

Abstract
Diabetic retinopathy is one of the most important causes of blindness. The underlying mechanisms of this disease include inflammatory changes and remodeling processes of the extracellular-matrix (ECM) leading to pericyte and vascular endothelial cell damage that affects the retinal circulation. In turn, this causes hypoxia leading to release of vascular endothelial growth factor (VEGF) to induce the angiogenesis process. Alpha-1 antitrypsin (AAT) is the most important circulating inhibitor of serine proteases (SERPIN). Its targets include elastase, plasmin, thrombin, trypsin, chymotrypsin, proteinase 3 (PR-3) and plasminogen activator (PAI). AAT modulates the effect of protease-activated receptors (PARs) during inflammatory responses. Plasma levels of AAT can increase 4-fold during acute inflammation then is so-called acute phase protein (APPs). Individuals with low serum levels of AAT could develop disease in lung, liver and pancreas. AAT is involved in extracellular matrix remodeling and inflammation, particularly migration and chemotaxis of neutrophils. It can also suppress nitric oxide (NO) by nitric oxide sintase (NOS) inhibition. AAT binds their targets in an irreversible way resulting in product degradation. The aim of this review is to focus on the points of contact between multiple factors involved in diabetic retinopathy and AAT resembling pleiotropic effects that might be beneficial.
Electronic supplementary material
The online version of this article (doi:10.1186/0717-6287-47-58) contains supplementary material, which is available to authorized users.
Keywords: Diabetic retinopathy, Alpha-1-antitrypsin, Diabetes, Endogenous anti-inflammatory agents, Retinal inflammation, NF-kB
Introduction
The overall prevalence of diabetic retinopathy (DR) in diabetic patients is about 34% worldwide and it is the leading cause of blindness in the working population (16–64 years old) [1]. The underlying mechanisms of this disease include degenerative and inflammatory changes as well as remodeling processes of the extracellular-matrix (ECM) leading to pericyte and vascular endothelial cell damage that severely affects the retinal microcirculation. In turn, this causes hypoxia, vascular endothelial growth factor (VEGF) release and angiogenesis [2–5]. Neovessels grow in the retina and also into the vitreous, and could induce hemorrhages due to their fragile walls [6, 7]. In advanced stages the development of vitreoretinal fibrosis promotes retinal traction and detachment [8]. It has widely been demonstrated that this process is one of the previous steps to blindness.
Unfortunately, the ophthalmic therapy for diabetic retinopathy is focused on severe stages of the disease. The treatment is carried out when it reaches the so-called pre-proliferative stage using pan-retinal photocoagulation; development of macular edema is treated with focal photocoagulation and anti-VEGF agents; presence of retinal detachment requires vitreoretinal surgery [9]. The development of molecules to treat diabetic retinopathy in early stages is scarcely explored. New insights into pharmaceutical molecules and the recent advances in regenerative medicine should be exploited in order to find a treatment for early DR.
Review
- AAT and inflammation
Protease-activated receptors
It is well known how alpha- 1 anti-trypsin (AAT) binds and inhibits serum serine proteases such as elastase, trypsin, thrombin and proteinase-3 (PR-3) [10]. These serin proteases are considered key mediators of the innate immune response [11, 12] and can activate specific receptors named protease-activated receptors (PARs) on the membrane of immune cells such as neutrophils, eosinophils and macrophages. PARs are a family of four receptors (PAR1-4) involved in the intracellular signaling cascade and PAR-1 and PAR-4 appear to be essential during inflammatory responses [13]. In neutrophils, cell activation is accompanied by Akt (also known as protein kinase B) phosphorylation, rise of intracellular Ca+2 and formation of actin filaments, leading to better cell motility [14]. The crucial role of PARs activation during disease progression was revealed in animal models of inflammation such as gastrointestinal diseases, neuroinflammatory and neurodegenerative processes, skin, or allergic responses [11] and insulin-deficient murine type 1 diabetes models [12]. Moreover, the expression of mRNA of the four members of PARs was found in the postnatal eye and in the retina of adult rat [15]. PAR-2 is expressed in a variety of cells, including neuronal tissue, leukocytes, and vascular endothelial cells [16] and it was found involved in neovascularization processes of proliferative retinopathies [17]. Furthermore, PAR-2 has a link between pro-inflammatory and pro-angiogenic effects mediated by TNF-α, via MEK/EK1/2 pathway in the retina [17]. In summary, the inhibition of serine proteases that activate PARs could contribute to decreasing the inflammatory and pro-angiogenic process.
Reactive oxygen and nitrogen species
It is known that reactive oxygen species (ROS) are generated during diabetic retinopathy [18, 19]. Particularly, superoxide anion production by polymorphonuclear cells (PMNs), was found to be higher in patients with DR than in patients without DR, suggesting that ROS may have a role in retinopathy development [20]. In eosinophils, a target of AAT, trypsin was able to induce superoxide anion production via PAR-2 [21]. Also reactive nitrogen species (RNS) such as nitric oxide (NO) could be modulated by AAT [22]. Du et al., observed a significant increase in superoxide, NO, cyclooxygenase (COX)-2 and leukostasis within retinal microvessels in a model of streptozotocin-treated diabetic rats. These effects were suppressed using a p38 mitogen-activated protein kinase (MAPK) inhibitor [23]. However, the role of AAT in the activation of p38 and ERK1/2 MAPK could not be demonstrated in in vitro studies of murine RAW 264.7 macrophagic cells stimulated with combined LPS and IFN-γ [23]. Therefore the relationship between AAT and superoxide anion production of NO seems to be partly regulated via MAPK in diabetic retinal microvessels, but not in cells of the innate immune system such as macrophages. However, some evidences suggest that the development of retinal neovessels requires the involvement of macrophages [24, 25]. The number of macrophages rises in the vitreous and in the retina of animals with oxygen induced retinopathy [26]. Also, a mutation of macrophage colony stimulator factor was reported to reduce retinal neovascularization [27]. These findings support the hypothesis that the activation and migration of macrophages contribute to the pathogenesis of retinal neovascularization.
Neutrophil chemotaxis
In the absence of any exogenous stimuli, AAT inactivates calcium-dependent cysteine protease calpain I (μ-calpain) and concomitantly induces random neutrophil migration and polarization. Moreover, rho GTPases are rapidly activated, and neutrophils show increase phosphorylation of ERK 1/2. Also, AAT inhibits neutrophil adhesion to fibrinogen [28]. Bergin et al. [14] have provided evidence that AAT modulates neutrophil chemotaxis by association with neutrophil membrane lipid rafts, interacting with the glycosylphosphatidylinositol linked (GPI-linked) membrane protein FcγRIIIb and inhibiting ADAM- 17 activity, a tumor necrosis factor alpha converting enzyme. Neutrophil migration is a process that occur due to chemotaxis [29], an event that is present in diabetic retinopathy [30].
On the other hand, glycosylated AAT can bind to IL-8, a ligand for CXCR1 (chemokine receptor 1), and the AAT-IL-8 complex formation can prevent IL-8 interaction with CXCR1 regulating neutrophil chemotaxis [14]. In response to IL-8, the cell is activated resulting in actin filament formation and cytoskeletal rearrangement, via Akt (also known as PKB) phosphorylation and Ca 2+flux. Thus, when IL-8 binds to AAT it cannot interact with CXCR1 and cell activation is inhibited (Figure 1).
Figure 1.
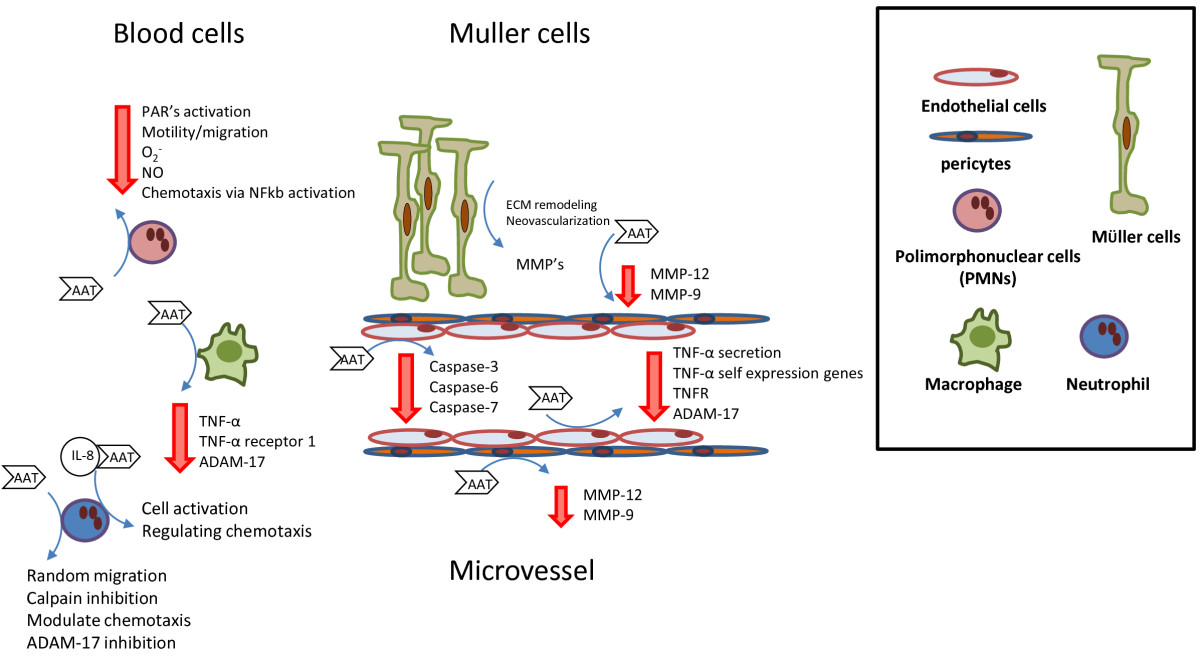
The interaction of AAT with Blood Cells and Müller Cells might influence the development of diabetic retinopathy.
CD40 and NFkB
It has been observed that CD154 (CD40 ligand) plays a key role in the production of pro-inflammatory cytokines and it has been linked to various autoimmune diseases with microvascular complications, like diabetes mellitus [31–33]. In vitro studies using Jurkat E6.1 T-cells demonstrated that the soluble form of CD154 (sCD154) is released from T-cells by ADAM10 and ADAM17 upon CD40 ligation [34]. Interestingly, a recent investigation performed in CD40 knock-out mice showed that these animals exhibited diminished inflammatory responses and they were protected from the development of diabetic retinopathy, suggesting that CD40 promotes the development of early diabetic retinopathy [35].
It was observed that AAT was able to inhibit nuclear transcriptional factor-kB (NF-kB) activation in a variety of animal models preventing PMN chemotaxis and the development of acute inflammation [36–38]. Activation of NF-kB induced by diabetes and high glucose regulates a pro-apoptotic program in retinal pericytes [39] and is well known that these cells are affected early in diabetic retinopathy [40].
Tumor necrosis factor-alpha and leucocytes
The effect of AAT on tumor necrosis factor alpha (TNF-α) was demonstrated in a microarray study in human endothelial lung cells. The co-administration of AAT inhibited 25% of genes up-regulated by TNF-α including TNF-α-induced self-expression. These effects were equally achieved when oxidized AAT, a modified form of AAT, lacking serine protease inhibitor activity was used [41]. AAT inhibited TNF-α receptor-1 up-regulation and significantly reduced TNF-α secretion. These results were associated with inhibition of TNF-α-converting enzyme activity or ADAM17. Furthermore, AAT inhibited calpain activity, whose activation by TNF-α contributed to decreasing intracellular AAT concentrations. All these data indicate that AAT initially facilitates acute responses of the endothelium to TNF-α, followed by selective inhibition of TNF-α-induced-self amplification, which may assist the vasculature in the resolution of chronic inflammation [42].
Intermittent infusions of alpha 1-antitrypsin were shown to be beneficial in the treatment of patients with alpha 1-antitrypsin deficiency [43] and augmentation therapy caused decreased neutrophil infiltration [44, 45]. Leukocytes and proteins that govern leukocyte adhesion to endothelial cells play a causal role in retinal abnormalities characteristic of the early stages of diabetic retinopathy, including diabetes-induced degeneration of retinal capillaries [46, 47]. These facts suggest a possible beneficial use of AAT in early stages of DR.
Protective effect on beta pancreatic cells
Non-functional circulating AAT (probably due to excessive non enzymatic glycation) was described in type 1 diabetes [48–53]. Additionally, levels of AAT in non-obese diabetic mice (NOD) were found to be half of those seen in the wild type strains [52]. These facts led to the development of gene therapy strategies using recombinant adeno-associated virus-(AAV) carrying murine AAT genes. As a result, AAV-AAT prevents type I diabetes in NOD mice [54]. Alpha-1-Antitrypsin (AAT) has been shown to reduce pro-inflammatory markers and protect pancreatic islets from autoimmune responses in pre-clinical studies [55]. Currently, clinical trials using recombinant AAT are being conducted in type 1 diabetic patients (Table 1). Preliminary results of one study showed better metabolic control probably through a protect effect on beta pancreatic cells that lead to a halt in disease progression. Diabetic retinopathy and other complications would benefit from this systemic therapy. In addition, visual scientists could consider the possibility to develop an ophthalmic treatment of AAT to further prevent or delay diabetic retinopathy.
Table 1.
Ongoing clinical trials using AAT in young patients with type 1 diabetes
| NCT | Phase | Age range (years) | Source/dose of AAT (mg) |
|---|---|---|---|
| 01304537 | II | 10 to 25 | Glassia®/40-60-80 |
| 01319331 | I | 6 to 45 | Aralast NP |
| 01183468 | II | 8 to 35 | Aralast NP |
- AAT role in cell death
Many studies have determined the ability of AAT to inhibit caspases. These are involved in cell death by apoptosis, as inducers or effectors [56]. The role of AAT in caspase-3 inhibition was described in murine lung endothelial cells and in murine pancreatic beta cells [57, 58]. Also, AAT was capable of inhibiting executing caspase-6 and −7 in lung microvascular endothelial cells [56]. Similar results were reported in animal models of diabetic retinopathy and also in diabetic patients. Activation of retinal caspases, particularly caspase-3, lead to apoptosis of endothelial cells and pericytes [59, 60]. The capacity of AAT to inhibit caspases could be exploited in order to protect microvasculature from early damage induced by DR (Figure 1).
- Potential interaction between AAT and Müller cells
Similary to brain astrocytes, Müller cells could produce factors that induce the formation of tight junctions conferring barrier properties to the retinal vessels [61]. They synthesize or store a number of growth factors with trophic or regulatory functions for various cell types in the retina. These characteristics make an assessment of Müller cell function in diabetes relevant to two well-known features of diabetic retinopathy: vascular leakage and capillary obliteration. Indeed, microvascular cell apoptosis occurs in human and experimental diabetic retinopathy [62], and one of the mechanisms leading to apoptosis is loss of survival signals provided by neighboring cells [63]. On the other hand, Müller cells might release metalloproteases (MMPs) that promote the degradation of extracellular matrix (ECM), along with the evidence that MMPs promote cell migration and proliferation. This strongly suggests that Müller cells play an important role in the control of cell and ECM interactions that, in turn, facilitate the development of retinal neovascularization (Figure 1).
It is noteworthy that Müller cells are currently being used in gene delivery. These cells transfected with plasmids or adeno-associated vectors (AAV) containing different constructions are a useful tool to explore different pathways. The retina is an attractive structure for gene therapy approaches because it is surgically approachable, isolated due to the presence of the blood-retinal barrier (BRB) and immunologically privileged. A study demonstrated that accumulation of hypoxia-inducible factor-1α in Müller cells induces the expression of VEGF, which in turn, promotes increased MMP-2 expression and activity in neighboring endothelial cells (EC). MMP-2 expression was detected in endothelial cells of retinal neovessels from proliferative diabetic retinopathy (PDR) patients, whereas MMP-2 protein levels were elevated in the aqueous humor of PDR patients compared with healthy patients [64]. The stability control of the microvasculature through regulation of the extracelullar matrix (ECM) in the retina is essential to avoid progressive development of the disease. AAT could be involved in the control of ECM because of its ability to inhibit MMP-12 and MMP-9. Furthermore, gene therapy using AAT could be a suitable tool for the inhibition of those changes.
The mRNA and protein levels of the complement receptor C5aR were measured in human Müller cells. C5aR was found constitutively expressed in human Müller cells. Up-regulated C5aR expression in Müller cells was promoted by, prostaglandin E2 and hyperglycemia, either individually or synergistically. Signaling through C5aR on Müller cells up-regulated production of IL-6 and VEGF, which promoted the proliferation of human retinal endothelial cells and increased their permeability [65]. Furthermore, IL-6 seems to be involved in the regulation of AAT since human hepatocyte exposure to IL-6 increased the expression levels of AAT [66]. A recent investigation also found increased IL-6 levels in diabetic animals [67]. This information suggests that complement plays a role in disease progression but how this could modulate the activity of AAT and the relationship between AAT and C5aR remains to be verified. However, the use of silencing strategies to reduce the availability of the receptor C5aR in the retina might be beneficial. Similar strategies have already been used in retinal Müller cells [68].
- AAT and extracellular matrix remodelling
MMPs are a family of enzymes capable of degrading essentially all ECM components [69]. The two major matrix degrading enzymes, known as MMP-2 and MMP-9 were found in the vitreous of eyes with proliferative DR [70]. The main source of these MMPs in vivo may be retinal pigment epithelial cells [71–73]. In the retina of diabetic rats the activation of cytosolic MMP-9 and MMP-2 is an early event, which is followed by their accumulation in the mitochondria [74]. In humans, it was found a positive correlation between vitreous levels of MMP-9 and VEGF with proliferative DR [75], and levels of AAT were found increased in different types of vitreoretinal diseases [76]. Besides, another study found higher vitreous levels of AAT in proliferative DR compared with vitreous levels seen in cases without diabetes mellitus [77]. Another MMP, MMP-12 is mainly produced by macrophages and called both metallo-elastase or macrophage-elastase [78]. An important factor in the development of vascular wall alterations is the degradation of the elastic fiber major protein-elastin [79]. It should be noted that hyperglycemia may directly disrupt elastin formation [80]. In diseases such as chronic obstructive pulmonary disease (COPD), it has been shown that AAT is capable of inhibiting the action of MMP-12. Besides, preliminary results on streptozotocin induced diabetes in rats intravitreally treated with human alpha-1 proteinase inhibitor Prolastin® have shown a higher expression of MMP-12 compared with controls (Ortiz et al. unpublished data). AAT also inhibited MMP-9 in a mouse model of the autoimmune disease bullous pemphigoid [81]. MMP-9 is an important IL-1 inducible protease that is suspected of contributing to the progression of various diseases such as cardiovascular disease, rheumatoid arthritis, COPD and multiple sclerosis [81, 82]. These evidences together suggest that progression of angiogenesis is associated with MMP’s and also with inflammation process in the vitreoretinal diseases. It is important to better understand these processes, to avoid the progression of the disease.
Recent studies on the role of epigenetic patterns in streptozotocin-induced diabetic rats reported an altered pattern of methylation of histone H3K4 H3K9 located in the promoter of MMP-9. The activity of Lysine-specific demethylase 1 (LSD1) was found elevated by 50% and gene and protein expression was 2-fold augmented. Gene activation markers, acetyl H3K9 and NF-kB (p65 subunit) recruitment were found to be increased by about 18-fold and 30-fold, respectively [83]. Epigenetic changes modify the expression pattern of MMP’s occurring at early stages in the development of DR. To ameliorate these changes the use of molecules that neutralize MMP’s action seems to be necessary.
The outgrowth of mouse retinal ganglion cells (RGCs) is co-regulated by MMP-2 and another membrane type 1 MMP (MT1-MMP) [84]. Furthermore, in an ex vivo retinal explant model MMPs were shown to be beneficial factors in axonal regeneration. On the other hand, CD44 proteolysis in T-cells is involved in migration and function of self-reactive T-cells, and a study using three MMP inhibitors in NOD mice found that MT1-MMP has a unique involvement in type 1 diabetes development [85].
- Vessel walls and capillaries might be protected by AAT
Pericyte loss and microaneurysm formation are hallmarks of early changes in the retinas of diabetic patients [86]. After induction of diabetes in rodents, reduction of pericyte number in retinal capillaries is the earliest morphological change, followed by the formation of increased number of acellular-occluded capillaries, occasional microaneurysms, and thickening of the vascular basement membrane [87]. With progressive vascular occlusions in the human diabetic eye, the retina responds with either a progressive increase of vascular permeability leading to retinal edema, or the formation of new vessels that finally proliferate into the vitreous [5].
Pericytes can control endothelial cell proliferation and angiogenesis, both under physiological and pathological conditions [88–94]. DR is morphologically characterized by pathological changes in the retinal capillaries. The primary and predominant characteristics are the loss of pericytes and the progressive occlusion of capillaries [3, 86]. Several research groups [39, 95, 96] have reported that cultured retinal pericytes exposed to high levels of glucose (25–30 mmol/l) for 7 days or more show a higher rate of apoptosis than cells grown at 5.5 mmol/l glucose. Besides, it has been found that retinal pericytes play a key role in the stabilization of endothelial cells protecting them from hypoxic insults and angiogenic stimuli [4].
Other research groups working on animals at 10 months post diabetes-induction have reported significant increases in the number of degenerate (acellular) capillaries and pericyte ghosts compared to non-diabetic animals. However, when the inhibitor of p38 MAPK was used, all these abnormalities were significantly diminished [23].
It is known that bone-marrow-stem-cells (BMSCs) appear to act primarily through their incorporation into the retina as endothelial cells, microglia, and photoreceptors [97–101]. Also, pericytes can be derived from BMSCs [102], but this does not appear to be a predominant differentiation pathway for these cells when injected into the eye [98, 103]. A recent study showed that pericytes obtained from adipose-derived stem cells (ASCs) protect against retinal vasculopathy. It is noteworthy that ASCs express pericyte-specific markers in vitro, and when they were intravitreally injected into the eye of a mouse model of oxygen-induced-retinopaty (OIR) they were capable of migrating and integrating in the vasculature [104].
The breakdown of the inner blood-retinal barrier (iBRB) is also a feature of experimental diabetes in animal models, being observed as early as 1-2-weeks post-diabetes induction in rodents [105, 106]. It is well established that this lesion occurs early in clinical diabetic retinopathy [107].
Advanced-glication-end products (AGEs) are known to induce expression of the potent angiogenic agent VEGF in the retina in vivo [108, 109] and in retinal cells in vitro [110, 111]. It has been demonstrated that in short-term diabetic rodents (3 weeks post induction of streptozotocin 165 mg/kg) inhibition of AGEs prevents disruption of iBRB [112]. Besides, AGEs mediated expression and secretion of TNF-α in rat retinal microglia [113].
We previously pointed out the capacity of AAT to inhibit protease-activated receptors, to diminish neutrophil chemotaxis, to hinder NFkB activation, to reduce the effect of TNF-alpha and also to inhibit caspases. Through these mechanisms described above AAT might protect the structures of the vessel walls of retinal capillaries that are damaged in DR development.Figure 2 schematizes the above data regarding the involvement of AAT in different pathways during DR progression.
Figure 2.
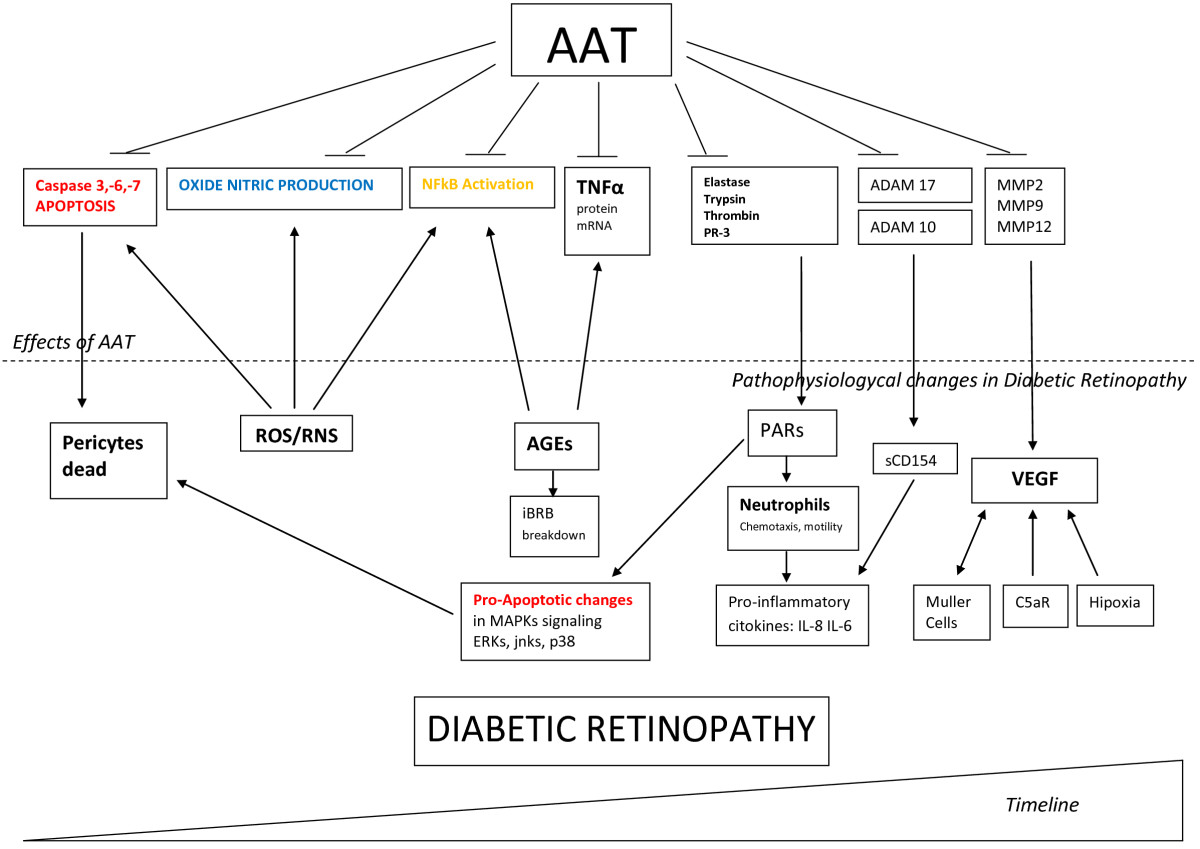
AAT might ameliorate DR progression inhibiting many key pathways of inflammation in early and advanced disease. AAT could inhibit several pathophysiological changes that occur during DR. In early stages AAT can inhibit effector caspases preventing the loss of pericytes. In turn, the resulting production of NO could be decreased. Both ROS and AGEs stimulate production of two proinflammatory key molecules: NFkb and TNF-α. Inactivation of these molecules may be performed partially by AAT. During chronic inflammatory processes AAT can inhibit activation of PARPs by blocking the action of serine proteases such as elastase, trypsin, thrombin and PR-3. Finally the process of neovascularization requires remodeling of the extracellular matrix, thereby inhibition of several MMP through AAT may partly decrease the action of VEGF. AAT: alpha 1 antitrypsin ROS: Reactive Oxygen Species RNS: Reactive Nitrogen Species NFkB: Nuclear Factor kappa beta TNF-α: Tumor Necrosis Factor alpha PR-3: Proteinase 3 AGEs: Advanced Glycation End products iBRB: Blood Retinal Barrier MAPKs: Mitogen-Activated Protein Kinases ERKs: Extracellular signal-regulated Kinases Jnks: c-Jun N-terminal kinases PARs: Protease Activated Receptors IL-6: Interleukin 6 IL-8: Interleukin 8 ADAM17: Metallopeptidase domain 17 ADAM10: Metallopeptidase domain 10 MMP-2: Matrix Metalloprotease 2 MMP-9: Matrix Metalloprotease 9 MMP-12: Matrix Metalloprotease 12 VEGF: Vascular Endothelial Growth Factor C5aR: Complement 5a Receptor.
Conclusions
The above data support the potential protective role of AAT in diabetic retinopathy as a result of its multiple activities and anti-inflammatory properties. AAT is able to inhibit key pro-inflammatory molecules such as NF-kB and TNF-α, as well as all serine proteases involved in activating PARs. Taking into account that activated PARs control neutrophil chemotaxis and motility, a hallmark of inflammatory chronic processes such as those present in diabetic retinopathy, AAT could be administered in the early or advanced stages of DR for the patients to achieve a therapeutic benefit.
Anti-apoptotic properties inhibiting caspase 3, 6, 7 could be beneficial in the pathogenesis of DR and any neurodegenerative process that may occur. Indirect anti-angiogenic features in the retinal microvasculature could decrease ECM remodeling. Because AAT could delay the damage induced by DR, early use of AAT therapy may be an effective strategy to prevent or hinder the progression of diabetic retinopathy.
Acknowledgements
We want to thank Mariana Malvicini for her invaluable help to carry out this review. Not only for her boundless intellectual capacity but also for her personal kindness, and Jorge Mancini for his interesting and never-ending discussions.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
GO carried out an extensive review of the bibliography regarding AAT and diabetes. JEG and GO participated in the design of the mini-review and both wrote the first draft of the manuscript. JPS participated in the discussion of the paper and design of figures. EHC helped to draft the manuscript and to make final corrections. All authors read and approved the final manuscript.
Contributor Information
Gustavo Ortiz, Email: gaodelacalle@gmail.com.
Juan P Salica, Email: jp.salica@gmail.com.
Eduardo H Chuluyan, Email: echuluyan@gmail.com.
Juan E Gallo, Email: jgallo06@gmail.com.
References
- 1.Yau JW, Rogers SL, Kawasaki R, Lamoureux EL, Kowalski JW, Bek T, Chen SJ, Dekker JM, Fletcher A, Grauslund J, Haffner S, Hamman RF, Ikram MK, Kayama T, Klein BE, Klein R, Krishnaiah S, Mayurasakorn K, O'Hare JP, Orchard TJ, Porta M, Rema M, Roy MS, Sharma T, Shaw J, Taylor H, Tielsch JM, Varma R, Wang JJ, Wang N, et al. Global prevalence and major risk factors of diabetic retinopathy. Diabetes Care. 2012;35:556–564. doi: 10.2337/dc11-1909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Egwuagu CE. Chronic intraocular inflammation and development of retinal degenerative disease. Adv Exp Med Biol. 2014;801:417–425. doi: 10.1007/978-1-4614-3209-8_53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hammes HP, Lin J, Renner O, Shani M, Lundqvist A, Betsholtz C, Brownlee M, Deutsch U. Pericytes and the pathogenesis of diabetic retinopathy. Diabetes. 2002;51:3107–3112. doi: 10.2337/diabetes.51.10.3107. [DOI] [PubMed] [Google Scholar]
- 4.Perdiguero EG, Galaup A, Durand M, Teillon J, Philippe J, Valenzuela DM, Murphy AJ, Yancopoulos GD, Thurston G, Germain S. Alteration of developmental and pathological retinal angiogenesis in angptl4-deficient mice. J Biol Chem. 2011;286:36841–36851. doi: 10.1074/jbc.M111.220061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Aiello LP, Gardner TW, King GL, Blankenship G, Cavallerano JD, Ferris FL, 3rd, Klein R. Diabetic retinopathy. Diabetes Care. 1998;21:143–156. doi: 10.2337/diacare.21.1.143. [DOI] [PubMed] [Google Scholar]
- 6.Abu El-Asrar AM, Nawaz MI, Kangave D, Siddiquei MM, Ola MS, Opdenakker G. Angiogenesis regulatory factors in the vitreous from patients with proliferative diabetic retinopathy. Acta Diabetol. 2013;50:545–551. doi: 10.1007/s00592-011-0330-9. [DOI] [PubMed] [Google Scholar]
- 7.Abu El-Asrar AM, Nawaz MI, Kangave D, Mairaj Siddiquei M, Geboes K. Angiogenic and vasculogenic factors in the vitreous from patients with proliferative diabetic retinopathy. J Diabetes Res. 2013;2013:539658. doi: 10.1155/2013/539658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Romano MR, Christoforidis J, Abu El-Asrar AM. Intravitreal inflammation: from benchside to bedside. Mediators Inflamm. 2013;2013:758035. doi: 10.1155/2013/758035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Abu El-Asrar AM. Evolving strategies in the management of diabetic retinopathy. Middle East Afr J Ophthalmol. 2013;20:273–282. doi: 10.4103/0974-9233.119993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Korkmaz B, Horwitz MS, Jenne DE, Gauthier F. Neutrophil elastase, proteinase 3, and cathepsin G as therapeutic targets in human diseases. Pharmacol Rev. 2010;62:726–759. doi: 10.1124/pr.110.002733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Vergnolle N. Clinical relevance of proteinase activated receptors (pars) in the gut. Gut. 2005;54:867–874. doi: 10.1136/gut.2004.048876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hyun E, Ramachandran R, Cenac N, Houle S, Rousset P, Saxena A, Liblau RS, Hollenberg MD, Vergnolle N. Insulin modulates protease-activated receptor 2 signaling: implications for the innate immune response. J Immunol. 2010;184:2702–2709. doi: 10.4049/jimmunol.0902171. [DOI] [PubMed] [Google Scholar]
- 13.Shpacovitch V, Feld M, Hollenberg MD, Luger TA, Steinhoff M. Role of protease-activated receptors in inflammatory responses, innate and adaptive immunity. J Leukoc Biol. 2008;83:1309–1322. doi: 10.1189/jlb.0108001. [DOI] [PubMed] [Google Scholar]
- 14.Bergin DA, Reeves EP, Meleady P, Henry M, McElvaney OJ, Carroll TP, Condron C, Chotirmall SH, Clynes M, O'Neill SJ, McElvaney NG. alpha-1 Antitrypsin regulates human neutrophil chemotaxis induced by soluble immune complexes and IL-8. J Clin Invest. 2010;120:4236–4250. doi: 10.1172/JCI41196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rohatgi T, Sedehizade F, Sabel BA, Reiser G. Protease-activated receptor subtype expression in developing eye and adult retina of the rat after optic nerve crush. J Neurosci Res. 2003;73:246–254. doi: 10.1002/jnr.10643. [DOI] [PubMed] [Google Scholar]
- 16.Macfarlane SR, Seatter MJ, Kanke T, Hunter GD, Plevin R. Proteinase-activated receptors. Pharmacol Rev. 2001;53:245–282. [PubMed] [Google Scholar]
- 17.Zhu T, Sennlaub F, Beauchamp MH, Fan L, Joyal JS, Checchin D, Nim S, Lachapelle P, Sirinyan M, Hou X, Bossolasco M, Rivard GE, Heveker N, Chemtob S. Proangiogenic effects of protease-activated receptor 2 are tumor necrosis factor-alpha and consecutively Tie2 dependent. Arterioscler Thromb Vasc Biol. 2006;26:744–750. doi: 10.1161/01.ATV.0000205591.88522.d4. [DOI] [PubMed] [Google Scholar]
- 18.Bonne C, Muller A, Villain M. Free radicals in retinal ischemia. Gen Pharmacol. 1998;30:275–280. doi: 10.1016/S0306-3623(97)00357-1. [DOI] [PubMed] [Google Scholar]
- 19.Giacco F, Brownlee M. Oxidative stress and diabetic complications. Circ Res. 2010;107:1058–1070. doi: 10.1161/CIRCRESAHA.110.223545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Abu El-Asrar AM, Soliman RT, Al-Amro SA, Al-Shammary FJ. Serum factor from diabetic patients with or without retinopathy stimulates superoxide anion production by normal polymorphonuclear leukocytes. Doc Ophthalmol. 1995;91:1–8. doi: 10.1007/BF01204618. [DOI] [PubMed] [Google Scholar]
- 21.Miike S, McWilliam AS, Kita H. Trypsin induces activation and inflammatory mediator release from human eosinophils through protease-activated receptor-2. J Immunol. 2001;167:6615–6622. doi: 10.4049/jimmunol.167.11.6615. [DOI] [PubMed] [Google Scholar]
- 22.Chan ED, Pott GB, Silkoff PE, Ralston AH, Bryan CL, Shapiro L. Alpha-1-antitrypsin inhibits nitric oxide production. J Leukoc Biol. 2012;92:1251–1260. doi: 10.1189/jlb.0212071. [DOI] [PubMed] [Google Scholar]
- 23.Du Y, Tang J, Li G, Berti-Mattera L, Lee CA, Bartkowski D, Gale D, Monahan J, Niesman MR, Alton G, Kern TS. Effects of p38 MAPK inhibition on early stages of diabetic retinopathy and sensory nerve function. Invest Ophthalmol Vis Sci. 2010;51:2158–2164. doi: 10.1167/iovs.09-3674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Davies MH, Eubanks JP, Powers MR. Microglia and macrophages are increased in response to ischemia-induced retinopathy in the mouse retina. Mol Vis. 2006;12:467–477. [PubMed] [Google Scholar]
- 25.Shen J, Xie B, Dong A, Swaim M, Hackett SF, Campochiaro PA. In vivo immunostaining demonstrates macrophages associate with growing and regressing vessels. Invest Ophthalmol Vis Sci. 2007;48:4335–4341. doi: 10.1167/iovs.07-0113. [DOI] [PubMed] [Google Scholar]
- 26.Naug HL, Browning J, Gole GA, Gobe G. Vitreal macrophages express vascular endothelial growth factor in oxygen-induced retinopathy. Clin Experiment Ophthalmol. 2000;28:48–52. doi: 10.1046/j.1442-9071.2000.00226.x. [DOI] [PubMed] [Google Scholar]
- 27.Kubota Y, Takubo K, Shimizu T, Ohno H, Kishi K, Shibuya M, Saya H, Suda T. M-CSF inhibition selectively targets pathological angiogenesis and lymphangiogenesis. J Exp Med. 2009;206:1089–1102. doi: 10.1084/jem.20081605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Al-Omari M, Korenbaum E, Ballmaier M, Lehmann U, Jonigk D, Manstein DJ, Welte T, Mahadeva R, Janciauskiene S. Acute-phase protein alpha1-antitrypsin inhibits neutrophil calpain I and induces random migration. Mol Med. 2011;17:865–874. doi: 10.2119/molmed.2011.00089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wagner JG, Roth RA. Neutrophil migration mechanisms, with an emphasis on the pulmonary vasculature. Pharmacol Rev. 2000;52:349–374. [PubMed] [Google Scholar]
- 30.Rangasamy S, McGuire PG, Das A. Diabetic retinopathy and inflammation: novel therapeutic targets. Middle East Afr J Ophthalmol. 2012;19:52–59. doi: 10.4103/0974-9233.92116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Pipi E, Marketou M, Tsirogianni A. Distinct clinical and laboratory characteristics of latent autoimmune diabetes in adults in relation to type 1 and type 2 diabetes mellitus. World J Diabetes. 2014;5:505–510. doi: 10.4239/wjd.v5.i4.505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lee MS. Role of innate immunity in the pathogenesis of type 1 and type 2 diabetes. J Korean Med Sci. 2014;29:1038–1041. doi: 10.3346/jkms.2014.29.8.1038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Szablewski L. Role of immune system in type 1 diabetes mellitus pathogenesis. Int Immunopharmacol. 2014;22:182–191. doi: 10.1016/j.intimp.2014.06.033. [DOI] [PubMed] [Google Scholar]
- 34.Yacoub D, Benslimane N, Al-Zoobi L, Hassan G, Nadiri A, Mourad W. CD154 is released from T-cells by a disintegrin and metalloproteinase domain-containing protein 10 (ADAM10) and ADAM17 in a CD40 protein-dependent manner. J Biol Chem. 2013;288:36083–36093. doi: 10.1074/jbc.M113.506220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Portillo JA, Greene JA, Okenka G, Miao Y, Sheibani N, Kern TS, Subauste CS. CD40 promotes the development of early diabetic retinopathy in mice. Diabetologia. 2014;57:2222–2231. doi: 10.1007/s00125-014-3321-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Churg A, Dai J, Zay K, Karsan A, Hendricks R, Yee C, Martin R, MacKenzie R, Xie C, Zhang L, Shapiro S, Wright JL. Alpha-1-antitrypsin and a broad spectrum metalloprotease inhibitor, RS113456, have similar acute anti-inflammatory effects. Lab Invest. 2001;81:1119–1131. doi: 10.1038/labinvest.3780324. [DOI] [PubMed] [Google Scholar]
- 37.Shapiro L, Pott GB, Ralston AH. Alpha-1-antitrypsin inhibits human immunodeficiency virus type 1. FASEB J. 2001;15:115–122. doi: 10.1096/fj.00-0311com. [DOI] [PubMed] [Google Scholar]
- 38.Zhou X, Shapiro L, Fellingham G, Willardson BM, Burton GF. HIV replication in CD4+ T lymphocytes in the presence and absence of follicular dendritic cells: inhibition of replication mediated by alpha-1-antitrypsin through altered IkappaBalpha ubiquitination. J Immunol. 2011;186:3148–3155. doi: 10.4049/jimmunol.1001358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Romeo G, Liu WH, Asnaghi V, Kern TS, Lorenzi M. Activation of nuclear factor-kappaB induced by diabetes and high glucose regulates a proapoptotic program in retinal pericytes. Diabetes. 2002;51:2241–2248. doi: 10.2337/diabetes.51.7.2241. [DOI] [PubMed] [Google Scholar]
- 40.Beltramo E, Porta M. Pericyte loss in diabetic retinopathy: mechanisms and consequences. Curr Med Chem. 2013;20:3218–3225. doi: 10.2174/09298673113209990022. [DOI] [PubMed] [Google Scholar]
- 41.Subramaniyam D, Virtala R, Pawlowski K, Clausen IG, Warkentin S, Stevens T, Janciauskiene S. TNF-alpha-induced self expression in human lung endothelial cells is inhibited by native and oxidized alpha1-antitrypsin. Int J Biochem Cell Biol. 2008;40:258–271. doi: 10.1016/j.biocel.2007.07.016. [DOI] [PubMed] [Google Scholar]
- 42.Lockett AD, Kimani S, Ddungu G, Wrenger S, Tuder RM, Janciauskiene SM, Petrache I. alpha(1)-Antitrypsin modulates lung endothelial cell inflammatory responses to TNF-alpha. Am J Respir Cell Mol Biol. 2013;49:143–150. doi: 10.1165/rcmb.2012-0515OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wewers MD, Casolaro MA, Sellers SE, Swayze SC, McPhaul KM, Wittes JT, Crystal RG. Replacement therapy for alpha 1-antitrypsin deficiency associated with emphysema. N Engl J Med. 1987;316:1055–1062. doi: 10.1056/NEJM198704233161704. [DOI] [PubMed] [Google Scholar]
- 44.Griese M, Latzin P, Kappler M, Weckerle K, Heinzlmaier T, Bernhardt T, Hartl D. alpha1-Antitrypsin inhalation reduces airway inflammation in cystic fibrosis patients. Eur Respir J. 2007;29:240–250. doi: 10.1183/09031936.00047306. [DOI] [PubMed] [Google Scholar]
- 45.Subramaniyam D, Steele C, Kohnlein T, Welte T, Grip O, Matalon S, Janciauskiene S. Effects of alpha 1-antitrypsin on endotoxin-induced lung inflammation in vivo. Inflamm Res. 2010;59:571–578. doi: 10.1007/s00011-010-0164-x. [DOI] [PubMed] [Google Scholar]
- 46.Veenstra AA, Tang J, Kern TS. Antagonism of CD11b with neutrophil inhibitory factor (NIF) inhibits vascular lesions in diabetic retinopathy. PLoS ONE. 2013;8:e78405. doi: 10.1371/journal.pone.0078405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Segel GB, Halterman MW, Lichtman MA. The paradox of the neutrophil’s role in tissue injury. J Leukoc Biol. 2011;89:359–372. doi: 10.1189/jlb.0910538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yaghmaei M, Hashemi M, Shikhzadeh A, Mokhtari M, Niazi A, Ghavami S. Serum trypsin inhibitory capacity in normal pregnancy and gestational diabetes mellitus. Diabetes Res Clin Pract. 2009;84:201–204. doi: 10.1016/j.diabres.2009.03.003. [DOI] [PubMed] [Google Scholar]
- 49.Hashemi M, Naderi M, Rashidi H, Ghavami S. Impaired activity of serum alpha-1-antitrypsin in diabetes mellitus. Diabetes Res Clin Pract. 2007;75:246–248. doi: 10.1016/j.diabres.2006.06.020. [DOI] [PubMed] [Google Scholar]
- 50.Lisowska-Myjak B, Pachecka J, Kaczynska B, Miszkurka G, Kadziela K. Serum protease inhibitor concentrations and total antitrypsin activity in diabetic and non-diabetic children during adolescence. Acta Diabetol. 2006;43:88–92. doi: 10.1007/s00592-006-0220-8. [DOI] [PubMed] [Google Scholar]
- 51.Bristow CL, Di Meo F, Arnold RR. Specific activity of alpha1proteinase inhibitor and alpha2macroglobulin in human serum: application to insulin-dependent diabetes mellitus. Clin Immunol Immunopathol. 1998;89:247–259. doi: 10.1006/clin.1998.4605. [DOI] [PubMed] [Google Scholar]
- 52.Sandler M, Gemperli BM, Hanekom C, Kuhn SH. Serum alpha 1-protease inhibitor in diabetes mellitus: reduced concentration and impaired activity. Diabetes Res Clin Pract. 1988;5:249–255. doi: 10.1016/S0168-8227(88)80059-7. [DOI] [PubMed] [Google Scholar]
- 53.Hall P, Tryon E, Nikolai TF, Roberts RC. Functional activities and nonenzymatic glycosylation of plasma proteinase inhibitors in diabetes. Clin Chim Acta. 1986;160:55–62. doi: 10.1016/0009-8981(86)90335-9. [DOI] [PubMed] [Google Scholar]
- 54.Lu Y, Tang M, Wasserfall C, Kou Z, Campbell-Thompson M, Gardemann T, Crawford J, Atkinson M, Song S. Alpha1-antitrypsin gene therapy modulates cellular immunity and efficiently prevents type 1 diabetes in nonobese diabetic mice. Hum Gene Ther. 2006;17:625–634. doi: 10.1089/hum.2006.17.625. [DOI] [PubMed] [Google Scholar]
- 55.Ashkenazi E, Baranovski BM, Shahaf G, Lewis EC. Pancreatic islet xenograft survival in mice is extended by a combination of alpha-1-antitrypsin and single-dose anti-CD4/CD8 therapy. PLoS ONE. 2013;8:e63625. doi: 10.1371/journal.pone.0063625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lockett AD, Van Demark M, Gu Y, Schweitzer KS, Sigua N, Kamocki K, Fijalkowska I, Garrison J, Fisher AJ, Serban K, Wise RA, Flotte TR, Mueller C, Presson RG, Jr, Petrache HI, Tuder RM, Petrache I. Effect of cigarette smoke exposure and structural modifications on the alpha-1 Antitrypsin interaction with caspases. Mol Med. 2012;18:445–454. doi: 10.2119/molmed.2011.00207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Petrache I, Fijalkowska I, Medler TR, Skirball J, Cruz P, Zhen L, Petrache HI, Flotte TR, Tuder RM. alpha-1 antitrypsin inhibits caspase-3 activity, preventing lung endothelial cell apoptosis. Am J Pathol. 2006;169:1155–1166. doi: 10.2353/ajpath.2006.060058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zhang B, Lu Y, Campbell-Thompson M, Spencer T, Wasserfall C, Atkinson M, Song S. Alpha1-antitrypsin protects beta-cells from apoptosis. Diabetes. 2007;56:1316–1323. doi: 10.2337/db06-1273. [DOI] [PubMed] [Google Scholar]
- 59.Kowluru RA, Koppolu P. Diabetes-induced activation of caspase-3 in retina: effect of antioxidant therapy. Free Radic Res. 2002;36:993–999. doi: 10.1080/1071576021000006572. [DOI] [PubMed] [Google Scholar]
- 60.Mohr S, Xi X, Tang J, Kern TS. Caspase activation in retinas of diabetic and galactosemic mice and diabetic patients. Diabetes. 2002;51:1172–1179. doi: 10.2337/diabetes.51.4.1172. [DOI] [PubMed] [Google Scholar]
- 61.Arthur FE, Shivers RR, Bowman PD. Astrocyte-mediated induction of tight junctions in brain capillary endothelium: an efficient in vitro model. Brain Res. 1987;433:155–159. doi: 10.1016/0165-3806(87)90075-7. [DOI] [PubMed] [Google Scholar]
- 62.Mizutani M, Kern TS, Lorenzi M. Accelerated death of retinal microvascular cells in human and experimental diabetic retinopathy. J Clin Invest. 1996;97:2883–2890. doi: 10.1172/JCI118746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Raff MC, Barres BA, Burne JF, Coles HS, Ishizaki Y, Jacobson MD. Programmed cell death and the control of cell survival: lessons from the nervous system. Science. 1993;262:695–700. doi: 10.1126/science.8235590. [DOI] [PubMed] [Google Scholar]
- 64.Rodrigues M, Xin X, Jee K, Babapoor-Farrokhran S, Kashiwabuchi F, Ma T, Bhutto I, Hassan SJ, Daoud Y, Baranano D, Solomon S, Lutty G, Semenza GL, Montaner S, Sodhi A. VEGF secreted by hypoxic Müller cells induces MMP-2 expression and activity in endothelial cells to promote retinal neovascularization in proliferative diabetic retinopathy. Diabetes. 2013;62:3863–3873. doi: 10.2337/db13-0014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Cheng L, Bu H, Portillo JA, Li Y, Subauste CS, Huang SS, Kern TS, Lin F. Modulation of retinal Müller cells by complement receptor C5aR. Invest Ophthalmol Vis Sci. 2013;54:8191–8198. doi: 10.1167/iovs.13-12428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Nakata K, Saitoh R, Amano J, Koshiyama A, Ichibangase T, Murao N, Ohta K, Aso Y, Ishigai M, Imai K. Alteration of intracellular secretory acute phase response proteins expressed in human hepatocyte induced by exposure with interleukin-6. Cytokine. 2012;59:317–323. doi: 10.1016/j.cyto.2012.04.025. [DOI] [PubMed] [Google Scholar]
- 67.Mohammad G, Mairaj Siddiquei M, Imtiaz Nawaz M, Abu El-Asrar AM. The ERK1/2 inhibitor U0126 attenuates diabetes-induced upregulation of MMP-9 and biomarkers of inflammation in the retina. J Diabetes Res. 2013;2013:658548. doi: 10.1155/2013/658548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zhang SX, Wang JJ, Gao G, Shao C, Mott R, Ma JX. Pigment epithelium-derived factor (PEDF) is an endogenous antiinflammatory factor. FASEB J. 2006;20:323–325. doi: 10.1096/fj.05-4313fje. [DOI] [PubMed] [Google Scholar]
- 69.Cornelius LA, Nehring LC, Harding E, Bolanowski M, Welgus HG, Kobayashi DK, Pierce RA, Shapiro SD. Matrix metalloproteinases generate angiostatin: effects on neovascularization. J Immunol. 1998;161:6845–6852. [PubMed] [Google Scholar]
- 70.Salzmann J, Limb GA, Khaw PT, Gregor ZJ, Webster L, Chignell AH, Charteris DG. Matrix metalloproteinases and their natural inhibitors in fibrovascular membranes of proliferative diabetic retinopathy. Br J Ophthalmol. 2000;84:1091–1096. doi: 10.1136/bjo.84.10.1091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.el-Shabrawi Y, Eckhardt M, Berghold A, Faulborn J, Auboeck L, Mangge H, Ardjomand N. Synthesis pattern of matrix metalloproteinases (MMPs) and inhibitors (TIMPs) in human explant organ cultures after treatment with latanoprost and dexamethasone. Eye (Lond) 2000;14(Pt 3A):375–383. doi: 10.1038/eye.2000.92. [DOI] [PubMed] [Google Scholar]
- 72.Padgett LC, Lui GM, Werb Z, LaVail MM. Matrix metalloproteinase-2 and tissue inhibitor of metalloproteinase-1 in the retinal pigment epithelium and interphotoreceptor matrix: vectorial secretion and regulation. Exp Eye Res. 1997;64:927–938. doi: 10.1006/exer.1997.0287. [DOI] [PubMed] [Google Scholar]
- 73.Alexander JP, Bradley JM, Gabourel JD, Acott TS. Expression of matrix metalloproteinases and inhibitor by human retinal pigment epithelium. Invest Ophthalmol Vis Sci. 1990;31:2520–2528. [PubMed] [Google Scholar]
- 74.Santos JM, Tewari S, Lin JY, Kowluru RA. Interrelationship between activation of matrix metalloproteinases and mitochondrial dysfunction in the development of diabetic retinopathy. Biochem Biophys Res Commun. 2013;438:760–764. doi: 10.1016/j.bbrc.2013.07.066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Abu El-Asrar AM, Mohammad G, Nawaz MI, Siddiquei MM, Van den Eynde K, Mousa A, De Hertogh G, Opdenakker G. Relationship between vitreous levels of matrix metalloproteinases and vascular endothelial growth factor in proliferative diabetic retinopathy. PLoS ONE. 2013;8:e85857. doi: 10.1371/journal.pone.0085857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Shitama T, Hayashi H, Noge S, Uchio E, Oshima K, Haniu H, Takemori N, Komori N, Matsumoto H. Proteome profiling of vitreoretinal diseases by cluster analysis. Proteomics Clin Appl. 2008;2:1265–1280. doi: 10.1002/prca.200800017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Gao BB, Chen X, Timothy N, Aiello LP, Feener EP. Characterization of the vitreous proteome in diabetes without diabetic retinopathy and diabetes with proliferative diabetic retinopathy. J Proteome Res. 2008;7:2516–2525. doi: 10.1021/pr800112g. [DOI] [PubMed] [Google Scholar]
- 78.Nenan S, Boichot E, Lagente V, Bertrand CP. Macrophage elastase (MMP-12): a pro-inflammatory mediator? Mem Inst Oswaldo Cruz. 2005;100(Suppl 1):167–172. doi: 10.1590/S0074-02762005000900028. [DOI] [PubMed] [Google Scholar]
- 79.Nicoloff G, Baydanoff S, Stanimirova N, Petrova C, Christova P. An association of anti-elastin IgA antibodies with development of retinopathy in diabetic children. Gen Pharmacol. 2000;35:83–87. doi: 10.1016/S0306-3623(01)00095-7. [DOI] [PubMed] [Google Scholar]
- 80.McMillan DE. Development of vascular complications in diabetes. Vasc Med. 1997;2:132–142. doi: 10.1177/1358863X9700200209. [DOI] [PubMed] [Google Scholar]
- 81.Liu Z, Zhou X, Shapiro SD, Shipley JM, Twining SS, Diaz LA, Senior RM, Werb Z. The serpin alpha1-proteinase inhibitor is a critical substrate for gelatinase B/MMP-9 in vivo. Cell. 2000;102:647–655. doi: 10.1016/S0092-8674(00)00087-8. [DOI] [PubMed] [Google Scholar]
- 82.Muroski ME, Roycik MD, Newcomer RG, Van den Steen PE, Opdenakker G, Monroe HR, Sahab ZJ, Sang QX. Matrix metalloproteinase-9/gelatinase B is a putative therapeutic target of chronic obstructive pulmonary disease and multiple sclerosis. Curr Pharm Biotechnol. 2008;9:34–46. doi: 10.2174/138920108783497631. [DOI] [PubMed] [Google Scholar]
- 83.Zhong Q, Kowluru RA. Regulation of matrix metalloproteinase-9 by epigenetic modifications and the development of diabetic retinopathy. Diabetes. 2013;62:2559–2568. doi: 10.2337/db12-1141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Gaublomme D, Buyens T, De Groef L, Stakenborg M, Janssens E, Ingvarsen S, Porse A, Behrendt N, Moons L. Matrix metalloproteinase 2 and membrane-type 1 matrix metalloproteinase co-regulate axonal outgrowth of mouse retinal ganglion cells. J Neurochem. 2014;129:966–979. doi: 10.1111/jnc.12703. [DOI] [PubMed] [Google Scholar]
- 85.Savinov AY, Strongin AY. Targeting the T-cell membrane type-1 matrix metalloproteinase-CD44 axis in a transferred type 1 diabetes model in NOD mice. Exp Ther Med. 2013;5:438–442. doi: 10.3892/etm.2012.821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Cogan DG, Toussaint D, Kuwabara T. Retinal vascular patterns. IV. Diabetic retinopathy. Arch Ophthalmol. 1961;66:366–378. doi: 10.1001/archopht.1961.00960010368014. [DOI] [PubMed] [Google Scholar]
- 87.Engerman RL. Pathogenesis of diabetic retinopathy. Diabetes. 1989;38:1203–1206. doi: 10.2337/diab.38.10.1203. [DOI] [PubMed] [Google Scholar]
- 88.Bauer HC, Steiner M, Bauer H. Embryonic development of the CNS microvasculature in the mouse: new insights into the structural mechanisms of early angiogenesis. EXS. 1992;61:64–68. doi: 10.1007/978-3-0348-7001-6_11. [DOI] [PubMed] [Google Scholar]
- 89.Betsholtz C. Insight into the physiological functions of PDGF through genetic studies in mice. Cytokine Growth Factor Rev. 2004;15:215–228. doi: 10.1016/j.cytogfr.2004.03.005. [DOI] [PubMed] [Google Scholar]
- 90.Egginton S, Zhou AL, Brown MD, Hudlicka O. The role of pericytes in controlling angiogenesis in vivo. Adv Exp Med Biol. 2000;476:81–99. doi: 10.1007/978-1-4615-4221-6_7. [DOI] [PubMed] [Google Scholar]
- 91.Gerhardt H, Betsholtz C. Endothelial-pericyte interactions in angiogenesis. Cell Tissue Res. 2003;314:15–23. doi: 10.1007/s00441-003-0745-x. [DOI] [PubMed] [Google Scholar]
- 92.Ozerdem U, Stallcup WB. Early contribution of pericytes to angiogenic sprouting and tube formation. Angiogenesis. 2003;6:241–249. doi: 10.1023/B:AGEN.0000021401.58039.a9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Wakui S, Yokoo K, Muto T, Suzuki Y, Takahashi H, Furusato M, Hano H, Endou H, Kanai Y. Localization of Ang-1, −2, Tie-2, and VEGF expression at endothelial-pericyte interdigitation in rat angiogenesis. Lab Invest. 2006;86:1172–1184. doi: 10.1038/labinvest.3700476. [DOI] [PubMed] [Google Scholar]
- 94.Antonelli-Orlidge A, Smith SR, D’Amore PA. Influence of pericytes on capillary endothelial cell growth. Am Rev Respir Dis. 1989;140:1129–1131. doi: 10.1164/ajrccm/140.4.1129. [DOI] [PubMed] [Google Scholar]
- 95.Li W, Liu X, He Z, Yanoff M, Jian B, Ye X. Expression of apoptosis regulatory genes by retinal pericytes after rapid glucose reduction. Invest Ophthalmol Vis Sci. 1998;39:1535–1543. [PubMed] [Google Scholar]
- 96.Naruse K, Nakamura J, Hamada Y, Nakayama M, Chaya S, Komori T, Kato K, Kasuya Y, Miwa K, Hotta N. Aldose reductase inhibition prevents glucose-induced apoptosis in cultured bovine retinal microvascular pericytes. Exp Eye Res. 2000;71:309–315. doi: 10.1006/exer.2000.0882. [DOI] [PubMed] [Google Scholar]
- 97.Otani A, Kinder K, Ewalt K, Otero FJ, Schimmel P, Friedlander M. Bone marrow-derived stem cells target retinal astrocytes and can promote or inhibit retinal angiogenesis. Nat Med. 2002;8:1004–1010. doi: 10.1038/nm744. [DOI] [PubMed] [Google Scholar]
- 98.Ritter MR, Banin E, Moreno SK, Aguilar E, Dorrell MI, Friedlander M. Myeloid progenitors differentiate into microglia and promote vascular repair in a model of ischemic retinopathy. J Clin Invest. 2006;116:3266–3276. doi: 10.1172/JCI29683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Grant MB, May WS, Caballero S, Brown GA, Guthrie SM, Mames RN, Byrne BJ, Vaught T, Spoerri PE, Peck AB, Scott EW. Adult hematopoietic stem cells provide functional hemangioblast activity during retinal neovascularization. Nat Med. 2002;8:607–612. doi: 10.1038/nm0602-607. [DOI] [PubMed] [Google Scholar]
- 100.Sengupta N, Caballero S, Mames RN, Butler JM, Scott EW, Grant MB. The role of adult bone marrow-derived stem cells in choroidal neovascularization. Invest Ophthalmol Vis Sci. 2003;44:4908–4913. doi: 10.1167/iovs.03-0342. [DOI] [PubMed] [Google Scholar]
- 101.Kicic A, Shen WY, Wilson AS, Constable IJ, Robertson T, Rakoczy PE. Differentiation of marrow stromal cells into photoreceptors in the rat eye. J Neurosci. 2003;23:7742–7749. doi: 10.1523/JNEUROSCI.23-21-07742.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Kielczewski JL, Hu P, Shaw LC, Li Calzi S, Mames RN, Gardiner TA, McFarland E, Chan-Ling T, Grant MB. Novel protective properties of IGFBP-3 result in enhanced pericyte ensheathment, reduced microglial activation, increased microglial apoptosis, and neuronal protection after ischemic retinal injury. Am J Pathol. 2011;178:1517–1528. doi: 10.1016/j.ajpath.2010.12.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Chan-Ling T, Baxter L, Afzal A, Sengupta N, Caballero S, Rosinova E, Grant MB. Hematopoietic stem cells provide repair functions after laser-induced Bruch’s membrane rupture model of choroidal neovascularization. Am J Pathol. 2006;168:1031–1044. doi: 10.2353/ajpath.2006.050697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Mendel TA, Clabough EB, Kao DS, Demidova-Rice TN, Durham JT, Zotter BC, Seaman SA, Cronk SM, Rakoczy EP, Katz AJ, Herman IM, Peirce SM, Yates PA. Pericytes derived from adipose-derived stem cells protect against retinal vasculopathy. PLoS ONE. 2013;8:e65691. doi: 10.1371/journal.pone.0065691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.El-Remessy AB, Behzadian MA, Abou-Mohamed G, Franklin T, Caldwell RW, Caldwell RB. Experimental diabetes causes breakdown of the blood-retina barrier by a mechanism involving tyrosine nitration and increases in expression of vascular endothelial growth factor and urokinase plasminogen activator receptor. Am J Pathol. 2003;162:1995–2004. doi: 10.1016/S0002-9440(10)64332-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Qaum T, Xu Q, Joussen AM, Clemens MW, Qin W, Miyamoto K, Hassessian H, Wiegand SJ, Rudge J, Yancopoulos GD, Adamis AP. VEGF-initiated blood-retinal barrier breakdown in early diabetes. Invest Ophthalmol Vis Sci. 2001;42:2408–2413. [PubMed] [Google Scholar]
- 107.Roy MS, Podgor MJ, Bungay P, Grunberger G, Carl J, Ellis D. Posterior vitreous fluorophotometry in diabetic patients with minimal or no retinopathy. Retina. 1987;7:170–176. doi: 10.1097/00006982-198700730-00006. [DOI] [PubMed] [Google Scholar]
- 108.Stitt AW, Bhaduri T, McMullen CB, Gardiner TA, Archer DB. Advanced glycation end products induce blood-retinal barrier dysfunction in normoglycemic rats. Mol Cell Biol Res Commun. 2000;3:380–388. doi: 10.1006/mcbr.2000.0243. [DOI] [PubMed] [Google Scholar]
- 109.Lu M, Kuroki M, Amano S, Tolentino M, Keough K, Kim I, Bucala R, Adamis AP. Advanced glycation end products increase retinal vascular endothelial growth factor expression. J Clin Invest. 1998;101:1219–1224. doi: 10.1172/JCI1277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Yamagishi S, Yonekura H, Yamamoto Y, Katsuno K, Sato F, Mita I, Ooka H, Satozawa N, Kawakami T, Nomura M, Yamamoto H. Advanced glycation end products-driven angiogenesis in vitro. Induction of the growth and tube formation of human microvascular endothelial cells through autocrine vascular endothelial growth factor. J Biol Chem. 1997;272:8723–8730. doi: 10.1074/jbc.272.13.8723. [DOI] [PubMed] [Google Scholar]
- 111.McFarlane S, Glenn JV, Lichanska AM, Simpson DA, Stitt AW. Characterisation of the advanced glycation endproduct receptor complex in the retinal pigment epithelium. Br J Ophthalmol. 2005;89:107–112. doi: 10.1136/bjo.2004.045914. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Canning P, Glenn JV, Hsu DK, Liu FT, Gardiner TA, Stitt AW. Inhibition of advanced glycation and absence of galectin-3 prevent blood-retinal barrier dysfunction during short-term diabetes. Exp Diabetes Res. 2007;2007:51837. doi: 10.1155/2007/51837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Wang AL, Yu AC, He QH, Zhu X, Tso MO. AGEs mediated expression and secretion of TNF alpha in rat retinal microglia. Exp Eye Res. 2007;84:905–913. doi: 10.1016/j.exer.2007.01.011. [DOI] [PubMed] [Google Scholar]