioning |
| Y |
0 |
0.0 |
Nuclear structure |
| V |
83 |
2.2 |
Defense mechanisms |
| T |
213 |
5.7 |
Signal transduction mechanisms |
| M |
237 |
6.3 |
Cell wall/membrane/envelope biogenesis |
| N |
193 |
5.1 |
Cell motility |
| Z |
0 |
0.0 |
Cytoskeleton |
| W |
0 |
0.0 |
Extracellular structures |
| U |
250 |
6.7 |
Intracellular trafficking, secretion, and vesicular transport |
| O |
140 |
3.7 |
Posttranslational modification, protein turnover, chaperones |
| C |
166 |
4.4 |
Energy production and conversion |
| G |
190 |
5.1 |
Carbohydrate transport and metabolism |
| E |
208 |
5.6 |
Amino acid transport and metabolism |
| F |
61 |
1.6 |
Nucleotide transport and metabolism |
| H |
131 |
3.5 |
Coenzyme transport and metabolism |
| I |
98 |
2.6 |
Lipid transport and metabolism |
| P |
258 |
6.9 |
Inorganic ion transport and metabolism |
| Q |
68 |
1.8 |
Secondary metabolites biosynthesis, transport and catabolism |
| R |
461 |
12.3 |
General function prediction only |
| S |
356 |
9.5 |
Function unknown |
| - | 1,645 | 33.8 | Not in COGs |
Insights from the genome sequence
Taxonomic classification vs. 16S rRNA gene analysis of Planctomyces and Schlesneria
Although it has only slowly been appreciated by taxonomists after Darwin had published his seminal works, the sole possible goal of a taxonomic classification is to summarize the genealogy of the organisms [37,51]. For this reason, it does not matter if a taxonomic classification contains less information than the empirical estimate of the phylogeny from which it was derived. But it does matter whether or not a classification can pretend to summarize the respective underlying genealogies – and this is never the case where classifications include obviously non-monophyletic groups [37,51-53].
As indicated earlier [19], in this respect the current classification of Planctomycetes is only partially satisfactory because with the description of the genus Schlesneria the genus Planctomyces now appears to be paraphyletic in the 16S rRNA gene trees (see Figure 1 and [19-21,54]), with P. limnophilus being more closely related to Schlesneria than to the other Planctomyces species. The two versions of the 16S rRNA gene sequence of Schlesneria paludicola MPL7T submitted to INSDC, NR_042466 and AM162407, are identical; AM162407 has been used in Figure 1. The previously published 16S rRNA gene sequences of P. limnophilus[19] and P. brasiliensis (this study) have also been found to be identical or almost identical to those from the respective genome-sequencing projects. For this reason, the biological identity of the Schlesneria and Planctomyces strains used can hardly be called into question, and a mix-up or contamination of cultures cannot explain the positioning of the strains in the phylogenetic analyses. In fact, that the establishment of the genus Schlesneria renders Planctomyces paraphyletic was, surprisingly, already visible in the phylogenetic tree presented in [55], even though support for the non-monophyly of Planctomyces is stronger in our analyses (Figure 1). (Since Kulichevskaya et al.[55] did not mention the issue of taxa being monophyletic this also cannot be considered to be a pre-requisite for their interpretation, even though it is then not entirely clear why they conducted a phylogenetic analysis in the first place.) This picture did not change after R + Y coding of the 16S rRNA gene alignment [56], which still yielded 97% support for the sister group of P. limnophilus and S. paludicola (data not shown).
To measure this phylogenetic conflict in detail, we conducted both unconstrained heuristic searches for the best tree under the maximum likelihood (ML) [57] and maximum parsimony (MP) criterion [58] as well as searches constrained for the monophyly of all genera (for details of the data matrix see the caption of Figure 1). The best-known ML tree had a log likelihood of −9,214.93, whereas the best tree found under the constraint had a log likelihood of −9,249.16. The constrained tree was significantly worse than the globally best one in the Shimodaira-Hasegawa test as implemented in RAxML [57] (α = 0.01). The best-known MP tree had a score of 1,519, whereas the best constrained trees found had a score of 1,541 and were significantly worse in the Kishino-Hasegawa test as implemented in PAUP* [58] (α = 0.05). (See, e.g. chapter 21 in [59] for an in-depth description of such paired-site tests.) Accordingly, the current classification within the family as used in, e.g., [54] is in significant conflict with the 16S rRNA gene data.
In the following we will assess whether these 16S rRNA gene results can be confirmed with genomic data.
Phylogenomic analysis and comparative genomics
For the phylogenomic analysis, protein sequences from all available Planctomycetaceae and outgroup (Phycisphaera mikurensis) genomes were retrieved from NCBI or IMG (Table 5).
Table 5.
Genomic G + C content of the Planctomycetaceae type strains, including Phycisphaera mikurensis as outgroup
| G + C content from species description [%] | G + C content calculated from genome sequence [%] | NCBI accessions/IMG taxon IDs (number of contigs) | |
|---|---|---|---|
|
Blastopirellula marina DSM 3645T |
53.6 - 57.4 [60] |
57.04* |
AANZ00000000 (67); NZ_AANZ00000000 (4)* |
|
Gemmata obscuriglobus UQM 2246T |
64.4 ± 1.0 [61] |
67.18 |
ABGO00000000 (923) |
|
Isosphaera pallida ATCC 43644T |
62.2 [62] |
62.49 [20] |
CP002353, CP002354 |
|
Phycisphaera mikurensis NBRC 102666T |
73.0 [63] |
73.22 |
AP012338, AP012339 |
|
Pirellula staleyi DSM 6068T |
56.4 ± 0.4 - 57.4 ± 0.3 [64] |
57.46 [21] |
CP001848 |
|
Planctomyces brasiliensis DSM 5305T |
55.1 - 57.7 [1] |
56.45 |
CP002546 |
|
Planctomyces limnophilus DSM 3776T |
53.24 ± 0.59 [2] |
53.68 [19] |
CP001744, CP001745 |
|
Planctomyces maris DSM 8797T |
50.5 [65] |
50.45 |
ABCE00000000 (126) |
|
Rhodopirellula baltica SH 1T |
55.0 [66] |
55.40 [67] |
BX119912 |
|
Schlesneria paludicola DSM 18645T |
56.3 [55] |
55.66* |
AHZR00000000 (112); NZ_AHZR00000000 (24)* |
|
Singulisphaera acidiphila DSM 18658T |
59.9 [67] |
62.23* |
AGRX00000000 (113); CP003364 - CP003367* |
| Zavarzinella formosa DSM 19928T | 62.5 [68] | 59.10* | IMG ID: 2548877000 (594); NZ_AIAB00000000 (106)* |
*Denote G + C content values calculated from the updated version of the genome sequence, which is also marked with an asterisk.
The G + C content calculated from the genome sequence of G. obscuriglobus was 67.18%, whereas the previously published value, determined using traditional techniques, is 64.4 ± 1.0% [61]. Similarly, the G + C content of S. acidiphila was given as 59.9% [66] and that of Z. formosa was given as 62.5% [68], whereas the analysis of the genome sequences yields 62.23% and 59.10%, respectively. The three strongly deviating G + C values discovered here were all obtained using thermal denaturation [61,67,68]. A recent study [69] has shown that when calculated from genome sequences the G + C content varies at most 1% within species (which holds even though no attempt was made to remove plasmid sequences) and that larger variances are caused by the inaccuracies of the traditional techniques. It has thus been recommended to conduct emendations of species descriptions in the case of discrepancies larger than 1%, and to also conduct emendations of genus descriptions if the species emendations yield values that do not fit into the range of the G + C content given in the literature for the respective genus [69].
The G + C content of the Planctomycetaceae, using exact calculations from the genome sequences where possible, ranges from 50% to 70% (Table 5 and [70,71]), similar to the range between 52% and 69% measured by Schlesner et al. using traditional methods [66]. However, the description of the order Planctomycetales as well as the family Planctomycetaceae by Schlesner and Stackebrandt 1986 [27], who could at that time only consider the Pirellula staleyi, Pirellula marina (formerly Pirella), Planctomyces maris and Planctomyces limnophilus, refers to a G + C content of 50 to 59%. Planctomyces and Schlesneria have a comparatively low G + C content between 50% and 56%, with P. brasiliensis and P. maris differing more strongly from each other than P. limnophilus and S. paludicola. Similarly, Schlesner et al. reported a G + C content of 50 to 58% for Planctomyces strains [66]. The G + C content of the group comprising Blastopirellula, Pirellula and Rhodopirellula ranges from 55% to 57%. Although the G + C content of Zavarzinella measured from the genome sequence is lower than the one given in the species description, the group containing Aquisphaera, Gemmata, Isosphaera, Singulisphaera, Telmatocola and Zavarzinella possesses a higher G + C content than the other Planctomycetaceae, between 59% and 70%.
The genome sequences (Table 5) were phylogenetically investigated using the DSMZ phylogenomics pipeline as previously described [72-75] using NCBI BLAST [76], TribeMCL [77], OrthoMCL [78], MUSCLE [79], RASCAL [80], GBLOCKS [81] and MARE [82] to generate gene- and ortholog-content matrices as well as concatenated alignments of distinct selections of genes. Maximum likelihood (ML) [83] and maximum-parsimony (MP) [84,85] trees were inferred from the data matrices with RAxML [57,86] and PAUP* [58], respectively, as previously described [72-75].
The MARE-filtered supermatrix ML tree is shown in Figure 4 together with ML and MP bootstrap support values from all phylogenomic analyses if larger than 60%. All trees, except for the core-genes supermatrix MP tree, were topologically identical. The MP tree inferred from the core-gene matrix showed a distinct grouping of the clade that contained Gemmata obscuriglobus and Zavarzinella formosa, i.e. as sister clade of all other taxa but the outgroup, with maximal bootstrap support. Given that the majority of analyses supports the topology in Figure 4, the different core-genes MP topology might be caused by long branch attraction between the outgroup and the clade comprising Gemmata and Zavarzinella.
Figure 4.

Phylogenetic tree inferred from the MARE-filtered supermatrix under the maximum likelihood (ML) criterion [82] and rooted with Phycisphaera mikurensis . The branches are scaled in terms of the expected number of substitutions per site. Numbers above the branches (from left to right) are bootstrapping support values [87] (if larger than 60%) from ML/MP MARE-filtered supermatrix; ML/MP unfiltered (full) supermatrix; ML/MP core-genes supermatrix; ML/MP gene-content matrix; ML/MP ortholog-content matrix. Values larger than 95% are shown in bold; dots indicate branches with maximum support under all settings.
The phylogenomic tree (Figure 4) is topologically identical to the 16S rRNA gene tree (Figure 1) except for the backbone of the trees, which showed no support in the 16S rRNA gene analysis. Furthermore, the phylogenomic analysis confirms with maximum support the 16S rRNA tree that P. limnophilus and S. paludicola are sister taxa and, thus, Planctomyces is paraphyletic. The genomic interrelationships between these four species are thus examined more closely in the following. A genome sequence-based phylogeny with the same branching order but less bootstrapping support (probably because a restricted set of genes was used) was recently reported by Gou et al. [88].
The genomes of the four genome-sequenced Planctomyces and Schlesneria species differ significantly in their size. The genomes of S. paludicola (8.6 Mb, 6,860 protein-coding genes) and P. maris (7.8 Mb, 6,480 protein-coding genes) are significantly larger in size than the genomes of P. brasiliensis (6.0 Mb, 4,811 protein-coding genes) and P. limnophilus (5.5 Mb, 4,304 protein-coding genes).
To estimate the overall similarity between the four Planctomycetaceae genomes the GGDC (Genome-to-Genome Distance Calculator) [89,90] was used. The system calculates the distances by comparing the genomes to obtain HSPs (high-scoring segment pairs) and interfering distances from a set of formulas (1, HSP length / total length; 2, identities / HSP length; 3, identities / total length). Table 6 shows the results of the pairwise comparison of the Planctomycetaceae species with formula 2, as this formula is robust against the use of draft genomes such as ABCE00000000 (P. maris) [91].
Table 6.
Pairwise comparison of the four Planctomycetaceae species using GGDC, formula 2 (DDH estimates based on identities / HSP length)*
| P. brasiliensis | P. limnophilus | P. maris | S. paludicola | |
|---|---|---|---|---|
|
P. brasiliensis |
100.00% |
18.60% ± 2.27 |
21.40% ± 2.34 |
20.90% ± 2.33 |
|
P. limnophilus |
18.60% ± 2.27 |
100.00% |
24.10% ± 2.39 |
23.20% ± 2.38 |
|
P. maris |
21.40% ± 2.34 |
24.10% ± 2.39 |
100.00% |
22.00% ± 2.35 |
| S. paludicola | 20.90% ± 2.33 | 23.20% ± 2.38 | 22.00% ± 2.35 | 100.00% |
*The confidence intervals indicate the inherent uncertainty in estimating DDH values from intergenomic distances based on models derived from empirical test data sets (which are always limited in size); see [91] for details. The distance formulas are explained in [89]; formula 2 is recommended, particularly for draft genomes.
This comparison of the genomes revealed that P. brasiliensis shows a slightly higher DDH estimate with P. maris, compared to those with P. limnophilus and Schlesneria paludicola. For Schlesneria paludicola, a higher DDH value was estimated with P. limnophilus, in contrast to the other genomes. These results are in accordance with the 16S rRNA (Figure 1) and phylogenomic analyses (Figure 4). However, given the confidence intervals (Table 6), the DDH estimates do not show significant differences.
The fraction of shared genes in the four genomes is shown in a Venn diagram (Figure 5). The numbers of pairwise shared genes were inferred from the TribeMCL analysis via counting the clusters of homologous genes shared by the genomes. 1,586 clusters of homologs are shared by all four Planctomycetaceae species.
Figure 5.

Venn diagram depicting the intersections of sets of homologous proteins of P. maris, P. brasiliensis, P. limnophilus and S. paludicola. Their cardinalities are given in parentheses; for the total number of proteins see Table 3 and the resources listed in Table 5. The Venn diagram was calculated with the corresponding R package [92].
P. brasiliensis shares 63.1% of its clusters with P. maris, but only 57.6% and 58.2% clusters with P. limnophilus and S. paludicola, respectively. Only 27.5% of the P. brasiliensis genes have no homologs in the other genomes. P. limnophilus shares 63.2% of its clusters with S. paludicola, whereas P. brasiliensis shares 58.2% and P. maris shares 50.4% clusters with S. paludicola.
P. brasiliensis shares many more clusters of homologous genes with P. maris than with the other genomes. Of the four compared genomes, P. limnophilus shares most clusters of homologous genes with S. paludicola. These results are in accordance with the 16S rRNA (Figure 1) and phylogenomic analyses (Figure 4) as well as the GGDC results (Table 6).
Jeske et al. [93] performed a comprehensive genome mining approach to identify secondary metabolite related genes or gene clusters within the Planctomycetes. Just as in the other studied Planctomycetes, a number of genes putatively related with the production of secondary metabolites were identified in P. brasiliensis, that is, bacteriocin, ectoine, lantipeptide, terpene and type 1 polyketide synthases [93]. The study revealed Planctomycetes as a rich source for small molecules that might ultimately lead to new antibiotics or drugs [93].
Phenotypic comparison
Kulichevskaya et al. [55] used fatty acids as one of the major characteristics that distinguish Schlesneria from the genus Planctomyces and thus included them in the genus description of Schlesneria. To assess whether there is any published evidence for distinguishing Planctomyces and Schlesneria on the basis of fatty acids as well as to study the reproducibility of the results, the fatty-acid measurements for Planctomycetes were comprehensively collected from the literature and re-evaluated (see Figure 6). Caution should however be exercised in the interpretation of the results since different methods have been used in different publications.
Figure 6.

Heat map of fatty-acid measurements (A[63], B [65], C [36], D [66], E [68], F [55], G [67], H [70], I [71], J [48]]) from the analyzed Planctomycetaceae (see above). The heat map was calculated with the R package opm [94] with arcsine-square root transformation and Ward clustering applied. Asterisks denote fatty-acid profiles whose sum deviates more than five percent from 100%.
The measurements from allegedly the same strains did not always cluster together. Blastopirellula marina, P. maris and Rhodopirellula baltica showed distinct groupings of their respective measurements. The sum of the B. marina measurements from Schlesner et al. [66] is substantially below 100%, whereas the measurements of P. maris from Kerger et al. [36] sum up to much less than 100%, indicating missing or overstated values (Figure 6, all strong deviations of the sum from 100% denoted with asterisks). Omitting the questionable data in this respect did not alter the relative positioning of the other measurements (data not shown). The differences between the R. baltica measurements from two studies [36,66] might be caused by the distinct growth conditions (temperature and medium). Except for these three species, the fatty-acid analyses of the Planctomycetes type strains appeared fairly reproducible (Figure 6).
The analysis of the fatty-acid measurements shows a group comprising P. brasiliensis, P. maris and S. paludicola (disregarding the incomplete P. maris data from [66]), whereas P. limnophilus does not cluster together with the other Planctomyces species. The analysis supports the earlier finding [55] that S. paludicola can be differentiated from P. limnophilus on the basis of the fatty acids. However, the analysis also shows that S. paludicola cannot be differentiated from P. brasiliensis and P. maris, whose fatty acids were not analyzed in [55]. Rather, the fatty-acid profile of P. limnophilus differs from that of the other Planctomyces species (and Schlesneria), hence the overall similarity in fatty-acid profiles does not entirely reflect the relationships inferred from 16S rRNA gene (Figure 1) and phylogenomic analyses (Figure 4) as well as the GGDC results (Table 6). Thus the evidence for distinguishing Planctomyces and Schlesneria on the basis of fatty acids given by Kulichevskaya et al. [55] cannot be confirmed once all three Planctomyces species with cultivated type strains are considered, since the published fatty-acid profile of S. paludicola cannot be differentiated from that of P. brasiliensis and P. maris.
Taxonomy of the Planctomyces-Schlesneria complex
Given the positioning of the Planctomyces and Schlesneria species in the phylogenetic trees (Figure 1, Figure 4), several taxonomic changes could be conducted to render all genera monophyletic. The taxonomy of the group is apparently hampered by the unavailability of a culture of the type species, P. bekefii, which was described 90 years ago and based on a few morphological characters [4]. In our view, the scarcity of published information on this species does not allow prediction of its phylogenetic position relative to the other species if a 16S rRNA gene sequence of P. bekefii could be obtained (Figure 1). For this reason, the only safeguard against the possibility that phylogenetic analysis of P. bekefii would keep Planctomyces monophyletic is to place all Planctomyces species with an already known 16S rRNA gene (or even genome) sequence into a genus of their own. Indeed, arguing against the removal of other Planctomyces species from the genus because the comparison with the type species using contemporary taxonomic methods is impossible would be illogical, because none of the other Planctomycetes genera with validly published names have been compared with P. bekefii using modern methods either. For instance, there is insufficient evidence against a sister-group relationship between P. bekefii and S. paludicola, which was described mainly based on the comparison with P. limnophilus[55].
Whether or not the three cultivated Planctomyces species and Schlesneria should be placed in one, two, three or four genera should also be assessed by comparing the divergence among the resulting taxa with those of others. For instance, it would be inconsistent to place all four species, or only two of them, in a single genus if less divergent groups within Planctomycetes existed that were nevertheless split into several genera. Divergence was measured directly from the phylogenetic trees (Figure 1, Figure 4) as the maximum subtree height (MaSH) of each subtree without its stem branch. This measure has the advantage that it is on the one hand equivalent to half the maximum pairwise (patristic) distance between organisms placed in a subtree of an ultrametric tree but on the other hand can also be applied if a phylogenetic tree is non-ultrametric, which is much more frequent (Figure 1, Figure 4). Indeed, pairwise distances can be taxonomically misleading because under non-ultrametric conditions less distant organisms need not be more closely related [38,59,69]. Using MaSH, the reference to subtrees guarantees by definition that only monophyletic groups are measured.
The results are shown in Table 7. In the 16S rRNA gene tree, the P. brasiliensis/maris and P. limnophilus/S. paludicola subtrees were among those subtrees with the smallest MaSH values, but these are more than twice as large as those for the two other single-genus subtrees (containing Blastopirellula and Singulisphaera, respectively). Further, the subtree containing Schlesneria and all Planctomyces species yielded a larger MaSH than a number of subtrees that contain up to three genera. In the ML trees inferred from genome-scale data, the Blastopirellula/Pirellula and the Gemmata/Zavarzinella subtrees showed in same cases larger MaSH values than the P. brasiliensis/maris and P. limnophilus/S. paludicola subtrees, but in other cases smaller MaSH values. These results indicate that there is no substantial difference between the divergence of the P. brasiliensis/maris group, the P. limnophilus/ S. paludicola group and Planctomycetes groups that comprise two genera. This argues against placing P. brasiliensis and P. maris in the same genus and against placing P. limnophilus/S. paludicola in the same genus, let alone placing all four species in a single genus.
Table 7.
MaSH values calculated for the ML trees inferred in this study
| Subtree | 16S rRNA gene | Full supermatrix | MARE supermatrix | Core-gene supermatrix | Gene content | Ortholog content |
|---|---|---|---|---|---|---|
|
S. acidiphila, S. rosea* |
0.027 |
~ |
~ |
~ |
~ |
~ |
|
B. cremea*, B. marina |
0.032 |
~ |
~ |
~ |
~ |
~ |
|
P. brasiliensis, P. maris |
0.072 |
0.414 |
0.272 |
0.287 |
0.166 |
0.185 |
|
P. limnophilus, Sc. |
0.074 |
0.382 |
0.240 |
0.245 |
0.193 |
0.215 |
|
Te.*, Za. |
0.086 |
~ |
~ |
~ |
~ |
~ |
|
Aq.*, Is. |
0.090 |
~ |
~ |
~ |
~ |
~ |
|
Aq.*, Is., S. acidiphila, S. rosea* |
0.100 |
0.422 |
0.305 |
0.309 |
0.177 |
0.209 |
|
B. cremea*, B. marina, Pi. |
0.101 |
0.378 |
0.272 |
0.267 |
0.164 |
0.171 |
|
Ge., Te.*, Za. |
0.119 |
0.346 |
0.280 |
0.275 |
0.213 |
0.236 |
|
Pl., Sc. |
0.121 |
0.480 |
0.338 |
0.333 |
0.198 |
0.220 |
|
B. cremea*, B. marina, Pi., Rh. |
0.125 |
0.539 |
0.353 |
0.382 |
0.379 |
0.347 |
|
Aq.*, Ge., Is., S. acidiphila, S. rosea*, Te.*, Za. |
0.245 |
0.633 |
0.579 |
0.522 |
0.381 |
0.448 |
|
Aq.*, Ge., Is., Pl., Sc., S. acidiphila, S. rosea*, Te.*, Za. |
0.297 |
0.658 |
0.523 |
0.522 |
0.410 |
0.382 |
|
Aq.*, B. cremea*, B. marina, Ge., Is., Pi., Pl., Rh., Sc., S. acidiphila, S. rosea*, Te.*, Za. |
0.308 |
0.734 |
0.682 |
0.588 |
0.427 |
0.495 |
| Aq.*, B. cremea*, B. marina, Ge., Is., Ph., Pi., Pl., Rh., Sc., S. acidiphila, S. rosea*, Te.*, Za. | 0.639 | 1.258 | 1.118 | 1.082 | 0.558 | 0.649 |
*Indicate organisms that were only present in the 16S rRNA gene tree.
Phenotypic differences between P. brasiliensis and P. maris on the one hand and P. limnophilus on the other hand also exist; for instance, the fatty-acid profiles of the former differ from that of the latter in the same way (Figure 6) as the S. paludicola profile differs from the one of P. limnophilus[55]. S. paludicola differs from all three Planctomyces species with respect to the number of flagella of the daughter cells [55]. Moreover, P. brasiliensis and P. maris contain sym-homospermidine as polyamine, whereas P. limnophilus contains putrescine instead (Table 8). P. maris differs from P. brasiliensis because only the latter contains spermidine (Table 8). Given that four Planctomycetes placed in distinct genera (Blastopirellula, Gemmata, Isosphaera, P. maris) possess the same polyamine pattern (Table 8) the differences between the Planctomyces species appear significant. Regarding polar lipids, P. maris contains phosphatidyl-monomethylethanolamine and phosphatidyl-dimethylethanolamine, which are not produced by P. brasiliensis (Table 8). Compared to Aquisphaera and Gemmata, between which no differences regarding their polar-lipid pattern have been reported [69,65], but which are placed in distinct genera, the differences between the Planctomyces species again appear significant.
Table 8.
Polyamines[96 ] and polar lipids [48,65,70,95] for several Planctomycetaceae as reported in the literature
| |
Polyamines |
Polar lipids
1
|
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cadaverine | Putrescine | Spermidine | sym -Homospermidine | DPG | Glyl | PC | DPE | PG | MPE | |
|
A. giovannonii[70] |
NR2 |
NR |
NR |
NR |
+ |
- |
+ |
- |
+ |
- |
|
B. cremea[95] |
NR |
NR |
NR |
NR |
- |
- |
- |
- |
+ |
- |
|
B. marina[40,65,66] |
- |
- |
- |
+ |
+ |
- |
- |
- |
+ |
- |
|
G. obscuriglobus[40,65] |
- |
- |
- |
+ |
+ |
- |
+ |
- |
+ |
- |
|
I. pallida[40] |
- |
- |
- |
+ |
NR |
NR |
NR |
NR |
NR |
NR |
|
P. staleyi[40,65,66] |
- |
- |
+ |
+ |
+ |
- |
- |
- |
+ |
+ |
|
P. brasiliensis[40,65] |
- |
- |
+ |
+ |
+ |
+ |
- |
- |
+ |
- |
|
P. limnophilus[40,65] |
- |
+ |
+ |
- |
+ |
+ |
- |
- |
+ |
- |
|
P. maris[40,65] |
- |
- |
- |
+ |
+ |
+ |
+ |
+ |
+ |
+ |
|
R. baltica[40,65,66] |
+ |
+ |
- |
+ |
+ |
+ |
+ |
+ |
+ |
- |
|
S. acidiphila[48] |
NR |
NR |
NR |
NR |
- |
- |
+ |
- |
+ |
- |
| S. rosea[48] | NR | NR | NR | NR | - | - | + | - | + | - |
1Phospholipids: DPG Diphosphatidylglygerol; Glyl Glycolipid; PC Phosphatidylcholine; DPE Phosphatidyl-dimethylethanolamine; PG Phosphatidylglycerol; MPE Phosphatidyl-monomethylethanolamine.
2Not reported in the literature.
We conclude that both genomic and phenotypic analyses according to both qualitative and quantitative criteria support the placement of the three Planctomyces species with cultivated type strains into genera separate from each other and from Schlesneria.
Taxonomic consequences
As explained in detail above, the differences in the reported G + C contents of G. obscuriglobus, S. acidiphila and Z. formosa from the ones calculated from their genome sequences justify an emendation of their species descriptions and in the case of Gemmata an emendation of the genus description (which referred to the G + C content).
Moreover, the range of G + C content values determined for the genera now placed within Planctomycetaceae (Planctomycetales) is in conflict with the description of the family (and order) as given by Schlesner and Stackebrandt 1986 [27]. In addition, an optimal growth temperature of 41°C is reported for Isosphaera[55], which is considerably higher than the one reported in the description of the family (and order). For Schlesneria, Singulisphaera, Telmatocola and Zavarzinella a slightly lower optimal growth temperature is reported, i.e. 20°C upward [48,55,67,68,71]. Furthermore, for the genera Schlesneria and Zavarzinella it was reported that the daughter cells are motile by means of two [55] or one to two flagella [68], respectively, whereas the description of the family (and order) only considered the presence of one flagellum.
Whereas these discrepancies call for an emendation of the family Planctomycetaceae, phylogenomic (Figure 4) and 16S rRNA gene analyses (Figure 1) show a monophyletic group comprising the genera with high G + C content. The remaining genera appear unresolved in the 16S rRNA gene tree (Figure 1) but as a maximally supported clade comprising the genera Planctomyces, Schlesneria, Blastopirellula, Pirellula and Rhodopirellula in the phylogenomic tree (Figure 4). This would support a splitting of the quite heterogeneous Planctomycetaceae into two families, but the unknown position of the type species, P. bekefii, of the type genus means that a more narrowly defined Planctomycetaceae may not include P. bekefii. For this reason, we refrain from generating a novel family to accommodate the high-G + C genera.
Emended description of the genus Gemmata Franzmann and Skerman (1985)
The description is the one given by Franzmann and Skerman [61] with the following modification.
The G + C content is about 67%.
Emended description of the species Gemmata obscuriglobus Franzmann and Skerman (1985)
The description of the species Gemmata obscuriglobus is the one given by Franzmann and Skerman [61], with the following modification.
The G + C content is 67%.
Emended description of the species Singulisphaera acidiphila Kulichevskaya et al. (2008)
The description of the species Singulisphaera acidiphila is the one given by Kulichevskaya et al. (2008) [67], with the following modification.
The G + C content is 62%.
Emended description of the species Zavarzinella formosa Kulichevskaya et al. 2009
The description of the species Zavarzinella formosa is the one given by Kulichevskaya et al. (2009) [68], with the following modification.
The G + C content is 59%.
Emended description of the order Planctomycetales Schlesner and Stackebrandt 1986
The description of the order Planctomycetales is the one given by Schlesner and Stackebrandt [27], with the following modification.
Daughter cells may be motile by means of one or two flagella. The G + C content ranges between about 50% and about 70%. The growth temperature optimum ranges between 20°C and 40°C.
Emended description of the family Planctomycetaceae Schlesner and Stackebrandt 1986
The description of the family Planctomycetaceae is the one given by Schlesner and Stackebrandt [27], with the following modification.
Daughter cells may be motile by means of one or two flagella. The G + C content ranges between about 50% and about 70%. The growth temperature optimum ranges approximately between 20°C and 40°C.
Description of Rubinisphaera gen. nov
Rubinisphaera (Ru.bi.ni.sphaer’a N. L. fem. n. Rubinia named in honor of Edward Rubin, an American geneticist and medical researcher, who played a key role in initiating the Genomic Encyclopedia of Archaea and Bacteria project at the DOE Joint Genome Institute (JGI); NL fem. n. sphaerae, a ball, globe, sphere; N. L. fem. n. Rubinisphaera, a spherical organism named after Edward Rubin).
The features are the ones of the type species. Characteristic polyamines are sym-homospermidine and spermidine. The polar lipids phosphatidyl-monomethylethanolamine and phosphatidyl-dimethylethanolamine are absent. Major fatty acids are C16:0 and C16:1 ω7c. The G + C content is about 56%.
The type species is Rubinisphaera brasiliensis. Rubinisphaera belongs to Planctomycetaceae.
Description of Rubinisphaera brasiliensis (Schlesner 1990) comb. nov.
Rubinisphaera brasiliensis (bra.si.lien’sis L. m. adj. pertaining to the country of Brazil).
Basonym: Planctomyces brasiliensis Schlesner 1990.
The genus Rubinispharea is comprised of one species, Rubinisphaera brasiliensis. The characteristics of the species are given in the genus description and the description given by Schlesner (1989) [1].
The type strain is IFAM 1448T (=ATCC 49424 = DSM 5305 = JCM 21570 = NBRC 103401).
Description of Planctopirus gen. nov.
Planctopirus (Planc.to.pi’rus Gr. adj. planktos, wandering, floating; L. maS. n. pirus, a pear tree, referring to the shape of the cells, which also occur in bundles; NL fem. n. Planctopirus, floating pear tree).
The features are the ones of the type species. Characteristic polyamines are putrescine and spermidine. Major fatty acids are C16:0, C16:1 ω7c and C18:1 ω9c. The G + C content is about 54%.
The type species is Planctopirus limnophilus. Planctopirus belongs to Planctomycetaceae.
Description of Planctopirus limnophilus (Hirsch and Müller 1986) comb. nov.
Planctopirus limnophilus (lim.no’phi.la Gr. n. limnos lake; Gr. adj. philus loving; NL adj. limnophila lake loving).
Basonym: Planctomyces limnophilus Hirsch and Müller 1986.
The genus Planctopirus is comprised of one species, Planctopirus limnophilus. The characteristics of the species are given in the genus description and the description given by Hirsch and Müller (1986) [2].
The type strain is Mü 290T (=ATCC 43296 = DSM 3776 = IFAM 1008).
Description of Gimesia gen. nov.
Gimesia (Gi.me’si.a N. L. fem. n. Gimesia named in honor of Nándor István Gimesi, a Hungarian plant physiologist and plant morphologist, who first reported organisms now placed in the phylum Planctomycetes).
The features are the ones of the type species. The only major polyamine component is sym-homospermidine. The polar lipids contain phosphatidyl-monomethylethanolamine and phosphatidyl-dimethylethanolamine. Major fatty acids are C16:0 and C16:1 ω7c. The G + C content is about 50%.
The type species is Gimesia maris. Gimesia belongs to Planctomycetaceae.
Description of Gimesia maris (ex Bauld and Staley 1976) comb. nov.
Gimesia maris (mar’is L. gen. noun maris of the sea).
Basonym: Planctomyces maris (ex Bauld and Staley 1976) Bauld and Staley 1980.
The genus Gimesia is comprised of one species Gimesia maris. The characteristics of the species are given in the genus description and the description given by Bauld and Staley (1976) [97] and Bauld and Staley (1980) [3].
The type strain is 534-30T (=ATCC 29201 = DSM 8797).
Conclusion
This study presents the genome sequence for the P. brasiliensis type strain IFAM 1448T, whose physiological and genomic features are reviewed in detail. Results from phylogenomic analyses including all available Planctomycetaceae genomes disagree with the present circumscription of the genera Planctomyces and Schlesneria. The revisited 16S rRNA gene and phenotypic data from the literature neither support the current classification. A quantitative comparison of phylogenetic and phenotypic aspects suggest the formation of three new genera (for which we propose the names Gimesia, Planctopirus and Rubinisphaera) to accommodate P. maris, P. limnophilus and P. brasiliensis, respectively. Considerable differences between the reported G + C content of Gemmata obscuriglobus, Singulisphaera acidiphila and Zavarzinella formosa and G + C content calculated from their genome sequences are found, suggesting the emendation of their species descriptions. The range of G + C values reported for the Planctomycetaceae (Planctomycetales) indicate that the descriptions of the family and the order should be emended.
Competing interests
The authors declare that they have no competing interests.
Author's contributions
CS, BA and MG conducted the phylogenomic studies. CS, BJT, RM, BA, PH, MG and HPK drafted the manuscript. MR performed the laboratory experiments. ML, MN, AL, JFC, LG, SP, MH, KL, IP, KM, NI, AP, AC, KP, CDJ, LH, ML, JCD, TW, JAE, VM and NCK sequenced, assembled and annotated the genome. All authors read and approved the final manuscript.
Contributor Information
Carmen Scheuner, Email: carmen.scheuner@dsmz.de.
Brian J Tindall, Email: brian.tindall@dsmz.de.
Megan Lu, Email: mlu@lbl.gov.
Matt Nolan, Email: mnolan@lbl.gov.
Alla Lapidus, Email: alapidus@lbl.gov.
Jan-Fang Cheng, Email: jfcheng@lbl.gov.
Lynne Goodwin, Email: lgoodwin@lbl.gov.
Sam Pitluck, Email: spitluck@lbl.gov.
Marcel Huntemann, Email: mhuntemann@lbl.gov.
Konstantinos Liolios, Email: kliolios@lbl.gov.
Ioanna Pagani, Email: ipagani@lbl.gov.
Konstantinos Mavromatis, Email: kmavromatis@lbl.gov.
Natalia Ivanova, Email: nivanova@lbl.gov.
Amrita Pati, Email: apati@lbl.gov.
Amy Chen, Email: achen@lbl.gov.
Krishna Palaniappan, Email: kpalaniappan@lbl.gov.
Cynthia D Jeffries, Email: cdjeffries@lbl.gov.
Loren Hauser, Email: lhauser@lbl.gov.
Miriam Land, Email: mland@lbl.gov.
Romano Mwirichia, Email: rkachiuru2000@yahoo.com.
Manfred Rohde, Email: manfred.rohde@helmholtz-hzi.de.
Birte Abt, Email: birte.abt@dsmz.de.
John C Detter, Email: jcdetter@lbl.gov.
Tanja Woyke, Email: twoyke@lbl.gov.
Jonathan A Eisen, Email: jaeisen@lbl.gov.
Victor Markowitz, Email: vmarkowitz@lbl.gov.
Philip Hugenholtz, Email: phugenholtz@lbl.gov.
Markus Göker, Email: markus.goeker@dsmz.de.
Nikos C Kyrpides, Email: nckyrpides@lbl.gov.
Hans-Peter Klenk, Email: hans-peter.klenk@dsmz.de.
Acknowledgements
We gratefully acknowledge the help of Helga Pomrenke for growing P. brasiliensis cultures and Evelyne-Marie Brambilla for DNA extraction and quality control (both at DSMZ). We are grateful to Erko Stackebrandt and Christian Jogler (both at DSMZ) for hints regarding Planctomycetes classification and to Edina Weidemann for information regarding the biography of N.I. Gimesi. This work was performed under the auspices of the US Department of Energy Office of Science, Biological and Environmental Research Program, and by the University of California, Lawrence Berkeley National Laboratory under contract No. DE-AC02-05CH11231, Lawrence Livermore National Laboratory under Contract No. DE-AC52-07NA27344, and Los Alamos National Laboratory under contract No. DE-AC02-06NA25396, UT-Battelle and Oak Ridge National Laboratory under contract DE-AC05-00OR22725, as well as German Research Foundation (DFG) INST 599/1-2.
References
- Schlesner H. Planctomyces brasiliensis sp. nov., a halotolerant bacterium from a salt pit. Syst Appl Microbiol. 1989;12:159–61. doi: 10.1016/S0723-2020(89)80008-6. http://dx.doi.org/10.1016/S0723-2020(89)80008-6. [DOI] [Google Scholar]
- Hirsch P, Müller M. Planctomyces limnophilus sp. nov., a stalked and budding bacterium from freshwater. Syst Appl Microbiol. 1985;6:276–80. doi: 10.1016/S0723-2020(85)80031-X. http://dx.doi.org/10.1016/S0723-2020(85)80031-X. [DOI] [Google Scholar]
- Bauld J, Staley JT. Planctomyces maris sp. nov., nom. rev. Int J Syst Bacteriol. 1980;30:657. doi: 10.1099/00207713-30-4-657. http://dx.doi.org/10.1099/00207713-30-4-657. [DOI] [Google Scholar]
- Gimesi N. Hydrobiologiai Tanulmanyok (Hydrobiologische Studien). I: Planctomyces bekefii Gim. nov. gen. et sp. [in Hungarian, with German translation]. Kiadja a Magyar Ciszterci Rend. Budapest, Hungary: Kiadja a Magyar Ciszterci Rend; 1924. pp. 1–8. [Google Scholar]
- Starr MP, Schmidt JM. Planctomyces stranskae (ex Wawrik 1952) sp. nov., nom. rev. and Planctomyces guttaeformis (ex Hortobagyi 1965) sp. nov., nom. rev. Int J Syst Bacteriol. 1984;34:470–7. doi: 10.1099/00207713-34-4-470. http://dx.doi.org/10.1099/00207713-34-4-470. [DOI] [Google Scholar]
- Ward N, Staley JT, Fuerst JA, Giovannoni S, Schlesner H, Stackebrandt E. The order Planctomycetales, including the genera Planctomyces, Pirellula, Gemmata and Isosphaera and the Candidatus genera Brocadia, Kuenenia and Scalindua . 3. Berlin, Germany: Springer-Verlag; 2006. [Google Scholar]
- Langó Z. Who has first observed Planctomyces. Acta Microbiol Immunol Hung. 2005;52:73–84. doi: 10.1556/AMicr.52.2005.1.3. http://dx.doi.org/10.1556/AMicr.52.2005.1.3. [DOI] [PubMed] [Google Scholar]
- Fuerst J, Gwilliam H, Lindsay M, Lichanska A, Belcher C, Vickers J, Hugenholtz P. Isolation and molecular identification of planctomycete bacteria from postlarvae of the giant tiger prawn, Penaeus monodon. Appl Environ Microbiol. 1997;63:254–62. doi: 10.1128/aem.63.1.254-262.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butler MK, Fuerst JA. Comparative analysis of ribonuclease P RNA of the planctomycetes. Int J Syst Evol Microbiol. 2004;54:1333–44. doi: 10.1099/ijs.0.03013-0. http://dx.doi.org/0.1099/ijs.0.03013-0. [DOI] [PubMed] [Google Scholar]
- Lindsay MR, Webb RI, Strous M, Jetten MS, Butler MK, Forde RJ, Fuerst JA. Cell compartmentalisation in planctomycetes: novel types of structural organisation for the bacterial cell. Arch Microbiol. 2001;175:413–29. doi: 10.1007/s002030100280. http://dx.doi.org/10.1007/s002030100280. [DOI] [PubMed] [Google Scholar]
- Fuerst JA. The PVC, superphylum: exceptions to the bacterial definition? Antonie Van Leeuwenhoek. 2013;104:451–66. doi: 10.1007/s10482-013-9986-1. http://dx.doi.org/10.1007/s10482-013-9986-1. [DOI] [PubMed] [Google Scholar]
- Devos DP. Re-interpretation of the evidence for the PVC cell plan supports a Gram-negative origin. Antonie Van Leeuwenhoek. 2014;105:271–4. doi: 10.1007/s10482-013-0087-y. http://dx.doi.org/10.1007/s10482-013-0087-y. [DOI] [PubMed] [Google Scholar]
- Jogler C, Glöckner FO, Kolter R. Characterization of Planctomyces limnophilus and development of genetic tools for its manipulation establish it as a model species for the phylum Planctomycetes. Appl Environ Microbiol. 2011;77:5826–9. doi: 10.1128/AEM.05132-11. http://dx.doi.org/10.1128/AEM.05132-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schreier HJ, Dejtisakdi W, Escalante JO, Brailo M. Transposon mutagenesis of Planctomyces limnophilus and analysis of a pckA mutant. Appl Environ Microbiol. 2012;78:7120–3. doi: 10.1128/AEM.01794-12. http://dx.doi.org/10.1128/AEM.01794-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klenk HP, Lapidus A, Chertkov O, Copeland A, Glavina del Rio T, Nolan M, Lucas S, Chen F, Tice H, Cheng JF, Han C, Bruce D, Goodwin LA, Pitluck S, Pati A, Ivanova N, Mavromatis K, Daum C, Chen A, Palaniappan K, Land M, Hauser L, Jeffries CD, Detter JC, Rohde M, Abt B, Pukall R, Göker M, Bristow J, Markowitz V. et al. Complete genome sequence of the thermophilic, hydrogen-oxydizing Bacillus tusciae type strain (T2T) and reclassification in the new genus, Kyrpidia gen. nov. as Kyrpidia tusciae comb. nov. and emendation of the family Alicyclobacillaceae da Costa and Rainey, 2010. Stand Genomic Sci. 2011;5:97–111. doi: 10.4056/sigs.2114901. http://dx.doi.org/10.4056/sigs.2144922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glatz RE, Lepp PW, Ward BB, Francis CA. Planktonic microbial community composition across steep physical/chemical gradients in permanently ice-covered Lake Bonney. Antarctica Geobiology. 2006;4:53–67. doi: 10.1111/j.1472-4669.2006.00057.x. http://dx.doi.org/10.1111/j.1472-4669.2006.00057.x. [DOI] [Google Scholar]
- Hess PN, De Moraes Russo CA. An empirical test of the midpoint rooting method. Biol J Linn Soc Lond. 2007;92:669–74. doi: 10.1111/j.1095-8312.2007.00864.x. http://dx.doi.org/10.1111/j.1095-8312.2007.00864.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pagani I, Liolios K, Jansson J, Chen IM, Smirnova T, Nosrat B, Markowitz VM, Kyrpides NC. The Genomes OnLine Database (GOLD) v. 4: status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Res. 2012;40:D571–9. doi: 10.1093/nar/gkr1100. http://dx.doi.org/10.1093/nar/gkr1100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LaButti K, Sikorski J, Schneider S, Nolan M, Lucas S, Rio TGD, Tice H, Cheng JF, Goodwin L, Pitluck S, Liolios K, Ivanova N, Mavromatis K, Mikhailova N, Pati A, Chen A, Palaniappan K, Land M, Hauser L, Chang YJ, Jeffries CD, Tindall BJ, Rohde M, Göker M, Woyke T, Bristow J, Eisen JA, Markowitz V, Hugenholtz P, Kyrpides NC. et al. Complete genome sequence of Planctomyces limnophilus type strain (Mü 290T) Stand Genomic Sci. 2010;3:47–56. doi: 10.4056/sigs.1052813. http://dx.doi.org/10.4056/sigs.1052813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Göker M, Cleland D, Saunders E, Lapidus A, Nolan M, Lucas S, Hammon N, Deshpande S, Cheng JF, Tapia R, Han C, Goodwin L, Pitluck S, Liolios K, Pagani I, Ivanova N, Mavromatis K, Pati A, Chen A, Palaniappan K, Land M, Hauser L, Chang YJ, Jeffries CD, Detter JC, Beck B, Woyke T, Bristow J, Eisen JA, Markowitz V. et al. Complete genome sequence of Isosphaera pallida type strain (IS1BT) Stand Genomic Sci. 2011;4:63–71. doi: 10.4056/sigs.1533840. http://dx.doi.org/10.4056/sigs.1533840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clum A, Tindall BJ, Sikorski J, Lucas S, Glavina del Rio T, Nolan M, Chen F, Tice H, Pitluck S, Cheng JF, Chertkov O, Brettin T, Han C, Detter JC, Kuske C, Bruce D, Goodwin L, Ovchinnikova G, Pati A, Mikhailova N, Chen A, Palaniappan K, Land M, Hauser L, Chang YJ, Jeffries CD, Chain P, Rohde M. et al. Complete genome sequence of Pirellula staleyi type strain (ATCC 27377T) Stand Genomic Sci. 2009;1:308–16. doi: 10.4056/sigs.51657. http://dx.doi.org/10.4056/sigs.51657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wegner CE, Richter-Heitmann T, Klindworth A, Richter M, Achstetter T, Glöckner FO, Harder J. Expression of sulfatases in Rhodopirellula baltica and diversity of sulfatases in the genus Rhodospirellula. Mar Genomics. 2013;9:51–61. doi: 10.1016/j.margen.2012.12.001. http://dx.doi.org/10.1016/j.margen.2012.12.001. [DOI] [PubMed] [Google Scholar]
- Field D, Garrity G, Gray T, Morrison N, Selengut J, Sterk P, Tatusova T, Thomson N, Allen MJ, Angiuoli SV, Ashburner M, Axelrod N, Baldauf S, Ballard S, Boore J, Cochrane G, Cole J, Dawyndt P, De Vos P, dePamphilis C, Edwards R, Faruque N, Feldman R, Gilbert J, Gilna P, Glöckner FO, Goldstein P, Guralnick R, Haft D, Hancock D. et al. The minimum information about a genome sequence (MIGS) specification. Nat Biotechnol. 2008;26:541–7. doi: 10.1038/nbt1360. http://dx.doi.org/10.1038/nbt1360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Field D, Amaral-Zettler L, Cochrane G, Cole JR, Dawyndt P, Garrity GM, Gilbert J, Glöckner FO, Hirschman L, Karsch-Mzrachi I, Klenk HP, Knight R, Kottmann R, Kyrpides N, Meyer F, San Gil I, Sansone SA, Schriml LM, Sterk P, Tatusova T, Ussery DW, White O, Wooley J. Clarifying concepts and terms in biodiversity informatics. PLoS Biol. 2011;9:e1001088. doi: 10.1371/journal.pbio.1001088. http://dx.doi.org/10.1371/journal.pbio.1001088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woese CR, Kandler O, Wheelis ML. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc Natl Acad Sci U S A. 1990;87:4576–9. doi: 10.1073/pnas.87.12.4576. http://dx.doi.org/10.1073/pnas.87.12.4576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrity GM, Holt JG. The Road Map to the Manual, Volume 1. 2. New York: Springer; 2001. pp. 119–66. [Google Scholar]
- Schlesner H, Stackebrandt E. Assignment of the genera Planctomyces and Pirella to a new family Planctomycetaceae fam. nov. and description of the order Planctomycetales ord. nov. Syst Appl Microbiol. 1986;8:174–6. doi: 10.1016/S0723-2020(86)80072-8. http://dx.doi.org/10.1016/S0723-2020(86)80072-8. [DOI] [Google Scholar]
- Judicial Commission of the International Committee on Systematics of Prokaryotes. The nomenclatural types of the orders Acholeplasmatales, Halanaerobiales, Halobacteriales, Methanobacteriales, Methanococcales, Methanomicrobiales, Planctomycetales, Prochlorales, Sulfolobales, Thermococcales, Thermoproteales and Verrucomicrobiales are the genera Acholeplasma, Halanaerobium, Halobacterium, Methanobacterium, Methanococcus, Methanomicrobium, Planctomyces, Prochloron, Sulfolobus, Thermococcus, Thermoproteus and Verrucomicrobium, respectively. Opinion 79. Int J Syst Evol Microbiol. 2005;55:517–8. doi: 10.1099/ijs.0.63548-0. http://dx.doi.org/10.1099/ijs.0.63548-0. [DOI] [PubMed] [Google Scholar]
- Validation of the publication of new names and new combinations previously effectively published outside the IJSB. Int J Syst Bacteriol. 1987;37:179–80. doi: 10.1099/00207713-46-2-625. Validation List no. 23. http://dx.doi.org/10.1099/00207713-37-2-179. [DOI] [PubMed] [Google Scholar]
- Skerman VBD, McGowan V, Sneath PHA. Approved lists of bacterial names. Int J Syst Bacteriol. 1980;30:225–420. doi: 10.1099/00207713-30-1-225. http://dx.doi.org/10.1099/00207713-30-1-225. [DOI] [PubMed] [Google Scholar]
- Hirsch P, Skuja HL. In: Bergey’s Manual of Determinative Bacteriology. 8. Buchanan RE, Gibbons NE, editor. Baltimore: The Williams and Wilkins Co; 1974. Genus Planctomyces Gimesi 1924, 4; pp. 162–3. [Google Scholar]
- Validation of the publication of new names and new combinations previously effectively published outside the IJSB. Int J Syst Bacteriol. 1990;40:105–6. doi: 10.1099/00207713-46-2-625. Validation List no. 32. http://dx.doi.org/10.1099/00207713-40-1-105. [DOI] [PubMed] [Google Scholar]
- Classification of bacteria and archaea in risk groups. p. 170. http://www.baue.de/ TRBA 466.
- Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M, Rubin GM, Sherlock G. Gene Ontology: tool for the unification of biology. Nat Genet. 2000;25:25–9. doi: 10.1038/75556. http://dx.doi.org/10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liesack W, König H, Schlesner H, Hirsch P. Chemical composition of the peptidoglycan-free cell envelopes of budding bacteria of the Pirelulla/Planctomyces group. Arch Microbiol. 1986;145:361–6. doi: 10.1007/BF00470872. http://dx.doi.org/10.1007/BF00470872. [DOI] [Google Scholar]
- Kerger BD, Mancuso CA, Nichols PD, White DC, Langworthy T, Sittig M, Schlesner H, Hirsch P. The budding bacteria, Pirellula and Planctomyces, with atypical 16S rRNA and absence of peptidoglycan, show eubacterial phospholipids and uniquely high proportions of long chain beta-hydroxy fatty acids in the lipopolysaccharide lipid A. Arch Microbiol. 1988;149:255–60. doi: 10.1007/BF00422014. http://dx.doi.org/10.1007/BF00422014. [DOI] [Google Scholar]
- Klenk HP, Göker M. En route to a genome-based classification of Archaea and Bacteria? Syst Appl Microbiol. 2010;33:175–82. doi: 10.1016/j.syapm.2010.03.003. http://dx.doi.org/10.1016/j.syapm.2010.03.003. [DOI] [PubMed] [Google Scholar]
- Göker M, Klenk HP. Phylogeny-driven target selection for large scale genome-sequencing (and other) projects. Stand Genomic Sci. 2013;8:360–74. doi: 10.4056/sigs.3446951. http://dx.doi.org/10.4056/sigs.3446951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu D, Hugenholtz P, Mavromatis K, Pukall R, Dalin E, Ivanova NN, Kunin V, Goodwin L, Wu M, Tindall BJ, Hooper SD, Pati A, Lykidis A, Spring S, Anderson IJ, D’haeseleer P, Zemla A, Singer M, Lapidus A, Nolan M, Copeland A, Han C, Chen F, Cheng JF, Lucas S, Kerfeld C, Lang E, Gronow S, Chain P, Bruce D. et al. A phylogeny-driven genomic encyclopaedia of Bacteria and Archaea. Nature. 2009;462:1056–60. doi: 10.1038/nature08656. http://dx.doi.org/10.1038/nature08656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mavromatis K, Land ML, Brettin TS, Quest DJ, Copeland A, Clum A, Goodwin L, Woyke T, Lapidus A, Klenk HP, Cottingham RW, Kyrpides NC. The fast changing landscape of sequencing technologies and their impact on microbial genome assemblies and annotation. PLoS ONE. 2012;7:e48837. doi: 10.1371/journal.pone.0048837. http://dx.doi.org/10.1371/journal.pone.0048837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- List of growth media used at DSMZ. http://www.dsmz.de/
- Gemeinholzer B, Dröge G, Zetzsche H, Haszprunar G, Klenk HP, Güntsch A, Berendsohn WG, The WJW. DNA. Bank Network: the start from a German initiative. Biopreserv Biobank. 2011;9:51–5. doi: 10.1089/bio.2010.0029. http://dx.doi.org/10.1089/bio.2010.0029. [DOI] [PubMed] [Google Scholar]
- The DOE Joint Genome Institute. http://www.jgi.doe.gov/
- Zerbino DR, Birney E. Velvet: algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008;18:821–9. doi: 10.1101/gr.074492.107. http://dx.doi.org/10.1101/gr.074492.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sims D, Brettin T, Detter JC, Han C, Lapidus A, Copeland A, Glavina Del Rio T, Nolan M, Chen F, Lucas S, Tice H, Cheng JF, Bruce D, Goodwin L, Pitluck S, Ovchinnikova G, Pati A, Ivanova N, Mavromatis K, Chen A, Palaniappan K, D'haeseleer P, Chain P, Bristow J, Eisen JA, Markowitz V, Hugenholtz P, Schneider S, Göker M, Pukall R. et al. Complete genome sequence of Kytococcus sedentarius type strain (541T) Stand Genomic Sci. 2009;1:12–20. doi: 10.4056/sigs.761. http://dx.doi.org/10.4056/sigs.761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lapidus A, LaButti K, Foster B, Lowry S, Trong S, Goltsman E. POLISHER: An Effective Tool for Using Ultra Short Reads in Microbial Genome Assembly and Finishing. Marco Island, FL: AGBT; 2008. [Google Scholar]
- Hyatt D, Chen GL, LoCascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics. 2010;11:119. doi: 10.1186/1471-2105-11-119. http://dx.doi.org/10.1186/1471-2105-11-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kulichevskaya IS, Detkova EN, Bodelier PLE, Rijpstra WIC, Damsté JSS, Dedysh SN. Singulisphaera rosea sp. nov., a planctomycete from acidic Sphagnum peat, and emended description of the genus Singulisphaera. Int J Syst Evol Microbiol. 2012;62:118–23. doi: 10.1099/ijs.0.025924-0. http://dx.doi.org/10.1099/ijs.0.025924-0. [DOI] [PubMed] [Google Scholar]
- Mavromatis K, Ivanova NN, Chen IM, Szeto E, Markowitz VM, Kyrpides NC. The DOE-JGI Standard operating procedure for the annotations of microbial genomes. Stand Genomic Sci. 2009;1:63–7. doi: 10.4056/sigs.632. http://dx.doi.org/10.4056/sigs.632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Markowitz VM, Ivanova NN, Chen IMA, Chu K, Kyrpides NC. IMG ER: a system for microbial genome annotation expert review and curation. Bioinformatics. 2009;25:2271–8. doi: 10.1093/bioinformatics/btp393. http://dx.doi.org/10.1093/bioinformatics/btp393. [DOI] [PubMed] [Google Scholar]
- Wiley EO, Lieberman BS. Phylogenetics. Theory and practice of phylogenetic systematics. Hoboken, NJ: Wiley-Blackwell; 2011. [Google Scholar]
- Farris JS. The information content of the phylogenetic system. Syst Zool. 1979;28:483–519. doi: 10.2307/2412562. [doi:10.2307/2412562] [DOI] [Google Scholar]
- Hennig W. Phylogenetic systematics. Annu Rev Entomol. 1965;10:97–116. doi: 10.1146/annurev.en.10.010165.000525. [doi:10.1146/annurev.en.10.010165.000525] [DOI] [Google Scholar]
- Yarza P, Ludwig W, Euzéby J, Amann R, Schleifer KH, Glöckner FO, Rosselló-Móra R. Update of the all-species living tree project based on 16S and 23S rRNA sequence analyses. Syst Appl Microbiol. 2010;33:291–9. doi: 10.1016/j.syapm.2010.08.001. http://dx.doi.org/10.1016/j.syapm.2010.08.001. [DOI] [PubMed] [Google Scholar]
- Kulichevskaya IS, Ivanova AO, Belova SE, Baulina OI, Bodelier PLE, Rijpstra WIC, Sinninghe Damste JS, Zavarzin GA, Dedysh SN. Schlesneria paludicola gen. nov., sp. nov., the first acidophilic member of the order Planctomycetales, from Sphagnum-dominated boreal wetlands. Int J Syst Evol Microbiol. 2007;57:2680–7. doi: 10.1099/ijs.0.65157-0. http://dx.doi.org/10.1099/ijs.0.65157-0. [DOI] [PubMed] [Google Scholar]
- Phillips M, Delsuc F, Penny D. Genome-scale phylogeny and the detection of systematic biases. Mol Biol Evol. 2004;21:1455–8. doi: 10.1093/molbev/msh137. http://dx.doi.org/10.1093/molbev/msh137. [DOI] [PubMed] [Google Scholar]
- Stamatakis A, Hoover P, Rougemont J. A rapid bootstrap algorithm for the RAxML web-servers. Syst Biol. 2008;57:758–71. doi: 10.1080/10635150802429642. http://dx.doi.org/10.1080/10635150802429642. [DOI] [PubMed] [Google Scholar]
- Swofford DL. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0 b10. Sunderland: Sinauer Associates; 2002. [Google Scholar]
- Felsenstein J. Inferring phylogenies. Sunderland, Massachusetts: Sinauer Associates Inc.; 2004. [Google Scholar]
- Schlesner H. Pirella marina sp. nov., a budding peptidoglycan-less bacterium from brackish water. Syst Appl Microbiol. 1986;8:177–80. doi: 10.1016/S0723-2020(86)80073-X. http://dx.doi.org/10.1016/S0723-2020(86)80073-X. [DOI] [Google Scholar]
- Franzmann PD, Skerman VBD. Gemmata obscuriglobus, a new genus and species of the budding bacteria. Antonie Van Leeuwenhoek. 1984;50:261–8. doi: 10.1007/BF02342136. http://dx.doi.org/10.1007/BF02342136. [DOI] [PubMed] [Google Scholar]
- Giovannoni SJ, Schabtach E, Castenholz RW. Isosphaera pallida, gen. and comb. nov., a gliding, budding eubacterium from hot springs. Arch Microbiol. 1987;147:276–84. doi: 10.1007/BF00463488. http://dx.doi.org/10.1007/BF00463488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukunaga Y, Kurahashi M, Sakiyama Y, Ohuchi M, Yokota A, Harayama S. Phycisphaera mikurensis gen. nov., sp. nov., isolated from a marine alga, and proposal of Phycisphaeraceae fam. nov., Phycisphaerales ord. nov. and Phycisphaerae classis nov. in the phylum Planctomycetes. J Gen Appl Microbiol. 2009;55:267–75. doi: 10.2323/jgam.55.267. http://dx.doi.org/10.2323/jgam.55.267. [DOI] [PubMed] [Google Scholar]
- Schlesner H, Hirsch P. Assignment of ATCC 27377 to Pirella gen. nov. as Pirella staleyi comb. nov. Int J Syst Bacteriol. 1984;34:492–5. doi: 10.1099/00207713-34-4-492. http://dx.doi.org/10.1099/00207713-34-4-492. [DOI] [Google Scholar]
- Sittig M, Schlesner H. Chemotaxonomic investigation of various prosthecate and/or budding bacteria. Syst Appl Microbiol. 1993;16:92–103. doi: 10.1016/S0723-2020(11)80253-5. http://dx.doi.org/10.1016/S0723-2020(11)80253-5. [DOI] [Google Scholar]
- Schlesner H, Rensmann C, Tindall BJ, Gade D, Rabus R, Pfeiffer S, Hirsch P. Taxonomic heterogeneity within the Planctomycetales as derived by DNA–DNA hybridization, description of Rhodopirellula baltica gen. nov., sp. nov., transfer of Pirellula marina to the genus Blastopirellula gen. nov. as Blastopirellula marina comb. nov. and emended description of the genus Pirellula. Int J Syst Evol Microbiol. 2004;54:1567–80. doi: 10.1099/ijs.0.63113-0. http://dx.doi.org/10.1099/ijs.0.63113-0. [DOI] [PubMed] [Google Scholar]
- Kulichevskaya IS, Ivanova AO, Baulina OI, Bodelier PLE, Damsté JSS, Dedysh SN. Singulisphaera acidiphila gen. nov., sp. nov., a non-filamentous, Isosphaera-like planctomycete from acidic northern wetlands. Int J Syst Evol Microbiol. 2008;58:1186–93. doi: 10.1099/ijs.0.65593-0. http://dx.doi.org/10.1099/ijs.0.65593-0. [DOI] [PubMed] [Google Scholar]
- Kulichevskaya IS, Baulina OI, Bodelier PLE, Rijpstra WIC, Damsté JSS, Dedysh SN. Zavarzinella formosa gen. nov., sp. nov., a novel stalked, Gemmata-like planctomycete from a Siberian peat bog. Int J Syst Evol Microbiol. 2009;59:357–64. doi: 10.1099/ijs.0.002378-0. http://dx.doi.org/10.1099/ijs.0.002378-0. [DOI] [PubMed] [Google Scholar]
- Meier-Kolthoff JP, Klenk HP, Göker M. Taxonomic use of the G+C content and DNA:DNA hybridization in the genomic aG. Int J Syst Evol Microbiol. 2014;64:352–6. doi: 10.1099/ijs.0.056994-0. http://dx.doi.org/10.1099/ijs.0.056994-0. [DOI] [PubMed] [Google Scholar]
- Bondoso J, Albuquerque L, Nobre MF, Lobo-da-Cunha A, da Costa MS, Lage OM. Aquisphaera giovannonii gen. nov., sp. nov., a planctomycete isolated from a freshwater aquarium. Int J Syst Evol Microbiol. 2011;61:2844–50. doi: 10.1099/ijs.0.027474-0. http://dx.doi.org/10.1099/ijs.0.027474-0. [DOI] [PubMed] [Google Scholar]
- Kulichevskaya IS, Serkebaeva YM, Kim Y, Rijpstra WIC, Damsté JSS, Liesack W, Dedysh SN. Telmatocola sphagniphila gen. nov., sp. nov., a novel dendriform planctomycete from Northern Wetlands. Front Microbiol. 2012;3:146. doi: 10.3389/fmicb.2012.00146. http://dx.doi.org/10.3389/fmicb.2012.00146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spring S, Scheuner C, Lapidus A, Lucas S, Glavina Del Rio T, Tice H, Copeland A, Cheng JF, Chen F, Nolan M, Saunders E, Pitluck S, Liolios K, Ivanova N, Mavromatis K, Lykidis A, Pati A, Chen A, Palaniappan K, Land M, Hauser L, Chang YJ, Jeffries CD, Goodwin L, Detter JC, Brettin T, Rohde M, Göker M, Woyke T, Bristow J. et al. The genome sequence of Methanohalophilus mahii SLPT reveals differences in the energy metabolism among members of the Methanosarcinaceae inhabiting freshwater and saline environments. Archaea. 2010;2010:690737. doi: 10.1155/2010/690737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson IJ, Scheuner C, Göker M, Mavromatis K, Hooper SD, Porat I, Klenk HP, Ivanova N, Kyrpides NC. Novel insights into the diversity of catabolic metabolism from ten haloarchaeal genomes. PLoS One. 2011;6:e20237. doi: 10.1371/journal.pone.0020237. http://dx.doi.org/10.1371/journal.pone.0020237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Göker M, Scheuner C, Klenk HP, Stielow JB, Menzel W. Codivergence of Mycoviruses with Their Hosts. PLoS One. 2011;6:e22252. doi: 10.1371/journal.pone.0022252. http://dx.doi.org/10.1371/journal.pone.0022252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abt B, Han C, Scheuner C, Lu M, Lapidus A, Nolan M, Lucas S, Hammon N, Deshpande S, Cheng JF, Tapia R, Goodwin L, Pitluck S, Liolios K, Pagani I, Ivanova N, Mavromatis K, Mikhailova N, Huntemann M, Pati A, Chen A, Palaniappan K, Land M, Hauser L, Brambilla EM, Rohde M, Spring S, Gronow S, Göker M, Woyke T. et al. Complete genome sequence of the termite hindgut bacterium Spirochaeta coccoides type strain (SPN1T), reclassification in the genus Sphaerochaeta as Sphaerochaeta coccoides comb. nov. and emendations of the family Spirochaetaceae and the genus Sphaerochaeta. Stand Genomic Sci. 2012;6:194–209. doi: 10.4056/sigs.2796069. http://dx.doi.org/10.4056/sigs.2796069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–402. doi: 10.1093/nar/25.17.3389. http://dx.doi.org/10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Enright AJ, Van Dongen SM, Ouzounis CA. An efficient algorithm for large-scale detection of protein families. Nucleic Acids Res. 2002;30:1575–84. doi: 10.1093/nar/30.7.1575. http://dx.doi.org/10.1093/nar/30.7.1575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li L, Stoeckert CJ Jr, Roos DS. OrthoMCL: identification of ortholog groups for eukaryotic genomes. Genome Res. 2003;13:2178–89. doi: 10.1101/gr.1224503. http://dx.doi.org/10.1101/gr.1224503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32:1792–7. doi: 10.1093/nar/gkh340. http://dx.doi.org/10.1093/nar/gkh340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson JD, Thierry JCC, Poch O. RASCAL: rapid scanning and correction of multiple sequence alignments. Bioinformatics. 2003;19:1155–61. doi: 10.1093/bioinformatics/btg133. http://dx.doi.org/10.1093/bioinformatics/btg133. [DOI] [PubMed] [Google Scholar]
- Castresana J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol. 2000;17:540–52. doi: 10.1093/oxfordjournals.molbev.a026334. http://dx.doi.org/10.1093/oxfordjournals.molbev.a026334. [DOI] [PubMed] [Google Scholar]
- Meusemann K, Von Reumont BM, Simon S, Roeding F, Strauss S, Kück P, Ebersberger I, Walzl M, Pass G, Breuers S, Achter V, Von Haeseler A, Burmester T, Hadrys H, Wägele JW, Misof B. A phylogenomic approach to resolve the arthropod tree of life. Mol Biol Evol. 2010;27:2451–64. doi: 10.1093/molbev/msq130. http://dx.doi.org/0.1093/molbev/msq130. [DOI] [PubMed] [Google Scholar]
- Felsenstein J. Evolutionary trees from DNA sequences: a maximum likelihood approach. J Mol Evol. 1981;17:368–76. doi: 10.1007/BF01734359. http://dx.doi.org/10.1007/BF01734359. [DOI] [PubMed] [Google Scholar]
- Fitch WM. Toward defining the course of evolution: minimum change on a specified tree topology. Syst Zool. 1971;20:406–16. doi: 10.2307/2412116. http://dx.doi.org/10.2307/2412116. [DOI] [Google Scholar]
- Goloboff PA. Parsimony, likelihood, and simplicity. Cladistics. 2003;19:91–103. doi: 10.1111/j.1096-0031.2003.tb00297.x. http://dx.doi.org/10.1111/j.1096-0031.2003.tb00297.x. [DOI] [Google Scholar]
- Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006;22:2688–90. doi: 10.1093/bioinformatics/btl446. http://dx.doi.org/10.1093/bioinformatics/btl446. [DOI] [PubMed] [Google Scholar]
- Pattengale ND, Alipour M, Bininda-Emonds ORP, Moret BME, Stamatakis A. How many bootstrap replicates are necessary? Lect Notes Comput Sci. 2009;5541:184–200. doi: 10.1007/978-3-642-02008-7_13. http://dx.doi.org/10.1007/978-3-642-02008-7_13. [DOI] [PubMed] [Google Scholar]
- Guo M, Thou Q, Thou Y, Yang L, Liu T, Yang J, Chen Y, Su K, Xu J, Chen J, Liu F, Chen J, Dai W, Ni P, Fang C, Yang R. Genomic evolution of 11 type strains within the family Planctomycetaceae. PLoS One. 2014;9:e86752. doi: 10.1371/journal.pone.0086752. http://dx.doi.org/10.1371/journal.pone.0086752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Auch AF, Von Jan M, Klenk HP, Göker M. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand Genomic Sci. 2010;2:117–34. doi: 10.4056/sigs.531120. http://dx.doi.org/10.4056/sigs.531120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Auch AF, Klenk HP, Göker M. Standard operating procedure for calculating genome-to-genome distances based on high-scoring segment pairs. Stand Genomic Sci. 2010;2:142–8. doi: 10.4056/sigs.541628. http://dx.doi.org/10.4056/sigs.541628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meier-Kolthoff JP, Auch AF, Klenk HP, Göker M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinformatics. 2013;14:60. doi: 10.1186/1471-2105-14-60. http://dx.doi.org/10.1186/1471-2105-14-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen H, Boutros PC. VennDiagram: a package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinformatics. 2011;12:35. doi: 10.1186/1471-2105-12-35. http://dx.doi.org/10.1186/1471-2105-12-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeske O, Jogler M, Petersen J, Sikorski J, Jogler C. From genome mining to phenotypic microarrays: Planctomycetes as source for novel bioactive molecules. Antonie Van Leeuwenhoek. 2013;104:551–67. doi: 10.1007/s10482-013-0007-1. http://dx.doi.org/10.1007/s10482-013-0007-1. [DOI] [PubMed] [Google Scholar]
- Vaas LAI, Sikorski J, Hofner B, Fiebig A, Buddruhs N, Klenk HP, Göker M. opm: an R package for analysing OmniLog® phenotype microarray data. Bioinformatics. 2013;29:1823–4. doi: 10.1093/bioinformatics/btt291. http://dx.doi.org/10.1093/bioinformatics/btt291. [DOI] [PubMed] [Google Scholar]
- Lee HW, Roh SW, Shin NR, Lee J, Whon TW, Jung MJ, Yun JH, Kim MS, Hyun DW, Kim D, Bae JW. Blastopirellula cremea sp. nov., isolated from a dead ark clam. Int J Syst Evol Microbiol. 2013;63:2314–9. doi: 10.1099/ijs.0.044099-0. http://dx.doi.org/10.1099/ijs.0.044099-0. [DOI] [PubMed] [Google Scholar]
- Griepenburg U, Ward-Rainey NL, Mohamed S, Schlesner H, Marxsen H, Rainey FA, Stackebrandt E, Auling G. Phylogenetic diversity, polyamine pattern and DNA base composition of members of the order Planctomycetales. Int J Syst Bacteriol. 1999;49:689–96. doi: 10.1099/00207713-49-2-689. http://dx.doi.org/10.1099/00207713-49-2-689. [DOI] [PubMed] [Google Scholar]
- Bauld J, Staley JT. Planctomyces maris sp. nov.: a marine isolate of the Planctomyces-Blastocaulis group of budding bacteria. J Gen Microbiol. 1976;97:45–55. doi: 10.1099/00221287-97-1-45. http://dx.doi.org/10.1099/00221287-97-1-45. [DOI] [Google Scholar]