

Abstract
We aimed to identify bone related markers in the peripheral blood of osteoporotic (OP) patients that pointed toward molecular mechanisms underlying late postmenopausal bone loss. Whole blood from 22 late postmenopausal OP patients and 26 healthy subjects was examined. Bone mineral density (BMD) was measured by DXA. Protein levels of p70-S6K, p21, MMP-9, TGFβ1, and caspase-3 were quantified by ELISA. Gene expression was measured using real-time RT-PCR. OP registered by low BMD indices in late postmenopausal patients was associated with a significant upregulation of autophagy protein ULK1, cyclin-dependent kinase inhibitor p21, and metalloproteinase MMP-9 gene expression in the blood compared to the healthy controls and in a significant downregulation of mTOR (mammalian target of rapamycin), RUNX2, and ALPL gene expression, while expression of cathepsin K, caspase-3, transforming growth factor (TGF) β1, interleukin- (IL-) 1β, and tumor necrosis factor α (TNFα) was not significantly affected. We also observed a positive correlation between TGFβ1 and RUNX2 expression and BMD at femoral sites in these patients. Therefore, bone loss in late postmenopausal OP patients is associated with a significant upregulation of survival-related genes (ULK1 and p21) and MMP-9, as well as the downregulation of mTOR and osteoblast differentiation-related genes (RUNX2 and ALPL) in the peripheral blood compared to the healthy controls.
1. Introduction
Osteoporosis (OP) is characterized by a low bone mass, poor bone quality, and an increased propensity to fracture. Osteoporotic fractures, which are often associated with decreased bone mineral density (BMD), are a major public health problem in the aging population [1]. However, the molecular mechanisms underlying OP related bone destruction are not completely understood at the present time. Recently it has been suggested that these mechanisms involve altered osteoblast and osteoclast differentiation and function [2].
Excessive bone degradation in OP is thought to result from an increased activity of resorptive osteoclasts, which express proteases such as metalloproteinase (MMP-9) and cathepsin K. Cathepsin K, a papain-like cysteine protease, is considered a key player in the process of bone resorption [3, 4]. Osteoclast differentiation and function are regulated by the tumor necrosis factor α (TNFα), receptor activator of nuclear factor kappa B ligand (RANKL), and macrophage colony-stimulating factor (M-CSF). These three cytokines may promote osteoclast survival by signaling through mammalian target of rapamycin (mTOR) as a common target [5]. mTOR is recognized as an evolutionarily conserved central coordinator of fundamental biological processes involving cell cycle progression, translational control, ribosomal biogenesis, transcription control, and autophagy [6]. Indeed, recent animal studies have shown that mTOR is capable of regulating osteoclastogenesis, while the downregulation of mTOR reduced osteoclast production and activity [7, 8].
Differentiation-related phenotypic alterations in osteoblasts include changes in specific gene expression and corresponding protein synthesis during the course of proliferation, extracellular matrix maturation, and mineralization [9, 10]. Initially, actively proliferating cells express cell cycle and cell growth regulating genes such as Runt-related transcription factor (RUNX) 2, an osteoblast transcriptional regulator, which also controls the expression of major bone matrix protein genes [11]. Osteoblast maturation is associated with production of the bone matrix from regularly and densely packed collagen fibrils and high mineralization. At this stage, the mechanical properties and composition of the bone matrix are regulated by transforming growth factor (TGF)β1 [12]. A decline in proliferative activity is accompanied by the enhanced expression of the CDK inhibitor p21(CIP1/WAF1) (p21) and alkaline phosphatase [9, 13]. As a caspase-3 substrate, physiological amounts of p21 are responsible for the apoptotic activity observed in many cell lines, including osteoblasts [14].
Alterations in mTOR signaling may also be involved in osteoblast phenotypic conversions [15, 16]. It has been shown that upregulation of mTOR signaling was associated with an increase in BMP2, RUNX2, and TGFβ expression in preosteoblasts [17]. In contrast, mTOR downregulation by rapamycin inhibited osteoblast proliferation and differentiation and reduced RUNX2, sialoprotein, and osterix gene expression, alkaline phosphatase activity, and mineralization capacity [18].
Furthermore, the downregulation of mTOR is accompanied by increased autophagy in many cell types [19]. Autophagy is a physiological cellular mechanism that degrades and recycles proteins to maintain an adequate amino acid level for survival purposes. It involves the formation of cytosolic double membrane vesicles (autophagosomes) associated with the upregulation of ULK (hATG)1–15 gene expression. Hyperautophagic conditions are capable of promoting caspase-dependent apoptotic cell death [20]. Recently, the importance of autophagy in OP development and progression was described [21, 22]. In particular, the regulation of autophagy (ROA) pathway was shown to be significantly associated with wrist and arm BMD [23]. Moreover, autophagic proteins are important for the generation of the osteoclast ruffled border, their secretory function, and bone resorption [24].
The interplay of anabolic and catabolic factors, which are involved in major metabolic pathways and are associated with osteoporotic bone loss, is currently unclear. Clinical studies might help clarify this issue. However, bone specimens from postmenopausal OP patients are largely unavailable. On the other hand, it is well established that the number of genes simultaneously expressed in various cell types is higher in developmentally related tissues. As immune and bone cells originate from the mesoderm and the formation of bone and adaptive immune systems are phylogenetically closely related, these systems might involve identical regulatory cytokines and growth factors [25]. Moreover, it was suggested recently that peripheral blood mononuclear cells (PBMCs) contain substantial numbers of T-lymphocytes, which are capable of producing the proinflammatory cytokines IL-1 and TNFα, which could be involved in postmenopausal bone loss [26, 27].
Here, we compared expression of the genes associated with bone cell differentiation and bone resorption (RUNX2, TGFβ1, ALPL, TNFα, IL-1β, MMP-9, and cathepsin K) and the genes responsible for global cell survival and functioning (mTOR, p21, caspase-3, and ULK1) in the peripheral blood of late postmenopausal OP patients and healthy subjects. We observed a significant upregulation of ULK1, p21, and MMP-9 gene expression in the blood of the OP women compared to the healthy controls. This was associated with a significant downregulation of mTOR, RUNX2, and alkaline phosphatase (ALPL) gene expression, while the expression of cathepsin K, caspase-3, IL-1β, and TNFα was not significantly different than that of the healthy subjects. We also observed a positive correlation between TGFβ1 and RUNX2 expression and the BMD at femoral sites in these patients. We concluded that late postmenopausal osteoporosis is associated with an upregulation of cell survival as well as a downregulation of cell growth/proliferation and osteoblast differentiation/function related gene expression as registered in the peripheral blood cells.
2. Patients and Methods
2.1. Ethics
The study protocol was approved by the Local Committee on the Ethics of Human Research and informed consent was obtained from all subjects.
2.2. Patients
The study included 22 consecutive, unrelated, late postmenopausal, Russian women with idiopathic osteoporosis who visited the outpatient clinic of the Nasonova Research Institute of Rheumatology. The average age of the OP patients was 66.1 ± 7.2 years, with a range of 53–76 years of age. The average menopause duration was 18.0 ± 4.8 years, with a range of 10–30 years. Individuals with disorders known to cause abnormalities in bone metabolism, including diabetes mellitus, renal diseases, rheumatoid arthritis, and thyroid, parathyroid, and other endocrinological diseases, were excluded from the study. Women that had taken drugs, such as estrogen, progesterone, glucocorticoids, bisphosphonates, and alfacalcidol, were also excluded.
Twenty-six age-matched postmenopausal healthy volunteers (average age 63.0 ± 12.2 years, with a range of 49–78 years of age) who did not have any serious diseases, including osteoarthritis, and had not taken drugs known to affect bone and calcium metabolism were also recruited in the Moscow area. The study protocol was approved by the Local Committee on the Ethics of Human Research and informed consent was obtained from all subjects. The study was conducted in full accordance with the current version (2008) of the Declaration of Helsinki.
2.3. Measurement of BMD
BMD of the lumbar spine (L1–L4), femoral neck, femoral trochanter, femoral intertrochanter, Ward triangle, and total femur was measured by dual-energy X-ray absorptiometry (DXA) using a QDR-4500w instrument (Hologic, USA) at the Nasonova Institute of Rheumatology. The diagnosis of osteoporosis was based on the criteria recommended by the World Health Organization [28], which included a T-score < −2.5 SD. According to this test, all the OP subjects examined in this study were diagnosed with osteoporosis (Table 1).
Table 1.
Average BMD values (±SD) in the examined postmenopausal osteoporotic women and healthy subjects.
| Healthy controls | T-score | OP patients | T-score | P | |
|---|---|---|---|---|---|
| (n = 26) | (n = 22) | ||||
| Lumbar spine L1–L4 |
1.052 ± 0.06 | 0.064 ± 0.62 | 0.712 ± 0.06 | −3.07 ± 0.60 | <0.001 |
| Femoral neck | 0.857 ± 0.06 | 0.064 ± 0.59 | 0.584 ± 0.08 | −2.407 ± 0.71 | <0.001 |
| Trochanter | 0.765 ± 0.11 | 0.633 ± 1.08 | 0.531 ± 0.07 | −1.703 ± 0.74 | <0.001 |
| Intertrochanter | 1.163 ± 0.10 | 0.525 ± 0.70 | 0.830 ± 0.16 | −1.696 ± 1.05 | <0.001 |
| Ward triangle | 0.727 ± 0.10 | −0.042 ± 0.91 | 0.378 ± 0.07 | −2.839 ± 1.24 | <0.001 |
| Total femur | 1.156 ± 0.12 | 0.533 ± 0.70 | 0.700 ± 0.11 | −1.853 ± 1.11 | <0.001 |
2.4. Peripheral Blood Mononuclear Cell (PBMC) Isolation
Peripheral blood (10 mL) was collected in Vacutainer tubes containing ethylenediaminetetraacetic acid (EDTA) (BDH, England). The blood samples were taken in a standardized manner in the morning (between 07:00 AM and 09:00 AM). Whole blood fractionation was performed using a Ficoll density gradient. Upon centrifugation the peripheral blood mononuclear cells (PBMCs) located in the interphase were collected and washed twice in phosphate-buffered saline (PBS) [29]. The obtained cell fractions were frozen and kept at −70°C prior to protein extraction.
2.5. Quantification of p70-S6K, p21, MMP-9, TGFβ1, and Caspase-3 Protein Levels
The concentrations of total p70-S6K (KHO0571), p21WAF1/Cip1 (KHO5421), MMP-9 (KHС3061), active caspase-3 (KHO1091) (Invitrogen, Camarillo, CA, USA), and TGFβ1 (BMS249/4) (eBioscience, Vienna, Austria) were determined in the isolated PBMCs using commercially available enzyme linked immunosorbent assay (ELISA) kits according to the manufacturer's instructions. For mTOR protein expression, we evaluated the levels of p70-S6K, an mTOR direct target for phosphorylation, which is used as a readout of mTOR activity [30, 31], as mTOR ELISA kits are not available in Russia.
The results were expressed per µg of protein measured in the PBMC lysates. The PBMC lysates were obtained using Cell Extraction Buffer containing 10 mM Tris, pH 7.4, 100 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM NaF, 20 mM Na4P2O7, 20 mM Na3VO4, 1% Triton X-100, 10% glycerol, 0.1% SDS, and 0.5% deoxycholate (Invitrogen, Camarillo, CA, USA) supplemented with Protease Inhibitor Cocktail (Sigma-Aldrich, Inc., St. Louis, USA) and 1 mM PMSF (Sigma-Aldrich, Inc., St. Louis, USA) according to the manufacturer's instructions. The total protein concentration in the cell lysates was quantified using the Bradford method [32].
2.6. Total RNA Isolation and Reverse Transcriptase (RT) Reaction
For detection of gene expression total RNA was isolated from 100 µL of whole blood immediately after withdrawal using Ribo-zol-A kit (InterLabService, Moscow, Russia) in accordance with the manufacturer's recommendations. Total RNA had an A260/290 > 1.9. The RT reaction was performed using a Reverta kit containing M-MLV Reverse Transcriptase, random hexanucleotide primers, and total RNA according to the manufacturer's recommendations (InterLabService, Moscow, Russia).
2.7. Real-Time Quantitative PCR
The following premade primers and probes were used for the TaqMan assay (Applied Biosystems, Foster City, CA, USA): mTOR (Hs00234522_m1), Unc-51-like kinase 1 (ULK1) (Hs00177504_m1), p21WAF1/Cip1 (p21) (Hs00355782_m1), caspase-3 (Hs00263337_m1), TNFα (Hs00174128_m1), TGFβ1 (Hs99999918_m1), RUNX2 (Hs00231692_m1), ALPL (Hs00758162_m1), cathepsin K (Hs00166156_m1), MMP-9 (Hs00234579_m1), and IL-1β (Hs00174097_m1). β-actin was used as an endogenous control.
The quantification of gene expression was conducted using a 7300 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) as described previously [33]. Briefly, 1 μL of RT product was subjected to real-time PCR in a 15 μL total reaction mixture containing 7.5 μL of TaqMan Universal PCR Master Mix (Applied Biosystems), 900 nM sense and antisense primers, 50 nM probe, and template cDNA. After a single step of 50°C for 2 min and an initial activation at 95°C for 10 min, the reaction mixtures were subjected to 40 amplification cycles (15 s at 95°C for denaturation and 1 min of annealing and extension at 60°C).
Relative mRNA expression was determined using the delta-delta CT method, as detailed by the manufacturer guidelines (Applied Biosystems) [34]. The delta CT value was calculated by subtracting the CT value for the housekeeping β-actin gene from the CT value for each sample. A delta-delta CT value was then calculated by subtracting the delta CT value of the control (each healthy patient) from the delta CT value of each OP patient. Each PCR was performed in duplicate. Three “no template” controls were consistently negative for each reaction.
2.8. Statistical Analysis
A Kolmogorov-Smirnov normality test showed that the data were distributed according to a Gaussian distribution curve. Therefore, for statistical evaluations, Pearson's rank correlations and unpaired Student's t-test were used for comparisons between the control subjects and the OP patients. As we compared only a control sample with OP sample in case of each gene and did not perform any multiple testing, no correction for multiple testing was made. Quantitative data were expressed as the mean ± SD. The Statistica 6 Software (StatSoft, Tulsa, OK, USA) was used for all statistical analyses. P values less than 0.05 were considered significant.
3. Results
3.1. Whole Blood Gene Expression
Examination of the gene expression in the whole blood of OP patients revealed that mTOR, RUNX2, and ALPL genes were significantly downregulated compared to the healthy subjects (Figure 1). In contrast the expression of the p21, autophagy-related ULK1, and MMP-9 genes was significantly upregulated in the same OP patients. No significant changes were observed in the expression of caspase-3, TNFα, IL-1β, TGFβ1, and cathepsin K.
Figure 1.
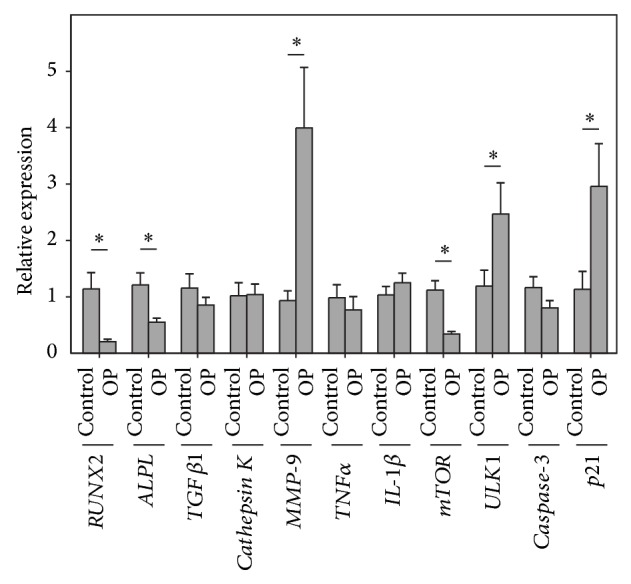
Relative expression of RUNX2, ALPL, TGFβ1, cathepsin K, MMP-9, TNFα, IL-1β, mTOR, ULK1, caspase-3, and p21 genes with reference to β-actin gene in the peripheral blood of 22 postmenopausal OP patients compared to 26 healthy age-matched control women (control). Each point represents the mean ± SD. Significant differences as compared to the control are indicated by asterisks.
3.2. Protein Levels of p70-S6K, p21, MMP-9, TGFβ1, and Caspase-3 in Isolated PBMC
To further investigate the clinical significance of mTOR, p21, MMP-9, TGFβ1, and caspase-3 gene expression in the whole blood of OP patients, we analyzed the protein levels of total p70-S6K serine/threonine kinase (a direct target of mTOR phosphorylation [30, 31]), p21, MMP-9, TGFβ1, and active caspase-3 in the PBMC fraction. These studies showed that OP patients demonstrated significantly lower p70-S6K protein concentrations in PBMCs compared to the healthy subjects (Figure 2). At the same time, p21 and MMP-9 protein levels were significantly increased, while no changes were observed in the amounts of active caspase-3 and TGFβ1 in PBMCs of the OP patients, as compared to the healthy patients.
Figure 2.

Protein concentrations of p70-S6K, p21 (a), MMP-9, TGFβ1, and caspase-3 (b) measured by ELISA in the PBMCs from OP patients (n = 22) compared with the control subjects (n = 26). Asterisks indicate significant differences from the healthy control subjects.
3.3. Association of Gene Expression with BMD
The analysis of bivariate correlations using Pearson's correlation coefficient for the expression of the examined genes showed that mTOR, TGFβ1, RUNX2, TNFα, IL1β, cathepsin K, caspase-3, and p21 positively (P < 0.05) correlated with each other (Table 2). However, no correlation was observed between ULK and ALPL gene expression and the other genes.
Table 2.
Correlation coefficients and their significance (P) are shown for the expression of cell growth and differentiation-related genes in relation to BMD in postmenopausal osteoporotic women.
| mTOR | TGFβ1 | RUNX2 | TNFα | IL-1β | Caspase-3 | |
|---|---|---|---|---|---|---|
| RUNX2 | 0.768 | 0.768 | ||||
| (P < 0.001) | (P < 0.001) | |||||
|
| ||||||
| TNFα | 0.664 | 0.504 | ||||
| (P = 0.005) | (P = 0.03) | |||||
|
| ||||||
| IL-1β | 0.424 | 0.696 | 0.496 | 0.494 | ||
| (P = 0.06) | (P < 0.001) | (P = 0.01) | (P = 0.03) | |||
|
| ||||||
| Caspase-3 | 0.666 | 0.481 | 0.519 | 0.867 | 0.637 | |
| (P = 0.003) | (P = 0.03) | (P = 0.01) | (P < 0.001) | (P = 0.002) | ||
|
| ||||||
| p21 | 0.580 | 0.868 | 0.680 | 0.715 | 0.675 | 0.715 |
| (P = 0.009) | (P < 0.001) | (P = 0.001) | (P = 0.002) | (P = 0.001) | (P = 0.001) | |
|
| ||||||
| TGFβ1 | 0.632 | |||||
| P = 0.004 | ||||||
|
| ||||||
| Cathepsin K | 0.542 | 0.516 | ||||
| (P = 0.03) | (P = 0.07) | |||||
|
| ||||||
| Lumbar spine | ||||||
|
| ||||||
| Femoral neck | ||||||
|
| ||||||
| Trochanter | 0.548 | |||||
| (P = 0.01) | ||||||
|
| ||||||
| Ward triangle | 0.516 | |||||
| (P = 0.02) | ||||||
|
| ||||||
| Total femur | 0.524 | 0.605 | ||||
| (P = 0.02) | (P = 0.005) | |||||
The expression of some of the examined genes also significantly correlated with BMD in the OP patients (Table 2). A positive correlation was noted between TGFβ1 and RUNX2 gene expression with BMD of the total femur and its compartments (trochanter and Ward triangle) in the examined OP patients.
4. Discussion
Recently, the skeletal and immune systems have been thought to interact more than previously believed, and this interaction primarily involves regulatory aspects [35]. Moreover, T- and B-lymphocytes are thought to be key regulators of osteoclast and osteoblast formation, lifespan, and activity [26]. Therefore, we suggested that changes in gene and protein expression in the peripheral blood cells of osteoporotic patients might also be associated with the disease related metabolic alterations observed in the bone cells. Here, we used the PBMCs of late postmenopausal OP patients and healthy controls to investigate the differences in the expression of genes related to the regulation of bone cell differentiation and bone resorption.
We show that the expression of osteoclast differentiation and function related genes was either unaltered (cathepsin K, TNFα, and IL-1β) or upregulated (MMP-9) in the peripheral blood of the examined OP women compared to the healthy subjects. Previous studies have also noted that osteoporotic women had serum IL-1β levels similar to those of normal controls [36]. In addition, no difference in the frequency of TNFα expression was observed in the bone tissue of OP versus healthy women [37]. Upregulation of MMP-9 gene and protein expression in the PBMCs of the examined OP women is supported by the in situ hybridization results of others, which showed an increase in MMP-9 mRNA in osteoporotic bone tissue versus the normal controls [3]. However, some studies also reported decreased MMP-9 expression in the bone of the postmenopausal OP patients [38]. Although MMP-9 expression was upregulated in the PBMCs of the examined OP women, its proteolytic activity in osteoclasts might be limited by the unaltered expression of cathepsin K, as both proteinases are required to be equally expressed for tissue proteolysis [39]. At the same time, the osteoclast proteolytic activity might not be directly pursued in the peripheral blood as we did not observe any correlation of the examined osteoclast differentiation and function related genes with the BMD indices.
In contrast, significant downregulation of osteoblast differentiation-related RUNX2 and ALPL gene expression in the PBMCs of OP women compared to the healthy age-matched controls, as well as a positive association of RUNX2 gene expression with the BMD at femoral sites, might indicate a significant reduction in bone formation during late postmenopausal OP. Previously, a decrease in RUNX2 and ALPL expression was also observed in the bone tissue of OP patients compared to healthy subjects [38, 40]. In addition, a lower RUNX2 expression and a subsequently increased bone loss were observed after treating of rodents with bone resorptive agents, such as rosiglitazone [41]. Therefore, bone loss during late postmenopausal OP might be associated with decreased bone formation rather than with increased bone resorption activity.
Our study also shows that late postmenopausal bone loss is associated with alterations in the non-tissue-specific gene and protein expression in the peripheral blood. It is well established that many human diseases occur when the precise regulation of cell growth (cell mass/size) and proliferation (rates of cell division) is compromised [42]. Therefore, the significant downregulation of mTOR gene expression that was associated with the decreased expression of osteoblast differentiation and function related genes (RUNX2 and ALPL) in the PBMCs of late postmenopausal OP women compared to healthy subjects was not surprising. Our observation is supported by the previous observations that the upregulation of mTOR signaling is associated with an increase in bone forming activity in rats and cultured human osteoblast-like cells [43, 44]. In contrast, systemic administration of the mTOR inhibitor FK506 caused dramatic OP in animals [45, 46], which was accompanied by the downregulation of RUNX2 expression [47]. Moreover, immunosuppressant FK506 therapy resulted in severe bone loss and an increase in fracture incidence in 65% of the patients [48]. Furthermore, a positive correlation between mTOR gene expression and the genes responsible for osteoblast (RUNX2 and TGFβ1) and osteoclast (cathepsin K, TNFα, and IL-1β) related differentiation and activity indicates the association of bone loss during OP with a general declination of cell growth and proliferation activity in the examined women.
Significant upregulation of autophagy-related ULK1 gene expression in the examined OP women might involve an increase in cell maintenance, as autophagy has been shown to be associated with increased survival in many cell types [49]. Indeed, increased osteoclast survival was previously observed in association with rapid bone loss in glucocorticoid-induced OP in animal studies [50]. Other animal studies have shown that the downregulation of autophagy resulted in the inhibition of MCPIP (zinc finger CCCH-type containing 12A) induced expression of osteoclastic markers in monocytic osteoclast precursors [51]. Therefore, increased osteoclast survival in the examined OP patients may explain the excessive bone loss in the absence of the upregulation of the genes related to osteoclast differentiation and activity, such as cathepsin K and TNFα.
The upregulation of p21, in association with the mTOR inhibition, which was observed in the PBMCs of the examined OP women, was also noted in cultured cells from various lineages following treatment with the mTOR inhibitor, rapamycin [52]. On the other hand, the upregulation of mTOR signaling in response to oscillatory shear stress was associated with a decreased expression of p21 in human osteoblast-like cells [44]. In addition, our observation of decreased RUNX2 gene expression associated with p21 upregulation in the blood of OP patients is supported by a previous study that showed p21 promoter repression by RUNX2 in osteoblast lineage cells [53]. Several other studies have also demonstrated an association between p21 upregulation and the inhibition of osteoblast proliferation and differentiation and an increase in the mononuclear precursor cell differentiation into osteoclasts [54–56].
5. Conclusion
Here, we show that peripheral blood cells of postmenopausal OP women exhibit significant upregulation of the ULK1, p21, and MMP-9 genes. This is associated with a significant downregulation of mTOR, RUNX2, and ALPL gene expression. Although the peripheral levels of cytokines and other factors regulating bone turnover do not always reflect the levels observed within the bone, the correlation of TGFβ1 and RUNX2 gene expression with BMD suggests that the expression of these gene counterparts in the bone tissue might be similarly affected. Therefore, bone resorption in late postmenopausal OP might be associated with decreased bone formation and an increased survival of the bone degrading cells. Further experiments should be performed to confirm our observations.
Acknowledgment
This study was funded by the Russian Foundation for Basic Research (Project nos. 15-04-00546-a and 12-04-00038a to Elena V. Tchetina).
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Diab D. L., Watts N. B. Postmenopausal osteoporosis. Current Opinion in Endocrinology, Diabetes and Obesity. 2013;20(6):501–509. doi: 10.1097/01.med.0000436194.10599.94. [DOI] [PubMed] [Google Scholar]
- 2.Hopwood B., Tsykin A., Findlay D. M., Fazzalari N. L. Gene expression profile of the bone microenvironment in human fragility fracture bone. Bone. 2009;44(1):87–101. doi: 10.1016/j.bone.2008.08.120. [DOI] [PubMed] [Google Scholar]
- 3.Zhao H., Cai G., Du J., Xia Z., Wang L., Zhu T. Expression of matrix metalloproteinase-9 mRNA in osteoporotic bone tissues. Journal of Tongji Medical University. 1997;17(1):28–31. doi: 10.1007/BF02887998. [DOI] [PubMed] [Google Scholar]
- 4.Troen B. R. The role of cathepsin K in normal bone resorption. Drug News and Perspectives. 2004;17(1):19–28. doi: 10.1358/dnp.2004.17.1.829022. [DOI] [PubMed] [Google Scholar]
- 5.Glantschnig H., Fisher J. E., Wesolowski G., Rodan G. A., Reszka A. A. M-CSF, TNFα and RANK ligand promote osteoclast survival by signaling through mTOR/S6 kinase. Cell Death & Differentiation. 2003;10(10):1165–1177. doi: 10.1038/sj.cdd.4401285. [DOI] [PubMed] [Google Scholar]
- 6.Santulli G., Totary-Jain H. Tailoring mTOR-based therapy: molecular evidence and clinical challenges. Pharmacogenomics. 2013;14(12):1517–1526. doi: 10.2217/pgs.13.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Cejka D., Hayer S., Niederreiter B., et al. Mammalian target of rapamycin signaling is crucial for joint destruction in experimental arthritis and is activated in osteoclasts from patients with rheumatoid arthritis. Arthritis and Rheumatism. 2010;62(8):2294–2302. doi: 10.1002/art.27504. [DOI] [PubMed] [Google Scholar]
- 8.Smink J. J., Leutz A. Rapamycin and the transcription factor C/EBPβ as a switch in osteoclast differentiation: implications for lytic bone diseases. Journal of Molecular Medicine. 2010;88(3):227–233. doi: 10.1007/s00109-009-0567-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Stein G. S., Lian J. B., Owen T. A. Relationship of cell growth to the regulation of tissue-specific gene expression during osteoblast differentiation. The FASEB Journal. 1990;4(13):3111–3123. doi: 10.1096/fasebj.4.13.2210157. [DOI] [PubMed] [Google Scholar]
- 10.Balint E., Lapointe D., Drissi H., et al. Phenotype discovery by gene expression profiling: mapping of biological processes linked to BMP-2-mediated osteoblast differentiation. Journal of Cellular Biochemistry. 2003;89(2):401–426. doi: 10.1002/jcb.10515. [DOI] [PubMed] [Google Scholar]
- 11.Long F., Ornitz D. M. Development of the endochondral skeleton. Cold Spring Harbor Perspectives in Biology. 2013;5(1) doi: 10.1101/cshperspect.a008334.a008334 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Alliston T. N. TGF-β regulation of osteoblast diferentiation and bone matrix properties. Journal of Musculoskeletal Neuronal Interactions. 2006;6(4):349–350. [PubMed] [Google Scholar]
- 13.Matsumoto T., Sowa Y., Ohtani-Fujita N., et al. p53-independent induction of WAF1/Cip1 is correlated with osteoblastic differentiation by vitamin D3. Cancer Letters. 1998;129(1):61–68. doi: 10.1016/S0304-3835(98)00080-9. [DOI] [PubMed] [Google Scholar]
- 14.Chan W. H., Wu H. J., Shiao N. H. Apoptotic signaling in methylglyoxal-treated human osteoblasts involves oxidative stress, c-Jun N-terminal kinase, caspase-3, and p21-activated kinase 2. Journal of Cellular Biochemistry. 2007;100(4):1056–1069. doi: 10.1002/jcb.21114. [DOI] [PubMed] [Google Scholar]
- 15.Singha U. K., Jiang Y., Yu S., et al. Rapamycin inhibits osteoblast proliferation and differentiation in MC3T3-E1 cells and primary mouse bone marrow stromal cells. Journal of Cellular Biochemistry. 2008;103(2):434–446. doi: 10.1002/jcb.21411. [DOI] [PubMed] [Google Scholar]
- 16.Tseng W.-P., Yang S.-N., Lai C.-H., Tang C.-H. Hypoxia induces BMP-2 expression via ILK, Akt, mTOR, and HIF-1 pathways in osteoblasts. Journal of Cellular Physiology. 2010;223(3):810–818. doi: 10.1002/jcp.22104. [DOI] [PubMed] [Google Scholar]
- 17.Kim J., Jung Y., Sun H., et al. Erythropoietin mediated bone formation is regulated by mTOR signaling. Journal of Cellular Biochemistry. 2012;113(1):220–228. doi: 10.1002/jcb.23347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tang C.-H., Lu D.-Y., Tan T.-W., Fu W.-M., Yang R.-S. Ultrasound induces hypoxia-inducible factor-1 activation and inducible nitric-oxide synthase expression through the integrin/integrin-linked kinase/Akt/mammalian target of rapamycin pathway in osteoblasts. The Journal of Biological Chemistry. 2007;282(35):25406–25415. doi: 10.1074/jbc.m701001200. [DOI] [PubMed] [Google Scholar]
- 19.Lum J. J., DeBerardinis R. J., Thompson C. B. Autophagy in metazoans: cell survival in the land of plenty. Nature Reviews Molecular Cell Biology. 2005;6(6):439–448. doi: 10.1038/nrm1660. [DOI] [PubMed] [Google Scholar]
- 20.Bell B. D., Leverrier S., Weist B. M., et al. FADD and caspase-8 control the outcome of autophagic signaling in proliferating T cells. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(43):16677–16687. doi: 10.1073/pnas.0808597105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Manolagas S. C., Parfitt A. M. What old means to bone. Trends in Endocrinology and Metabolism. 2010;21(6):369–374. doi: 10.1016/j.tem.2010.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hocking L. J., Whitehouse C., Helfrich M. H. Autophagy: a new player in skeletal maintenance? Journal of Bone and Mineral Research. 2012;27(7):1439–1447. doi: 10.1002/jbmr.1668. [DOI] [PubMed] [Google Scholar]
- 23.Zhang L., Guo Y.-F., Liu Y.-Z., et al. Pathway-based genome-wide association analysis identified the importance of regulation-of-autophagy pathway for ultradistal radius BMD. Journal of Bone and Mineral Research. 2010;25(7):1572–1580. doi: 10.1002/jbmr.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.DeSelm C. J., Miller B. C., Zou W., et al. Autophagy proteins regulate the secretory component of osteoclastic bone resorption. Developmental Cell. 2011;21(5):966–974. doi: 10.1016/j.devcel.2011.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Müller B. Cytokine imbalance in non-immunological chronic disease. Cytokine. 2002;18(6):334–339. doi: 10.1006/cyto.2002.0882. [DOI] [PubMed] [Google Scholar]
- 26.Teitelbaum S. L. Postmenopausal osteoporosis, T cells, and immune dysfunction. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(48):16711–16712. doi: 10.1073/pnas.0407335101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Loverro G., Cantatore F. P., Sasanelli B., Carrozzo M., Boscia F. M., Selvaggi L. Role of interleukin 1-alpha and insulin like growth factor in post menopausal osteoporosis. Minerva Ginecologica. 1994;46(1-2):15–20. [PubMed] [Google Scholar]
- 28.World Health Organization Study Group. Assessment of Fracture Risk and Its Application for Screening for Postmenopausal Osteoporosis. Geneva, Switzerland: WHO; 1994. [PubMed] [Google Scholar]
- 29.Son B. K., Roberts R. L., Ank B. J., Stiehm E. R. Effects of anticoagulant, serum, and temperature on the natural killer activity of human peripheral blood mononuclear cells stored overnight. Clinical and Diagnostic Laboratory Immunology. 1996;3(3):260–264. doi: 10.1128/cdli.3.3.260-264.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Isotani S., Hara K., Tokunaga C., Inoue H., Avruch J., Yonezawa K. Immunopurified mammalian target of rapamycin phosphorylates and activates p70 S6 kinase αin vitro . The Journal of Biological Chemistry. 1999;274(48):34493–34498. doi: 10.1074/jbc.274.48.34493. [DOI] [PubMed] [Google Scholar]
- 31.Fingar D. C., Salama S., Tsou C., Harlow E., Blenis J. Mammalian cell size is controlled by mTOR and its downstream targets S6K1 and 4EBP1/eIF4E. Genes and Development. 2002;16(12):1472–1487. doi: 10.1101/gad.995802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bradford M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Analytical Biochemistry. 1976;72(1-2):248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 33.Tchetina E. V., Poole A. R., Zaitseva E. M., et al. Differences in Mammalian target of rapamycin gene expression in the peripheral blood and articular cartilages of osteoarthritic patients and disease activity. Arthritis. 2013;2013:14. doi: 10.1155/2013/461486.461486 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Livak K. J. ABI Prism 7700 Sequence Detection System. Foster City, Calif, USA: PE Applied Biosystems; 1997. Comparative Ct method. (User Bulletin no. 2). [Google Scholar]
- 35.Arron J. R., Choi Y. Bone versus immune system. Nature. 2000;408(6812):535–536. doi: 10.1038/35046196. [DOI] [PubMed] [Google Scholar]
- 36.Khosla S., Peterson J. M., Egan K., Jones J. D., Riggs B. L. Circulating cytokine levels in osteoporotic and normal women. The Journal of Clinical Endocrinology and Metabolism. 1994;79(3):707–711. doi: 10.1210/jc.79.3.707. [DOI] [PubMed] [Google Scholar]
- 37.Ralston S. H. Analysis of gene expression in human bone biopsies by polymerase chain reaction: evidence for enhanced cytokine expression in postmenopausal osteoporosis. Journal of Bone and Mineral Research. 1994;9(6):883–890. doi: 10.1002/jbmr.5650090614. [DOI] [PubMed] [Google Scholar]
- 38.Balla B., Kósa J. P., Kiss J., et al. Different gene expression patterns in the bone tissue of aging postmenopausal osteoporotic and non-osteoporotic women. Calcified Tissue International. 2008;82(1):12–26. doi: 10.1007/s00223-007-9092-3. [DOI] [PubMed] [Google Scholar]
- 39.Tanaka H., Tanabe N., Kawato T., et al. Nicotine affects bone resorption and suppresses the expression of cathepsin K, MMP-9 and vacuolar-type H+-ATPase d2 and actin organization in osteoclasts. PLoS ONE. 2013;8(3) doi: 10.1371/journal.pone.0059402.e59402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Patsch J. M., Kohler T., Berzlanovich A., et al. Trabecular bone microstructure and local gene expression in iliac crest biopsies of men with idiopathic osteoporosis. Journal of Bone and Mineral Research. 2011;26(7):1584–1592. doi: 10.1002/jbmr.344. [DOI] [PubMed] [Google Scholar]
- 41.Ali A. A., Weinstein R. S., Stewart S. A., Parfitt A. M., Manolagas S. C., Jilka R. L. Rosiglitazone causes bone loss in mice by suppressing osteoblast differentiation and bone formation. Endocrinology. 2005;146(3):1226–1235. doi: 10.1210/en.2004-0735. [DOI] [PubMed] [Google Scholar]
- 42.Tee A. R., Blenis J. mTOR, translational control and human disease. Seminars in Cell and Developmental Biology. 2005;16(1):29–37. doi: 10.1016/j.semcdb.2004.11.005. [DOI] [PubMed] [Google Scholar]
- 43.Ford-Hutchinson A. F., Ali Z., Lines S. E., Hallgrímsson B., Boyd S. K., Jirik F. R. Inactivation of Pten in osteo-chondroprogenitor cells leads to epiphyseal growth plate abnormalities and skeletal overgrowth. Journal of Bone and Mineral Research. 2007;22(8):1245–1259. doi: 10.1359/jbmr.070420. [DOI] [PubMed] [Google Scholar]
- 44.Lee D.-Y., Li Y.-S. J., Chang S.-F., et al. Oscillatory flow-induced proliferation of osteoblast-like cells is mediated by αvβ3 and β1 integrins through synergistic interactions of focal adhesion kinase and Shc with phosphatidylinositol 3-kinase and the Akt/mTOR/p70S6K pathway. The Journal of Biological Chemistry. 2010;285(1):30–42. doi: 10.1074/jbc.m109.010512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sass D. A., Bowman A. R., Yuan Z., Ma Y., Jee W. S. S., Epstein S. Alendronate prevents cyclosporin A-induced osteopenia in the rat. Bone. 1997;21(1):65–70. doi: 10.1016/s8756-3282(97)00071-9. [DOI] [PubMed] [Google Scholar]
- 46.Cvetkovic M., Mann G. N., Romero D. F., et al. The deleterious effects of long-term cyclosporine A, cyclosporine G, and FK506 on bone mineral metabolism in vivo . Transplantation. 1994;57(8):1231–1237. doi: 10.1097/00007890-199404270-00016. [DOI] [PubMed] [Google Scholar]
- 47.Sun L., Blair H. C., Peng Y., et al. Calcineurin regulates bone formation by the osteoblast. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(47):17130–17135. doi: 10.1073/pnas.0508480102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Epstein S., Inzerillo A. M., Caminis J., Zaidi M. Disorders associated with acute rapid and severe bone loss. Journal of Bone and Mineral Research. 2003;18(12):2083–2094. doi: 10.1359/jbmr.2003.18.12.2083. [DOI] [PubMed] [Google Scholar]
- 49.Neufeld T. P. Contribution of Atg1-dependent autophagy to TOR-mediated cell growth and survival. Autophagy. 2007;3(5):477–479. doi: 10.4161/auto.4348. [DOI] [PubMed] [Google Scholar]
- 50.Weinstein R. S., Chen J.-R., Powers C. C., et al. Promotion of osteoclast survival and antagonism of bisphosphonate-induced osteoclast apoptosis by glucocorticoids. The Journal of Clinical Investigation. 2002;109(8):1041–1048. doi: 10.1172/jci200214538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Wang K., Niu J., Kim H., Kolattukudy P. E. Osteoclast precursor differentiation by MCPIP via oxidative stress, endoplasmic reticulum stress, and autophagy. Journal of Molecular Cell Biology. 2011;3(6):360–368. doi: 10.1093/jmcb/mjr021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Costa L. F., Balcells M., Edelman E. R., Nadler L. M., Cardoso A. A. Proangiogenic stimulation of bone marrow endothelium engages mTOR and is inhibited by simultaneous blockade of mTOR and NF-κB. Blood. 2006;107(1):285–292. doi: 10.1182/blood-2005-06-2208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Westendorf J. J., Zaidi S. K., Cascino J. E., et al. Runx2 (Cbfa1, AML-3) interacts with histone deacetylase 6 and represses the p21CIP1/WAF1 promoter. Molecular and Cellular Biology. 2002;22(22):7982–7992. doi: 10.1128/mcb.22.22.7982-7992.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Bellosta P., Masramon L., Mansukhani A., Basilico C. p21WAF1/CIP1 acts as a brake in osteoblast differentiation. Journal of Bone and Mineral Research. 2003;18(5):818–826. doi: 10.1359/jbmr.2003.18.5.818. [DOI] [PubMed] [Google Scholar]
- 55.Kwak H. B., Jin H.-M., Ha H., et al. Tumor necrosis factor-alpha induces differentiation of human peripheral blood mononuclear cells into osteoclasts through the induction of p21 (WAF1/Cip1) Biochemical and Biophysical Research Communications. 2005;330(4):1080–1086. doi: 10.1016/j.bbrc.2005.03.085. [DOI] [PubMed] [Google Scholar]
- 56.Sankar U., Patel K., Rosol T. J., Ostrowski M. C. RANKL coordinates cell cycle withdrawal and differentiation in osteoclasts through the cyclin-dependent kinase inhibitors p27KIP1 and p21 CIP1. Journal of Bone and Mineral Research. 2004;19(8):1339–1348. doi: 10.1359/jbmr.040321. [DOI] [PubMed] [Google Scholar]