

Abstract
Salinity is one of the most brutal environmental factors limiting the productivity of crop plants because most of the crop plants are sensitive to salinity caused by high concentrations of salts in the soil, and the area of land affected by it is increasing day by day. For all important crops, average yields are only a fraction – somewhere between 20% and 50% of record yields; these losses are mostly due to drought and high soil salinity, environmental conditions which will worsen in many regions because of global climate change. A wide range of adaptations and mitigation strategies are required to cope with such impacts. Efficient resource management and crop/livestock improvement for evolving better breeds can help to overcome salinity stress. However, such strategies being long drawn and cost intensive, there is a need to develop simple and low cost biological methods for salinity stress management, which can be used on short term basis. Microorganisms could play a significant role in this respect, if we exploit their unique properties such as tolerance to saline conditions, genetic diversity, synthesis of compatible solutes, production of plant growth promoting hormones, bio-control potential, and their interaction with crop plants.
Keywords: Salinity, Salt-tolerant, PGPR, Microorganisms
1. Introduction
The beginning of 21st century is marked by global scarcity of water resources, environmental pollution and increased salinization of soil and water. Increasing human population and reduction in land available for cultivation are two threats for agricultural sustainability (Shahbaz and Ashraf, 2013). Various environmental stresses viz. high winds, extreme temperatures, soil salinity, drought and flood have affected the production and cultivation of agricultural crops, among these soil salinity is one of the most devastating environmental stresses, which causes major reductions in cultivated land area, crop productivity and quality (Yamaguchi and Blumwald, 2005; Shahbaz and Ashraf, 2013). A saline soil is generally defined as one in which the electrical conductivity (EC) of the saturation extract (ECe) in the root zone exceeds 4 dS m−1 (approximately 40 mM NaCl) at 25 °C and has an exchangeable sodium of 15%. The yield of most crop plants is reduced at this ECe, though many crops exhibit yield reduction at lower ECes (Munns, 2005; Jamil et al., 2011). It has been estimated that worldwide 20% of total cultivated and 33% of irrigated agricultural lands are afflicted by high salinity. Furthermore, the salinized areas are increasing at a rate of 10% annually for various reasons, including low precipitation, high surface evaporation, weathering of native rocks, irrigation with saline water, and poor cultural practices. It has been estimated that more than 50% of the arable land would be salinized by the year 2050 (Jamil et al., 2011).
Water and soil management practices have facilitated agricultural production on soil marginalized by salinity but an additional gain from these approaches seems problematic (Zahir et al., 2008). Impacted soils are a major limiting production factor worldwide for every major crop (Bacilio et al., 2004; Shannon and Grieve, 1999). A significant increase (an estimated 50%) in grain yields of major crop plants such as rice (Oryza sativa L.), wheat (Triticum aestivum L.) and maize (Zea mays L.) is required to fulfill the food supply requirements for the projected population by 2050 (Godfray et al., 2010). The urgency of feeding the world’s growing population while combating soil pollution, salinization, and desertification has given plant and soil productivity research vital importance. Under such circumstances, it requires suitable biotechnology not only to improve crop productivity but also to improve soil health through interactions of plant roots and soil microorganisms (Lugtenberg et al., 2002).
Salt stressed soils are known to suppress the growth of plants (Paul, 2012). Plants in their natural environment are colonized both by endocellular and intracellular microorganisms (Gray and Smith, 2005). Rhizosphere microorganisms, particularly beneficial bacteria and fungi, can improve plant performance under stress environments and, consequently, enhance yield both directly and indirectly (Dimkpa et al., 2009). Some plant growth-promoting rhizobacteria (PGPR) may exert a direct stimulation on plant growth and development by providing plants with fixed nitrogen, phytohormones, iron that has been sequestered by bacterial siderophores, and soluble phosphate (Hayat et al., 2010). Others do this indirectly by protecting the plant against soil-borne diseases, most of which are caused by pathogenic fungi (Lutgtenberg and Kamilova, 2009). The problem of soil salinization is a scourge for agricultural productivity worldwide. Crops grown on saline soils suffer on an account of high osmotic stress, nutritional disorders and toxicities, poor soil physical conditions and reduced crop productivity. The present review focuses on the enhancement of productivity under stressed conditions and increased resistance of plants against salinity stress by application of plant growth promoting microorganisms.
2. Problem of soil salinization
Soil salinity is an enormous problem for agriculture under irrigation. In the hot and dry regions of the world the soils are frequently saline with low agricultural potential. In these areas most crops are grown under irrigation, and to exacerbate the problem, inadequate irrigation management leads to secondary salinization that affects 20% of irrigated land worldwide (Glick et al., 2007). Irrigated agriculture is a major human activity, which often leads to secondary salinization of land and water resources in arid and semi-arid conditions. Salts in the soil occur as ions (electrically charged forms of atoms or compounds). Ions are released from weathering minerals in the soil. They may also be applied through irrigation water or as fertilizers, or sometimes migrate upward in the soil from shallow groundwater. When precipitation is insufficient to leach ions from the soil profile, salts accumulate in the soil resulting soil salinity (Blaylock et al., 1994). All soils contain some water-soluble salts. Plants absorb essential nutrients in the form of soluble salts, but excessive accumulation strongly suppresses the plant growth. During the last century, physical, chemical and/or biological land degradation processes have resulted in serious consequences to global natural resources (e.g. compaction, inorganic/organic contamination, and diminished microbial activity/diversity). The area under the affected soils continues to increase each year due to introduction of irrigation in new areas (Patel et al., 2011).
Salinization is recognized as the main threats to environmental resources and human health in many countries, affecting almost 1 billion ha worldwide/globally representing about 7% of earth’s continental extent, approximately 10 times the size of a country like Venezuela or 20 times the size of France (Metternicht and Zinck, 2003; Yensen, 2008). It has been estimated that an approximate area of 7 million hectares of land is covered by saline soil in India (Patel et al., 2011). Most of which occurs in indogangetic plane that covers the states of Punjab, Haryana, U.P. Bihar and some parts of Rajasthan. Arid tracts of Gujarat and Rajasthan and semi-arid tracts of Gujarat, Madhya Pradesh, Maharashtra, Karnataka and Andhra Pradesh are also largely affected by saline lands.
3. Impact of salinity on plants
Agricultural crops exhibit a spectrum of responses under salt stress. Salinity not only decreases the agricultural production of most crops, but also, effects soil physicochemical properties, and ecological balance of the area. The impacts of salinity include—low agricultural productivity, low economic returns and soil erosions, (Hu and Schmidhalter, 2002). Salinity effects are the results of complex interactions among morphological, physiological, and biochemical processes including seed germination, plant growth, and water and nutrient uptake (Akbarimoghaddam et al., 2011; Singh and Chatrath, 2001). Salinity affects almost all aspects of plant development including: germination, vegetative growth and reproductive development. Soil salinity imposes ion toxicity, osmotic stress, nutrient (N, Ca, K, P, Fe, Zn) deficiency and oxidative stress on plants, and thus limits water uptake from soil. Soil salinity significantly reduces plant phosphorus (P) uptake because phosphate ions precipitate with Ca ions (Bano and Fatima, 2009). Some elements, such as sodium, chlorine, and boron, have specific toxic effects on plants. Excessive accumulation of sodium in cell walls can rapidly lead to osmotic stress and cell death (Munns, 2002). Plants sensitive to these elements may be affected at relatively low salt concentrations if the soil contains enough of the toxic element. Because many salts are also plant nutrients, high salt levels in the soil can upset the nutrient balance in the plant or interfere with the uptake of some nutrients (Blaylock et al., 1994). Salinity also affects photosynthesis mainly through a reduction in leaf area, chlorophyll content and stomatal conductance, and to a lesser extent through a decrease in photosystem II efficiency (Netondo et al., 2004). Salinity adversely affects reproductive development by inhabiting microsporogenesis and stamen filament elongation, enhancing programed cell death in some tissue types, ovule abortion and senescence of fertilized embryos. The saline growth medium causes many adverse effects on plant growth, due to a low osmotic potential of soil solution (osmotic stress), specific ion effects (salt stress), nutritional imbalances, or a combination of these factors (Ashraf, 2004). All these factors cause adverse effects on plant growth and development at physiological and biochemical levels (Munns and James, 2003), and at the molecular level (Tester and Davenport, 2003).
In order to assess the tolerance of plants to salinity stress, growth or survival of the plant is measured because it integrates the up- or down-regulation of many physiological mechanisms occurring within the plant. Osmotic balance is essential for plants growing in saline medium. Failure of this balance results in loss of turgidity, cell dehydration and ultimately, death of cells. On the other hand, adverse effects of salinity on plant growth may also result from impairment of the supply of photosynthetic assimilates or hormones to the growing tissues (Ashraf, 2004). Ion toxicity is the result of replacement of K+ by Na+ in biochemical reactions, and Na+ and Cl− induced conformational changes in proteins. For several enzymes, K+ acts as cofactor and cannot be substituted by Na+. High K+ concentration is also required for binding tRNA to ribosomes and thus protein synthesis (Zhu, 2002). Ion toxicity and osmotic stress cause metabolic imbalance, which in turn leads to oxidative stress (Chinnusamy et al., 2006). The adverse effects of salinity on plant development are more profound during the reproductive phase. Wheat plants stressed at 100–175 mM NaCl showed a significant reduction in spikelets per spike, delayed spike emergence and reduced fertility, which results in poor grain yields. However, Na+ and Cl− concentrations in the shoot apex of these wheat plants were below 50 and 30 mM, respectively, which is too low to limit metabolic reactions (Munns and Rawson, 1999). Hence, the adverse effects of salinity may be attributed to the salt-stress effect on the cell cycle and differentiation. Salinity arrests the cell cycle transiently by reducing the expression and activity of cyclins and cyclin-dependent kinases that results in fewer cells in the meristem, thus limiting growth. The activity of cyclin-dependent kinase is diminished also by post-translational inhibition during salt stress. Recent reports also show that salinity adversely affects plant growth and development, hindering seed germination, seedling growth, enzyme activity (Seckin et al., 2009), DNA, RNA, protein synthesis and mitosis (Tabur and Demir, 2010; Javid et al., 2011).
4. Amelioration of salinity
Salinization can be restricted by leaching of salt from root zone, changed farm management practices and use of salt tolerant plants. Irrigated agriculture can be sustained by better irrigation practices such as adoption of partial root zone drying methodology, and drip or micro-jet irrigation to optimize use of water. The spread of dry land salinity can be contained by reducing the amount of water passing beyond the roots. This can be done by re-introducing deep rooted perennial plants that continue to grow and use water during the seasons that do not support annual crop plants. This may restore the balance between rainfall and water use, thus preventing rising water tables and the movement of salt to the soil surface (Manchanda and Garg, 2008). Farming systems can change to incorporate perennials in rotation with annual crops (phase farming), in mixed plantings (alley farming, intercropping), or in site-specific plantings (precision farming) (Munns et al., 2002). Although the use of these approaches to sustainable management can ameliorate yield reduction under salinity stress, implementation is often limited because of cost and availability of good water quality or water resource. Evolving efficient, low cost, easily adaptable methods for the abiotic stress management is a major challenge. Worldwide, extensive research is being carried out, to develop strategies to cope with abiotic stresses, through development of salt and drought tolerant varieties, shifting the crop calendars, resource management practices etc. (Venkateswarlu and Shanker, 2009) as shown in Fig. 1.
Figure 1.
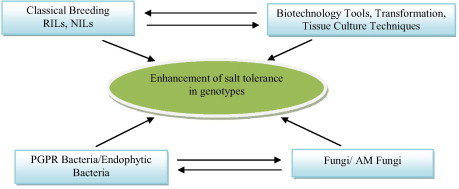
Different approaches for improvement of salt tolerance in agricultural crops.
5. Use of salt tolerant crops and transgenics
Using the salt-tolerant crops is one of the most important strategies to solve the problem of salinity. Tolerance will be required for the “de-watering” species, but also for the annual crops to follow, as salt will be left in the soil when the water table is lowered. Salt tolerance in crops will also allow the more effective use of poor quality irrigation water. To increase the plant salt-tolerance, there is a need for understanding the mechanisms of salt limitation on plant growth and the mechanism of salt tolerance at the whole-plant, organelle, and molecular levels. Under saline conditions, there is a change in the pattern of gene expression, and both qualitative and quantitative changes in protein synthesis. Although it is generally agreed that salt stress brings about quantitative changes in protein synthesis, there is some controversy as to whether salinity activates specialized genes that are involved in salt stress. Salt tolerance does not appear to be conferred by unique gene(s) (Manchanda and Garg, 2008). When a plant is subjected to abiotic stress, a number of genes are turned on, resulting in increased levels of several metabolites and proteins, some of which may be responsible for conferring a certain degree of protection to these stresses (Bhatnagar-Mathur et al., 2008). Efforts to improve crop performance by transgenic approach under environmental stresses have not been that fruitful because the fundamental mechanisms of stress tolerance in plants remain to be completely understood.
Development of salt-tolerant crops has been a major objective of plant breeding programs for decades in order to maintain crop productivity in semiarid and saline lands. Although several salt-tolerant varieties have been released, the overall progress of traditional breeding has been slow and has not been successful as only few major determinant genetic traits of salt tolerance have been identified (Schubert et al., 2009; Dodd and Perez-Alfocea, 2012). 25 years ago Epstein et al. (1980) described the technical and biological constraints to solving the problem of salinity. Although there has been some success with technical solutions to the problem, the biological solutions have been more difficult to develop because a pre-requisite for the development of salt tolerant crops is the identification of key genetic determinants of stress tolerance. The existence of salt-tolerant plants (halophytes) and differences in salt tolerance between genotypes within salt-sensitive plant species (glycophytes) indicates that there is a genetic basis to salt response (Yamaguchi and Blumwald, 2005). Although a lot of approaches have been done for development of salt tolerant plants by transgenics complete success is not achieved yet. The assessment of salt tolerance in transgenic experiments has been mostly carried out using a limited number of seedlings or mature plants in laboratory experiments. In most of the cases, the experiments were carried out in greenhouse conditions where the plants were not exposed to those conditions that prevail in high-salinity soils (e.g. alkaline soil pH, high diurnal temperatures, low humidity, and presence of other sodic salts and elevated concentrations of selenium and/or boron). The salt tolerance of the plants in the field needs to be evaluated and, more importantly, salt tolerance needs to be evaluated as a function of yield. The evaluation of field performance under salt stress is difficult because of the variability of salt levels in field conditions (Richards, 1983) and the potential for interactions with other environmental factors, including soil fertility, temperature, light intensity and water loss due to transpiration. Evaluating tolerance is also made more complex because of variation in sensitivity to salt during the life cycle. For example, in rice, grain yield is much more affected by salinity than in vegetative growth (Khatun and Flowers, 1995). In tomato, the ability of the plants to germinate under conditions of high salinity is not always correlated with the ability of the plant to grow under salt stress because both are controlled by different mechanisms (Foolad and Lin, 1997), although some genotypes might display similar tolerance at germination and during vegetative growth (Foolad and Chen, 1999). Therefore, the assessment of stress tolerance in the laboratory often has little correlation to tolerance in the field. Although there have been many successes in developing stress-tolerant transgenics in model plants such as tobacco, Arabidopsis or rice (Grover et al., 2003), there is an urgent need to test these successes in other crops. There are several technical and financial challenges associated with transforming many of the crop plants, particularly the monocots. First, transformation of any monocot other than rice is still not routine and to develop a series of independent homozygous lines is costly, both in terms of money and time. Second, the stress tolerance screens will need to include a field component because many of the stress tolerance assays used by basic researchers involve using nutrient-rich media (which in some cases include sucrose). This type of screen is unlikely to have a relationship to field performance. Third, because saline soils are often complex and can include NaCl, CaCl2, CaSO4, Na2SO4, high boron concentrations and alkaline pH, plants that show particular promise will eventually have to be tested in all these environments (Joseph and Jini, 2010).
6. Microbes: abiotic stress alleviation tool in crops
Several strategies have been developed in order to decrease the toxic effects caused by high salinity on plant growth, including plant genetic engineering (Wang et al., 2003), and recently the use of plant growth-promoting bacteria (PGPB) (Dimkpa et al., 2009). The role of microorganisms in plant growth promotion, nutrient management and disease control is well known and well established. These beneficial microorganisms colonize the rhizosphere/endorhizosphere of plants and promote growth of the plants through various direct and indirect mechanisms (Nia et al., 2012; Ramadoss et al., 2013). Previous studies suggest that utilization of PGPB has become a promising alternative to alleviate plant stress caused by salinity (Yao et al., 2010) and the role of microbes in the management of biotic and abiotic stresses is gaining importance. The subject of PGPR elicited tolerance to abiotic stresses has been reviewed recently (Dodd and Perez-Alfocea, 2012; Yang et al., 2009).
The term Induced Systemic Tolerance (IST) has been proposed for PGPR-induced physical and chemical changes that result in enhanced tolerance to abiotic stress. PGPR facilitate plant growth indirectly by reducing plant pathogens, or directly by facilitating the nutrient uptake through phytohormone production (e.g. auxin, cytokinin and gibberellins), by enzymatic lowering of plant ethylene levels and/or by production of siderophores (Kohler et al., 2006). It has been demonstrated that inoculations with AM (arbuscular mycorrhizal) fungi improves plant growth under salt stress (Cho et al., 2006). Kohler et al., 2006 demonstrated the beneficial effect of PGPR Pseudomonas mendocina strains on stabilization of soil aggregate. The three PGPR isolates P. alcaligenes PsA15, Bacillus polymyxa BcP26 and Mycobacterium phlei MbP18 were able to tolerate high temperatures and salt concentrations and thus confer on them potential competitive advantage to survive in arid and saline soils such as calcisol (Egamberdiyeva, 2007). Kohler et al., 2009 investigated the influence of inoculation with a PGPR, P. mendocina, alone or in combination with an AM fungus, Glomus intraradices or G. mosseae on growth and nutrient uptake and other physiological activities of Lactuca sativa affected by salt stress. The plants inoculated with P. mendocina had significantly greater shoot biomass than the controls and it is suggested that inoculation with selected PGPR could be an effective tool for alleviating salinity stress in salt sensitive plants. Bacteria isolated from different stressed habitats possess stress tolerance capacity along with the plant growth-promoting traits and therefore are potential candidates for seed bacterization. When inoculated with these isolates, plants show enhanced root and shoot length, biomass, and biochemical levels such as chlorophyll, carotenoids, and protein (Tiwari et al., 2011). Investigations on interaction of PGPR with other microbes and their effect on the physiological response of crop plants under different soil salinity regimes are still in incipient stage. Inoculations with selected PGPR and other microbes could serve as the potential tool for alleviating salinity stress in salt sensitive crops. Therefore, an extensive investigation is needed in this area, and the use of PGPR and other symbiotic microorganisms, can be useful in developing strategies to facilitate sustainable agriculture in saline soils.
7. Alleviation of abiotic stress in plants by rhizospheric bacteria
Besides developing mechanisms for stress tolerance, microorganisms can also impart some degree of tolerance to plants towards abiotic stresses like drought, chilling injury, salinity, metal toxicity and high temperature. In the last decade, bacteria belonging to different genera including Rhizobium, Bacillus, Pseudomonas, Pantoea, Paenibacillus, Burkholderia, Achromobacter, Azospirillum, Microbacterium, Methylobacterium, Variovorax, Enterobacter etc. have been reported to provide tolerance to host plants under different abiotic stress environments (Grover et al., 2011). Use of these microorganisms per se can alleviate stresses in agriculture thus opening a new and emerging application of microorganisms. Microbial elicited stress tolerance in plants may be due to a variety of mechanisms proposed from time to time based on studies done. Production of indole acetic acid, gibberellins and some unknown determinants by PGPR, results in increased root length, root surface area and number of root tips, leading to an enhanced uptake of nutrients thereby improving plant health under stress conditions (Egamberdieva and Kucharova, 2009). Plant growth promoting bacteria have been found to improve growth of tomato, pepper, canola, bean and lettuce under saline conditions (Barassi et al., 2006; Yildirim and Taylor, 2005).
Some PGPR strains produce cytokinin and antioxidants, which result in abscisic acid (ABA) accumulation and degradation of reactive oxygen species. High activities of antioxidant enzymes are linked with oxidative stress tolerance (Stajner et al., 1997). Another PGPR strain, Achromobacter piechaudii ARV8 which produced 1-aminocyclopropane-1-carboxylate (ACC) deaminase, conferred IST against drought and salt in pepper and tomato (Mayak et al., 2004). Many aspects of plant life are regulated by ethylene levels and the biosynthesis of ethylene is subjected to tight regulation, involving transcriptional and post-transcriptional factors regulated by environmental cues, including biotic and abiotic stresses (Hardoim et al., 2008). Under stress conditions, the plant hormone ethylene endogenously regulates plant homoeostasis and results in reduced root and shoot growth. In the presence of ACC deaminase producing bacteria, plant ACC is sequestered and degraded by bacterial cells to supply nitrogen and energy. Furthermore, by removing ACC, the bacteria reduce the deleterious effect of ethylene, ameliorating stress and promoting plant growth (Glick, 2007). The complex and dynamic interactions among microorganisms, roots, soil and water in the rhizosphere induce changes in physicochemical and structural properties of the soil (Haynes and Swift, 1990). Microbial polysaccharides can bind soil particles to form microaggregates and macroaggregates. Plant roots and fungal hyphae fit in the pores between microaggregates and thus stabilize macroaggregates. Plants treated with Exo-poly saccharides (EPS) producing bacteria display increased resistance to water and salinity stress due to improved soil structure (Sandhya et al., 2009). EPS can also bind to cations including Na+ thus making it unavailable to plants under saline conditions. Chen et al., 2007 correlated proline accumulation with drought and salt tolerance in plants. Introduction of proBA genes derived from B. subtilis into A. thaliana resulted in production of higher levels of free proline resulting in increased tolerance to osmotic stress in the transgenic plants. Increased production of proline along with decreased electrolyte leakage, maintenance of relative water content of leaves and selective uptake of K ions resulted in salt tolerance in Zea mays coinoculated with Rhizobium and Pseudomonas (Bano and Fatima, 2009). Rhizobacteria inhabiting the sites exposed to frequent stress conditions, are likely to be more adaptive or tolerant and may serve as better plant growth promoters under stressful conditions. Moreover Yao et al., 2010 reported that inoculation with P. putida Rs 198 promoted cotton growth and germination under conditions of salt stress. Tank and Saraf (2010) showed that PGPRs which are able to solubilize phosphate, produce phytohormones and siderophores in salt condition promote growth of tomato plants under 2% NaCl stress.
In a study carried out by Naz et al., 2009, it was shown that strains isolated from Khewra salt range of Pakistan exhibited their tolerance when tested on saline media simulated by rhizosphere soil filtrate. Noteworthy, the isolates produced ABA in a concentration much higher than that of previous reports. Furthermore production of proline, shoot/root length, and dry weight was also higher in soybean plants inoculated with these isolates under induced salt stress. Likewise Upadhyay et al., 2011 studied the impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions and reported that co-inoculation with B. subtilis and Arthrobacter sp. could alleviate the adverse effects of soil salinity on wheat growth with an increase in dry biomass, total soluble sugars and proline content. Jha et al., 2011 reported that P. pseudoalcaligenes, an endophytic bacterium in combination with a rhizospheric B. pumilus in paddy was able to protect the plant from abiotic stress by induction of osmoprotectant and antioxidant proteins than by the rhizospheric or endophytic bacteria alone at early stages of growth. Plants inoculated with endophytic bacterium P. pseudoalcaligenes showed a significantly higher concentration of glycine betaine-like quaternary compounds and higher shoot biomass at lower salinity levels. While at higher salinity levels, a mixture of both P. pseudoalcaligenes and B. pumilus showed better response against the adverse effects of salinity. Nia et al., 2012 studied the effect of inoculation of Azospirillum strains isolated from saline or non-saline soil on yield and yield components of wheat in salinity and they observed that inoculation with the two isolates increased salinity tolerance of wheat plants; the saline-adapted isolate significantly increased shoot dry weight and grain yield under severe water salinity. The component of grain yield most affected by inoculation was grains per plant. Plants inoculated with saline-adapted Azospirillum strains had higher N concentrations at all water salinity levels.
Sadeghi et al., 2012 studied the plant growth promoting activity of an auxin and siderophore producing isolate of Streptomyces under saline soil conditions and reported increases in growth and development of wheat plant. They observed significant increases in germination rate, percentage and uniformity, shoot length and dry weight compared to the control. Applying the bacterial inocula increased the concentration of N, P, Fe and Mn in wheat shoots grown in normal and saline soil and thus concluded that Streptomyces isolate has potential to be utilized as biofertilizers in saline soils. More recently Ramadoss et al., 2013 studied the effect of five plant growth promoting halotolerant bacteria on wheat growth and found that inoculation of those halotolerant bacterial strains to ameliorate salt stress (80, 160 and 320 mM) in wheat seedlings produced an increase in root length of 71.7% in comparison with uninoculated positive controls. In particular, Hallobacillus sp. and B. halodenitrificans showed more than 90% increase in root elongation and 17.4% increase in dry weight when compared to uninoculated wheat seedlings at 320 mM NaCl stress indicating a significant reduction of the deleterious effects of NaCl. These results indicate that halotolerant bacteria isolated from saline environments have potential to enhance plant growth under saline stress through direct or indirect mechanisms and would be most appropriate as bioinoculants under such conditions. The isolation of indigenous microorganisms from the stress affected soils and screening on the basis of their stress tolerance and PGP traits may be useful in the rapid selection of efficient strains that could be used as bioinoculants for stressed crops. Some of the advances and researches carried out in evaluating role of rhizobacteria as salinity stress remediators have been summarized in Table 1.
Table 1.
Role of plant growth promoting bacteria in salinity stress alleviation in plants.
| Plant growth promoting bacterial species | Crop plant | Effect | References |
|---|---|---|---|
| Achromobacter piechaudii | Tomato (Lycopersicon esculentum) | Reduced levels of ethylene and improved plant growth | Mayak et al. (2004) |
| Azospirillum | Maize (Zea mays) | Restricted Na + uptake and increased K + and Ca2 + uptake along with increased nitrate reductase and nitrogenase activity | Hamdia et al. (2004) |
| Aeromonas hydrophila/caviae Bacillus insolitus, Bacillus sp. | Wheat (Triticum aestivum) | Exopolysaccharide production | Ashraf (2004) |
| Pseudomonas syringae, Pseudomonas fluorescens, Enterobacter aerogenes | Maize (Zea mays) | ACC deaminase activity | Nadeem et al. (2007) |
| Pseudomonas fluorescens | Groundnut (Arachis hypogea) | Enhanced ACC deaminase activity | Saravanakumar and Samiyappan (2007) |
| Bacillus subtilis | Arabidopsis thaliana | Tissue specific regulation of sodium transporter HKT1 | Zhang et al. (2008) |
| Pseudomonas mendocina | Lettuce (L. sativa L. cv. Tafalla) | ACC deaminase activity and enhanced uptake of essential nutrients | Kohler et al. (2009) |
| Rhizobium, Pseudomonas | Maize | Decreased electrolyte leakage and, increase in proline production, maintenance of relative water content of leaves, and selective uptake of K ion | Bano and Fatima (2009) |
| Pseudomonas pseudoalcaligenes, Bacillus pumilus | Rice (Oryza sativa) | Increased concentration of glycine betaine (compatible solute) | Jha et al. (2011) |
| Pseudomonas putida Rs-198 | Cotton | Increase the absorption of the Mg2+, K+ and Ca2+ and decrease the uptake of the Na2+ from the soil | Yao et al. (2010) |
| PGPR (Mk1, Pseudomonas syringae; Mk20, Pseudomonas fluorescens; and Mk25, Pseudomonas fluorescens biotype G) and Rhizobium phaseoli strains M1, M6, and M9 | Mung bean | ACC deaminase activity and improvement in growth and nodulation in mung bean | Ahmad et al. (2011) |
| Raoultella planticola Rs-2 | Cotton | ACC deaminase activity | Wu et al. (2012) |
| Brachybacterium saurashtrense (JG-06), Brevibacterium casei (JG-08), and Haererohalobacter (JG-11) | Groundnut (Arachis hypogaea L.) | High K+/Na+ ratio and higher Ca2+, phosphorus, and nitrogen content | Shukla et al. (2012) |
| Rhizobium phaseoli and PGPR (Pseudomonas syringae, Mk1; Pseudomonas fluorescens, Mk20 and Pseudomonas fluorescens Biotype G, Mk25) | Mung bean (Vigna radiata L.) | ACC deaminase activity and increased water use efficiency | Ahmad et al. (2012) |
| Rhizobium and Pseudomonas | Mung bean (Vigna radiata L.) | IAA production and ACC deaminase activity | Ahmad et al. (2013) |
| Pseudomonas putida, Enterobacter cloacae, Serratia ficaria, and Pseudomonas fluorescens | Wheat | Enhanced germination percentage, germination rate, and index and improved the nutrient status of the wheat plants | Nadeem et al. (2013) |
| Pseudomonas pseudoalcaligenes and Bacillus pumilus | Salt sensitive rice GJ-17 | Reduce lipid peroxidation and superoxide dismutase activity | Jha and Subramanian, 2014 |
| Acinetobacter spp. and Pseudomonas sp. | Barley and oats | Production of ACC deaminase and IAA | Chang et al. (2014) |
| Streptomyces sp. strain PGPA39 | ‘Micro tom’ tomato | ACC deaminase activity and IAA production and phosphate solubilization | Palaniyandi et al. (2014) |
8. Conclusion
An ideal sustainable agricultural system is one which maintains and improves human health, benefits producers and consumers both economically and spiritually, protects the environment, and produces enough food for an increasing world population. One of the most important constraints to agricultural production in world is abiotic stress conditions prevailing in the environment. Plant-associated microorganisms can play an important role in conferring resistance to abiotic stresses. These organisms could include rhizoplane, rhizosphere and endophytic bacteria and symbiotic fungi and operate through a variety of mechanisms like triggering osmotic response, providing growth hormones and nutrients, acting as biocontrol agents and induction of novel genes in plants. The development of stress tolerant crop varieties through genetic engineering and plant breeding is essential but a long drawn and expensive process, whereas microbial inoculation to alleviate stresses in plants could be a more cost effective environmental friendly option which could be available in a shorter time frame. Taking the current leads available, concerted future research is needed in this area, particularly on field evaluation and application of potential organisms as biofertilizers in stressed soil.
Footnotes
Peer review under responsibility of King Saud University.
References
- Ahmad M., Zahir Zahir A., Naeem Asghar H., Asghar M. Inducing salt tolerance in mung bean through coinoculation with rhizobia and plant-growth-promoting rhizobacteria containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2011;57(7):578–589. doi: 10.1139/w11-044. [DOI] [PubMed] [Google Scholar]
- Ahmad M., Zahir Z.A., Asghar H.N., Arshad M. The combined application of rhizobial strains and plant growth promoting rhizobacteria improves growth and productivity of mung bean (Vigna radiata L.) under salt-stressed conditions. Ann. Microbiol. 2012;62:1321–1330. [Google Scholar]
- Ahmad M., Zahir Zahir A., Nazli Farheen, Akram Fareeha, Muhammad Arshad Khalid M. Effectiveness of halo-tolerant, auxin producing pseudomonas and Rhizobium strains to improve osmotic stress tolerance in mung bean (Vigna radiata L.) Braz. J. Microbiol. 2013;44(4):1341–1348. doi: 10.1590/s1517-83822013000400045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akbarimoghaddam H., Galavi M., Ghanbari A., Panjehkeh N. Salinity effects on seed germination and seedling growth of bread wheat cultivars. Trakia J. Sci. 2011;9(1):43–50. [Google Scholar]
- Ashraf M. Some important physiological selection criteria for salt tolerance in plants. Flora. 2004;199:361–376. [Google Scholar]
- Bacilio M., Rodriguez H., Moreno M., Hernandez Juan-Pablo, Bashan Y. Mitigation of salt stress in wheat seedlings by a gfp-tagged Azospirillum lipoferum. Biol. Fertility Soils. 2004;40:188–193. [Google Scholar]
- Bano A., Fatima M. Salt tolerance in Zea mays (L.) following inoculation with Rhizobium and Pseudomonas. Biol. Fertility Soils. 2009;45:405–413. [Google Scholar]
- Barassi C.A., Ayrault G., Creus C.M., Sueldo R.J., Sobero M.T. Seed inoculation with Azospirillum mitigates NaCl effects on lettuce. Sci. Horticulturae (Amsterdam) 2006;109:8–14. [Google Scholar]
- Bhatnagar-Mathur P., Vadez V., Sharma K.K. Transgenic approaches for abiotic stress tolerance in plants: retrospect and prospects. Plant Cell Rep. 2008;27:411–424. doi: 10.1007/s00299-007-0474-9. [DOI] [PubMed] [Google Scholar]
- Blaylock, A.D., 1994. Soil salinity, salt tolerance and growth potential of horticultural and landscape plants. Co-operative Extension Service, University of Wyoming, Department of Plant, Soil and Insect Sciences, College of Agriculture, Laramie, Wyoming.
- Chang P., Gerhardt K.E., Huang Xiao-Dong, Yu Xiao-Ming, Glick B.R., Gerwing P.D., Greenberg B.M. Plant growth-promoting bacteria facilitate the growth of barley and oats in salt-impacted soil: implications for phytoremediation of saline soils. Int. J. Phytorem. 2014;16(11):1133–1147. doi: 10.1080/15226514.2013.821447. [DOI] [PubMed] [Google Scholar]
- Chen M., Wei H., Cao J., Liu R., Wang Y., Zheng C. Expression of Bacillus subtilis proAB genes and reduction of feedback inhibition of proline synthesis increases proline production and confers osmotolerance in transgenic Arabdopsis. J. Biochem. Mol. Biol. 2007;40(3):396–403. doi: 10.5483/bmbrep.2007.40.3.396. [DOI] [PubMed] [Google Scholar]
- Chinnusamy V., Zhu J., Zhu Jian-Kang. Gene regulation during cold acclimation in plants. Physiol. Plant. 2006;126(1):52–61. [Google Scholar]
- Cho K., Toler H., Lee J., Owenley B., Stutz J.C., Moore J.L., Auge R.M. Mycorrhizal symbiosis and response of sorghum plants to combined drought and salinity stresses. J. Plant Physiol. 2006;163:517–528. doi: 10.1016/j.jplph.2005.05.003. [DOI] [PubMed] [Google Scholar]
- Dimkpa C., Weinand T., Ash F. Plant-rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 2009;32:1682–1694. doi: 10.1111/j.1365-3040.2009.02028.x. [DOI] [PubMed] [Google Scholar]
- Dodd I.C., Perez-Alfocea F. Microbial amelioration of crop salinity stress. J. Exp. Bot. 2012;63(9):3415–3428. doi: 10.1093/jxb/ers033. [DOI] [PubMed] [Google Scholar]
- Egamberdieva D., Kucharova Z. Selection for root colonizing bacteria stimulating wheat growth in saline soils. Biol. Fertility Soil. 2009;45:563–571. [Google Scholar]
- Egamberdiyeva D. The effect of plant growth promoting bacteria on growth and nutrient uptake of maize in two different soils. Appl. Soil Ecol. 2007;36:184–189. [Google Scholar]
- Epstein E.J.D., Norlyn D.W., Rush R.W., Kinsbury D.B., Kelly G.A., Cunningham, Wrona A.F. Saline culture of crops: a genetic approach. Science. 1980;210:399–404. doi: 10.1126/science.210.4468.399. [DOI] [PubMed] [Google Scholar]
- Foolad M.R., Chen F.Q. RFLP mapping of QTLs conferring salt tolerance during the vegetative stage in tomato. Theor. Appl. Genet. 1999;99:235–243. [Google Scholar]
- Foolad M.R., Lin G.Y. Absence of a genetic relationship between salt tolerance during seed germination and vegetative growth in tomato. Plant Breed. 1997;116:363–367. [Google Scholar]
- Glick B.R. Promotion of plant growth by bacterial ACC deaminase. Crit. Rev. Plant Sci. 2007;26:227–242. [Google Scholar]
- Glick B.R., Cheng Z., Czarny J., Duan J. Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur. J. Plant Pathol. 2007;119:329–339. [Google Scholar]
- Godfray H.C.J., Beddington J.R., Crute I.R., Haddad L., Lawrence D., Muir J.F., Pretty J., Robinson S., Thomas S.M., Toulmin C. Food security: the challenge of feeding 9 billion people. Science. 2010;327:812–818. doi: 10.1126/science.1185383. [DOI] [PubMed] [Google Scholar]
- Gray E.J., Smith D.L. Intracellular and extracellular PGPR: commonalities and distinctions in the plant-bacterium signalling processes. Soil Biol. Biochem. 2005;37:395–412. [Google Scholar]
- Grover A., Aggarwal P.K., Kapoor A., Katiyar-Agarwal S., Agarwal M., Chandramouli A. Addressing abiotic stresses in agriculture through transgenic technology. Curr. Sci. 2003;84:355–367. [Google Scholar]
- Grover M., Ali Sk.Z., Sandhya V., Rasul A., Venkateswarlu B. Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World J. Microbiol. Biotechnol. 2011;27:1231–1240. [Google Scholar]
- Hamdia M.A., Shaddad M.A.K., Doaa M.M. Mechanism of salt tolerance and interactive effect of Azospirillum bransilense inoculation on maize cultivars grown under salt stress conditions. Plant Growth Regul. 2004;44:165–174. [Google Scholar]
- Hardoim P.R., van Overbeek S.V., van Elsas J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008;16:463–471. doi: 10.1016/j.tim.2008.07.008. [DOI] [PubMed] [Google Scholar]
- Hayat R., Ali S., Amara U., Khalid R., Ahmed I. Soil beneficial bacteria and their role in plant growth promotion: a review. Ann. Microbiol. 2010;60:579–598. [Google Scholar]
- Haynes R.J., Swift R.S. Stability of soil aggregates in relation to organic constituents and soil water content. J. Soil Sci. 1990;41:73–83. [Google Scholar]
- Hu Y., Schmidhalter U. Limitation of salt stress to plant growth. In: Hock B., Elstner C.F., editors. Plant Toxicology. Marcel Dekker Inc.; New York: 2002. pp. 91–224. [Google Scholar]
- Jamil A., Riaz S., Ashraf M., Foolad M.R. Gene expression profiling of plants under salt stress. Crit. Rev. Plant Sci. 2011;30(5):435–458. [Google Scholar]
- Javid M.G., Sorooshzadeh A., Moradi F., Sanavy Seyed A.M.M., Allahdadi I. The role of phytohormones in alleviating salt stress in crop plants. AJCS. 2011;5(6):726–734. [Google Scholar]
- Jha Y., Subramanian R.B. PGPR regulate caspase-like activity, programmed cell death, and antioxidant enzyme activity in paddy under salinity. Physiol. Mol. Biol. Plants. 2014;20(2):201–207. doi: 10.1007/s12298-014-0224-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jha Y., Subramanian R.B., Patel S. Combination of endophytic and rhizospheric plant growth promoting rhizobacteria in Oryza sativa shows higher accumulation of osmoprotectant against saline stress. Acta Physiol. Plant. 2011;33:797–802. [Google Scholar]
- Joseph B., Jini D. Salinity induced programmed cell death in plants: challenges and opportunities for salt tolerant plants. J. Plant Sci. 2010:1–15. [Google Scholar]
- Khatun S., Flowers T.J. Effects of salinity on seed set in rice. Plant Cell Environ. 1995;18:61–67. [Google Scholar]
- Kohler J., Caravaca F., Carrasco L., Roldan A. Contribution of Pseudomonas mendocina and Glomus intraradices to aggregates stabilization and promotion of biological properties in rhizosphere soil of lettuce plants under field conditions. Soil Use Manage. 2006;22:298–304. [Google Scholar]
- Kohler J., Hernandez J.A., Caravaca F., Roldan A. Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ. Exp. Bot. 2009;65:245–252. [Google Scholar]
- Lugtenberg B., Chin-A-Woeng T., Bloemberg G. Microbe-plant interactions: principles and mechanisms. Antonie Van Leeuwenhoek. 2002;81:373–383. doi: 10.1023/a:1020596903142. [DOI] [PubMed] [Google Scholar]
- Lutgtenberg B., Kamilova F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009;63:541–556. doi: 10.1146/annurev.micro.62.081307.162918. [DOI] [PubMed] [Google Scholar]
- Manchanda G., Garg N. Salinity and its effects on the functional biology of legumes. Acta Physiol. Plant. 2008;30:595–618. [Google Scholar]
- Mayak S., Tirosh T., Glick B.R. Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol. Biochem. 2004;42:565–572. doi: 10.1016/j.plaphy.2004.05.009. [DOI] [PubMed] [Google Scholar]
- Metternicht G.I., Zinck J.A. Remote sensing of soil salinity: potentials and constraints. Remote Sens. Environ. 2003;85:1–20. [Google Scholar]
- Munns R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002;25:239–250. doi: 10.1046/j.0016-8025.2001.00808.x. [DOI] [PubMed] [Google Scholar]
- Munns R. Genes and salt tolerance: bringing them together. New Phytol. 2005;167:645–663. doi: 10.1111/j.1469-8137.2005.01487.x. [DOI] [PubMed] [Google Scholar]
- Munns R., James R.A. Screening methods for salinity tolerance: a case study with tetraploid wheat. Plant Soil. 2003;253:201–218. [Google Scholar]
- Munns R., Rawson H.M. Effect of salinity on salt accumulation and reproductive development in the apical meristem of wheat and barley. Aust. J. Plant Physiol. 1999;26:459–464. [Google Scholar]
- Munns R., Husain S., Rivelli A.R., James R.A., Condon A.G., Lindsay M.P., Lagudah E.S., Schachtman D.P., Hare R.A. Avenues for increasing salt tolerance of crops, and the role of physiologically based selection traits. Plant Soil. 2002;247(1):93–105. [Google Scholar]
- Nadeem S.M., Zahir Z.A., Naveed M., Arshad M. Preliminary investigation on inducing salt tolerance in maize through inoculation with rhizobacteria containing ACC-deaminase activity. Can. J. Microbiol. 2007;53:1141–1149. doi: 10.1139/W07-081. [DOI] [PubMed] [Google Scholar]
- Nadeem S.M., Zaheer Z.A., Naveed M., Nawaz S. Mitigation of salinity-induced negative impact on the growth and yield of wheat by plant growth-promoting rhizobacteria in naturally saline conditions. Ann. Microbiol. 2013;63(1):225–232. [Google Scholar]
- Naz I., Bano A., Tamoor-ul-Hassan. Isolation of phytohormones producing plant growth promoting rhizobacteria from weeds growing in Khewra salt range, Pakistan and their implication in providing salt tolerance to Glycine max L. Afr. J. Biotechnol. 2009;8(21):5762–5766. [Google Scholar]
- Netondo G.W., Onyango J.C., Beck E. Sorghum and salinity: II. Gas exchange and chlorophyll fluorescence of sorghum under salt stress. Crop Sci. 2004;44:806–811. [Google Scholar]
- Nia S.H., Zarea M.J., Rejali F., Varma A. Yield and yield components of wheat as affected by salinity and inoculation with Azospirillum strains from saline or non-saline soil. J. Saudi Soc. Agric. Sci. 2012;11:113–121. [Google Scholar]
- Palaniyandi S.A., Damodharan K., Yang S.H., Suh J.W. Streptomyces sp. strain PGPA39 alleviates salt stress and promotes growth of ‘Micro Tom’ tomato plants. J. Appl. Microbiol. 2014;117:766–773. doi: 10.1111/jam.12563. [DOI] [PubMed] [Google Scholar]
- Patel B.B., Patel Bharat.B., Dave R.S. Studies on infiltration of saline–alkali soils of several parts of Mehsana and Patan districts of north Gujarat. J. Appl. Technol. Environ. Sanitation. 2011;1(1):87–92. [Google Scholar]
- Paul D. Osmotic stress adaptations in rhizobacteria. J. Basic Microbiol. 2012;52:1–10. doi: 10.1002/jobm.201100288. [DOI] [PubMed] [Google Scholar]
- Ramadoss D., Lakkineni V.K., Bose P., Ali S., Annapurna K. Mitigation of salt stress in wheat seedlings by halotolerant bacteria isolated from saline habitats. Springer Plus. 2013;2(6):1–7. doi: 10.1186/2193-1801-2-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richards R.A. Should selection for yield in saline regions be made on saline or non-saline soils. Euphytica. 1983;32:431–438. [Google Scholar]
- Sadeghi A., Karimi E., Dahaji P.A., Javid M.G., Dalvand Y., Askari H. Plant growth promoting activity of an auxin and siderophore producing isolate of Streptomyces under saline soil conditions. World J. Microbiol. Biotechnol. 2012;28:1503–1509. doi: 10.1007/s11274-011-0952-7. [DOI] [PubMed] [Google Scholar]
- Sandhya V., Ali Sk.Z., Grover M., Reddy G., Venkateswarlu B. Alleviation of drought stress effects in sunflower seedlings by exopolysaccharides producing Pseudomonas putida strain P45. Biol. Fertility Soil. 2009;46:17–26. [Google Scholar]
- Saravanakumar D., Samiyappan R. ACC deaminase from Pseudomonas fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J. Appl. Microbiol. 2007;102:1283–1292. doi: 10.1111/j.1365-2672.2006.03179.x. [DOI] [PubMed] [Google Scholar]
- Schubert S., Neubert A., Schierholt A., Sumer A., Zorb C. Development of salt-resistant maize hybrids: the combination of physiological strategies using conventional breeding methods. Plant Sci. 2009;177:196–202. [Google Scholar]
- Seckin B., Sekmen A.H., Turkan I. An enhancing effect of exogenous mannitol on the antioxidant enzyme activities in roots of wheat under salt stress. J. Plant Growth Regul. 2009;28:12–20. [Google Scholar]
- Shahbaz M., Ashraf M. Improving salinity tolerance in cereals. Crit. Rev. Plant Sci. 2013;32:237–249. [Google Scholar]
- Shannon M.C., Grieve C.M. Tolerance of vegetable crops to salinity. Sci. Horticulturae. 1999;78:5–38. [Google Scholar]
- Shukla P.S., Agarwal P.K., Jha B. Improved Salinity tolerance of Arachis hypogaea (L.) by the interaction of halotolerant plant-growth-promoting rhizobacteria. J. Plant Growth Regul. 2012;31:195–206. [Google Scholar]
- Singh K.N., Chatrath R. Salinity tolerance. In: Reynolds M.P., Monasterio J.I.O., McNab A., editors. Application of Physiology in Wheat Breeding. CIMMYT; Mexico, DF: 2001. pp. 101–110. [Google Scholar]
- Stajner D., Kevresan S., Gasic O., Mimica-Dukic N., Zongli H. Nitrogen and Azotobacter chroococcum enhance oxidative stress tolerance in sugar beet. Biol. Plantarum. 1997;39(3):441–445. [Google Scholar]
- Tabur S., Demir K. Role of some growth regulators on cytogenetic activity of barley under salt stress. Plant Growth Regul. 2010;60:99–104. [Google Scholar]
- Tank N., Saraf M. Salinity-resistant plant growth promoting rhizobacteria ameliorates sodium chloride stress on tomato plants. J. Plant Interact. 2010;5:51–58. [Google Scholar]
- Tester M., Davenport R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003;91:503–507. doi: 10.1093/aob/mcg058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tiwari S., Singh P., Tiwari R., Meena K.K., Yandigeri M., Singh D.P., Arora D.K. Salt-tolerant rhizobacteria-mediated induced tolerance in wheat (Triticum aestivum) and chemical diversity in rhizosphere enhance plant growth. Biol. Fertility Soils. 2011;47:907–916. [Google Scholar]
- Upadhyay S.K., Singh J.S., Saxena A.K., Singh D.P. Impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions. Plant Biol. 2011;14:605–611. doi: 10.1111/j.1438-8677.2011.00533.x. [DOI] [PubMed] [Google Scholar]
- Venkateswarlu B., Shanker A.K. Climate change and agriculture: adaptation and mitigation strategies. Indian J. Agron. 2009;54:226–230. [Google Scholar]
- Wang W., Vinocur B., Altman A. Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta. 2003;218:1–14. doi: 10.1007/s00425-003-1105-5. [DOI] [PubMed] [Google Scholar]
- Wu Z., Yue H., Lu J., Li C. Characterization of rhizobacterial strain Rs-2 with ACC deaminase activity and its performance in promoting cotton growth under salinity stress. World J. Microbiol. Biotechnol. 2012;28:2383–2393. doi: 10.1007/s11274-012-1047-9. [DOI] [PubMed] [Google Scholar]
- Yamaguchi T., Blumwald E. Developing salt-tolerant crop plants: challenges and opportunities. Trends Plant Sci. 2005;10(12):615–620. doi: 10.1016/j.tplants.2005.10.002. [DOI] [PubMed] [Google Scholar]
- Yang J., Kloepper J.W., Ryu C.M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009;14:1–4. doi: 10.1016/j.tplants.2008.10.004. [DOI] [PubMed] [Google Scholar]
- Yao L., Wu Z., Zheng Y., Kaleem I., Li C. Growth promotion and protection against salt stress by Pseudomonas putida Rs-198 on cotton. Eur. J. Soil Biol. 2010;46:49–54. [Google Scholar]
- Yensen N.P. Halophyte uses for the twenty-first century. In: Khan M.A., Weber D.J., editors. Ecophysiology of High Salinity Tolerant Plants. Springer; Dordrecht: 2008. pp. 367–396. [Google Scholar]
- Yildirim E., Taylor A.G. Effect of biological treatments on growth of bean plans under salt stress. Ann. Rep. Bean Improvement Cooperative. 2005;48:176–177. [Google Scholar]
- Zahir Z.A., Munir A., Asghar H.N., Arshad M., Shaharoona B. Effectiveness of rhizobacteria containing ACC-deaminase for growth promotion of peas (Pisum sativum) under drought conditions. J. Microbiol. Biotechnol. 2008;18(5):958–963. [PubMed] [Google Scholar]
- Zhang H., Kim M.S., Sun Y., Dowd S.E., Shi H., Pare P.W. Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol. Plant Microbe Interact. 2008;21:731–744. doi: 10.1094/MPMI-21-6-0737. [DOI] [PubMed] [Google Scholar]
- Zhu J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Bol. 2002;53:247–273. doi: 10.1146/annurev.arplant.53.091401.143329. [DOI] [PMC free article] [PubMed] [Google Scholar]