

Abstract
We used multilevel modeling to examine dynamic changes in respiratory sinus arrhythmia (RSA) and observer-coded interactive synchrony for mother-child dyads engaged in a laboratory interaction, to characterize parenting-at-risk. Seventy-nine preschooler-mother dyads including a subset with documented child maltreatment (CM; n=43) were observed completing a joint puzzle task while physiological measures were recorded. Dyads led by CM mothers showed decreases in positive synchrony over time, whereas no variation was observed in non-CM dyads. Growth models of maternal RSA indicated that mothers who maintained high levels of positive interactive synchrony with their child evidenced greater RSA reactivity, characterized by an initial withdrawal followed by augmentation as the task progressed, after accounting for CM group status. These results help to clarify patterns of RSA responding in the context of caregiver-child interactions, and demonstrate the importance of modeling dynamic changes in physiology over time in order to better understanding biological correlates of parenting-at-risk.
Keywords: respiratory sinus arrhythmia, RSA, parenting, child maltreatment
Research on parenting-at-risk has documented adverse effects of harsh parenting on the development of a child's ability to self-regulate their autonomic physiology and behavior (Calkins, et al., 2008; Rothbaum & Weisz, 1994). At the extreme end of this continuum, child maltreatment (CM) has been linked to an increased risk for emotional and behavioral dysregulation (Cicchetti & Toth, 2005), disruption of stress neurophysiology (Pollak, 2008), and adverse mental health outcomes (Kaplow & Widom, 2007). Likewise, impairments in self-regulation constitute a significant contributing factor for CM parenting and have been reported in perpetrators of CM and those at risk for CM (Deater-Deckard, Sewell, Petrill, & Thompson, 2010; Stern & Azar, 1998). Parents at risk for CM display greater reactivity to child-specific and neutral stimuli, difficulties regulating their emotions and behavior, and higher resting heart rate and greater sympathetic activation than do non-CM parents (Joosen, Mesman, Bakermans-Kranenburg, & van IJzendoorn, 2013; McCanne & Hagstrom, 1996). Thus, understanding the nature of regulatory deficits associated with parenting at-risk could be greatly informative to interventions aimed at preventing the hostile, controlling, and intrusive interaction styles characteristic of CM parenting (e.g., Wilson, Rack, Shi, & Norris, 2008). Physiological measures of self-regulation are particularly well-suited for investigating this question because they reflect real-time reactivity to context and can forecast moment-to-moment shifts in the quality of parenting observed during the act of parenting (e.g., Skowron et al., 2013). This study was designed to characterize the quality of maternal-child dyadic interactions and the trajectories of respiratory sinus arrhythmia (RSA) for mothers and children during a joint puzzle task, with a specific focus on the potential moderating role of CM status.
Research using observations of structured interactions in laboratory and household settings to investigate CM family interactions has revealed rich insights that help to inform critical targets of intervention. For example, interactions between CM parents and their children are less positive and devolve into negative exchanges that escalate over time (e.g., Reid et al., 1981; Knutson, DeGarmo, Koeppl, & Reid, 2005). CM parents display more consistent, contingent negative responses to their children's negative behaviors and less contingent positive responding to their children's prosocial behavior (e.g., Dumas & Wahler, 1985; Wahler, Williams, & Cerezo, 1990), and are more likely to respond to their preschoolers' efforts to assert age-appropriate autonomy with strict, harsh control (Skowron et al., 2011). Although meta-analysis indicates that interventions for CM lead to self-reported improvements in parent and child functioning, most interventions fail to result in improvements in the quality of observed parenting (e.g., Skowron & Reinemann, 2005; but see Chaffin et al., 2004; 2011, for important exceptions).
Positive interactive synchrony in mother-child dyads is defined as the temporal coordination of micro-level positive social behavior (Feldman, 2007) and is characterized by reciprocity and mutually contingent responsiveness (Harrist & Waugh, 2002). In infancy, the experience of positive synchrony with one's caregiver is thought to serve relational, homeostatic regulatory functions (Tronick, 1989), whereas in the preschool years, the experience of positive interactive synchrony supports and enables children's developing autonomy, self-control, and social skills (Harrist & Waugh, 2002). Research in normative samples shows that healthy mother-infant and mother-toddler interactions are characterized by high rates of positive interactive synchrony, interspersed with brief ruptures that are successfully repaired, returning the dyad back into positive synchrony (e.g., Cohn & Tronick, 1987, 1988; Harrist et al., 1994; Rocissano, Slade, & Lynch, 1987). In contrast, mothers at high risk for CM show greater rates of interactive miscoordination, characterized by mother-initiated ruptures, and fewer successful repairs, relative to low-risk families (Skowron, Kozlowski, & Pincus, 2010). Critically, the degree of positive interactive synchrony children experienced with a caregiver is linked with individual differences in children's physiological and behavioral regulation abilities (Gianino & Tronick, 1988; Porter, 2003), but no published studies to date have begun to clarify the role of maternal physiology in achieving positive dyadic synchrony with their child.
The parasympathetic nervous system (PNS) has been proposed as a biological system underlying many of these crucial facets of behavior, particularly emotional and physiological reactivity and regulation in the context of social engagement (e.g., Porges, 1995; 2001; Thayer & Lane, 2000). The PNS has been implicated in the down-regulation of autonomic arousal via the influence of the vagus nerve on heart rate variability, often indexed by rhythmic fluctuations in heart periods at the respiratory frequency, known as respiratory sinus arrhythmia (RSA; Berntson, Cacioppo, & Quigley, 1993). According to Porges' polyvagal theory (Porges, 1995; 2001), PNS control is mediated by neural mechanisms that allow rapid responses to changing demands in the environment, in particular the ability to dynamically regulate physiological arousal (e.g., cardiac output) to foster engaging or disengaging with situational demands. Phasic changes in RSA are often couched in terms of RSA augmentation and withdrawal from baseline levels, due to their association with increases and decreases in activation of the parasympathetic nervous system, respectively (e.g., Beauchaine, 2001). Decreases in RSA index the withdrawal of PNS influence on the heart, leading to cardiac acceleration to support the metabolic requirements involved in mobilization and attentional engagement, while increases in RSA index an augmentation of PNS influence on the heart, resulting in a slowed heart rate that promotes calm behavioral states, self-soothing, and social engagement (e.g., Porges, 2001; Berntson et al., 1993). Indeed, a number of findings have documented an association between augmented RSA and elevated levels of social engagement and positive emotionality (Geisler, Kubiak, Siewert, & Weber, 2013; Kok & Frederickson, 2010; Holt-Lunstad et al., 2007). Neuroimaging studies further support the role of this system in social behavior, as medial prefrontal regions implicated in fluctuations in PNS activity overlap with regions commonly observed in social neuroscience research (see Thayer et al., 2012).
Much of the existing research on RSA has characterized the role of baseline, or resting values, and task difference scores (e.g., Beauchaine, 2001; Berntson, Cacioppo, & Quigley, 1993; Porges, 1995; Thayer & Lane, 2009). In children and adults, higher levels of baseline RSA have been associated with greater regulatory capacity and attentional control (Beauchaine, 2001; Fox, 1989; Mezzacappa, Kindlon, Saul, & Earls, 1998), whereas lower values of resting RSA have been linked to increased risk for emotion dysregulation and a broad range of psychopathologies (Beauchaine, 2001; Graziano & Derefinko, 2013; Light, Kothandapini, & Allen, 1998; Thayer et al., 1996). When examining RSA during a given task relative to baseline levels, increases in RSA (i.e., RSA augmentation) typically relate to increased efforts to regulate one's emotions (Butler, Wilhelm, & Gross, 2006; Ingjaldsson et al, 2003; Skowron et al., 2011). Conversely, decreases in RSA (i.e., RSA withdrawal) have been associated with psychological stress (Berntston, Cacioppo, Quigley, & Fabro, 1994) and negative emotionality (Beauchaine, 2001).
In the context of parenting, most research on RSA has focused on the impact on child RSA, with few studies examining the role of RSA in parents. For children interacting with their mothers, increases in child RSA have been consistently linked to greater social engagement and maternal care (Hastings et al., 2008; Moore et al., 2009; Moore & Calkins, 2004; Feldman, Singer, & Zagoory, 2010). RSA findings for parents are less clear, with conflicting results about whether optimal parenting is associated with RSA increases (e.g., Skowron et al., 2011; Weisman, Zagoory-Sharon, Feldman, 2012; Hill-Soderlund et al., 2008) or decreases (e.g., Mills-Koonce et al., 2009; Moore et al., 2009). Higher baseline RSA in mothers has been associated with greater sensitivity to their infant during free play (Musser, Ablow, & Measelle, 2012), and a recent study showed that increases in parent-child engagement induced by oxytocin administration were associated with increased RSA during free play in parents and their children (Weisman, Zagoory-Sharon, & Feldman, 2012). In contrast, others have reported that mothers rated high in maternal sensitivity show larger RSA decreases when reuniting with their infant during the still-face paradigm than less sensitive mothers (Moore et al., 2009). These conflicting results may be due to dynamic coupling between physiology and behavior that fluctuates over the course of an interaction and varies in directionality based on context and/or individual differences among the parents in question (Skowron et al., 2013). Given that the present experimental context of a mother and child seated together with a puzzle toy is more similar to a free play period than a stress-induction task such as the still-face paradigm, it follows that mothers in this study would likely show RSA increases in association with more positively-rated interactions with their child (e.g., Musser et al., 2012).
The primary goal of this study was to clarify whether increases or decreases in maternal RSA observed during interactions with one's child would be associated with positive or maladaptive parenting, by quantifying trajectories of changes in RSA during the course of a recording session. Such an approach moves beyond examining a single value of RSA change from baseline values and can be used to provide a finer-grained characterization of the dynamic nature of physiological changes during an interpersonal interaction (Fortunato, Gatzke-Kopp, & Ram, 2013; Oppenheimer, Measelle, Laurent, & Ablow, 2013; Miller, Chocol, Nuselovici, Utendale, Simard, & Hastings, 2011; Brooker & Buss, 2010). For example, Fortunato and colleagues (2013) found that RSA responses dynamically varied over the course of emotion-inducing film clips, and that these variations reflected individual differences not captured by using static change scores alone (Fortunato et al., 2013). Brooker and Buss (2010) examined RSA reactivity in high-fear toddlers during a fear-eliciting task, and found that static RSA change scores predicted behaviorally-coded boldness only among low-fear control children; however, dynamic, non-linear changes in RSA predicted individual differences in shyness behavior among high-fear toddlers. Given similar findings of nonlinear patterns of RSA reactivity in children (Miller et al., 2013) and in mothers interacting with their children (Oppenheimer et al. 2013), a more fine-grained temporal characterization of RSA dynamics represents a compelling next step in understanding the role of physiology in mother-child dyadic synchrony. This may be particularly true in at-risk populations who show disruptions in the functioning of self-regulatory systems during interactions (e.g., Skowron et al., 2013). We adopted this approach to examine growth models of RSA reactivity in children and their mothers during a social interaction.
The current study combines this microanalytic acquisition approach to quantifying the quality of mother-child interactions and a growth modeling analytic approach to quantify RSA change over time in mothers and their children as it relates to dyadic interactive synchrony. Specifically, moment-to-moment interactions were microcoded at the speaking turn level and quantified within 30-second blocks of time over the course of a 5-minute joint puzzle task. During this time, RSA was recorded simultaneously in mothers and their children, and also quantified within 30-second blocks over the course of the task. Multilevel modeling was used to examine growth trajectories of positive interactive synchrony and RSA throughout the task. Based on previous findings (Hastings et al., 2008; Skowron et al., 2010), we investigated whether (1) mother-child dyads with a history of CM would demonstrate lower levels of positive interactive synchrony than non-CM dyads, and whether (2) levels of positive dyadic synchrony would relate to distinct growth patterns in RSA for mothers and children. Regarding the direction of the relationship between positive synchrony and RSA, several outcomes could be expected based on the mixed results in the existing literature. For example, greater interactive synchrony might be associated with increases in parasympathetic tone, reflecting efforts to regulate emotion during a positive social interaction (e.g., Butler et al., 2006; Hastings et al., 2008); conversely, greater interactive synchrony could be associated with decreases in parasympathetic tone, reflecting greater attentional focusing and engagement during the joint task (Beauchaine, 2001). We predicted that among the lower risk (i.e., non-CM) dyads in the sample, the extent of positive interactive synchrony observed may be unrelated to mothers' parasympathetic tone, following the notion that healthy parent-child dyadic states may be achieved independent of mother's physiological arousal in parents at low risk for child maltreatment (Skowron et al., 2013).
Method
Participants
Mother-child dyads were recruited, where the mother was 18 years of age or older, spoke fluent English, and lived with her preschool child. Initial recruitment efforts targeted families involved with Child Protective Services (CPS), specifically mothers who were documented perpetrators of CM. For comparison with the CM group, a sociodemographically similar sample was recruited from department of public welfare agencies and a database featuring information on birth announcements in local newspapers. Physiological recordings from both mother and her child during the lab visit were available from a total of 155 dyads. Given the novelty of the present statistical modeling approach, we excluded dyads that had missing RSA data for any single 30-second epoch during the joint puzzle task. This conservative approach was taken to reduce the likelihood of spurious results related to patterns of missing data, as ECG data is often unable to be processed during a given epoch due to movement-related artifact by the participant, also reducing the potential influence of motor activity on changes in RSA (e.g., Bush et al., 2011). As such, continuous RSA data throughout the task was available for a total of 79 mother-child dyads, split into groups of CM (n=43) and non-CM (n=36) families. Importantly, dyads included in final analyses did not differ from the dyads excluded for missing physiological data on any of the key variables of interest, specifically child gender, child age, maternal age, maternal years of education, CM status, child baseline and task RSA, maternal baseline and task RSA (all ps > .05). Comorbidity of CM subtypes was observed in 65.1% of this sample, consistent with previous reports (Belsky, 1993), and thus were collapsed across subtypes into a single CM group. The two groups, CM and non-CM dyads, were matched on child age (control, M = 3.75, SD = .73, range = 3 to 5 years; maltreatment, M = 3.88, SD = .79, range = 3 to 5 years), t(77) = -.77, p = .44, maternal age (control, M = 31.22, SD = 5.11, range = 23 to 42 years; maltreatment, M = 29.49, SD = 6.67, range = 20 to 4 5 years), t(77) = 1.28, p = .21, and child gender, t(77) = -.55, p = .59. Despite efforts to recruit a socioeconomically-comparable sample to the CM group, there were group differences in maternal education, t(77) = 4.261, p < .001. Non-CM mothers reported more years of schooling (M = 14.92, SD = 2.92) than CM mothers (M = 12.53, SD = 2.03). For statistical analyses, maternal years of education was used as a proxy variable for socioeconomic status.
Procedure
All materials and components of the procedure were approved by the Office for Research Protections. Families were recruited as part of a larger project investigating parenting behaviors and the development of children's self-regulation in families with and without a history of CM (e.g. Skowron et al., 2011). Mother-child dyads completed a three-visit protocol over a 2-3 week period, consisting of two at-home assessments and a 2.5 hour laboratory session. Families were paid $150 to complete the three visits, and in addition were provided transportation, snacks, and children's toys. The results presented herein only concern the laboratory session.
Upon arrival to the laboratory, both mother and child were fitted with electrodes for psychophysiological data collection. Then dyads completed a 5-minute resting baseline period, while seated together on a comfortable couch and viewing a relaxing Baby Einstein video segment (http://www.babyeinstein.com). After the baseline period, mother and child were then seated together at a small table, and the child was provided with an assembled duplo-block object and 12 disassembled blocks to construct a replica. Mothers were instructed to verbally aid their child in building the object as they might typically, but were not allowed to physically touch any of the pieces. The task was stopped after 5 minutes of activity.
Observational coding of dyadic synchrony
A key focus of this study was to examine variation in the level of positive interactive synchrony within mother and child dyads over the course of the joint puzzle task1. To operationalize positive synchrony, observational data from the mother-child joint task were coded using the Structural Analysis of Social Behavior (SASB; Benjamin, 1996, see Figure 1). SASB coding provides a framework for characterizing the extent of positive dyadic interactive synchrony observed across sequentially ordered individual mother and child “speaking” turns. Mothers and children's videotaped interactions during the joint task were transcribed, unitized by speaking turn to delineate each coding unit, and subjected to SASB coding by a team of two trained coders blind to the family's maltreatment status. SASB coding began with the first codable event (i.e., first utterance by mother or child) in the sequence of mother-child interaction, and ended with the last codable behavior that occurred. Coders received 60 hours of training using practice tapes to achieve sufficient reliability (weighted kappa > .7) prior to coding. Interrater reliability was calculated using Cohen's weighted kappa, a conservative method of computing agreement between raters on categorical codes that controls for the likelihood of chance agreement and penalizes judges for the magnitude of disagreement (Cohen, 1968).
Figure 1.
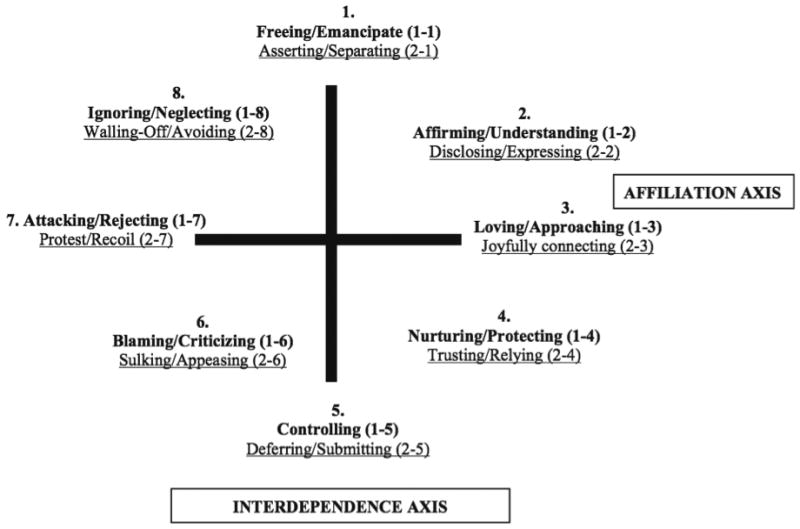
SASB simplified cluster model. Labels in bold print describe actions directed toward another person. Labels in underline print describe actions in response to the other. Adapted from Interpersonal Diagnosis and Treatment of Personality Disorders (2nd ed.) by L.S. Benjamin, 1996, New York, NY: Guilford Press. Copyright © 1996 by Guilford Press. Reprinted with permission.
The process of SASB coding a unit of behavior involves three steps: determining focus, degree to warmth/affiliation, and degree of interdependence (Humphrey & Benjamin, 1989). As shown in Figure 1, SASB Clusters 2, 3, and 4, form a constellation of “positive” behaviors: (2: Affirm/Understand, Disclose/Express), (3: Love/Approach, Joyfully Connect), (4: Nurture/Protect, Trust/Rely). Conversely, relationships characterized by transactional patterns containing behaviors in Clusters 6, 7, and 8 (6: Blame/Criticize, i.e., harsh control, and Sulk/Appease, i.e., hostile submission), (7: Attack/Reject, Protest/Recoil), (8: Ignore/Neglect, and Wall-Off/Avoid behaviors), form those categorized as “negative” behaviors. Each speaking turn by the mother and child was assigned a SASB cluster code, following which, codes from proximal speaking turns were reviewed to characterize sequences of the higher-order dyadic state variable, positive interactive synchrony, defined as a minimum 3-step sequence of positive mother-child behaviors (i.e., M+, C+, M+; i.e., 3 mother-child transactions SASB-coded in Clusters 2, 3, or 4). The proportion of dyadic interaction spent in positive synchrony was calculated in terms of (a) epoch-level positive synchrony for each 30-second epoch, yielding 10 sequentially-ordered scores across the joint task (level 1 variable), and (b) task-average positive synchrony scores, or the average across all 30-second epochs (entered as a level 2 variable in the multilevel growth models). As reported previously with this dataset (Skowron et al., 2013), interrater reliability was calculated on 15% of the full sample, with weighted kappas ranging from .52 to 1.0 (M = .74), where values at .41 to .60 are considered “moderate”, values between .61 to .80 are “substantial”, and .81 to 1.00 are “almost perfect” (Landis & Koch, 1977).
Physiological data acquisition
Disposable Ag/AgCl electrodes were placed on mothers' and children's chests in a modified Lead II placement, on the distal end of the right clavicle, lower left rib cage, and lower abdomen. Electrocardiogram (ECG) data were acquired via Mindware Technologies (Gahanna, OH) ambulatory electrocardiograph MW1000A at a sampling rate of 500 Hz, and transmitted via a wireless signal to a computer monitored by a research assistant. The ECG signal was synchronized at acquisition with video recordings of behavior to enable time-synchronization of behavioral codes and RSA values. ECG data were then processed offline using Mindware Technologies HRV 2.6 analysis program. Trained research assistants visually inspected 30-second epochs for erroneously identified or missing heartbeats, and manually deleted or inserted as appropriate. The resulting interbeat interval time series was then subjected to a fast-Fourier transformation, and power in the respiratory frequency band was derived from the spectral density function to estimate RSA values. The RSA frequency band was set from 0.24-1.04 for children, and 0.12-0.40 for mothers. It should be noted that respiration data were only obtained from a small subset of participants from whom cardiac impedance data was collected (N = 13 dyads), thus RSA values in this manuscript are only estimates of RSA based on activity in the respective frequency bands for children and adults.
For baseline data, RSA values were averaged across 30-second epochs of the 5-minute task to create a single score. For RSA data collected during the joint puzzle task, multilevel modeling was used to analyze the values obtained from each 30-second epoch over the course of the task relative to baseline RSA. For each mother and child's RSA data, within-task RSAchange scores were calculated for each 30-second epoch of the 5-minute joint task as the difference in the RSA score during an epoch of the joint puzzle task, relative to the person's average RSAbaseline score (i.e., RSA change = RSA baseline – RSA task epoch). Although more dyads participated in the full study, only 79 dyads had clean, available RSA data for mother and child in both the baseline period and all epochs of the joint-task.
Analytic Strategy
Multilevel modeling was employed to examine the moderating impact of CM status on growth models of positive dyadic synchrony and RSA during the joint puzzle task. Models were run separately for positive dyadic synchrony, mother RSA, and child RSA, and focused on identifying the extent of linear and/or quadratic change in the outcome of interest as a function of CM history, while controlling for person-level averages of positive synchrony and SES2. Multilevel models were specified within Hierarchical Linear Modeling (HLM) software (Raudenbush & Bryk, 2002), with growth parameters entered at Level-1 (within person or dyad) and CM group status and other covariates at Level-2 (between-person or dyads). Linear and quadratic coefficients for time were entered uncentered as the epoch number (1-10) and the square of each epoch number, respectively.
| Level 1: | Synchronyij = β0j + β1j*(Timeij) + β2j*(Time2ij) + rij |
| Level 2: | β0j = γ00 + γ01*(Groupj) + γ02*(Avg. Synchronyj) + γ03*(SESj) + u0j |
| β1j = γ10 + γ11*(Groupj) + γ12*(Avg. Synchronyj) + γ13*(SESj) + u1j | |
| β2j = γ20 + γ21*(Groupj) + γ22*(Avg. Synchronyj) + γ23*(SESj) + u2j |
Similar models were run with mother and child RSA as DVs to examine growth trajectories of RSA and the moderating impact of variables of interest. In these models, baseline RSA and overall task RSA were entered as level-2 predictors, along with CM group status and covariates3. The outcome variable in these models was RSA reactivity measured via within-task RSAchange scores.
| Level 1: | RSAchangeij = β0j + β1j*(Timeij) + β2j*(Time2ij) + rij |
| Level 2: | β0j, β1j β2j = γ00 + γ01*(Groupj) + γ02*(Avg.Synchronyj) + γ03*(RSA baselinej) + γ04*(RSA taskj) + u0j |
Results
Growth model of dyadic synchrony by CM status
There was a significant main effect of group status on positive dyadic synchrony (γ01 = .22, t(75) = 2.12, p < .001), controlling for all other effects listed in the model (Table 2). This main effect was qualified by significant group × time and group × time2 interactions; CM dyads had a greater linear decrease in positive synchrony over the course of the task (γ11 = -.11, t(75) = -2.22, p = .030) and a more pronounced quadratic change in positive synchrony (γ21 = .01, t(75) = 2.04, p = .045). As seen in Figure 2, positive synchrony for CM dyads sharply decreased at the onset of the task, followed by a return to task-average levels. In contrast, non-CM positive synchrony showed a relatively flatter trajectory of change during the task. It should be noted that overall task levels showed no CM group differences in the proportion of mother-child interactions spent in positive synchrony averaged across the task (non-CM = .58, CM = .52; t(77) = .92, p = .36), yet CM group differences emerge in the dynamic levels of positive synchrony observed over the course of the task.
Table 2. Multilevel Model of Linear and Quadratic Growth in Positive Dyadic Synchrony.
| Coefficient | SE | |
|---|---|---|
| Intercept, π0 | ||
| Intercept, β00 | -0.410 | 0.300 |
| Group, β01 | 0.224 | 0.106* |
| Synchrony, β02 | 0.870 | 0.169*** |
| SES, β03 | 0.035 | 0.020 |
| Linear Growth, π1 | ||
| Linear slope, β10 | 0.224 | 0.145 |
| Linear × Group, β11 | -0.113 | 0.051* |
| Linear × Synchrony, β12 | 0.029 | 0.081 |
| Linear × SES, β13 | -0.019 | 0.010 |
| Quadratic Growth, π2 | ||
| Quadratic slope, β20 | -0.018 | 0.015 |
| Quadratic × Group, β21 | 0.010 | 0.005* |
| Quadratic × Synchrony, β22 | -0.004 | 0.008 |
| Quadratic × SES, β23 | 0.002 | 0.001 |
p < .05;
p < .001
Figure 2.
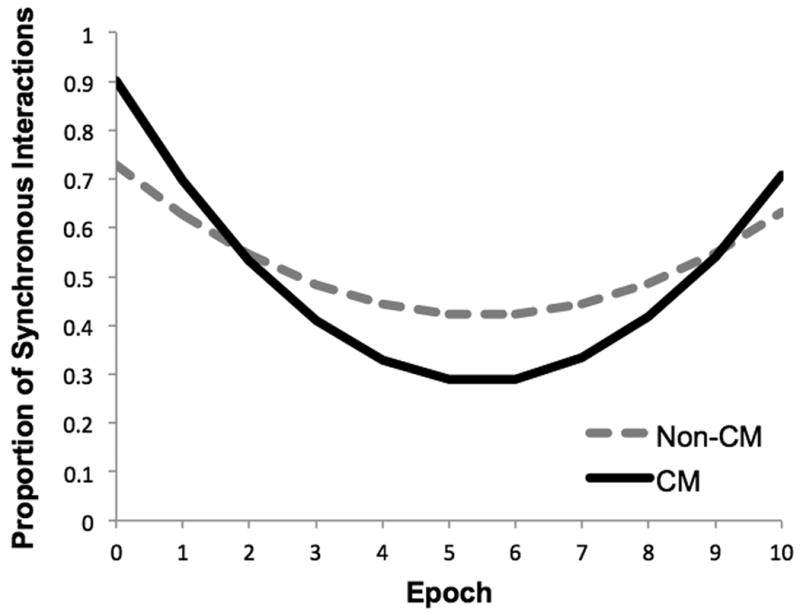
The Moderating Impact of Child Maltreatment on Growth Models of Dyadic Synchrony During the Joint Puzzle Task.
Growth models of RSA reactivity in mothers and children
Given that growth models of positive dyadic synchrony indicated that there is meaningful variance across risk groups in the quality of mother-child interactions throughout a joint puzzle task, we ran similar growth models for fluctuations in mother's and child's within-task RSAchange scores (i.e., relative to baseline RSA) as a function of CM status and overall positive synchrony during the task (Table 3). Additionally, these models controlled for the given individual's baseline RSA and average task RSA at level-2.
Table 3. Multilevel Model of Linear and Quadratic Growth in RSA for Mothers and Children.
| Mothers | Children | |||
|---|---|---|---|---|
| Coefficient | SE | Coefficient | SE | |
| Intercept, π0 | ||||
| Intercept, β00 | 0.972 | 0.742 | -0.637 | 0.698 |
| Group, β01 | -0.050 | 0.234 | 0.142 | 0.218 |
| Average Synchrony, β02 | -1.101 | 0.397* | 0.245 | 0.379 |
| Task RSA Average, β03 | 1.064 | 0.164*** | 1.129 | 0.141*** |
| Baseline RSA Average, β03 | -1.092 | 0.157*** | -1.031 | 0.126*** |
| Linear Growth, π1 | ||||
| Linear slope, β10 | -0.448 | 0.324 | 0.234 | 0.315 |
| Linear × Group, β11 | -0.001 | 0.068 | -0.048 | 0.057 |
| Linear × Synchrony, β12 | 0.458 | 0.175* | -0.159 | 0.098 |
| Linear × Task RSA, β13 | 0.014 | 0.072 | -0.009 | 0.172 |
| Linear × Baseline RSA, β13 | 0.013 | 0.068 | -0.020 | 0.065 |
| Quadratic Growth, π2 | ||||
| Quadratic slope, β20 | -0.018 | 0.015 | -0.016 | 0.029 |
| Quadratic × Group, β21 | 0.010 | 0.005 | 0.003 | 0.009 |
| Quadratic × Synchrony, β22 | -0.036 | 0.016* | 0.017 | 0.016 |
| Quadratic × Task RSA, β23 | -0.004 | 0.007 | -0.002 | 0.006 |
| Quadratic × Baseline RSA, β23 | 0.001 | 0.006 | 0.004 | 0.005 |
p <.05;
p <.01;
p <.001
For growth models of mother's within-task (i.e., dynamic) RSAchange, there was a main effect of overall task RSA (γ04 = 1.06, t(74) = 6.51, p < .001) and a main effect of baseline RSA (γ01 = -1.09, t(74) = -6.97, p < .001), such that higher baseline values were associated with lower mother RSAchange values. There were no effects of CM group status and mothers' dynamic RSAchange scores (all ps > .83). A main effect of task-average positive synchrony was observed (γ03 = -1.10, t(74) = -2. 77, p = .007), and this effect was qualified by significant interactions between synchrony × time (γ13 = .46, t(74) = 2.61, p = .011) and synchrony × time2 (γ23 = -.04, t(74) = -2.26, p = .027). Interpretation of the linear effects indicates that mothers in dyads posting high levels of positive dyadic synchrony showed the lowest initial RSAchange values (i.e., greater RSA withdrawal) at the task outset, followed by a trajectory of linear increase over time in mother RSAchange values (i.e., to RSA augmentation). Interpretation of the quadratic effects, shown in Figure 3, indicate that mothers from dyads high in positive dyadic synchrony showed the largest RSA withdrawal to task onset, followed by an abrupt RSA augmentation before returning to baseline levels at the task's end. In contrast, the RSA growth trajectories of mothers in dyads posting low levels of positive synchrony showed the least reactivity over the course of the task.
Figure 3.

Growth Models of Mother's Respiratory Sinus Arrhythmia (RSA) Reactivity as a Function of Task-Average Dyadic Synchrony.
Growth models of children's RSA reactivity during the joint task did not show predictable linear or quadratic changes over time (ps > .45). After accounting for overall task RSA (γ04 = 1.06, t(74) = 6.51, p < .001) and baseline RSA (γ01 = -1.09, t(74) = -6.97, p < .001), there were no effects of task-average positive synchrony (all ps > .28) or CM group status (all ps > .51) on children's within-task RSAchange scores.
Discussion
The present study aimed to clarify relations between positive dyadic interactions and dynamic RSA measures in mothers and their preschool children. We examined profiles of variation in positive dyadic synchrony and mother and child RSA over the course of a laboratory task designed to give caregivers an opportunity to assist their child in completing a challenging objective. We were primarily interested in how the proportion of positive synchrony displayed by a dyad would relate to dynamic changes in RSA for mothers and children over time during the joint task.
Growth models of RSA reported here indicate that mothers who displayed greater positive interactive coordination with their preschool child at the micro-level of social behavior also showed greater autonomic reactivity over the course of their dyadic interaction. These findings are consistent with biological models that posit high levels of variability in physiological systems like the PNS reflect healthy processes, whereas low variation tends to characterize more pathological states (Friedman, 2007; Thayer & Lane, 2000), and suggests that mothers who achieved high levels of positive synchrony with their child during the joint task may have withdrawn RSA at the task onset (relative to their task average) to orient and focus on the task, and then augmented RSA levels through the remainder of the joint task to support positive social engagement and joint problem-solving with their child (e.g., Porges, 2001). In contrast, mothers who achieved low positive synchrony with their child displayed blunted RSA reactivity over the course of the task. The relationships reported here between positive dyadic synchrony and dynamic changes in mother's RSA in the act of parenting apply across the spectrum of risk in the sample.
Previous findings on RSA reactivity have been mixed with regard to what constitutes a healthy response in the context of parenting. Some have suggested that RSA withdrawal is optimal, as a decrease in parasympathetic tone facilitates elevated heart rate and greater attentional focus and engagement with stimuli (Beauchaine, 2001), and RSA decreases have been observed in mothers who are rated as more sensitive (Moore et al., 2009) and less intrusive (Mills-Koonce et al., 2009) with their distressed infants. In contrast, others have documented that RSA augmentation (from baseline) is concordant with efforts to regulate emotional expressiveness during social interactions (Butler et al., 2006; Smith et al., 2011). Our findings highlight the utility of examining dynamic changes in RSA in the context of parenting, and appear consistent with both sets of findings. Specifically, mothers who achieved greater positive synchrony with their preschooler showed larger RSA withdrawals at the onset of the task, presumably reflecting greater engagement and focus on the task objectives, followed by greater increases in RSA over the remainder of the task, suggesting a higher degree of social-emotional regulation while engaged with their child to complete the puzzle together. These results are also consistent with Graziano and Derefinko's (2013) recent meta-analysis concluding that decreases in vagal tone are more optimal during stressful interactions that call for greater effort to engage within a difficult context, while increased vagal tone may be more optimal in facilitating calm, social connections that call for regulation of emotional expression (e.g., Porges, 2001).
In another finding, observational coding revealed larger decreases in interactive synchrony among CM dyads over the course of the puzzle task than in non-CM dyads; however, CM status did not moderate the relationship between dyadic synchrony and RSA growth trajectories. One interpretation of this finding could be that the quality of mother-child dyadic coordination and difficulties associated with CM status are not uniquely related to autonomic physiology, but are due to other sources of variance, such as individual differences in child characteristics and maternal attributions about child intentions, comorbid psychopathology, life stressors, and parent representations of their own early experiences with caregivers (e.g., Bugental, 2009). It should be noted that CM and non-CM mother-child dyads were recruited from demographically comparable populations of relatively low SES and elevated risk status, and future studies should examine the patterns of dyadic synchrony in families with greater economic and social resources, to ascertain whether associations between physiology and interactive behavior observed here would generalize.
It is important to note that the RSA values reported here were not controlled for respiration values. Changes in respiration may confound the interpretation of RSA values as an indicator of vagal tone, instead reflecting movement-related activity which can heavily impact both respiration and RSA (for a review, see Grossman & Taylor, 2007). However, others have concluded that the confound of respiration only applies for relatively extreme values such as during exercise (Houtveen, Rietveld, & de Geus, 2002), and that respiration frequency has little impact on RSA amplitude in seated participants (Denver, Reed, & Porges, 2007). Given that the dyads in the present study were seated at all times in a relatively quiet laboratory environment, it does not seem likely that the RSA fluctuations reported here were primarily driven by changes in respiratory parameters, although this is a caveat that should be addressed in future studies. Another notable limitation of the current study is that claims about causality cannot be drawn about the relationship between the extent of dyadic synchrony and variations over time in RSA during the joint puzzle task. Additionally, we did not collect information on factors that could influence RSA, such as smoking status, caffeine or alcohol use, or BMI; thus, effects of group membership reported here between CM and non-CM mothers could reflect more general health differences. Nonetheless, our findings implicate blunted RSA reactivity in mothers as a risk factor for achieving low positive synchrony with their child. Given evidence that self-report from mothers and children may not be a reliable indicator of the quality of mother-child relationships (Skowron & Reinemann, 2005), identifying basic biometric measures of potential risk for maltreatment is an important step for the development of interventions aiming to identify malleable factors that can support greater positive coordination between at-risk parents and children in the early years. Ongoing studies in our lab are examining the role of more proximal effects between dyadic coordination, rupture, and repair processes, and patterns of RSA responding that may prove helpful in elucidating the neurobehavioral mechanisms underlying parenting in early childhood in order to optimize and tailor family interventions for at-risk populations. One potential direct implication of this work would be for interventions to analyze RSA in parents while interacting with their children and determine whether coaching methods designed to increase parasympathetic reactivity would improve the proportion of positive exchanges that parent would have with their child. Additionally, identifying parents who show a blunted parasympathetic response while interacting with their children could better inform interventionists about individuals who might need more training with emotion recognition and regulation strategies in order to support positive parenting practices.
Table 1. Descriptive Statistics for Mother-Child Dyads by Maltreatment Group Status.
| Maltreatment (N=43) | Control (N=36) | |
|---|---|---|
| Participant Characteristics | ||
| Proportion of Female Children | .53 (.50) | .47 (.51) |
| Child Age | 3.88 (.79) | 3.75 (.73) |
| Mother Age | 29.49 (6.67) | 31.22 (5.11) |
| Maternal Education (Years) | 12.53 (2.03)* | 14.92 (2.92)* |
| Joint-Task Outcomes | ||
| Proportion Who Completed Task | .35 (.48) | .33 (.48) |
| Number of Epochs (Task Length) | 8.56 (1.83) | 8.78 (1.59) |
| Proportion of Positive Synchrony | .52 (.28) | .58 (.31) |
| Baseline Child RSA | 6.43 (1.24) | 6.35 (1.34) |
| Baseline Mom RSA | 5.57 (1.48)† | 6.18 (1.27)† |
| On-task Child RSA | 5.68 (1.09) | 5.66 (1.20) |
| On-task Mom RSA | 5.47 (1.36) | 5.82 (1.29) |
Note: Values shown are means (and standard deviations).
Significant group difference at p < .01, two-tailed;
marginal trend at p < .07.
Highlights.
Changes in RSA and positive synchrony were modeled over time in mother-child dyads.
History of child maltreatment associated with lower positive synchrony.
Growth models of maternal RSA related to extent of positive synchrony.
Greater positive synchrony associated with larger RSA responsivity in mothers.
Footnotes
During the joint puzzle task, dyads spent a majority of their time either in positive synchrony (M = 54%) or in a state of dyadic mismatch (M = 42%). A small minority of the observed interactions classified as negative synchrony (M = 4%), with no difference between groups (p > .8). While positive and negative synchrony are clearly defined as periods of three or more consecutive speaking turns of positive or negative exchanges, dyadic mismatch encompasses a heterogeneous group of parent-child exchanges (for more detail on the coding of behaviors underlying dyadic states, see Skowron et al., 2011).
Maternal education (number of years) was used as a proxy variable for socioeconomic status. Additional exploratory models were run to test for the effects of other covariates, such as child age and whether the duplo-block task was successfully completed. No interactions with group status or dyadic synchrony were observed, thus these covariates were left out of presented models in the interest of parsimony. Overall, correlations between average positive dyadic synchrony, baseline RSA, and task RSA were non-significant for both mother and child RSA.
Child age, SES, and task completion were entered as covariates in exploratory models but demonstrated no significant effects. For parsimony, we only report reduced models here.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 73.Barnett D, Manly JT, Cicchetti D. Defining child maltreatment: The interface between policy and research. In: Cicchetti D, Toth SL, editors. Child abuse, child development, and social policy. Norwood, NJ: Ablex; 1993. p. 7. [Google Scholar]
- Belsky J. Etiology of child maltreatment: A developmental€ cological analysis. Psychological bulletin. 1993;114(3):413. doi: 10.1037/0033-2909.114.3.413. [DOI] [PubMed] [Google Scholar]
- Berntson GG, Cacioppo JT, Quigley KS. Respiratory sinus arrhythmia: autonomic origins, physiological mechanisms, and psychophysiological implications. Psychophysiology. 1993;30(2):183–196. doi: 10.1111/j.1469-8986.1993.tb01731.x. [DOI] [PubMed] [Google Scholar]
- Berntson GG, Cacioppo JT, Quigley KS, Fabro VT. Autonomic space and psychophysiological response. Psychophysiology. 1994;31(1):44–61. doi: 10.1111/j.1469-8986.1994.tb01024.x. [DOI] [PubMed] [Google Scholar]
- Beauchaine T. Vagal tone, development, and Gray's motivational theory: Toward an integrated model of autonomic nervous system functioning in psychopathology. Development and psychopathology. 2001;13(02):183–214. doi: 10.1017/s0954579401002012. [DOI] [PubMed] [Google Scholar]
- Benjamin LS. A clinician-friendly version of the Interpersonal Circumplex: Structural Analysis of Social Behavior (SASB) Journal of Personality Assessment. 1996;66(2):248–266. doi: 10.1207/s15327752jpa6602_5. [DOI] [PubMed] [Google Scholar]
- Brooker RJ, Buss KA. Dynamic measures of RSA predict distress and regulation in toddlers. Developmental psychobiology. 2010;52(4):372–382. doi: 10.1002/dev.20432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bugental DB. Predicting & preventing child maltreatment: A biocognitive transactional approach. In: Sameroff A, editor. The transactional model of development: How children and contexts shape each other. Washington, DC: American Psychological Association; 2009. pp. 97–115. [Google Scholar]
- Bush NR, Alkon A, Obradović J, Stamperdahl J, Boyce WT. Differentiating challenge reactivity from psychomotor activity in studies of children's psychophysiology: Considerations for theory and measurement. Journal of experimental child psychology. 2011;110(1):62–79. doi: 10.1016/j.jecp.2011.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Butler EA, Wilhelm FH, Gross JJ. Respiratory sinus arrhythmia, emotion, and emotion regulation during social interaction. Psychophysiology. 2006;43:612–622. doi: 10.1111/j.1469-8986.2006.00467.x. [DOI] [PubMed] [Google Scholar]
- Calkins SD, Graziano PA, Berdan LE, Keane SP, Degnan KA. Predicting cardiac vagal regulation in early childhood from maternal-child relationship quality during toddlerhood. Developmental Psychobiology. 2008;50(8):751–766. doi: 10.1002/dev.20344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaffin M, Funderburk B, Bard D, Valle LA, Gurwitch R. A combined motivation and parent–child interaction therapy package reduces child welfare recidivism in a randomized dismantling field trial. Journal of Consulting and Clinical Psychology. 2011;79(1):84. doi: 10.1037/a0021227. [DOI] [PubMed] [Google Scholar]
- Chaffin M, Silovsky JF, Funderburk B, Valle LA, Brestan EV, Balachova T, Bonner BL. Parent-child interaction therapy with physically abusive parents: efficacy for reducing future abuse reports. Journal of consulting and clinical psychology. 2004;72(3):500. doi: 10.1037/0022-006X.72.3.500. [DOI] [PubMed] [Google Scholar]
- Cicchetti D, Toth SL. Child Maltreatment. Annual Review of Clinical Psychology. 2005;1:409–438. doi: 10.1146/annurev.clinpsy.1.102803.144029. [DOI] [PubMed] [Google Scholar]
- Cohen J. Weighted kappa: Nominal scale agreement provision for scaled disagreement or partial credit. Psychological bulletin. 1968;70(4):213. doi: 10.1037/h0026256. [DOI] [PubMed] [Google Scholar]
- Cohn JF, Tronick EZ. Mother–infant face-to-face interaction: The sequence of dyadic states at 3, 6, and 9 months. Developmental Psychology. 1987;23(1):68. [Google Scholar]
- Cohn JF, Tronick EZ. Mother-infant face-to-face interaction: Influence is bidirectional and unrelated to periodic cycles in either partner's behavior. Developmental psychology. 1988;24(3):386. [Google Scholar]
- Deater-Deckard K, Sewell MD, Petrill SA, Thompson LA. Maternal working memory and reactive negativity in parenting. Psychological Science. 2010;21:75–79. doi: 10.1177/0956797609354073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denver JW, Reed SF, Porges SW. Methodological issues in the quantification of respiratory sinus arrhythmia. Biological Psychology. 2007;74:286–294. doi: 10.1016/j.biopsycho.2005.09.00. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dumas JE, Wahler RG. Indiscriminate mothering as a contextual factor in aggressive-oppositional child behavior:“Damned if you do and damned if you don't”. Journal of Abnormal Child Psychology. 1985;13(1):1–17. doi: 10.1007/BF00918368. [DOI] [PubMed] [Google Scholar]
- Feldman R. Parent–Infant Synchrony Biological Foundations and Developmental Outcomes. Current directions in psychological science. 2007;16(6):340–345. [Google Scholar]
- Feldman R, Singer M, Zagoory O. Touch attenuates infants' physiological reactivity to stress. Developmental Science. 2010;13:271–278. doi: 10.1111/j.1467-7687.2009.00890.x. [DOI] [PubMed] [Google Scholar]
- Fortunato CK, Gatzke-Kopp LM, Ram N. Associations between respiratory sinus arrhythmia reactivity and internalizing and externalizing symptoms are emotion specific. Cognitive, Affective, & Behavioral Neuroscience. 2013;13(2):238–251. doi: 10.3758/s13415-012-0136-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox NA. Heart-rate variability and behavioral reactivity: Individual differences in autonomic patterning and their relation to infant and child temperament. In: Reznick JS, editor. Perspectives on behavioral inhibition The John D and Catherine T MacArthur Foundation Series on Mental Health and Development. Chicago, IL: University of Chicago Press; 1989. pp. 177–195. [Google Scholar]
- Friedman BH. An autonomic flexibility–neurovisceral integration model of anxiety and cardiac vagal tone. Biological psychology. 2007;74(2):185–199. doi: 10.1016/j.biopsycho.2005.08.009. [DOI] [PubMed] [Google Scholar]
- Gianino A, Tronick EZ. The mutual regulation model: The infant's self and interactive regulation and coping and defensive capacities 1988 [Google Scholar]
- Geisler F, Kubiak T, Siewert K, Weber H. Cardiac vagal tone is associated with social engagement and self-regulation. Biological psychology. 2013;93(2):279–286. doi: 10.1016/j.biopsycho.2013.02.013. [DOI] [PubMed] [Google Scholar]
- Graziano P, Derefinko K. Cardiac vagal control and children's adaptive functioning: A meta-analysis. Biological psychology. 2013;94(1):22–37. doi: 10.1016/j.biopsycho.2013.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grossman P, Taylor EW. Toward understanding respiratory sinus arrhythmia: Relations to cardiac vagal tone, evolution and biobehavioral functions. Biological Psychology. 2007;74:263–285. doi: 10.1016/j.biopsycho.2005.11.014. [DOI] [PubMed] [Google Scholar]
- Harrist AW, Pettit GS, Dodge KA, Bates JE. Dyadic synchrony in mother-child interaction: Relation with children's subsequent kindergarten adjustment. Family Relations. 1994:417–424. [Google Scholar]
- Harrist AW, Waugh RM. Dyadic synchrony: Its structure and function in children's development. Developmental Review. 2002;22(4):555–592. [Google Scholar]
- Hastings PD, Nuselovici JN, Utendale WT, Coutya J, McShane KE, Sullivan C. Applying the polyvagal theory to children's emotion regulation: Social context, socialization, and adjustment. Biological Psychology. 2008;79(3):299–306. doi: 10.1016/j.biopsycho.2008.07.005. [DOI] [PubMed] [Google Scholar]
- Hill-Soderlund AL, Mills-Koonce WR, Propper C, Calkins SD, Granger DA, Moore GA, Cox MJ. Parasympathetic and sympathetic responses to the strange situation in infants and mothers from avoidant and securely attached dyads. Developmental Psychobiology. 2008;50(4):361–376. doi: 10.1002/dev.20302. [DOI] [PubMed] [Google Scholar]
- Holt-Lunstad J, Uchino BN, Smith TW, Hicks A. On the importance of relationship quality: the impact of ambivalence in friendships on cardiovascular functioning. Annals of Behavioral Medicine. 2007;33:278–290. doi: 10.1007/BF02879910. [DOI] [PubMed] [Google Scholar]
- Houtveen JH, Rietveld S, de Geus EJC. Contribution of tonic vagal modulation of heart rate, central respiratory drive, respiratory depth, and respiratory frequency to respiratory sinus arrhythmia during mental stress and physical exercise. Psychophysiology. 2002;39:427–436. doi: 10.1017.S0048577202394022. [DOI] [PubMed] [Google Scholar]
- Humphrey LL, Benjamin LS. Using structural analysis of social behavior to assess critical but elusive family processes: A new solution to an old problem. American Psychologist. 1986;41(9):979. doi: 10.1037//0003-066x.41.9.979. [DOI] [PubMed] [Google Scholar]
- Ingjaldsson J, Laberg JC, Thayer JF. Reduced heart rate variability in chronic alcohol abuse: relationship with negative mood, chronic thought suppression, and compulsive drinking. Biological Psychiatry. 2003;54:1427–1436. doi: 10.1016/s0006-3223(02)01926-1. [DOI] [PubMed] [Google Scholar]
- Joosen KJ, Mesman J, Bakermans-Kranenburg MJ, van IJzendoorn MH. Maternal overreactive sympathetic nervous system responses to repeated infant crying predicts risk for impulsive harsh discipline of infants. Child Maltreatment. 2013;18(4) doi: 10.1177/1077559513494762. [DOI] [PubMed] [Google Scholar]
- Kaplow JB, Widom CS. Age of onset of child maltreatment predicts long-term mental health outcomes. Journal of Abnormal Psychology. 2007;116(1):176–187. doi: 10.1037/0021-843X.116.1.176. [DOI] [PubMed] [Google Scholar]
- Knutson JF, DeGarmo D, Koeppl G, Reid JB. Care neglect, supervisory neglect, and harsh parenting in the development of children's aggression: A replication and extension. Child maltreatment. 2005;10(2):92–107. doi: 10.1177/1077559504273684. [DOI] [PubMed] [Google Scholar]
- Kok BE, Fredrickson BL. Upward spirals of the heart: Autonomic flexibility, as indexed by vagal tone, reciprocally and prospectively predicts positive emotions and social connectedness. Biological Psychology. 2010;85:432–436. doi: 10.1016/j.biopsycho.2010.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landis JR, Koch GG. The measurement of observer agreement for categoricaldata. Biometrics. 1977;33:159–174. [PubMed] [Google Scholar]
- Light KC, Kothandapani RV, Allen MT. Enhanced cardiovascular and catecholamine responses in women with depressive symptoms. International Journal of Psychophysiology. 1998;28(2):157–166. doi: 10.1016/s0167-8760(97)00093-7. [DOI] [PubMed] [Google Scholar]
- MacLeod J, Nelson G. Programs for the promotion of family wellness and the prevention of child maltreatment: A meta-analytic review. Child abuse & neglect. 2000;24(9):1127–1149. doi: 10.1016/s0145-2134(00)00178-2. [DOI] [PubMed] [Google Scholar]
- McCanne TR, Hagstrom AH. Physiological hyperreactivity to stressors in physical child abusers and individuals at risk for being physically abusive. Aggression and Violent Behavior. 1996;1:345–358. [Google Scholar]
- Mezzacappa E, Kindlon D, Saul JP, Earls F. Executive and motivational control of performance task behavior, and autonomic heart-rate regulation in children: physiologic validation of two-factor solution inhibitory control. Journal of Child Psychology and Psychiatry. 1998;39(04):525–531. [PubMed] [Google Scholar]
- Miller JG, Chocol C, Nuselovici JN, Utendale WT, Simard M, Hastings PD. Children's dynamic RSA change during anger and its relations with parenting, temperament, and control of aggression. Biological psychology. 2013;92(2):417–425. doi: 10.1016/j.biopsycho.2012.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mills-Koonce WR, Propper C, Gariepy JL, Barnett M, Moore GA, Calkins S, Cox MJ. Psychophysiological correlates of parenting behavior in mothers of young children. Developmental psychobiology. 2009;51(8):650–661. doi: 10.1002/dev.20400. [DOI] [PubMed] [Google Scholar]
- Moore GA, Calkins SD. Infants' vagal regulation in the still-face paradigm is related to dyadic coordination of mother-infant interaction. Developmental Psychology. 2004;40:1068–1080. doi: 10.1037/0012-1649.40.6.1068. [DOI] [PubMed] [Google Scholar]
- Moore GA, Hill-Soderlund AL, Propper CB, Calkins SD, Mills-Koonce WR, Cox MJ. Mother-infant vagal regulation in the face-to-face still-face paradigm is moderated by maternal sensitivity. Child Development. 2009;80:209–223. doi: 10.1111/j.1467-8624.2008.01255.x. [DOI] [PubMed] [Google Scholar]
- Musser ED, Ablow JC, Measelle JR. Predicting maternal sensitivity: The roles of postnatal depressive symptoms and parasympathetic dysregulation. Infant Mental Health Journal. 2012;33(4):350–359. doi: 10.1002/imhj.21310. [DOI] [PubMed] [Google Scholar]
- Oppenheimer JE, Measelle JR, Laurent HK, Ablow JC. Mothers' vagal regulation during the still-face paradigm: normative reactivity and impact of depression symptoms. Infant Behavior and Development. 2013;36(2):255–267. doi: 10.1016/j.infbeh.2013.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollak SD. Mechanisms linking early experience and the emergence of emotions. Psychological Science. 2008;17(6):370–375. doi: 10.1111/j.1467-8721.2008.00608.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollak SD, Cicchetti D, Hornung K, Reed A. Recognizing emotion in faces: Developmental effects of child abuse and neglect. Developmental Psychology. 2000;36:679–688. doi: 10.1037/0012-1649.36.5.679. [DOI] [PubMed] [Google Scholar]
- Porges SW. Orienting in a defensive world: Mammalian modifications of our evolutionary heritage. A polyvagal theory. Psychophysiology. 1995;32(4):301–318. doi: 10.1111/j.1469-8986.1995.tb01213.x. [DOI] [PubMed] [Google Scholar]
- Porges SW. The polyvagal theory: phylogenetic substrates of a social nervous system. International Journal of Psychophysiology. 2001;42(2):123–146. doi: 10.1016/s0167-8760(01)00162-3. [DOI] [PubMed] [Google Scholar]
- Porter CL. Coregulation in mother-infant dyads: links to infants' cardiac vagal tone. Psychological Reports. 2003;92(1):307–319. doi: 10.2466/pr0.2003.92.1.307. [DOI] [PubMed] [Google Scholar]
- Raudenbush SW, Bryk AS. Hierarchical Linear Models: Applications and Data Analysis Methods. Newbury Park, CA: Sage; 2002. p. 2002. [Google Scholar]
- Reid JB, Patterson GR, Loeber R. Nebraska symposium on motivation. University of Nebraska Press; 1981. The abused child: Victim, instigator, or innocent bystander? [PubMed] [Google Scholar]
- Rocissano L, Slade A, Lynch V. Dyadic synchrony and toddler compliance. Developmental Psychology. 1987;23(5):698. [Google Scholar]
- Rothbaum F, Weisz JR. Parental caregiving and child externalizing behavior in nonclinical samples: A meta-analysis. Psychological Bulleting. 1994;116:55–74. doi: 10.1037/0033-2909.116.1.55. [DOI] [PubMed] [Google Scholar]
- Skowron EA, Cipriano-Essel E, Benjamin LS, Pincus AL, Van Ryzin MJ. Cardiac vagal tone and quality of parenting show concurrent and time-ordered associations that diverge in abusive, neglectful, and non-maltreating mothers. Couple and Family Psychology: Research and Practice. 2013;2(2):95–115. doi: 10.1037/cfp0000005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skowron EA, Kozlowski JM, Pincus AL. Differentiation, self–other representations, and rupture–repair processes: Predicting child maltreatment risk. Journal of counseling psychology. 2010;57(3):304. doi: 10.1037/a0020030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skowron EA, Loken E, Gatzke-Kopp LM, Cipriano-Essel EA, Woehrle PL, Van Epps JJ, Gowda A, Ammerman RT. Mapping cardiac physiology and parenting prcoesses in maltreating mother-child dyads. Journal of Family Psychology. 2011;25(5):663–674. doi: 10.1037/a0024528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skowron E, Reinemann DH. Effectiveness of Psychological Interventions for Child Maltreatment: A Meta-Analysis. Psychotherapy: Theory, research, practice, training. 2005;42(1):52. [Google Scholar]
- Smith TW, Cribbet MR, Nealey-Moore JB, Uchino BN, Williams PG, MacKenzie J, Thayer JF. Matters of the variable heart: Respiratory sinus arrhythmia response to marital interaction and associations with marital quality. Journal of Personality and Social Psychology. 2012;100:103–119. doi: 10.1037/a0021136. [DOI] [PubMed] [Google Scholar]
- Stern SB, Azar ST. Integrating cognitive strategies into behavioral treatment for abusive parents and families with aggressive adolescents. Clinical Child Psychology and Psychiatry. 1998;3(3):387–403. [Google Scholar]
- Thayer JF, Lane RD. A model of neurovisceral integration in emotion regulation and dysregulation. Journal of affective disorders. 2000;61(3):201–216. doi: 10.1016/s0165-0327(00)00338-4. [DOI] [PubMed] [Google Scholar]
- Tronick EZ. Emotions and emotional communication in infants. American psychologist. 1989;44(2):112. doi: 10.1037//0003-066x.44.2.112. [DOI] [PubMed] [Google Scholar]
- Wahler RG, Williams AJ, Cerezo A. The compliance and predictability hypotheses: Sequential and correlational analyses of coercive mother-child interactions. Behavioral Assessment 1990 [Google Scholar]
- Weisman O, Zagoory-Sharon O, Feldman R. Oxytocin administration to parent enhances infant physiological and behavioral readiness for social engagement. Biological psychiatry. 2012;72(12):982–989. doi: 10.1016/j.biopsych.2012.06.011. [DOI] [PubMed] [Google Scholar]
- Whiteman J, Fanshel D, Grundy JF. Cognitive-behavioral interventions aimed at anger of parents at risk of child abuse. Social Work. 1987;32:469–474. [Google Scholar]
- Wilson SR, Rack JJ, Shi X, Norris AM. Comparing physically abusive, neglectful, and non-maltreating parents during interactions with their children: A meta-analysis of observational studies. Child Abuse & Neglect. 2008;32(9):897–911. doi: 10.1016/j.chiabu.2008.01.003. [DOI] [PubMed] [Google Scholar]