

Abstract
Herein we compare the effects of perturbations in the Kölliker–Fuse nucleus (KFN) and the lateral (LPBN) and medial (MPBN) parabrachial nuclei on the coordination of breathing and swallowing. Cannula was chronically implanted in goats through which ibotenic acid (IA) was injected while awake. Swallows in late expiration (E) always reset while swallows in early inspiration (I) never reset the respiratory rhythm. Before cannula implantation, all other E and I swallows did not reset the respiratory rhythm, and had small effects on E and I duration and tidal volume (VT). However, after cannula implantation in the MPBN and KFN, E and I swallows reset the respiratory rhythm and increased the effects on I and E duration and VT. Subsequent injection of IA into the KFN eliminated the respiratory phase resetting of swallows but exacerbated the effects on I and E duration and VT. We conclude that the KFN and to a lesser extent the MPBN contribute to coordination of breathing and swallowing.
1. Introduction
In animals which use a common oropharyngolaryngeal passage for breathing and swallowing, the coordination of each activity is of vital importance to sustaining life and airway protection. Proper synchronization of functionally shared passages is reliant upon common neurological pathways originating from brainstem nuclei (Bolser et al., 2006), with loss of regulation potentially leading to a range of negative sequelae including aspiration, malnutrition, and dehydration (Prasse and Kikano, 2009; Shaker et al., 1992). Indeed, presumably impaired airway protection resulting in aspiration pneumonia is the leading cause of death in Parkinson’s and Alzheimer’s diseases (Kalia, 2003) underscoring the importance of the coordination of these behaviors.
We have previously reported evidence supporting site-specific cardiorespiratory effects of glutamate agonist injections into the dorsolateral pons (DLP) in awake goats. For example, injections of the glutamate receptor excitatory neurotoxin ibotenic acid (IA, Curtis et al., 1979) into the Kölliker–Fuse nucleus (KFN) elicited biphasic ventilatory and cardiovascular responses, whereas there were little or no effects of IA injection within other DRG sites (Bonis et al., 2010b). Chamberlin and Saper (1998) also reported site-specific effects, with glutamate injections into the DLP in rat eliciting site-dependent hyperpnea and hypopnea (Chamberlin et al., 1994). Tract-tracing experiments suggested the rostral pons may play an important role in reflexive respiratory responses to airway stimuli (Chamberlin, 2004). Radulovacki et al. (2003) reported in anesthetized, freely breathing rats that the role of the intertrigeminal region in the control of breathing is to attenuate vagal reflex apneas and dampen respiratory instabilities (Radulovacki et al., 2003). These data taken collectively imply a possible site-specific regulation of breathing and swallowing in rostral pontine areas.
We also previously reported (Bonis et al., 2011) data supporting the concept that neurons in the KFN contribute to coordination of breathing and swallowing in awake goats. These data came from studies in which IA was injected into the KFN, but we recognized that due to possible diffusion of IA to other sites and/or physical damage of the cannula implantation, the observed effects may not have been due entirely to perturbation of the KFN. Accordingly, our objective herein was to gain insight into whether the effects of IA injections into the DLP are site-specific; thus, we expanded the analyses of breathing and spontaneously occurring swallows in animals instrumented with chronically placed cannulae into the lateral (LPBN) and medial (MPBN) parabrachial nuclei for comparison to the KFN. We hypothesized that: (1) IA injections into the DLP would attenuate any resetting of respiratory phase in a site-specific manner and (2) injection effects would increase in magnitude with progressive lesion toward the KFN.
2. Methods
Adult goats were studied as their large size permitted chronic implantation of: (1) stainless steel cannulas into the brainstem and enabled microinjection into target sites during the awake state and (2) muscle electromyographic (EMG) electrodes. Physiologic data were acquired from 11 female adult goats, weighing 46.1 ± 8.6 kg. Six additional goats were used for histological purposes only (44.1 ± 2.9 kg). Goats were housed and studied in an environmental chamber with a fixed ambient temperature and photoperiod and free access to hay and water, except during periods of study. The study was reviewed and approved by the Medical College of Wisconsin Animal Care Committee before studies were initiated.
2.1. Experimental design, protocols, and surgical procedures
Data reported herein were obtained as part of a larger experimental series that determined the effects on breathing of attenuating cholinergic modulation with atropine (Bonis et al., 2010a) and effects of neurotoxic lesions in the dorsolateral pons (Bonis et al., 2010b). Thus, the experimental design and surgical procedures have been reported previously (Bonis et al., 2010b). Briefly, in an initial surgery electrodes were implanted into upper airway and pump muscles for detection of swallows. At least 2 weeks later, in a second surgery cannula were bilaterally implanted just dorsal to the LPBN, MPBN or KFN. These implantations required a single occipital craniotomy created through a posterior midline incision to expose the posterior cerebellum and medulla for visualization of obex. The dorsal medullary surface, obex, and midline were used to determine the dorsoventral, rostrocaudal, and mediolateral planes, respectively. The coordinates for implantations varied with the goat’s size and ranged from 0 to 2 mm ventral, 4 to 5 mm lateral, and 20 to 24 mm rostral to respective reference points. After placement, cannulas were anchored to the surrounding bone using screws and dental acrylic.
After both surgeries, animals were allowed to recover for ≥2 weeks followed by studies to obtain control data which after the second surgery included monitoring of breathing and swallowing over 5½ h while breathing room air. Subsequently, injection protocols consisted of a 30 min control period followed by two unilateral (ipsilateral, then contralateral) microinjections (1 or 10 µl) through the chronically implanted cannulas of IA (50 mM), an irreversible glutamate receptor agonist and excitotoxin (Curtis et al., 1979). Small injection tubes were preloaded with IA and inserted into the cannulas, such that the injection was made at the distal-most aspect of the cannulas without penetrating the tissue. These injections were separated by at least an hour. The total study time was 5½ h, and the 1 and then 10 µl IA injections were separated by 1 week. Seven days after the last IA injection, the animals were euthanized and histological analyses were performed on the brainstem (Bonis et al., 2010b).
2.2. Data analyses
Pulmonary ventilation (ÏI; l/min), breathing frequency (f; breaths/min), tidal volume (VT; l/breath), expiratory (TE; s) and inspiratory (TI; s) times, and diaphragm (DIA), posterior cricoarytenoid (PCA), and at least one other airway muscle activities (mV) were analyzed on a breath-by-breath basis. Muscle activity was recorded via Windaq (Windows Data Acquisition) using the chronically implanted EMG electrodes. The airflow signal was calibrated against a known airflow value. From the calibrated airflow values, VT was calculated. The airway muscle EMG signal provided a signal for detection of swallows. The airflow and EMG signals were converted to a .txt file and input into a custom-designed program that output all parameters on a breath-by-breath basis.
Ventilation was analyzed using the processed airflow signal. Initiation of inspiration (I) was defined as when airflow was greater than 0.01 l/s for at least 0.2 s in duration, and expiration (E) was when airflow returned to zero l/s for at least 0.3 s. The respiratory cycle was defined as an I followed by an E, with the total time (TTot) calculated as the sum of TI and TE. In addition to the respiratory variables reported above, for each breath the presence of one or more swallows and the timing parameters of the swallow(s) relative to the corresponding respiratory cycle were also analyzed.
The activity of the PCA muscle was used for analysis of swallows simply because this was the only airway muscle activity that was recorded in all goats. At least one additional airway muscle was also recorded to verify that the PCA activity reliably identified the swallow. A swallow was considered to have occurred when the PCA signal was greater than 0.01 mV for 0.2–0.5 s in duration. The start of the swallow was defined as the beginning of the signal which met these criteria. To be considered solitary, a single swallow must have occurred within the fourth breath of a six-breath set (Fig. 1). This breath containing the swallow was labeled the n breath, with preceding (n − 1, n − 2 and n − 3) and succeeding (n + 1 and n + 2) breaths labeled accordingly (Fig. 1). The first, second, and sixth breaths served as controls. Swallows associated with coughing, mastication, eructation, etc. were not included in analysis.
Fig. 1.

Depiction of swallow detection and derivation of variables for phase analysis. Swallows were considered solitary if the three breaths preceding (n − 1, n − 2, and n − 3) and the two breaths succeeding (n + 1 and n + 2) the breath of the swallow (n) did not contain swallows. A respiratory cycle was considered to begin with inspiration (I) followed by expiration (E). The n − 3, n − 2, and n + 2 breaths served as control breaths. The vertical line indicates the start of a swallow as evidenced by the raw and moving time average posterior cricoarytenoid (PCA) and thyropharyngeus (TP) muscles signals. Swallows were characterized according to their occurrence within the respiratory cycle as either E, late-E, I, or early-I (Panels A–D, respectively). Old phase (ϕ) was defined as the time from the start of the swallow to the beginning of the n breath. Cophases (θ) were defined as the time from the start of the swallow to the beginning of the preceding breath (θn−1) and three succeeding breaths (θn+1, θn+2, θn+3). Values are normalized as a fraction of control breaths. Panels (A–D) were acquired from the same study in a KFN goat. DIA, diaphragm.
A phase analysis was performed to examine the coordination of breathing and swallowing and investigate whether swallows affected the respiratory rhythm generator. The old phase (ϕ) is the time from the beginning of the n breath to the start of the swallow (Fig. 1). Cophases (θ) were calculated as the time from the beginning of the n − 1 (θn−1), n + 1 (θn+1), n + 2 (θn+2), and n + 3 (θn+3) breaths to the start of the swallow (Fig. 1). ϕ and θ were expressed as a ratio of the average TTot of the control breaths. Solitary swallows were subdivided into 4 subtypes according to their time of occurrence within the respiratory cycle labeled as E, late-E, I or early-I (Fig. 1A–D, respectively). Swallows that occurred during the E phase with a θn+1/ϕ ratio <0.35 were classified as late-E, while those with a θn+1/ϕ ratio >0.35 were classified as E. Similarly, swallows that occurred during the I phase with a θn+1/ϕ ratio >20 were classified as early-I, while those with a θn+1/ϕ ratio <20 were classified as I. We recognize that swallows occurring during the I phase (Fig. 1C and D) created a deglutition apnea, but these temporary pauses in airflow generally did not exceed the 0.3 s duration criteria necessary to trigger a newly defined E phase, and thus used an “I” nomenclature to swallows occurring during the I phase.
2.3. Statistical analysis
To determine if the occurrence of swallows significantly changed over time (5½ h), or condition (control, 1 and 10 µl IA injections), we used a two-way ANOVA with repeated measures and Tukey’s post hoc analysis on the number of swallows per 30 min bin as a percent of the control periods. Similarly, condition effects were determined with the same tests for: (1) the number of swallows occurring within the phases of the respiratory cycle (number of swallows per 10% respiratory phase bin), (2) when swallows occurred with respect to ϕ (number of swallows per 10% ϕ bin), (3) the difference between cophases with respect to ϕ (number of swallows per 10% ϕ bin), and (4) ventilatory variables with respect to ϕ (ventilatory variables expressed as a percent of control breaths per 10% ϕ bin). A threshold for significance was set to P < 0.05.
2.4. Histological analyses
Histological analyses have been reported previously (Bonis et al., 2010b) for these lesioned and control goats. Briefly, after euthanization the head was perfused and fixed with 4% paraformaldehyde in PBS. The brainstem was excised, frozen and serially sectioned (25 µm) from obex to the superior colliculi. Every fourth sections in the rostral pons were stained for Nissl substance and juxtaposed sections were immunostained for detection of muscarinic receptors. The Nissl sections were imaged every 200 µm at 4× magnification utilizing a Nikon Eclipse E400 microscope after Kohler alignment, flat field subtraction and while-balance correction. About 20 image files per slide were then photomerged in Adobe Photoshop, calibrated, and analyzed after importation into MetaMorph Offline v. 7.1.3.0. The KFN, MPBN, and LPBN stain positive for M2 receptors (Mallios et al., 1995); thus, the count region for each was determined using the anti-M2 receptor antibody staining. The boundaries of these regions were then transferred to adjacent Nissl-stained sections for quantification made every 200 µm as previously described.
3. Results
3.1. Histology
The most distal tract created by the implanted cannula was visualized in thinly sectioned pontine tissues and was used to determine the affected anatomic site. The cannulae in 11 goats were located within three pontine respiratory group sub-nuclei including: LPBN (n = 3), MPBN (n = 4), and KFN (n = 4). Histologic observations made obvious a progressively greater lesion resulting from increasing physical (cannulation to a greater depth) and neurotoxic lesions (Fig. 2). For example, cannulation of the MPBN required penetration of the cannulae through the LPBN destroying neurons and dendritic connections within the LPBN, and similarly cannulation of the KFN required penetration of the cannulae through both the LPBN and MPBN, resulting in physical damage in both and additional neurotoxic destruction within the KFN (Fig. 2). As shown in Fig. 2, the area devoid of neurons extended at least 1 mm beyond the cannula medially and laterally. Consistent with this concept, neuronal counts decrease in a manner reflective of the aforementioned phenomenon, with the smallest deficits in the goats implanted in the LPBN and the greatest in the goats implanted in the KFN (Table 1). However, the effects of IA injections were likely on neurons immediately distal to the cannula tip, as injection needles were inserted to a depth that equaled the length of the guide cannulae. Nonetheless, diffusion probably occurred in all directions, and consequently the acute and chronic effects of IA injections are likely cumulative and not solely due to neurons distal to the cannula tract.
Fig. 2.

Histochemical and immunohistochemical staining for Nissl substance and M2 receptors of hemisections from a control goat and a goat with cannula implanted into the KFN. Included also is a schematic to emphasize the rostral–caudal changes in the LPBN, MPBN, and KFN beginning 1 mm caudal to the peak in number of KFN neurons and extending 3 mm rostral. Blue shaded area illustrates the orientation of the nuclei to the superior cerebellar peduncle. The tract of the cannula (gray) extends over 2 mm, and the area devoid of neurons (red) extends over 3 mm in the rostral–caudal direction. Note particularly the halo surrounding and extending beyond the cannula tip indicating tissue damage likely due primarily to ibotenic acid.
Table 1.
Number of neurons and intact area of LPBN, MPBN, and KFN at cannula implantation sites in goats.
| Cannula implantation site | Neurons |
Area |
||||
|---|---|---|---|---|---|---|
| LPBN | MPBN | KFN | LPBN | MPBN | KFN | |
| LPBN | 73 ± 12 | 88 ± 15 | 92 ± 9 | 74 ± 8* | 91 ± 2 | 88 ± 5 |
| MPBN | 61 ± 10 | 62 ± 11 | 80 ± 14 | 63 ± 11* | 75 ± 9* | 96 ± 6 |
| KFN | 52 ± 12* | 51 ± 16* | 51 ± 20* | 62 ± 10* | 63 ± 17* | 59 ± 21* |
Values are the number of neurons and intact area (expressed as a percentage of that in control goats) in the lateral (LPBN) and medial (MPBN) parabrachial nuclei and the Kölliker–Fuse nucleus (KFN) in goats with implanted cannulas. Significant differences between lesioned and control goats are indicated, but because of variations among both lesioned and control goats, several of the deficits in the LPBN, MPBN, and KFN groups were not significantly different from controls.
P< 0.05
3.2. Spontaneous, solitary swallows
The number of breaths and swallows analyzed, stratified by cannula implantation site and study condition (pre-implant control, pre-IA control, and the first 5 h after 1 and 10 µl IA injections) are shown in Table 2. Solitary swallows are further characterized as expiratory (E), inspiratory (I), late-E, or early-I. Of 185,332 total breaths, spontaneous swallows occurred during 37,184 breaths (20% of total breaths). Of these, 17,007 (9% of total breaths) were solitary and met additional selection criteria for inclusion (see Section 3). E, I, late-E, and early-I swallows accounted for 9% (1530), 25% (4202), 62% (10,619), and 4% (656) of solitary swallows, respectively.
Table 2.
Swallow characterization in LPBN, MPBN, and KFN goats.
| Pre-implant control | Pre-IA control | 1 μl IA injection | 10 μl IA injection | Total | |
|---|---|---|---|---|---|
| LPBN | |||||
| Breaths (#) | 5625 | 17,211 | 17,317 | 18,497 | 58,650 |
| Swallows | 848 (15) | 3403 (20) | 3687 (21) | 3169 (17) | 11,107 (19) |
| Solitary sw | 517 (9) | 1805 (10) | 1702 (10) | 1625 (9) | 5649 (10) |
| E | 82 (16) | 240 (13) | 152 (9) | 255 (16) | 729 (13) |
| I | 126 (24) | 479 (27) | 523 (31) | 458 (28) | 1586 (28) |
| Late-E | 309 (60) | 963 (53) | 936 (55) | 845 (52) | 3053 (54) |
| Early-I | 0 (0) | 123 (7) | 91 (5) | 67 (4) | 281 (5) |
| MPBN | |||||
| Breaths (#) | 4435 | 16,884 | 24,396 | 20,216 | 65,931 |
| Swallows | 529 (12) | 4589 (27) | 4910 (20) | 4149 (21) | 14,177 (22) |
| Solitary sw | 301 (7) | 1355 (8) | 1649 (7) | 1758 (9) | 5063 (8) |
| E | 23 (8) | 223 (16) | 231 (14) | 157 (9) | 634 (13) |
| I | 60 (20) | 184 (14) | 357 (21) | 426 (24) | 1027 (20) |
| Late-E | 216 (72) | 888 (66) | 1058 (64) | 1162 (66) | 3324 (66) |
| Early-I | 2 (1) | 60 (4) | 3 (1) | 13 (1) | 78 (2) |
| KFN | |||||
| Breaths (#) | 7816 | 20,392 | 20,615 | 11,928 | 60,751 |
| Swallows | 972 (12) | 3832 (19) | 4315 (21) | 2781 (23) | 11,900 (20) |
| Solitary sw | 590 (8) | 2076 (10) | 2398 (12) | 1231 (10) | 6295 (10) |
| E | 23 (4) | 50 (2) | 23 (1) | 71 (6) | 167 (3) |
| I | 178 (30) | 526 (25) | 697 (29) | 188 (15) | 1589 (25) |
| Late-E | 387 (65) | 1411 (68) | 1554 (65) | 890 (72) | 4242 (67) |
| Early-I | 2 (1) | 89 (4) | 124 (5) | 82 (7) | 297 (5) |
Total and solitary swallow (sw) are expressed as number and (% of total breaths). Expiratory (E), inspiratory (I), late-E and early-I swallows are expressed as number and (% of solitary sw).
3.3. Effects of ibotenic acid on swallowing
The frequency of swallows was not affected by 1 µl IA injection at any DLP site (data not shown). Similarly, 10 µl IA injections into the LPBN and MPBN did not affect the frequency of swallows (Fig. 3). However, injection of 10 µl IA into the KFN increased (P < 0.05) the occurrence of swallows to 175% of control levels within 60–90 min (Fig. 3). Within an hour of this transient increase, the frequency of swallows was attenuated (P < 0.05) relative to the initial increase and it was lower (P > 0.10) than the control value. Injection of 10 µl IA into the KFN, but not LPBN or MPBN, decreased (P < 0.05) the overall number of solitary swallows compared to pre-IA controls and 1 µl IA studies (data not shown). The large standard error bars are indicative of large variation in these responses in all 3 groups, which is implicit with a labile response of swallow frequency.
Fig. 3.

Frequency of swallows over 5 h after injecting 10 µl ibotenic acid (IA) bilaterally into the LPBN (filled diamonds), MPBN (open squares), or KFN (gray triangles) of awake goats. Ipsilateral and contralateral injections (arrows) were made at min 30 and 90, respectively. With injection into the LPBN or MPBN, occurrence of swallows did not significantly differ from control levels. Within 60–90 min after injection into the KFN, the occurrence of swallows was significantly (*P < 0.05) increased to 175% of control levels. Following this increase, the occurrence of swallows was significantly († P < 0.05) attenuated relative to the initial transient increase. Data plotted as a percent of control levels ± SE.
The occurrence of swallows within different phases (E and I) of the respiratory cycle was remarkably consistent between cannula implantation sites (Fig. 4). Peak swallow occurrence was between 80 and 90% of the E phase for all cannula implantation sites, reflective of the majority late-E swallows which were E-terminating. All other swallows were relatively evenly distributed across the respiratory cycle, with a characteristic, yet conserved pattern of occurrence across all cannula implantation sites.
Fig. 4.

Histogram of solitary swallow (normalized to total number of breaths) occurrence with the respiratory cycle during pre-implant and pre-IA injection control (Panels A and B) and 1 and 10 µl IA injection (Panels C and D) protocols. The phase transition between inspiration (I) and expiration (E) (IE trans; denoted by gray bar) is between an old phase (ϕ) of 0.4 and 0.5 in Panel (A), but subsequently widened to between 0.3 and 0.5 in Panels (B–D). Note the conserved magnitude of swallow occurrence across all experimental conditions and cannula implantation sites, as well as the peak occurrence of I swallows at ϕ = 0.2 and E swallows at ϕ = 1 (Panels A–D). Open symbols indicate I swallows, while closed symbols indicate E swallows. ϕ values greater than 1 are possible because some respiratory cycles are longer than the total respiratory time (TTot) of control breaths, some presumably due to the effects of swallowing on respiratory timing parameters (see Fig. 8).
3.4. Respiratory rhythm: a phase analysis
A phase analysis was performed to determine whether swallows affected the respiratory rhythm and subsequently whether injection of IA within different DLP subnuclei altered these effects in a site-dependent manner. To complement the previous results highlighting the timing of swallows with the E or I phases (Table 2), Fig. 4 underscores the timing of swallows within the respiratory cycle (TTot) relative to control breaths (n − 3, n − 2, n + 2). This allows calibration for the effects of the swallow on respiratory rhythm, irrespective of when the swallow occurred during E or I. Fig. 4 depicts the relationship of I or E swallows vs. old phase (ϕ), or the representative TTot. During pre-implant controls, the transition from I to E swallows occurred within a ϕ of 0.4 and 0.5, but the IE transitions broadened to a ϕ of 0.3–0.5 post-cannula implantation and after the 1 and 10 µl IA injections (Fig. 4). The normalized swallow occurrence was conserved across all experimental conditions at all cannula implantation sites (Fig. 4). The peak occurrence of I (ϕ = 0.2) and E (ϕ = 1) swallows was similarly well conserved (Fig. 4).
Fig. 5A illustrates an idealized scenario – a series of parallel lines with a slope of −1 and an amplitude difference of θ = 1 – in which swallows do not affect the respiratory rhythm in the breaths preceding or succeeding the swallow. Deviation above or below these parallel lines indicate a phase advance or delay, respectively, and the resetting of respiratory rhythm. During the pre-implant control study of a KFN goat as shown in Fig. 5B, the θn−1 series (y = −1.0282x − 0.9966; gray data points) closely approximates the idealized θn−1 series in Fig. 5A (y = −1x − 1), establishing that there was no respiratory phase resetting prior to swallows, or ‘anticipation’ of the occurrence of a swallow. In contrast, when including all swallows for the θn+1 series in Fig. 5B, then many data points deviated from that in Fig. 5A, suggesting phase resetting in the breath following swallows. Furthermore, this phase resetting continued through the θn+2 and θn+3 series, suggesting the phase resetting in the θn+1 series was not a transient rhythmic perturbation, but rather a persistent and complete resetting of the respiratory rhythm following a swallow.
Fig. 5.

Depiction of the phase analysis of an idealized example (Panel A) and data during a pre-implant control study in a KFN goat (Panel B). In Panel (A), the idealized plot of θn versus ϕ depicts swallows having no effect on respiratory rhythm indicated by the respective points organizing into series of parallel lines with a slope of −1 and an amplitude difference where θ = 1. Thus, respiratory rhythm was not reset. In Panel (B), when considering effect of all swallow during the control study, the pattern of n + 1 breaths deviated from that in Panel (A) and persisted through subsequent breaths (n + 2 and n + 3). Thus, in this case the swallows appeared to reset respiratory rhythm, with amplitude changes where θ < 1 indicating phase advances and θ > 1 indicating phase delays. The linear regression for θn−1is y = −1.0282x − 0.9966, confirming no anticipation of the phase shift prior to swallows.
However, the relationship among ϕ and θn+1 (and other subsequent breaths) appears to be non-linear, with the data apparently ‘clustering’ in different groups. To gain further insight into this relationship, we separated the swallows based on their occurrence within the respiratory cycle. After this separation, it became apparent that any phase resetting by early-I and late-E swallows had a predictable and consistent effect on e. Early-I swallows largely did not affect the respiratory rhythm, while late-E swallows with a ϕ < 1 caused phase advancing (below idealized line), at a ϕ = 1 did not affect phase, and at a ϕ > 1 caused phase delay (above idealized line; Fig. 6). In contrast, when considering only I and E swallows, a linear regression analysis for the data from LPBN, MPBN, and KFN goats before cannula implantation suggested minimal resetting (Fig. 6). Respective θn+1 slopes for LPBN, MPBN, and KFN animals were near the idealized slope of −1 at −0.7657, −0.9457, and −0.6551 respectively and all y-intercepts were near the ideal of 1 (Fig. 6). This is in contrast to our own previously reported (Bonis et al., 2011, Fig. 9) findings indicating near complete resetting of respiratory rhythm after implanting the cannulas but before 10 µl IA injection into the KFN.
Fig. 6.

The phase analysis for n + 1 breaths during pre-implant control studies in LPBN (Panel A), MPBN (Panel B), and KFN (Panel C) goats. The swallows are categorized as expiration (E; orange), inspiration (I; blue), late-E (purple), and early-I (gray). A trendline is fitted to I and E swallows. The linear regressions for Panels (A–C) are as follows: (A) y = −0.7657x + 1.0003, (B) y = −0.9457x + 1.0992, and (C) y = −0.6551x + 0.9811. The slope of the θn+1 trendlines is similar across cannula implantation sites and near the idealized θn+1 series (y = −1x + 1), indicating minimal phase resetting under intact control, unoperated conditions. Early-I and late-E swallows phase shift in predictable and consistent manners, and were thus not analyzed in this fashion.
Fig. 9.

Physical and chemical destruction altered the effect of swallows on breathing in a site-dependent manner. In the n + 1 breath inspiratory time (TI; Panel A) was decreased (*P < 0.05) in LPBN goats with cannula implantation and KFN goats with 1 µl ibotenic acid (IA) injection. In the n breath expiratory time (TE ; Panel B) was decreased (*P < 0.05) in LPBN animals with 1 µl IA injection and KFN goats with all subsequent conditions. Also in the n breath, TE was decreased († P < 0.05) in a site-dependent manner with 1 µl IA injection in KFN versus MPBN goats. In the n + 1 breath tidal volume (VT; Panel C) was decreased (*P < 0.05) in LPBN and KFN animals with 1 µl IA injections. There was no anticipation or perpetuation of the effects of swallows on breathing, as the n − 1 and n + 2 breaths did not deviate from baseline. The four conditions are (1) pre-implant controls, (2) pre-IA controls, (3) 1 µl IA injections, and (4) 10 µl IA injections. Asterisks denote significant (P < 0.05) conditional differences between bracketed values. The dagger denotes a significant (P < 0.05) implantation site difference between bracketed values.
Plotted in Fig. 7 are the changes in the θn+1 slope relative to pre-implant control for post-implant control and 1 and 10 µl IA injections. There was no phase resetting after I and E swallows in LPBN goats, as indicated by the unchanging θn+1 slope over all studies (Fig. 7). However, as indicated by the increased (less negative) θn+1 slopes relative to pre-implant control levels, there was marked phase resetting after I and E swallows in MPBN and KFN goats after the implants and 1 µl IA injections (Fig. 7). The progression from the absence of phase resetting in LPBN goats to early, marked phase resetting in KFN goats physiologically parallels the progressive physical and chemical lesions, respectively. Note that all goats are at or near pre-implant controls levels following 10 µl IA injections (Fig. 7). These findings suggest that the effects of I and E swallows on respiratory rhythm changes from no resetting at baseline to near-complete resetting, and back to little or no resetting. Furthermore these data suggest plasticity in the site-specific effects of swallows on respiratory rhythm after progressive lesions in the DLP.
Fig. 7.
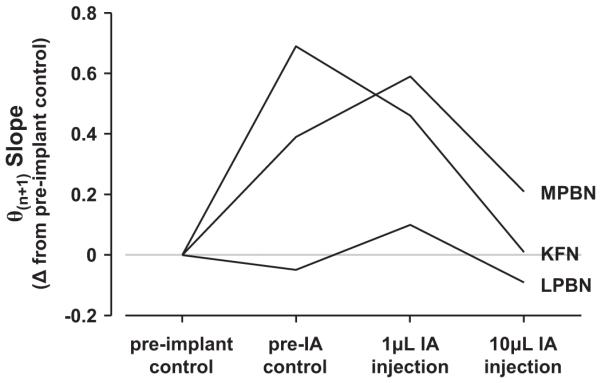
The θn+1trendline slopes (as changed from pre-implant controls) across experimental conditions for all cannula implantation sites. In LPBN goats, the slope remained unchanged from pre-implant control levels, indicating the absence of phase resetting. In MPBN and KFN animals, the slope increased (became less negative) from pre-implant control levels, indicating marked phase resetting.
3.5. Effects of swallows on breathing
Fig. 8 illustrates the effects of swallows on TI, TE, and VT with respect to ϕ after the 10 µl IA injections. Swallows had no significant effects in the n − 1 and n + 2 breaths regardless of ϕ, condition, or implantation site (data not shown), demonstrating a lack of any anticipatory or perpetuated effect on breathing. The majority of effects were found in I swallows (ϕ ≈ 0–40%); in post-implant control studies and after IA injections, where TI was decreased in n and n + 1 breaths of LPBN and KFN goats, TE was decreased in n breaths of all goats, and VT was decreased in n and n + 1 breaths of all animals (P < 0.05; Fig. 8). For all conditions and cannula implantation sites, swallows occurring early in E (ϕ ≈ 40–90%) decreased TE in the n breath, and decreased TI and VT in the n + 1 breath (P < 0.05; Fig. 8). Additionally, swallows occurring late in E (ϕ > 110%) increased TE in the n breath (P < 0.05; Fig. 8).
Fig. 8.

In LPBN and KFN goats, physical and chemical destruction significantly (P < 0.05) decreased inspiratory time (TI) in the n and n + 1 breaths of I swallows (ϕ ≈ 0–40%). In all 3 goat groups, physical and chemical destruction significantly (P < 0.05) decreased expiratory time (TE) and tidal volume (VT) in the n and n + 1 breaths of I swallows (ϕ ≈ 0–40%). For all conditions and cannula implantation sites, early-E swallows (ϕ ≈ 40–90%) decreased TE in the n breath, and decreased TI and VT in the n + 1 breath. Additionally, late-E swallows (ϕ > 110%) increased TE in the n breath. Asterisks denote significant (P < 0.05) differences of respective pre-implant controls from pre-IA control, 1 µl IA injection and/or 10 µl IA injection values by two-way RM ANOVA.
To better characterize the conditional and site-specific effects of swallows on breathing, ventilatory data were plotted without respect to ϕ (Fig. 9). There were no significant effects in the n − 1 and n + 2 breaths, eliminating an anticipatory or a perpetuated effect of swallowing on breathing. Inspiratory time (TI; Fig. 9A) was decreased (P < 0.05) in the n + 1 breath of LPBN goats with cannula implantation and KFN goats with 1 µl IA injection. TE (Fig. 9B) was decreased (P < 0.05) in the n breath of LPBN goats with 1 µl IA injection and KFN goats with all subsequent conditions. TE was decreased (†P < 0.05) in the n breath in a site-dependent manner with 1 µl IA injection in KFN versus MPBN goats (Fig. 9B). Tidal volume (VT; Fig. 9C) was decreased (*P < 0.05) in the n + 1 breath of LPBN and KFN goats with 1 µl IA injections.
4. Discussion
The results of this retrospective analysis support the concept that in awake goats the DLP contributes to the coordination of breathing and swallowing in a site-specific manner, with the effects of perturbations in the KFN greater than at the LPBN and MPBN. Our first hypothesis regarding attenuation of phase resetting was not validated, and in fact proved opposite of that previously reported (Bonis et al., 2011). Herein we report that physical (cannula implantation) and chemical (IA injections) perturbations in the MPBN and KFN resulted in respiratory phase resetting following inspiratory and expiratory swallows, which did not occur with swallows before cannula implantation. Our second hypothesis regarding site-specificity was validated in that progressive physical and chemical lesions produced marked respiratory phase resetting in KFN goats and to a lesser extent in MPBN goats and no phase resetting in LPBN goats.
4.1. Limitations
The retrospective nature of this study is a potential limitation, as it was not designed specifically to examine the coordination of swallows and breathing. However, after now expanding the analyses, the design has proven near ideal to test our present hypotheses. Our previous report (Bonis et al., 2011) was limited by lack of pre-implant control data, which has been addressed and provides significant insight to the present findings. The inability to unequivocally define the anatomic regions affected by IA injections has been previously reported (Bonis et al., 2010b) and is a limitation of this study. A second potential limitation of the study is the relatively small (3–4) number of goats in each group. On the other hand, complete and comprehensive data sets over four different conditions on the same 3 or 4 goats (11 in total) has provided a unique perspective on the role of these pontine sites in coordination of breathing and swallowing. The progressive physiologic effects – most pronounced in KFN goats – observed in parallel to equivalent physical and chemical lesioning provides convincing evidence for the role of a primary site (KFN) and a lesser contribution by the MPBN.
4.2. Site-specificity of effects of perturbations in the DLP
Breathing frequency and heart rate increase within the first 30 min of a 10 µl IA injection into the KFN of awake goats, but both decrease significantly thereafter (Bonis et al., 2010b). The temporal pattern of the occurrence of swallows following 10 µl IA injections into the KFN (Fig. 3) closely resembles the pattern of effects on the minute ventilation and heart rate (Bonis et al., 2010b). In contrast, we found no consistent effects of 10 µl IA injections on breathing, heart rate or the number of swallows when injected into the MPBN or LPBN. Thus, it seems reasonable to conclude the KFN serves a distinct role in modulation of physiologic functions relative to other DLP subnuclei (Chamberlin and Saper, 1998; Chamberlin, 2004). It is important to note that the total number of swallows during pre-implant control studies were similar in all groups of goats suggesting that the effects on swallows was specific to IA injection (Table 2).
Are the effects on multiple physiologic functions of IA injections into the KFN coordinated or coincident? Is the primary response to chemical stimulation of the KFN with 10 µl IA the biphasic ventilatory response, and the occurrence of swallows simply follows as a function of breathing, perhaps for the purpose of increased mucociliary clearance concomitant with increased breathing? This possibility seems unlikely as there are other stimuli such as hypercapnia that increase breathing without affecting swallowing occurrence (Feroah et al., 2000). A second possibility is that there are changes in the sensitivity of the swallowing reflex that account for waxing and waning in the number of swallows, and that the pattern of swallow occurrence is parallel but mechanistically controlled independently from the biphasic ventilatory response. A third possibility is that another behavior followed a similar pattern of stimulation followed by inhibition, such as a primary increase in salivation and licking, leading to a biphasic swallowing response secondary to this behavior. Indeed, anecdotally we have observed increases in salivation and licking after 10 µl IA injections into the KFN of some goats which followed a time course that seemed to be associated with the periods when swallowing and breathing were below control. The mechanism(s) that underlie these parallel responses to chemical stimulation of the KFN remain speculative. Whatever the mechanism, the KFN likely coordinates multiple physiologic functions through excitatory and/or inhibitory outputs to neuronal networks generating specific sequences of muscle patterns for breathing and swallowing.
4.3. Resetting of the respiratory rhythm generator by swallows
Resetting is defined as a shift in phase between that predicted by pre-swallow cycles and the actual post-swallow phase (Fig. 5 and Paydarfar et al., 1995). We emphasize that late-E swallows always reset the respiratory phase as they are E-terminating, whereas early-I swallows never reset respiratory rhythm as they are always succeeded by the inspiratory phase. Prior to cannula implantation, non-transition (I or E) swallows did not cause phase resetting in all goats. These data suggests that the neural network coordinating breathing and swallowing was capable of accommodating a swallow without resetting the respiratory rhythm. Co-equivalency between pre-implant θn+1 slopes (for E and I swallows, see Fig. 6) in all goats and the idealized θn+1 slope (y = −1x + 1) supports this conclusion (Fig. 7). However, significant phase resetting occurred after cannula implantation into the MPBN and particularly into the KFN prior to the generation of IA-induced lesions. Evidence of phase resetting persisted following the 1 µl IA injections, but returned toward pre-implant control slopes following 10 µl IA injections in both groups. The progression of change in θn+1 slope from LPBN to MPBN to KFN goats (Fig. 7) occurring in parallel to progressive physical and chemical lesions in these respective subnuclei are suggestive of site-specificity of effects. In other words, the phase resetting resulted from progressive lesioning in a dorsal to ventral manner upon implantation into the LPBN to the MPBN to the KFN followed by additional neurotoxic lesions with increasing volumes of IA. Collectively, these findings demonstrate that physical and chemical destruction within the DLP transiently altered the effect of I and E swallows to reset the respiratory rhythm in a site-dependent manner, with the KFN serving a major role. In addition, the data suggest that the 10 µl IA injection induced sufficient plasticity to eliminate the respiratory phase resetting that occurred after implantation of the cannula.
4.4. The effect of swallows on TI, TE, and VTVT
Under pre-implant conditions, swallows have small effects on TI, TE, and VT of the breath during and the breath after a swallow (Figs. 8 and 9 and Feroah et al., 2002a). However, these effects were increased by the physical lesion of implanting the cannula and by 1 and 10 µl IA injections into the KFN but not after injections into the LPBN and MPBN. In other words, there seems to be flexibility within the control network so that ventilatory variables are normally minimally compromised by a swallow. However, perturbations in the DLP (particularly in the KFN) affect this flexibility or coordination within the control network. Nevertheless, in spite of the perturbation-induced deficits in the control network, the system was still able to accommodate as we rarely observed coughs in these goats and they had no problems during feeding.
4.5. Concepts on coordination of multiple behaviors utilizing common anatomical structures
For years, the central pattern generator for swallowing was considered as a group of neurons dedicated to swallowing (Bianchi et al., 1995; Jean et al., 1996). However, recently a holarchical system has been proposed whereby a distributed neural network is capable of reorganizing to provide motor output for multiple behaviors (Bolser et al., 2006; Davenport et al., 2011; Pitts et al., 2012). Evidence from reduced preparations supports the KFN as a prominent anatomical site contributing to such a system. Indeed, perturbations within the KFN have been shown to modulate not only respiratory rhythms, but also other behaviors such as coughing, swallowing, vomiting, and vocalization (Gestreau et al., 2005; Bolser et al., 2006; Dutschmann and Herbert, 2006). The consequences of disrupted coordination of these behaviors are exemplified by aspiration pneumonia prevalent in Parkinson’s and Alzheimer’s patients leading to death (Kalia, 2003). Autopsies on these patients have revealed cytoskeletal anomalies (tauopathy) in the LPBN, MPBN, and KFN that conforms with the neurofibrillary tangles and neuropil treads observed in the cortex of these patients (Rüb et al., 2001, 2002). Moreover, the tau protein which normally functions in assembly of microtubules in neurons, is hyperphosphorylated in transgenic mice with a mutation in the tau gene and these mice develop tauopathy in the KFN that is correlated with a progressive upper airway dysfunction including inadequate laryngeal closure during swallowing (Dutschmann et al., 2010). These data provide evidence of how the pathology in the KFN of Parkinson’s and Alzheimer’s patients leads to aspiration pneumonia.
Our findings are consistent with the recent concepts on coordination of breathing and swallowing and provide strong evidence regarding the unique contributions among the rostral pontine sites of the KFN to this coordination. Finally, we emphasize our data in this and previous studies (Bonis et al., 2011; Feroah et al., 2002a, 2002b) were obtained during wakefulness; thus, concerns regarding the relevance of data obtained from reduced preparations should be mitigated.
4.6. Conclusions
Several conclusions are warranted by the data presented herein. First, KFN neurons affect neural networks determining the frequency of multiple physiologic functions. Second, in intact goats, except for late expiratory swallows, the coordination of breathing and swallowing is accomplished without a swallow resetting the respiratory rhythm or greatly compromising TI, TE, or VT. Third, perturbation of the KFN and to a lesser extent the MPBN, reduce the capability of the neural network to accommodate a swallow without affecting respiratory variables. Fourth, the pontine site or mechanism that affects frequency of breathing, swallowing, and heart rate is separate from the site affecting respiratory phase resetting. This conclusion is based on the finding that the frequency of these functions was affected only by IA injections into the KFN but phase resetting was affected by physical and chemical lesions in the KFN and MPBN. Fifth, the mechanism and/or site of phase resetting are distinct from the effects on TI, TE, and VT. This conclusion is based on the finding that the latter effects were exacerbated by progressive lesioning while the effect on phase resetting peaked with the physical lesion and then returned to normal with IA injections. Finally, all the above provide evidence regarding site-specific effects of perturbations within the DLP.
Acknowledgement
This study was supported by the Department of Veterans Affairs and National Heart, Lung, and Blood Institute Grants HL-25739 and HL-007852.
References
- Bianchi AL, Denavit-Saiboe M, Champagnat J. Central control of breathing in mammals: neuronal circuitry, membrane properties and neurotransmitters. Physiological Reviews. 1995 Jan 1;75:1–45. doi: 10.1152/physrev.1995.75.1.1. [DOI] [PubMed] [Google Scholar]
- Bolser DC, Poliacek I, Jakus J, Fuller DD, Davenport PW. Neurogenesis of cough, other airway defensive behaviors and breathing: a holarchical system? Respiratory Physiology and Neurobiology. 2006 Jul 3;152:255–265. doi: 10.1016/j.resp.2006.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonis JM, Neumueller SE, Krause KL, Kiner T, Smith A, Marshall BD, Qian B, Pan LG, Forster HV. A role for the Kölliker–Fuse nucleus in cholinergic modulation of breathing at night during wakefulness and NREM sleep. Journal of Applied Physiology. 2010a Jul 1;109:159–170. doi: 10.1152/japplphysiol.00933.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonis JM, Neumueller SE, Krause KL, Kiner T, Smith A, Marshall BD, Qian B, Pan LG, Forster HV. Site-specific effects on respiratory rhythm and pattern of ibotenic acid injections in the pontine respiratory group of goats. Journal of Applied Physiology. 2010b Jul 1;109:171–188. doi: 10.1152/japplphysiol.00934.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonis JM, Neumueller SE, Marshall BD, Krause KL, Qian B, Pan LG, Hodges MR, Forster HV. The effects of lesions in the dorsolateral pons on the coordination of swallowing and breathing in awake goats. Respiratory Physiology and Neurobiology. 2011 Feb 2;175:272–282. doi: 10.1016/j.resp.2010.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chamberlin NL. Functional organization of the parabrachial complex and intertrigeminal region in the control of breathing. Respiratory Physiology and Neurobiology. 2004 Nov 2–3;143:115–125. doi: 10.1016/j.resp.2004.03.015. [DOI] [PubMed] [Google Scholar]
- Chamberlin NL, Saper CB. A brainstem network medicating apneic reflexes in the rat. Journal of Neuroscience. 1998 Aug 15;18:6048–6056. doi: 10.1523/JNEUROSCI.18-15-06048.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Curtis DR, Lodge D, McLennan H. The excitation and depression of spinal neurons by ibotenic acid. Journal of Physiology. 1979 Jun;291:19–28. doi: 10.1113/jphysiol.1979.sp012796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davenport PW, Bolser DC, Morris KF. Swallow remodeling of respiratory neural networks. Head and Neck. 2011 Oct;33:S8–S13. doi: 10.1002/hed.21845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dutschmann M, Herbert H. The Kölliker–Fuse nucleus gates the postinspiratory phase of the respiratory cycle to control inspiratory off-switch and upper airway resistance in rat. European Journal of Neuroscience. 2006 Aug 4;24:1071–1084. doi: 10.1111/j.1460-9568.2006.04981.x. [DOI] [PubMed] [Google Scholar]
- Dutschmann M, Menuet C, Stettner GM, Gestreau C, Borghgraef P, Devijver H, Gielis L, Hilaire G, Van Leuven F. Upper airway dysfunction of Tau-P301L mice correlates with tauopathy in midbrain and ponto-medullary brainstem nuclei. Journal of Neuroscience. 2010;30:1810–1821. doi: 10.1523/JNEUROSCI.5261-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feroah TR, Forster HV, Pan LG, Rice T. Reciprocal activation of hypopharyngeal muscles and their effect on upper airway area. Journal of Applied Physiology. 2000 Feb 2;88:611–626. doi: 10.1152/jappl.2000.88.2.611. [DOI] [PubMed] [Google Scholar]
- Feroah TR, Forster HV, Fuentes CG, Lang IM, Beste D, Martino P, Pan L, Rice T. Effects of spontaneous swallows on breathing in awake goats. Journal of Applied Physiology. 2002a;92(5):1923–1935. doi: 10.1152/japplphysiol.01079.2000. [DOI] [PubMed] [Google Scholar]
- Feroah TR, Forster HV, Fuentes CG, Wenninger J, Martino P, Hodges M, Pan L, Rice T. Contributions from rostral medullary nuclei to coordination of swallowing and breathing in awake goats. Journal of Applied Physiology. 2002b;93(2):581–591. doi: 10.1152/japplphysiol.01268.2001. [DOI] [PubMed] [Google Scholar]
- Gestreau C, Dutschmann M, Obled S, Bianchi AL. Activation of XII motoneurons and premotor neurons during various oropharyngeal behaviors. Respiratory Physiology and Neurobiology. 2005 Jul 2–3;147:159–176. doi: 10.1016/j.resp.2005.03.015. [DOI] [PubMed] [Google Scholar]
- Jean A, Car A, Kessler JP. Neural Control of Respiratory Muscles. CRC; New York: 1996. Brainstem organization of swallowing and its interaction with respiration. [Google Scholar]
- Kalia M. Dysphagia and spiration pneurmonia in patients with Alzheimer’s disease. Metabolism: Clinical and Experimental. 2003 Oct 10;52(Suppl. 2):36–38. doi: 10.1016/s0026-0495(03)00300-7. [DOI] [PubMed] [Google Scholar]
- Mallios V, Lydic R, Baghdoyan H. Muscarinic receptor subtypes are differentially distributed across brain stem respiratory nuclei. American Journal of Physiology. 1995 Jun 6;268:L941–L949. doi: 10.1152/ajplung.1995.268.6.L941. Pt 1. [DOI] [PubMed] [Google Scholar]
- Pitts T, Morris K, Lindsey B, Davenport P, Poliacek I, Bolser D. Coordination of cough and swallow in vivo and in silico. Experimental Physiology. 2012 Apr 4;97:469–473. doi: 10.1113/expphysiol.2011.063362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paydarfar D, Gilbert RJ, Poppel CS, Nassab PF. Respiratory phase resetting and airflow changes induced by swallowing in humans. Journal of Physiology. 1995;483:273–288. doi: 10.1113/jphysiol.1995.sp020584. Pt 1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prasse JE, Kikano GE. An overview of pediatric dysphagia. Clinical Pediatrics. 2009;48:247–251. doi: 10.1177/0009922808327323. [DOI] [PubMed] [Google Scholar]
- Radulovacki M, Pavlovic S, Saponjic J, Carley DW. Intertrigeminal region attenuates reflex apnea and stabilizes respiratory pattern in rats. Brain Research. 2003 Jun 1–2;975:66–72. doi: 10.1016/s0006-8993(03)02587-3. [DOI] [PubMed] [Google Scholar]
- Rüb U, Del Tredici K, Schultz C, Thai DR, Braak E, Braak H. The autonomic higher order processing nuclei of the lower brain stem are among the early targets of the Alzheimer’s disease-related cytoskeletal pathology. Acta Neuropathologica. 2001;101:555–564. doi: 10.1007/s004010000320. [DOI] [PubMed] [Google Scholar]
- Rüb U, Del Tredici K, Schultz C, de Vos RAI, Jansen ENH, Arai K, Braak H. Progressive supranuclear palsy: neuronal and glial cytoskeletal pathology in the higher order processing autonomic nuclei of the lower brainstem. Neuropathology and Applied Neurobiology. 2002;28:12–22. doi: 10.1046/j.0305-1846.2001.00374.x. [DOI] [PubMed] [Google Scholar]
- Shaker R, Li Q, Ren J, Townsend WF, Dodds WJ, Martin BJ, Kern MK, Rynders A. Coordination of deglutition and phases of respiration: effect of aging, tachypnea, bolus volume, and chronic obstructive pulmonary disease. American Journal of Physiology. 1992 Nov 5;263:G750–G755. doi: 10.1152/ajpgi.1992.263.5.G750. Pt. 1. [DOI] [PubMed] [Google Scholar]