

Abstract
Studies of implicit perceptual-motor sequence learning have often shown learning to be inflexibly tied to the training conditions during learning. Since sequence learning is seen as a model task of skill acquisition, limits on the ability to transfer knowledge from the training context to a performance context indicates important constraints on skill learning approaches. Lack of transfer across contexts has been demonstrated by showing that when task elements are changed following training, this leads to a disruption in performance. These results have typically been taken as suggesting that the sequence knowledge relies on integrated representations across task elements (Abrahamse, Jiménez, Verwey, & Clegg, Psychon Bull Rev 17:603–623, 2010a). Using a relatively new sequence learning task, serial interception sequence learning, three experiments are reported that quantify this magnitude of performance disruption after selectively manipulating individual aspects of motor performance or perceptual information. In Experiment 1, selective disruption of the timing or order of sequential actions was examined using a novel response manipulandum that allowed for separate analysis of these two motor response components. In Experiments 2 and 3, transfer was examined after selective disruption of perceptual information that left the motor response sequence intact. All three experiments provided quantifiable estimates of partial transfer to novel contexts that suggest some level of information integration across task elements. However, the ability to identify quantifiable levels of successful transfer indicates that integration is not all-or-none and that measurement sensitivity is a key in understanding sequence knowledge representations.
Introduction
The acquisition of skilled performance depends critically on repetitive practice that hones execution of learned sequences of actions. Implicit learning contributes importantly to this process and can be seen in the frequent absence of awareness of the exact knowledge learned via repetition. However, for practice-based training to be effective, the improvements gained must transfer from training conditions to the potentially novel performance context. Because implicit learning is frequently observed to be inflexible (Dienes & Berry, 1997; Reber, Knowlton, & Squire, 1996), there is a risk that learning may become bound to the training context (e.g., Shea & Wright, 1995) and pose a challenge for obtaining transfer to the performance context.
In the laboratory, the degree to which implicit learning is inflexible and context-bound can be measured using traditional implicit learning paradigms with training followed by transfer tests in which specific elements of the task are changed. If less knowledge is expressed on the transfer test than in training, then we can conclude that the learning acquired during training could not be accessed or applied fully to the transfer context. This approach can be used to assess transfer as either succeeding (full transfer of knowledge) or failing (absence of any transfer) but could also be used to attempt to quantify amount of transfer as a percentage of the original learning if our measures of learning are sufficiently precise.
Perceptual-motor sequence learning tasks are productive paradigms for characterizing transfer across contexts because they have motor and perceptual task elements that can be independently manipulated. These tasks serve as a model of acquiring fluid performance of action sequences, a key piece of complex skill learning. A frequently used paradigm for studying the acquisition of sequential skill performance and transfer to novel conditions is the serial reaction time task (SRT; Nissen & Bullemer, 1987). More recently, we have introduced the serial interception sequence learning task (SISL; Sanchez, Gobel, & Reber, 2010) which adds movement to the perceptual cues and requires a precisely timed motor response. In the SRT task, cues appear in one of four response locations and participants respond to the appearance of a cue with a corresponding keypress as quickly as possible. Participants are not told that the cues appear in a predictable manner, typically a repeating sequence 10–12 items long, but exhibit knowledge of the sequence by increasingly rapid response times. Sequence-specific learning is demonstrated by a slowing of responses when the order of the cues is changed, showing that some of the learned increase in response speed is specific to the trained sequence and does not transfer to novel, untrained sequences. The SISL task also has four response locations, but cues appear at the top of the screen moving vertically down towards a target zone. Participants attempt to time a motor response to the cue arriving at the target zone (an “interception” response) and performance is measured by accuracy. Sequence-specific learning is likewise measured by a drop in performance when the embedded repeating sequence is surreptitiously changed.
For both tasks, the improvement in performance is often independent of awareness of the repeating sequence, indicating the importance of implicit learning. Healthy participants often acquire some explicit sequence knowledge (e.g., Willingham, Greeley, & Bardone, 1993; Sanchez et al., 2010). However, memory-disordered patients with damage to the medial temporal lobe memory system exhibit intact learning (Nissen & Bullemer, 1987; Reber & Squire, 1994, 1998; Gobel, Parrish, & Reber, 2011), indicating that this explicit knowledge is not necessary in perceptual-motor sequence learning. The SISL task was recently shown to be particularly resistant to influence from explicit knowledge (Sanchez & Reber, 2013) in that even with full explicit knowledge of the repeating sequence provided at training, participants are not able to improve basic task performance. Both tasks provide a method for examining the inflexibility of the implicit learning process due to the fact that most of the task improvement relies on implicit learning of the embedded cue-response sequence.
The fact that implicit learning operates outside of awareness likely contributes to the observations of inflexibility in knowledge use. Because the practiced sequence cannot be brought to mind explicitly, the improvement in performance can only occur if the task parameters automatically trigger the participant’s internal knowledge. The SRT task has been a fertile paradigm for mapping out characteristics of sequence knowledge and conditions in which transfer is or is not seen. In their excellent review of a wide range of studies of implicit learning with the SRT task, Abrahamse et al. (2010b) showed that there is good evidence for learning of multiple response and perceptual sequence components, such as timing, order and stimulus shape. Through extension of the dual-process model (Keele, Ivry, Mayr, Hazeltine, & Hauer, 2003), they argue that integration across covaried elements during training is a key factor in implicit sequence knowledge representations. Likewise, some suggest that these multiple sources of covaried information are crucial for implicit learning to occur (Meier & Cock, 2010).
Integration across elements suggests that disruption to one task element might make the learned knowledge entirely unavailable. In this latter case, transfer should generally be observed to be all-or-none, reflecting cases where information can be brought to bear (full transfer) or cannot (no transfer). In many studies of transfer using the SRT task, transfer has been reported to be of this all-or-none type. Willingham, Wells, Farrell, and Stemwedel (2000) examined performance in a condition that changed just the response locations and found no transfer. Similarly, Schwarb & Schumacher (2010) manipulated S-R associations between the cue and planned response to observe no transfer when this learning was disrupted. Abrahamse and Verwey (2008) found that changing even task-irrelevant features such as the shape of the perceptual cue led to no transfer of sequence knowledge. Jiménez, Vaquero, and Lupiáñez (2006) likewise found that changing perceptual aspects of the response task interfered with the expression of implicit sequence knowledge (but not explicit). These reports showing an absence of transfer to novel conditions are consistent with the idea of inflexible learning and findings in motor learning (Wright & Shea, 1991) showing that changing context impairs performance.
Although the perceptual and response location manipulations appeared to lead to an absence of transfer to the new conditions, two prior studies examined changing the temporal pacing of the cue appearance in the SRT task and found evidence of partial transfer. Both Shin and Ivry (2002) and O’Reilly, McCarthy, Capizzi, and Nobre (2008) found that changing just the inter-cue timing resulted in some transfer of sequence knowledge, suggesting that the timing and response order were represented independently and that response order was more important to accurate responding. However, when we previously examined this question using the SISL task (Gobel, Sanchez, & Reber, 2011), disrupting either source of information led to an apparent absence of transfer again. Even with functional neuroimaging (Gobel et al., 2011a) we did not observe evidence of partial transfer of either source of information on transfer tests or in changes to evoked neural activity. These findings may indicate a discrepancy between the paradigms with the precisely timed response required during SISL performance forcing an integration of the two sources of information during learning. Alternately, it could be that embedding multiple types of sequence information (timing, order) into a single motor response integrates these sources of information during performance.
Here we present three experiments that quantify the degree of transfer in a novel context when one element of a trained sequence representation is altered. In Experiment 1, we replicated the timing-transfer paradigm with SISL but used a new manipulandum that allowed for separating the action order and timing information elements of the response. Separating the order and timing elements of the response made it possible to observe partial (but weak) transfer after selectively manipulating timing information, suggesting that some loss of performance during transfer is due to integration of information sources. We follow this with two additional experiments that examine transfer in contexts with novel perceptual changes, where partial transfer is again found. This suggests that the SISL task is well-suited to quantitative estimation of the magnitude of transfer across conditions. We have previously shown that the measurement of sequence knowledge in SISL is sufficiently sensitive to identify learning in individual participants (Sanchez et al., 2010) and to quantify the learning rate as a function of sequence repetitions (Sanchez & Reber, 2012). Here, across three experiments, transfer conditions were examined in which information about the trained sequence was partially disrupted. In each case, performance on the transfer test was considered as a fraction of the size of the original learning effect to provide an estimate of the amount of knowledge successfully brought to the transfer test.
Experiment 1
Method
Participants
Thirty-five right-handed adults participated in the study (21 female, Mage = 21.8). Nine were Northwestern University undergraduates who received course credit for participation, and 26 were healthy adults recruited by flyer from the Northwestern University community who received $10/h for participation.
Materials
Devices
The guitar-shaped manipulandum was borrowed from music rhythm video games and provided a natural, effective method for separating the order (left hand) and timing (right hand) responses across hands. The guitar-shaped body (RedOctane) was connected via Bluetooth to the computer with a Nintendo® Wii Remote. Responses were recorded into MATLAB© (Version 7.9.0) (2009) (The Mathworks Inc., 2009) using the WiiLab (Brindza, Szweda, Liao, Jiang, & Striegel, 2009) toolbox package.
The serial interception sequence learning (SISL) task
All experiments were written in MATLAB© (The Mathworks Inc., 2009) using the Psychophysics Toolbox extensions (Brainard, 1997). Participants observed circular cues scrolling vertically down a monitor in one of four horizontal locations towards corresponding yellow target rings located near the bottom of the screen. Participants were instructed to intercept the cues when they overlapped a target ring by making a bimanual response with a guitar-shaped manipulandum. Interception responses were made by pressing and holding down the corresponding key with the left hand and subsequently strumming with the right hand (see Fig. 1). The keys were color-coded and were laid out so that the finger placement on the manipulandum corresponded to the horizontal locations on the screen when the palm of the left hand was facing the participant (position/key color/finger: left/green/index; left-middle/red/middle; right-middle/yellow/ring; right/blue/pinky). A correct, coordinated response required the corresponding button to be held down with the left hand while the strum bar was triggered with the right hand. If the strum was outside of the target time window or the incorrect button was held down (or multiple, or no buttons) during the strum, the trial was scored as incorrect. The target time window was set as a given distance around the target ring location; half a cue-length before or after the optimal target location for the class-credit participants, and three-quarters a cue-length before or after for the paid participants. If a coordinated response was correct, the cue disappeared. A performance meter and numerical score provided participants with constant feedback about the overall level of performance.
Fig. 1.

Serial interception sequence learning task with guitar-shaped manipulandum. Participants time a bimanually-coordinated response to intercept circular cues as they scroll through a corresponding target ring. The two-part bimanual response consists of first pressing the corresponding button on the ‘neck’ of the manipulandum with the left hand, followed by an accurately-timed ‘strum’ response by depressing a bar with the right hand. The four target rings correspond to color-coded buttons, mapped (from left to right) to the index, middle, ring, and pinky fingers of the left hand. The button (order) can be pressed any time prior to the strum response, but must be held during the strum. The strum response (timing) was required to be made when a cue was within a given vertical distance from the target ring
During SISL training, the speed of the moving cues was adaptively adjusted based on performance to keep accuracy at around 70 % correct. These adjustments are made to prevent ceiling effects in performance due to general task learning. Subtly different initial speed and adjustment parameters were used for the 9 class-credit participants and 21 paid participants but in both cases the effect was to keep performance at a constant level where a change in performance would eventually be detectable when the sequence was changed. The moving cues initially traversed the screen in 1.25 or 2.0 s and performance was assessed every 12 or 30 trials to determine if speed adjustment was needed. Adjustments were made in small increments (1.6 or 2.5 %) to avoid disrupting performance. Speed was increased slightly when performance was over 70–75 % (a double increase was made at >85 % for the class-credit group whose performance was assessed only every 30 trials) and decreased slightly when performance was <50–70 % correct. Although there were no performance differences between the two groups (on either overall speed or percent correct on the repeating sequence trials), potential differences between the groups were assessed in all data analyses to ensure that the minor variations did not affect any of the reported results.
Sequences were constructed so that the order of responses followed second-order conditional (SOC) structure (see Reed & Johnson, 1994). SOC structure restricts cues from repeating (e.g. 1-1) and prevents paired cues (e.g. 1-3) from appearing more than once per sequence, making a trigram (e.g. 1-3-2) the smallest statistically predictable structure. The sequence timing structure embedded along with cue order followed a specific timing pattern of short and long inter-stimulus intervals (ISIs). Each timing pattern had equal numbers (6) of short and long ISIs, and no more than two of the same ISI lengths would appear consecutively (e.g. 4S2S3L1S4L1L2S4L3S2-S1L3L; S = short ISI, L = long ISI). The short and long ISIs were initially set as 300 and 600 ms, respectively (500/1,000 ms for paid participants), and adjusted with the cue velocity (e.g. faster velocities led to shorter ISIs). This ensured that faster velocities led to more motor responses per second, and also allowed the inter-cue distance on screen to remain stable across velocity adjustments.
Procedure
To familiarize themselves with the task and manipulandum, all participants completed a short demonstration of the SISL task, which included 24 randomly ordered cues. Participants were assigned a randomly selected 12-item repeating sequence of timing and order for training. Trained sequence order was selected from a pool of 256 unique 12-item SOC order sequences. The order and timing used in the trained sequence was not used for any other novel or foil sequences during training or test. The training portion of the SISL task contained six blocks of 40 sequence presentations (480 cues), for 240 sequence presentations total (2,880 cues). Each block consisted of 32 repetitions of the training sequence of timing and order, and 8 presentations of novel non-repeating sequences. Therefore, participants received a total of 192 training sequence repetitions. The blocks were constructed such that for every five sequence presentations, four were repetitions of the training sequence and one was a novel non-repeating sequence. Novel sequences during training never repeated, and were not used as foils during the implicit or explicit knowledge post-training tests. All sequences (trained, novel, or test foils) followed SOC-structure. In between blocks, participants were offered a thirty-second break that could be bypassed by pressing the space bar.
To assess the expression of sequence knowledge when trained order and timing were both maintained, and order and timing were independently maintained, test consisted of four different sequence conditions. The practiced order practiced timing (OPTP) condition retained both the practiced order and timing, while the practiced order novel timing (OPTN) condition selectively changed the ISI timing sequence and the novel order practiced timing (ONTP) condition selectively changed the cue order. A novel order novel timing (ONTN) condition served as the baseline performance condition with completely novel components of both order and timing. The performance advantage for the trained sequence of order and timing was assessed by comparing percentage correct performance on the trained condition (OPTP) to performance on the completely novel condition (ONTN). Performance transfer of independent order or timing information was quantified by assessing performance on the OPTN and ONTP conditions compared to the completely novel condition (ONTN), respectively.
Test consisted of 80 sequence presentations (960 cues), divided into two 40-presentation blocks. The test blocks followed directly after training with no indication that they were different from the preceding training blocks. The test blocks were structured such that every five sequence presentations represented one of the four test conditions. This allowed for the test conditions to be pseudo-randomly intermixed and counter-balanced in order to avoid any order or fatigue effects. Thus, each condition was presented for a total of 20 sequence presentations.
Upon completion of the SISL test, participants were informed that a repeating sequence was present in the task they had just completed. All participants then completed both a recognition test and a cue-order recall test to assess explicit knowledge of the trained sequence. For the recognition test, participants observed and performed two sequence repetitions of the SISL task with their trained sequence and four completely novel order and timing sequences. After each sequence participants were asked to rate whether or not the sequence they had just performed was the repeating sequence from the training trials. Participants rated their confidence on a scale from −10 (absolutely not the sequence) to 10 (absolutely was the sequence).
After the recognition test, participants completed an explicit recall task in which they saw only the yellow target rings on the screen and were instructed to attempt to generate the trained sequence using the same button-strum response from the SISL task. The recall test ended after a participant entered 24 responses. Recall performance was measured by identifying the longest matching subsequence between the participant’s order response and the trained sequence order. To assess baseline recall performance, the generated sequence was also compared to the remaining 201 novel SOC sequences (of 256, 55 had already been used for novel training sequences and tests) and the average matching subsequence to these novel foil sequences was calculated.
Results
Two participants did not understand the SISL task instructions (strummed constantly, as opposed to a single strum response per cue) and were excluded from the analysis. Data from three additional participants were excluded for excessively poor performance during the SISL test (under 15 % correct during a 60-trial sub-block of any of the four conditions). The data reported are from 30 total participants (18 female, Mage = 22.1 years), 9 credit and 21 paid. To measure any potential effects of the method differences between the credit and paid participants, the between-subjects term of ‘method group’ was added to all key ANOVAs examining SISL performance.
Coordinated performance
Sequence-specific learning during training was measured by the difference in performance (percent correct) during the sequence repetitions and the interspersed non-repeating segments (20 % of trials). A mixed ANOVA of training block (one through six) by method group (credit, paid) revealed a significant linear trend in sequence-specific performance, F(1,29) = 31.30, p <0.001, , suggesting that knowledge expression increased throughout training (Fig. 2a). Neither the main effect of method group (F <1) nor the interaction (p = 0.13) reached significance. Non-sequence specific learning, assessed as the cue velocity (time-to-target) at which participants were capable of performing the SISL task, also increased in a linear trend across training, F(1,29) = 53.89, p <0.001. By the onset of the SISL test, cues reached the target 1.16 s (SE = 0.05) after first appearing on screen, and this did not differ across groups, t <1. Although cue velocity is affected by learning of the repeating sequence, it serves as the best measure of general task performance as participants become familiar with the task.
Fig. 2.
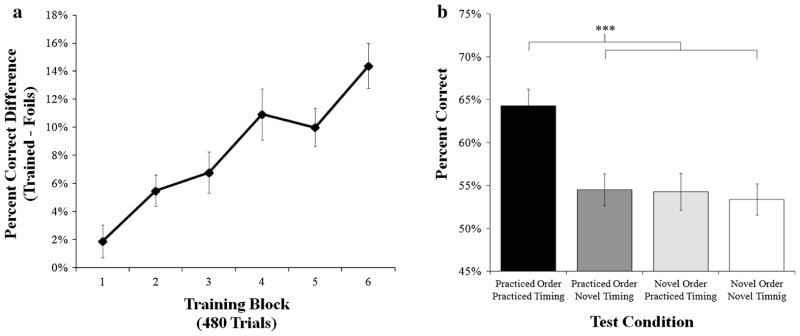
Sequence-specific learning curve and SISL test performance with coordinated bimanual responses in Experiment 1. a The learning curve was calculated as the percent correct difference between the trained sequence trials and the non-repeating novel segment trials for each block of training, which leads to a sequence-specific performance advantage that increased over training. b The SISL test results showed that participants had a sequence-specific performance advantage at test for the trained sequence of timing and order. Sequence-specific performance benefits at test were not present if either the order or timing component of the sequence was changed
The performance effect of changing the practiced sequence order, timing, or both at test was assessed by a mixed 2 × 2 × 2 ANOVA of order (practiced, novel), timing (practiced, novel), and the method group (credit, paid). There were main effects of order, F(1,28) = 21.04, p <0.001, , and timing, F(1,28) = 16.45, p <0.001, , and a significant interaction between order and timing, F(1,28) = 25.37, p <0.001, , reflecting that performance was best when the practiced order and timing were retained (see Fig. 2b). As in Gobel et al. (2011b), performance during the practiced sequence of order and timing was significantly better than the other three conditions, ts(29) >5.78, ps <0.001, which were all performed at similarly disrupted levels, F <1. Using the completely novel sequence of order and timing (ONTN) as a baseline performance measure, the sequence-specific performance benefit when order or timing was selectively retained was negligible (OPTN: M = 1.13 %, SE = 1.17 %; ONTP: M = 0.89 %, SE = 1.47 %), compared to when both sequence characteristics were kept the same (OPTP, M = 10.86 %, SE = 1.88 %), suggesting a lack of partial transfer if order or timing was selectively maintained. There was no main effect and no significant interactions between method groups, all ps >0.1.
Separate performance
Using the novel manipulandum, the coordinated response for each trial can be broken down into two separate elements: the left-hand order response and the right-hand timing response. The bimanual response was scored with respect to the left hand to assess partial transfer of order knowledge (was the correct key pressed) or just the right hand to assess partial transfer of timing knowledge (was the timing correct).
Left-hand/order
Order performance was assessed independently of the right hand (timing strum) by comparing the order of button-press responses against the order of cues appearing on the screen. The response characteristics of the task promote early button presses, in order to prepare the timed strum response, so this scoring tended to include correct key responses made before the cue intercepted the target zone. However, this method did penalize extra LH order responses not paired with a RH strum response that were ignored in the standard bimanual response scoring. For instance, a D keypress not followed by a strum was ignored in the bimanual scoring, but considered as a response in the LH order performance scoring.
As can be seen in Fig. 3a, the overall percent correct was higher with this scoring method (as expected because only the button press response had to be correct). A 2 × 2 × 2 mixed ANOVA of order (practiced, novel), timing (practiced, novel), and method group (credit, paid) revealed main effects of order, F(1,28) = 22.41, p <0.001, , and timing, F(1,28) = 4.69, p <0.05, , although the order-timing interaction was not significant, F <1. There was neither a main effect nor significant interaction of method group, all ps >0.1.
Fig. 3.
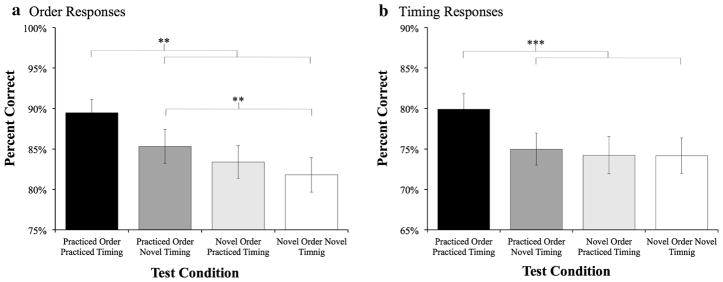
SISL test performance separately assessed by order and timing for Experiment 1. Left hand order responses (button presses) were examined separately of right hand timing responses (strums). a Order performance in the practiced order practiced timing condition showed the largest performance benefit, with a smaller, yet significant, performance benefit for the practiced order novel timing condition, suggesting partial transfer or sequence order knowledge. No performance benefit was found in the novel order practiced timing condition. b When right-hand timing responses (strum) were examined separately, participants expressed a sequence-specific performance advantage for only the practiced order practiced timing condition, with no signs of transfer of knowledge to the other conditions
The performance benefits of retaining both trained order and timing, just order, and just timing as compared to completely novel order and timing were examined to assess the a priori hypotheses that retaining both sequence components leads to the highest sequence-specific benefit while retaining order will show some degree of performance transfer. Similar to coordinated responding, order responses were significantly better in the OPTP condition, ts >3.26, ps <0.01, compared to the other three conditions, and featured the highest sequence-specific benefit (M = 7.67 %, SE = 1.59 %) compared to the completely novel (ONTN) condition. However, in contrast to the coordinated performance analysis, a sequence-specific performance benefit in the OPTN condition compared to the novel condition was found (M = 3.51 %, SE = 1.15 %), t(29) = 3.05, p <0.01, suggesting successful transfer of sequential order knowledge. No performance transfer of sequential order knowledge was found when timing was selectively retained, as performance during the ONTP condition was only marginally better (M = 1.58 %, SE = 1.59 %) than the novel condition, t <1.
Right-hand/timing
Timing responses were scored as correct if a single, appropriately timed strum was made to the upcoming cue, regardless of the correctness of the left hand (button selection) action. Timing was considered correct if the strum was made while the cue was within. 75 cue-lengths either before or after the target zone. Multiple strums or strums made outside of the necessary timing window were considered incorrect.
As can be seen in Fig. 3b, the overall percent correct was higher as the correct button did not have to be pressed or held down for a strum to be properly timed. A 2 × 2 × 2 mixed ANOVA of order (practiced, novel), timing (practiced, novel), and method group (credit, paid) revealed results very similar to the coordinated performance, with main effects of order, F(1,28) = 7.35, p <0.05, , and timing, F(1,28) = 16.77, p <0.001, , and a significant interaction, F(1,28) = 9.99, p <0.01, . Again, strum responses were significantly better in the OPTP condition, ts >4.21, ps <0.001, compared to the other three conditions, and featured the highest sequence-specific benefit (M = 5.74 %, SE = 1.20 %) compared to baseline. The other three conditions did not differ in performance, F <1, reflecting a lack of performance transfer when sequence timing was selectively retained in the ONTP condition. A main effect of method group, F(1,28) = 7.39, p <0.05, , reflected the higher percent of accurate strums in the paid condition (M = 83.28 %, SE = 2.88 %) compared to credit condition (M = 72.62 %, SE = 2.24 %)—likely due to the minor differences in administration. Importantly, method group did not interact with any other terms, all ps >0.1.
Explicit knowledge
Participants developed some explicit knowledge of their trained sequence, as measured by both recognition and recall. The confidence ratings provided to the trained sequence (M = 6.40, SE = 0.68) were significantly higher than the confidence ratings provided to the foil sequences (M = −2.93, SE = 0.83) during the recognition test, t(29) = 8.70, p <0.001. Likewise, the sequences generated by the participants during the recall test matched the trained sequence (longest matching subsequence; M = 5.27, SE = 0.42) slightly better than novel sequences (M = 4.18, SE = 0.09), t(29) = 2.74, p <0.05.
To assess the potential effect of explicit knowledge on the performance transfer of ordinal sequence knowledge in the OPTN test condition in the left-hand scoring analysis, participants were median-split based on their recognition scores into a high explicit knowledge group and a low explicit knowledge group. The difference in recognition ratings provided to the trained sequence and foil sequences in the low explicit knowledge group (M = 4.60, SE = 0.81), was much lower than in the high explicit knowledge group (M = 14.05, SE = 0.96). The transfer exhibited by the low explicit knowledge group (M = 3.39 %, SE = 1.68 %) was nearly identical to the transfer exhibited by the high explicit knowledge group (M = 3.63 %, SE = 1.64 %), t <1, and recognition was not correlated with transfer, r = −0.02, suggesting that sequence performance transfer was not driven by explicit knowledge.
Discussion
When correct responding requires a coordinated response based on cue timing and sequential order, the sequence-specific performance advantage obtained through practice did not transfer, even partially, to conditions where just order or timing were selectively changed. This result is consistent with the prior report with the SISL task (Gobel et al., 2011b). However, when separately analyzing action and order response elements (enabled by the novel manipulandum) partial transfer was observed such that action selection based on sequence order was enhanced in the OPTN condition when the timing information was selectively changed. This result indicates that separate action sequence learning was occurring independent of timing information. Transfer was not found in the ONTP condition even though the individual timing response characteristics did not change from the trained condition. The partial performance transfer of order (when timing was changed) but not timing (when order was changed) fits with previous SRT work suggesting that timing is merely a component of an action order sequence which is not learned independently (O’Reilly et al., 2008; Shin & Ivry, 2002).
These results suggest that the integration of sequence components is not necessarily all-or-none, such that separating the timing and action components across hands allowed for partial transfer of ordinal knowledge. The lack of measureable increases in trained sequence performance in just the timing-response hand (RH) may suggest that learning of a timing sequence independent of ordinal action responding is not occurring here, and is in line with previous reports (O’Reilly et al., 2008; Shin & Ivry, 2002). Alternately, the lack of transfer may be related to the fact that the timing response is made after the initial order response since the left-hand button press (order) is the first part of a two-part response which precedes an accurately timed right-hand strum. When the order sequence and LH responses were changed in the ONTP condition, this may have blocked the subsequent ability to produce a correctly timed strum even if there had been independent learning of the inter-response timing sequence.
As a side effect to manipulating the timing and order of the motor response, the transfer conditions in Experiment 1 altered the visuo-spatial cue pattern that participants viewed during training. The SISL task presents multiple (3–4) upcoming cues simultaneously and this spatial information may be a perceptual component that is integrated into the learning process and could also affect transfer from training to test. Thus, in Experiment 1, the spatial layout of cues on-screen was altered across each condition. Although a change in perceptual information could not be the only factor leading to a lack of transfer, because the degree of transfer was specific to the sequence element (ordinal transfer only), it is possible that the change in perceptual information was a factor that led to low performance transfer.
To examine the selective impact of changing perceptual information, in Experiment 2, participants trained on the same repeating sequence under one of two perceptual conditions, followed by tests including both the trained and alternate perceptual conditions. For this experiment, the motor response demands were the more traditional forced-choice keypress on a keyboard we have used in previous reports (e.g. Sanchez et al., 2010). Across the two conditions used for training and transfer test, participants were performing an identical precisely timed motor response sequence. Abrahamse and Verwey (2008) found that changing apparently irrelevant perceptual task features disrupted transfer with the SRT task. Here we will assess whether perceptual information is similarly embedded in SISL learning and like Experiment 1, attempt to quantify the degree of transfer observed.
Experiment 2
Method
Participants
Forty-four Northwestern University undergraduates received course credit for participation (24 female, Mage = 18.8, 39 right-handed).
Materials
SISL task
Participants intercepted scrolling cues by making a keypress as they overlapped target rings. A novel SISL task variant was used in which the scrolling cues moved down the screen in a Single column and contained visible color and letter information about the appropriate button to press (Single-column condition; Fig. 4). This modification removed the spatial mapping between cue position and response, making the task more difficult, while also changing the perceptual appearance of the cue sequence so that transfer across perceptual characteristics of the task could be assessed. Participants also performed the SISL task with the traditional perceptual presentation of Experiment 1 (Quad-column condition). For both versions of the task, participants used a standard computer keyboard and responded to color-coded circular cues labeled with the corresponding keyboard response (D, F, J or K). The same repeating sequences can be covertly embedded in both task versions so that the identical motor response sequence can be trained with either perceptual cueing.
Fig. 4.

SISL task in Single- and Quad-column perceptual variants. a In the Single-column version, the cues scroll down the same middle column towards the center ring, and participants must respond with the correct keypress based on the color and letter of the perceptual cue. b In the Quad-column version of the SISL task, the cues scroll down one of four columns towards the four outer rings, corresponding to the four horizontal response locations on the keyboard (D, F, J, K). No cues scroll towards the center ring in the Quad-column condition. In both of these instances, a participant must be responding with the “D” key, and be preparing for the upcoming “K” and “F” responses
Positive and negative feedback were provided by turning the corresponding ring green if the response was correct or red if the response was incorrect. The feedback was provided to both the four outer rings and central ring in both conditions, so that visual feedback was constant across conditions. Response feedback and cue velocity adjustments were based on the correctness of both order and timing. A response provided positive feedback and counted towards an increase in cue velocity if the correct button was pressed while the cue was within half a cue-length either before or after the target location. Incorrect keypresses, keypresses made outside of the timing window, and multiple keypresses made within a single timing window were all considered incorrect responses. Incorrect responses elicited negative feedback, and counted towards a decrease in cue velocity. Performance was assessed every 12 trials and percent correct over 75 % led to an increase in speed of 2.5 % and performance of ≤50 % correct led to a decrease of the speed by 2.5 %. In contrast to Experiment 1, adaptive velocity was extended through training and into the test portion of the SISL task as well. The timing between trials was kept constant (initially 700 ms) so that the Single-column condition would contain no unique spatial information between cues.
Procedure
Experiment 2 was conducted over two sessions, 48 h apart, in order to provide additional training that was necessary for participants to become successful at performing the more difficult Single-column task. Participants first completed short demonstrations of both variations of the SISL task, which each included 24 random cues. Participants were then randomly assigned to either the Quad-column or Single-column training condition, and were assigned a 12-item SOC training sequence. For session 1, participants first completed six training blocks in their assigned condition. Each block had 40 sequence presentations—32 presentations of the training sequence, and 8 presentations of novel, non-repeating sequences. This was immediately followed by a block of 40 novel, non-repeating sequences in the other perceptual condition to allow participants to familiarize themselves with the transfer condition that they would be tested on in the second session. Sequence concatenation was accomplished in a way to prevent any items from repeating (e.g. no 4-4 or 2-2 was allowed at sequence junctures). All non-repeating SOC sequences were randomly chosen and never re-used during the experiment.
At the onset of the second session, participants received two more sequence training blocks in their assigned condition, resulting in a total of 256 repetitions of the trained sequence. Directly following training were SISL tests of both the Single- and Quad-column conditions (test order was counterbalanced across participants). Each test began with 15 novel, non-repeating sequence presentations (to allow for cue velocity adjustments to bring task performance into the targeted range). This was followed by 75 total sequence presentations, consisting of the trained sequence and two novel foils (25 repetitions of each 12-item sequence). Similar to Experiment 1, each test was divided into two blocks (45 sequence presentations), and sequences were presented in five-repetition sub-blocks in order to pseudo-randomly intermix the sequence types. Participants received 60-s self-terminated rest breaks between all training and test blocks to reduce fatigue. After SISL training and tests, participants completed a recognition test and cue-order recall test, as in Experiment 1. These tests were counterbalanced for order effects and were conducted in the participants’ trained SISL condition (Single- or Quad-column).
Results
SISL performance
After study completion, it was found that the colored and labeled cues caused a slight display rendering issue (dropped frames) that produced occasional uncertainty in the position of the moving cues. To accommodate for the imprecise screen draw timing, the response-timing window for the temporally-sensitive SISL task was slightly expanded for performance assessments. Thus, responses were scored as correct as long as the correct keypress was made when the current cue was closer to the target than any other cue. Data from two participants were excluded for excessively poor performance during the SISL test (under 15 % correct during a 60-trial sub-block of any test condition). Three additional participants were excluded due to exceptionally low training performance (2 participants responded to less than half the trials of an entire 480-trial training block, and one participant performed consistently below the 50 % velocity adjustment window). The remaining 39 participants (21 female, Mage = 18.8 years, 34 right-handed) were included in the final data analysis.
During training, sequence-specific performance improvements were calculated as the difference in percentage correct responses for the trained sequence and the novel, non-repeating segments. A 2 × 8 mixed ANOVA (training condition × training block) revealed that the sequence-specific benefit increased in a linear trend across training, F(1,37) = 21.57, p <0.001, . At the end of training, the sequence-specific performance benefit was 8.74 % (SE = 1.48 %) in the Quad-column condition and 5.14 % (SE = 2.20 %) in the Single-column condition, but the interaction did not reach significance, F(7,259) = 1.81, p = 0.09. Correct responding was similar across groups as there was no main effect of training condition, F <1.
Test performance was assessed with a 2 × 2 × 2 mixed ANOVA of training condition (Quad-column, Single-column), sequence type (trained, novel), and test type (same perceptual condition, transfer perceptual condition). As seen in Fig. 5, participants exhibited significant sequence-specific performance benefits at test for the trained sequence, as reflected by the main effect for sequence type, F(1,37) = 56.48, p <0.001, . The sequence-specific benefit at test was similar across training conditions, but was higher during the same test condition (Quad: M = 4.35 %, SE = 0.89 %; Single: M = 5.49 %, SE = 1.36 %) compared to the transfer test condition (Quad: M = 1.85 %, SE = 0.72 %; Single: M = 1.54 %, SE = 1.20 %), evidenced by a significant interaction between sequence type and test type, F(1,37) = 7.46, p <0.01, . Sequence expression was significant for both groups in the same test condition, ts >4.04, ps <0.001. Transfer expression was only significant for the participants in the Quad-column training condition, t(20) = 2.58, p <0.05, although there was a trend in the same direction for the Single-column condition, t(17) = 1.28, p = 0.22. Sequence type did not interact with training condition, nor was there a three-way interaction, Fs <1.
Fig. 5.

Sequence-specific performance advantage during training and test in Experiment 2. Both the Single- and Quad-column training conditions show a linear increase in sequence-specific performance improvements over training, with significantly more sequence-specific performance benefit expressed during test with the trained perceptual condition, compared to the transfer perceptual condition
In regards to general SISL performance at test, there was a main effect of test type, F(1,37) = 9.82, p <0.01, , and a large interaction effect, F(1,37) = 213.64, p <0.001, , reflecting the higher performance during the Quad-column test for participants in both the Quad-column training (M = 92.16 %, SE = 1.61 %) and Single-column training conditions (M = 89.75 %, SE = 1.43 %), as compared to the Single-column test (Quad-column training: M = 73.76 %, SE = 1.51 %; Single-column training: M = 77.85 %, SE = 1.85 %). There was no main effect of training condition, F <1.
Cue velocity, measured as the time-to-target in seconds, decreased across training in both conditions in a linear trend, F(1,37) = 127.81, p <0.001 , but participants were performing the SISL task at a much faster time-to-target (M = 0.73 s, SE = 0.03 s) in the Quad-column condition, compared to the Single-column condition (M = 1.18 s, SE = 0.04 s), t(37) = 8.80, p <0.001. Likewise, the time-to-target during the Quad-column test was much faster (M = 0.85 s, SE = 0.03 s) than during the Single-column test (M = 1.27 s, SE = 0.03 s), t(38) = 14.90, p <0.001, suggesting that the SISL task was performed slower during the Single-column perceptual version. Cue velocity was extremely stable across the tests, as the time-to-target during the first 180-trials of each test (Quad: M = 0.85 s, SE = 0.04 s; Single: M = 1.25 s, SE = 0.03 s) was nearly identical to the time-to-target during the last 180 test trials (Quad: M = 0.85 s, SE = 0.04 s; Single: M = 1.26 s, SE = 0.03 s).
Explicit knowledge
Participants in both conditions exhibited an ability to recognize their training sequence, providing higher confidence ratings to the trained sequence (M = 3.72, SE = 0.93) than to the foil sequences (M = 0.34, SE = 0.59), F(1,37) = 8.19, p <0.01, , on the −10 to 10 scale. There was neither a main effect of training condition nor an interaction effect (both n.s.). The sequences generated by the participants during the recall test had a slightly longer matching sub-sequence to the trained sequence (M = 4.95 items, SE = 0.27) compared to novel foil sequences (M = 4.47 items, SE = 0.04), but the difference did not reach significance F(1,37) = 2.95, p = 0.09, . Both training conditions had similar recall knowledge, as there was neither a main effect of training type nor a significant interaction (Fs <1).
To assess the potential effect of explicit knowledge on the ability to transfer sequence performance across conditions, the two groups were median split based on their recognition scores into a high explicit knowledge group and a low explicit knowledge group. The difference in confidence ratings provided to the trained sequence and foil sequences in the low explicit knowledge group was actually negative (M = −2.45, SE = 0.97), suggesting they provided higher confidence ratings to the foil sequences than to the trained sequence. The high explicit knowledge group, in contrast had a very high difference in confidence ratings between the trained sequence and foil sequences (M = 8.91, SE = 0.99). However, the amount of performance transfer exhibited by the low explicit knowledge group was actually higher (M = 2.33 %, SE = 1.01 %) than the transfer exhibited by the high explicit knowledge group (M = 1.43 %, SE = 0.88 %), t <1, and recognition was not correlated with transfer, r = −0.07, suggesting that sequence performance transfer was not driven by explicit knowledge.
Discussion
In both perceptual conditions, participants exhibited similar sequence-specific performance advantages during the post-training test even though the perceptual presentation of the response cues was different. During the transfer test when participants switched to the untrained perceptual condition, participants exhibited a significant drop in performance in spite of the fact that they were performing an identical motor response sequence. In the transfer condition, SISL performance was still better for the trained sequence than for novel foils, indicating that there was partial transfer of sequence knowledge of approximately 35 % of the magnitude of the original learning. As in Experiment 1, transfer was not observed to be all-or-none. Partial transfer implies that much, but not all, of the learning during the initial training was tied to the method of perceptual presentation. Interpretation of the effect of changing the structure of the perceptual cues in Experiment 2 is potentially complicated by the fact that the Single-column condition disrupted the natural spatial mapping between the cues and motor responses. As a result, this task was more difficult for participants than the usual Quad-column condition, as might be expected (Simon, 1969). Previous SRT work has suggested that changing the spatial compatibility between the stimuli and responses can significantly affect learning (Werheid, Ziessler, Nattkemper, & Yves von Cramon, 2003). However, these effects of spatial-mapping have not been completely consistent across studies and may be affected by explicit knowledge in healthy participants (Abrahamse, Jiménez, Deroost, van den Broek, & Clegg, 2010a; Deroost & Soetens, 2006b; Koch, 2007). However, if the spatial compatibility mapping between the stimuli and response locations is a learned component of the sequence representation, altering this information could have been part of the reduction in performance on the transfer test.
In Experiment 3, perceptual information was manipulated across conditions but the spatial compatibility was maintained between the cue and response locations. In addition, as in Experiment 2, the response sequence to be executed by the participants was identical across conditions. The amount of transfer obtained was again estimated quantitatively to measure how much of the knowledge of the repeating sequence could be applied when only the perceptual cueing structure of the task was changed between training and test.
Experiment 3
Method
Participants
Twenty-eight participants from the Northwestern University community received $15 for 90-min of participation (19 female, Mage = 20.5, 26 right-handed).
Materials
SISL task
Participants intercepted scrolling cues by making a keypress as they overlapped target rings. The cues scrolled down four separate columns towards four horizontally-spaced target rings that spatially corresponded to the four locations on the keyboard. Positive and negative feedback were provided by turning the corresponding ring green if the response was correct or red if the response was incorrect. Response feedback and cue velocity adjustments were based on the correctness of both order and timing. A response provided positive feedback and counted towards an increase in cue velocity if the correct button was pressed within half a short ISI length (initial short ISI, 400 ms) either before or after the cue was optimally-lined up with the target location. If the wrong button was pressed or the correct button was pressed outside of this timing window, then negative feedback was provided and it counted towards a decrease in cue velocity. Performance was assessed every 12 trials throughout training and test, and percent correct over 75 % led to an increase in speed of 5.0 % and performance of 50 % correct or worse decreased the speed by 5.0 %.
The Standard perceptual version of the SISL task (similar to the display in Experiment 1 and the Quad-column condition of Experiment 2) has multiple cues visible on screen at a time (about 3). For this experiment, a new Isolated variant only has a single cue visible on the screen at a time, and the timing between responses is maintained from the Standard version by increasing the overall cue velocity. Thus, the physical distance between cues differs between the two conditions in order to alter how many cues are visible on screen at any given time, but the cue velocity varies between conditions in order to keep the timing between sequence items consistent (see Fig. 6). For instance, the distance between the tops of two cues separated by a short ISI is 130 pixels (two cue lengths), and the initial time it takes the cues to travel across the screen is 1,180 ms. However, in the Isolated version the distance is 465 pixels (7.2 cue lengths) and the initial time it takes the cues to travel across the screen is 330 ms. In both cases, the short ISI was 400 ms and the correct response timing window was kept constant. Thus, the response rate and timing accuracy was constant across both conditions. The presentation, responses and feedback for the Isolated cue variant was otherwise identical to the Standard version of the SISL task.
Fig. 6.
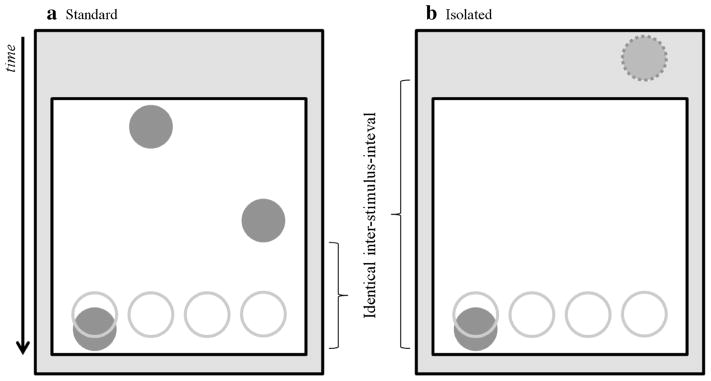
SISL task in Standard and Isolated cue variants. In both versions of the task, the cues scroll down one of four columns towards four rings that are displayed in a spatially-compatible layout that correspond to the response locations on the keyboard (D, F, J, K). In both of the task variants above, only stimuli within the white space are visible to the participant. a In the Standard version of the task, multiple cues are visible on the screen at a time. b In the Isolated cue version of the task, only a single cue is visible at a time. This change is made by increasing the physical distance between the cues while also increasing the velocity the cues are scrolling at, so that the inter-stimulus timing between cues remains the same between conditions. Note that the cue with the dashed-outline in the grey area of the figure is not visible to the participant, but is being used to display how the distance between cues changes between conditions
Procedure
Participants first completed short demonstrations of both variations of the SISL task, which each included 24 random cues. Participants were then randomly assigned to the Standard (multiple-visible cue) condition or the Isolated (single-visible cue) condition for training, and were assigned a 12-item SOC training sequence. SISL training consisted of six blocks, which each featured 32 repetitions of the training sequence and 8 presentations of novel, non-repeating sequences (192 repetitions of the trained sequence, total). Directly after training, participants completed SISL tests of both the Standard and Isolated cue conditions, which were counterbalanced across participants to control for order effects. The SISL tests followed directly after training, and no indication that a test was being administered was provided to the participants. The test procedure for Experiment 3 was identical to Experiment 2. Each test began with 15 presentations of novel, non-repeating sequences to allow for cue velocity adjustments, followed by 75 total sequence presentations, including the trained sequence and two foils (25 presentations of each sequence). The test was divided into two 45-presentation blocks, and structured so that every five repetitions represented performance on one of the three sequences. Participants received 60-s self-terminated rest breaks between all training and test blocks to reduce fatigue. After SISL training and tests, participants completed a recognition test and cue-order recall test, as in Experiments 1 and 2. These tests were counterbalanced for order effects and were conducted in the participants’ trained SISL condition (Standard or Isolated).
Results
SISL Performance
Sequence-specific performance improvements were calculated as the percentage correct difference between the trained sequence and the novel, non-repeating segments across training. A mixed 2 × 6 ANOVA of condition (Standard, Isolated) and training block (1 through 6) revealed that the sequence-specific benefit increased in a linear trend across training, F(1,26) = 21.94, p <0.001, , and that there was also a main effect of training condition, F(1, 26) = 7.64, p <0.05, , and a significant interaction effect, F(5, 130) = 2.32, p <0.05, , suggesting that learning rates differed between training groups (see Fig. 7). To further examine the high sequence-specific performance advantage in the Isolated condition (M = 32.89 %, SE = 6.05 %) compared to the Standard condition (M = 18.09 %, SE = 3.54 %) at the end of training, the performance on the training sequence and novel non-repeating sequences was assessed. The trained sequence performance at the end of training in the Isolated condition (M = 69.83 %, SE = 0.74 %) was similar to the trained sequence performance in the Standard condition (M = 68.51 %, SE = 0.93 %), t(26) = 1.09, p = 0.29. However, the novel non-repeating sequences were performed much worse in the Isolated condition (M = 36.94 %, SE = 5.45 %) compared to the Standard condition (M = 50.42 %, SE = 2.97 %), t(26) = 2.25, p <0.05.
Fig. 7.
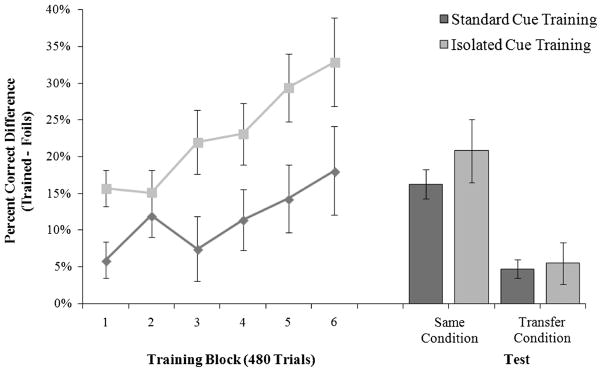
Sequence-specific performance advantage during training and test in Experiment 3. Both the Standard and Isolated cue training conditions show a linear increase in sequence-specific performance improvements over training, with significantly more sequence-specific performance benefit expressed during test with the trained condition, compared to the transfer condition
Sequence performance at test was assessed with a 2 × 2 × 2 mixed ANOVA of training condition (Standard, Isolated), sequence type (trained, novel), and test type (same perceptual condition, transfer perceptual condition). As seen in Fig. 7, the pattern of results is similar to Experiment 2 in that participants exhibited a significant sequence-specific performance advantage at test, evidenced by a main effect of sequence type, F(1,26) = 58.14, p <0.001, . However, a significant interaction between sequence type and test type, F(1,26) = 35.66, p <0.001, , reflects that both training groups exhibited a higher sequence-specific benefit during the condition that matched training (Standard: M = 16.28 %, SE = 2.03; Isolated: M = 20.82 %, SE = 4.30 %), when compared to the transfer test condition (Standard: M = 4.74 %, SE = 1.28; Isolated: M = 5.49 %, SE = 2.82 %). Sequence knowledge expression was robust in the condition that matched training, ts >4.85, ps <0.001, and while transfer was significant for participants in the Standard condition, t(14) = 3.43, p <0.01, the performance transfer in the Isolated condition trended towards significance, t(12) = 1.95, p = 0.08. There was a main effect of test type, F(1,26) = 10.12, p <0.01, , reflecting better general performance in the same condition than in the transfer condition. However, there was no main effect of training condition and it did not interact with any other terms, Fs <1.
Non-sequence specific learning was assessed as the cue velocity set by the adaptive velocity adjustments. Cue velocity has previously been measured as the time-to-target, but because this variable was manipulated across conditions in order to maintain sequential inter-stimulus-interval timing, the velocity measure reported here is the short ISI. A 2 × 6 mixed ANOVA (training condition, training block) revealed that the short ISI decreased in a linear trend across training in both conditions, F(1,26) = 20.49, p <0.001, . There was also a main effect of training condition, F(1,26) = 47.75, p <0.001, , but the interaction did not reach significance, F <1, suggesting that while both groups had increases in general task performance, the overall velocity at which the participants performed the task differed across groups. From the initial short ISI of 400 ms, by the end of training participants in the Isolated condition were performing the task with a short ISI of 345 ms (SE = 16) while participants in the Standard condition had a mean short ISI of 226 ms (SE = 5.8). This difference in velocity was reflected during test across all participants as well, as the Standard version of the SISL task was performed with a much faster short ISI (M = 254 ms, SE = 15), compared to the Isolated version of the SISL task (M = 402 ms, SE = 7.7). Similar to Experiment 2, cue velocity was extremely stable across the tests, as the ISI during the first 180-trials of each test (Standard: M = 236 ms, SE = 11 ms; Isolated: M = 406 ms, SE = 9 ms) was very similar to the ISI during the last 180 test trials (Standard: M = 253 ms, SE = 11 ms; Isolated: M = 407 ms, SE = 12 ms).
Explicit knowledge
A mixed 2 × 2 ANOVA of sequence type (trained, foils) and training condition (Standard, Isolated) on the recognition test revealed a main effect of sequence type, F(1,26) = 38.64, p <0. 001, , suggesting that participants were capable of recognizing their trained sequence. However, a significant interaction effect, F(1,26) = 5.98, p <0.05, , reflects that the difference in confidence ratings provided by the Standard group to the trained sequence (M = 4.13, SE = 1.19) and foil sequences (M = −0.15, SE = 1.11) was not as large as the difference in confidence ratings by the Isolated group (trained: M = 6.46, SE = 1.25; foils: M = −3.35, SE = 1.29), indicating that participants in the Isolated training condition were significantly better at recognizing their trained sequence. The main effect of training condition was not significant, F <1.
A 2 × 2 ANOVA of the recall data showed significant main effects for both sequence type (trained, foils), F(1,26) = 8.54, p <0. 01, , and training condition (Standard, Isolated), F(1,26) = 5.68, p <0.05, and a significant interaction effect, F(1,26) = 4.59, p <0.05, . Participants in the Isolated condition produced more of the trained sequence (M = 6.92 items, SE = 1.00) than the foil sequences (M = 4.41 items, SE = 0.06), t(12) = 2.49, p <0.05, while the participants in the Standard condition recalled a subsequence that matched both the trained sequence (M = 4.67 items, SE = 0.32) and foil sequences (M = 4.28 items, SE = 0.08) at roughly similar levels, t(15) = 1.29, p = 0.22 (chance level of recall).
Similar to the previous experiments, to assess the potential effect of explicit knowledge on the ability to transfer sequence performance across conditions, the two groups were median split based on their recognition scores into a high explicit knowledge group (n = 15) and a low explicit knowledge group (n = 13). The difference in confidence ratings provided to the trained sequence and foil sequences in the low explicit knowledge group was much lower (M = 1.58, SE = 0.67) than participants in the high explicit knowledge group (M = 11.42, SE = 1.37). However, the amount of performance transfer exhibited in the low explicit knowledge group (M = 4.95 %, SE = 2.00 %) was similar to the transfer exhibited by the high explicit knowledge group (M = 5.21 %, SE = 2.14 %), t <1, and there was no correlation between recognition and transfer, r = 0.06, suggesting that sequence performance transfer was not driven by explicit knowledge.
Discussion
Despite consistent spatial compatibility between conditions, when participants had to transfer their learned sequence knowledge to a novel perceptual variant of the SISL task, there was a significant cost in knowledge expression. Importantly, this significant cost was not complete, and participants in both conditions exhibited a similar ~25–30 % partial transfer of sequence knowledge. The overall results are very similar to Experiment 2, such that the cost in sequence-specific performance expression when the perceptual display is changed is much higher than would be expected if perceptual learning of the stimulus display is a separate and equal component of the sequence representation learned in parallel, suggesting there is a degree of integration across components. However, the integration is not absolute because changing the perceptual information during a transfer test did not completely wipe out sequence expression in the SISL task, as it has in other perceptual-motor tasks (e.g. Abrahamse & Verwey, 2008). Again, these results support a model of learning based on integration across the component processes supporting performance during the perceptual-motor sequence learning task, but do not suggest complete integration such that any slight change disallows performance transfer.
General discussion
Across three experiments, the ability to maximally express knowledge acquired during practice was dependent on performing under conditions that matched the precise conditions during training. When aspects of either the motor response or perceptual display were changed, relatively low levels of partial transfer were observed although there was notably still a reliable benefit from training. Theories of sequence learning have debated about what component processes are recruited for sequence learning, from stimulus (Clegg, 2005) and response location (Willingham et al., 2000) processing to a more complex rule about their relation (Schwarb & Schumacher, 2010). The current results suggest that learning is occurring across multiple components and, further, that there is evidence for integration of information across sensory and motor domains. Although the low levels of transfer observed here suggest that learning is specifically and inflexibly tied to the training context, the resulting ability to identify performance transfer suggests that it is possible to partially apply information sources that are maintained from training to test performance contexts.
The timing and order transfer results of Experiment 1 demonstrate the potential complexity of the integration component in sequence learning. During the coordinated responding that required response correctness to be dependent on both order and timing information, no transfer was found in any condition that differed from training. This integration result is similar to prior SISL results where a single keypress was used (Gobel et al., 2011a, b). However, when performance was assessed separately for timing and order across hands, partial transfer of ordinal—but not temporal—information was revealed, replicating similar results with the SRT task (O’Reilly et al., 2008; Shin & Ivry, 2002). These results suggest that the integration of individual sequence components (e.g. timing and order) may be dependent on the response characteristics of how this information is used. Thus, the knowledge representation may differ based on the goal-oriented response characteristics of the task that is used for learning (Abrahamse et al., 2010b).
Experiments 2 and 3 showed that integration extends cross-modally, such that perceptual cue information was partially integrated with the motor response sequence. Although the motor expression of sequence knowledge has been argued in some reports to not depend on perceptual information (Willingham, 1999; Willingham et al., 2000) several other reports have found perceptual information to be a robustly learned component (Deroost & Soetens, 2006a; Remillard, 2003; Song, Howard, & Howard, 2008). Here, by quantifying the magnitude of transfer across conditions where perceptual information is varied we find evidence for both a key role of perceptual structure in learning and also some ability to apply the practiced motor sequence (at 25–35 % efficiency) when the cues were changed. Similar to Experiment 1, these results imply that the integration of sequential elements is complex. Previous SRT work suggests that learning of the perceptual component is sensitive to procedural task characteristics (Deroost & Soetens, 2006a). This claim fits with the results found here in that the SISL task has a robust perceptual component that may recruit perceptual learning for optimal sequence learning and performance.
These quantitative estimates also provide a starting point for developing models of sequence learning in which there is learning within each of the task element components but also a critical integration component. The additional performance benefit provided by the integrated learning leads to the observation that total sequence-specific increase in performance is greater than the sum of the parts of the learning of individual component processes, such as previous reports of the facilitation of ordinal learning with a coordinated temporal sequence (O’Reilly et al., 2008; Shin & Ivry, 2002). Likewise, the utilization of the novel SISL task here demonstrates the necessity of using different types of tasks in order to develop a more robust understanding of sequence learning. The SRT task has been the canonical task for studying sequence learning for years (Nissen & Bullemer, 1987) and has developed quite a legacy. However, as noted in Schwarb and Schumacher (2012), to appropriately understand and model this learning mechanism, it is necessary to examine learning in a range of tasks in order to reconcile disparate results that cannot be explained when only a single task is used.
The findings across the three studies reported here support the idea of the relative inflexibility of implicit learning (Dienes & Berry, 1997) compared with explicit knowledge that can be consciously brought to mind. Inflexibility in knowledge use is typically considered a hallmark of implicit learning (Cleeremans, Destrebecqz, & Boyer, 1999) and has been observed in domains from probabilistic classification (Reber et al., 1996) to priming (Schacter, Dobbins, & Schnyer, 2004). Although concomitant explicit knowledge was found across the experiments, it is unlikely to have had an impact on the rates of knowledge transfer. Explicit knowledge was not found to have an effect on performance or the degree of transfer in any of the experiments, and is not likely to have been capable of being applied in order to support performance given the rapid response characteristics of the SISL task. Additionally, the utilization of explicit knowledge is typically recruited in a top-down fashion when executive control is called upon through conscious task demands or rule changes (e.g. Jiménez et al., 2006) and in the transfer conditions used here, participants were merely asked to intercept falling cues with motor responses—there was no additional complexity or rule change that would have implied or warranted use of explicit knowledge.
Despite a significant drop in sequence knowledge expression when any element of the sequence was changed, the observation of at least partial transfer indicates that sequential learning is not completely bound to the training context, which would pose a problem for training in the real world. Since there is generally a benefit from practice even when practice and performance contexts differ, a complete lack of transfer could have indicated a problem with the paradigms used to capture implicit learning in the laboratory. Our observations of very limited transfer may reflect the fact that participants had a fairly limited amount of practice with the skill learning task compared to the many hours devoted to the development of expertise. For example, changes in the underlying representation over extended practice (e.g., Korman, Raz, Flash, & Karni, 2003) might eventually allow learners to avoid impairments in applying skilled motor knowledge due to differences in the perceptual context of the kind examined here. However, it is also notable that skill acquisition training programs often seek to mimic the performance conditions during practice, seeming to implicitly acknowledge the potential difficulty of transfer. The current findings provide a method for quantitatively estimating the magnitude of transfer effects (and likewise the decrement in performance across contexts) that may serve to guide future research aimed at optimizing training conditions to maximize transfer.
Acknowledgments
This research was funded in part by an American Psychological Association Diversity Program in Neuroscience Fellowship and a National Institutes of Health Training Grant T32 NS047987 (DJS), NSF CNS-1228357 (PJR), and a Northwestern Posner Summer Fellowship and a Northwestern Undergraduate Research Grant (ENY). The authors would like to thank Dr. Konrad Koerding and Ian Stevenson for their assistance with the guitar manipulandum interface.
Contributor Information
Daniel J. Sanchez, Department of Psychology, Northwestern University, 2029 Sheridan Road, Evanston, IL 60208, USA
Eric N. Yarnik, Department of Psychology, Northwestern University, 2029 Sheridan Road, Evanston, IL 60208, USA
Paul J. Reber, Email: preber@northwestern.edu, Department of Psychology, Northwestern University, 2029 Sheridan Road, Evanston, IL 60208, USA
References
- Abrahamse EL, Jiménez L, Deroost N, van den Broek EL, Clegg BA. Controlled response selection benefits explicit, but not implicit sequence learning. In: Abrahamse EL, editor. Serial action and perception. Enschede: University of Twente; 2010a. pp. 131–154. [Google Scholar]
- Abrahamse EL, Jiménez L, Verwey WB, Clegg BA. Representing serial action and perception. Psychonomic Bulletin and Review. 2010b;17(5):603–623. doi: 10.3758/PBR.17.5.603. [DOI] [PubMed] [Google Scholar]
- Abrahamse EL, Verwey WB. Context dependent learning in the serial RT task. Psychological Research. 2008;72(4):397–404. doi: 10.1007/s00426-007-0123-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brainard DH. The psychophysics toolbox. Spatial Vision. 1997;10(4):433–436. [PubMed] [Google Scholar]
- Brindza J, Szweda J, Liao Q, Jiang Y, Striegel A. WiiLab: bringing together the Nintendo Wiimote and MATLAB. Frontiers in education conference, 2009. FIE’09. 39th IEEE; IEEE; 2009. pp. 1–6. [Google Scholar]
- Cleeremans A, Destrebecqz A, Boyer M. Implicit learning: news from the front. Trends in Cognitive Sciences. 1998;2(10):406–416. doi: 10.1016/s1364-6613(98)01232-7. [DOI] [PubMed] [Google Scholar]
- Clegg BA. Stimulus-specific sequence representation in serial reaction time tasks. The Quarterly Journal of Experimental Psychology Section A. 2005;58(6):1087–1101. doi: 10.1080/02724980443000485. [DOI] [PubMed] [Google Scholar]
- Deroost N, Soetens E. Perceptual or motor learning in SRT tasks with complex sequence structures. Psychological Research. 2006a;70(2):88–102. doi: 10.1007/s00426-004-0196-3. [DOI] [PubMed] [Google Scholar]
- Deroost N, Soetens E. The role of response selection in sequence learning. Quarterly Journal of Experimental Psychology. 2006b;59(3):449–456. doi: 10.1080/17470210500462684. [DOI] [PubMed] [Google Scholar]
- Dienes Z, Berry D. Implicit learning: below the subjective threshold. Psychonomic Bulletin and Review. 1997;4(1):3–23. [Google Scholar]
- Gobel EW, Parrish TB, Reber PJ. Neural correlates of skill acquisition: decreased cortical activity during a serial interception sequence learning task. NeuroImage. 2011a;58(4):1150–1157. doi: 10.1016/j.neuroimage.2011.06.090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gobel EW, Sanchez DJ, Reber PJ. Integration of temporal and ordinal information during serial interception sequence learning. Journal of Experimental Psychology Learning, Memory, and Cognition. 2011b;37(4):994–1000. doi: 10.1037/a0022959. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiménez L, Vaquero JMM, Lupiáñez J. Qualitative differences between implicit and explicit sequence learning. Journal of Experimental Psychology Learning, Memory, and Cognition. 2006;32(3):475–490. doi: 10.1037/0278-7393.32.3.475. [DOI] [PubMed] [Google Scholar]
- Keele SW, Ivry R, Mayr U, Hazeltine E, Heuer H. The cognitive and neural architecture of sequence representation. Psychological Review. 2003;110(2):316–339. doi: 10.1037/0033-295x.110.2.316. [DOI] [PubMed] [Google Scholar]
- Koch I. Anticipatory response control in motor sequence learning: evidence from stimulus-response compatibility. Human Movement Science. 2007;26(2):257–274. doi: 10.1016/j.humov.2007.01.004. [DOI] [PubMed] [Google Scholar]
- Korman M, Raz N, Flash T, Karni A. Multiple shifts in the representation of a motor sequence during the acquisition of skilled performance. Proceedings of the National Academy of Sciences. 2003;100(21):12492–12497. doi: 10.1073/pnas.2035019100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MATLAB (Version 7.9.0) Natick: The MathWorks Inc; 2009. [Google Scholar]
- Meier B, Cock J. Are correlated streams of information necessary for implicit sequence learning? Acta Psychologica. 2010;133(1):17–27. doi: 10.1016/j.actpsy.2009.08.001. [DOI] [PubMed] [Google Scholar]
- Nissen MJ, Bullemer P. Attentional requirements of learning: evidence from performance measures. Cognitive Psychology. 1987;19(1):1–32. [Google Scholar]
- O’Reilly JX, McCarthy KJ, Capizzi M, Nobre AC. Acquisition of the temporal and ordinal structure of movement sequences in incidental learning. Journal of Neurophysiology. 2008;99(5):2731–2735. doi: 10.1152/jn.01141.2007. [DOI] [PubMed] [Google Scholar]
- Reber PJ, Knowlton BJ, Squire LR. Dissociable properties of memory systems: differences in the flexibility of declarative and nondeclarative knowledge. Behavioral Neuroscience. 1996;110(5):861–871. doi: 10.1037//0735-7044.110.5.861. [DOI] [PubMed] [Google Scholar]
- Reber PJ, Squire LR. Parallel brain systems for learning with and without awareness. Learning and Memory. 1994;1(4):217–229. [PubMed] [Google Scholar]
- Reber PJ, Squire LR. Encapsulation of implicit and explicit memory in sequence learning. Journal of Cognitive Neuroscience. 1998;10(2):248–263. doi: 10.1162/089892998562681. [DOI] [PubMed] [Google Scholar]
- Reed J, Johnson P. Assessing implicit learning with indirect tests: determining what is learned about sequence structure. Journal of Experimental Psychology Learning, Memory, and Cognition. 1994;20(3):585–594. [Google Scholar]
- Remillard G. Pure perceptual-based sequence learning. Journal of Experimental Psychology Learning, Memory, and Cognition. 2003;29(4):581–597. doi: 10.1037/0278-7393.29.4.581. [DOI] [PubMed] [Google Scholar]
- Sanchez DJ, Gobel EW, Reber PJ. Performing the unexplainable: implicit task performance reveals individually reliable sequence learning without explicit knowledge. Psychonomic Bulletin and Review. 2010;17(6):790–796. doi: 10.3758/PBR.17.6.790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanchez DJ, Reber PJ. Operating characteristics of the implicit learning system supporting serial interception sequence learning. Journal of Experimental Psychology: Human Perception and Performance. 2012;38(2):439–452. doi: 10.1037/a0026347. [DOI] [PubMed] [Google Scholar]
- Sanchez DJ, Reber PJ. Explicit pre-training instruction does not improve implicit perceptual-motor sequence learning. Cognition. 2013;126(3):341–351. doi: 10.1016/j.cognition.2012.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schacter DL, Dobbins IG, Schnyer DM. Specificity of priming: a cognitive neuroscience perspective. Nature Reviews Neuroscience. 2004;5(11):853–862. doi: 10.1038/nrn1534. [DOI] [PubMed] [Google Scholar]
- Schwarb H, Schumacher E. Implicit sequence learning is represented by stimulus-response rules. Memory and Cognition. 2010;38(6):677–678. doi: 10.3758/MC.38.6.677. [DOI] [PubMed] [Google Scholar]
- Schwarb H, Schumacher EH. Generalized lessons about sequence learning from the study of the serial reaction time task. Advances in Cognitive Psychology. 2012;8(2):165–178. doi: 10.2478/v10053-008-0113-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shea CH, Wright DL. Contextual dependencies: influence on response latency. Memory. 1995;3(1):81–95. doi: 10.1080/09658219508251498. [DOI] [PubMed] [Google Scholar]
- Shin JC, Ivry RB. Concurrent learning of temporal and spatial sequences. Journal of Experimental Psychology-Learning Memory and Cognition. 2002;28(3):445–457. doi: 10.1037//0278-7393.28.3.445. [DOI] [PubMed] [Google Scholar]
- Simon JR. Reactions toward source of stimulation. Journal of Experimental Psychology. 1969;81(1):174–176. doi: 10.1037/h0027448. [DOI] [PubMed] [Google Scholar]
- Song S, Howard JH, Howard DV. Perceptual sequence learning in a serial reaction time task. Experimental Brain Research. 2008;189(2):145–158. doi: 10.1007/s00221-008-1411-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Werheid K, Ziessler M, Nattkemper D, Yves von Cramon D. Sequence learning in Parkinson’s disease: the effect of spatial stimulus–response compatibility. Brain and Cognition. 2003;52(2):239–249. doi: 10.1016/s0278-2626(03)00076-9. [DOI] [PubMed] [Google Scholar]
- Willingham DB. Implicit motor sequence learning is not purely perceptual. Memory and Cognition. 1999;27(3):561–572. doi: 10.3758/bf03211549. [DOI] [PubMed] [Google Scholar]
- Willingham DB, Greeley T, Bardone AM. Dissociation in a serial response time task using a recognition measure: comment on Perruchet and Amorim (1992) Journal of Experimental Psychology Learning, Memory, and Cognition. 1993;19(6):1424–1430. [Google Scholar]
- Willingham DB, Wells LA, Farrell JM, Stemwedel ME. Implicit motor sequence learning is represented in response locations. Memory and Cognition. 2000;28(3):366–375. doi: 10.3758/bf03198552. [DOI] [PubMed] [Google Scholar]
- Wright DL, Shea CH. Contextual dependencies in motor skills. Memory and Cognition. 1991;19(4):361–370. doi: 10.3758/bf03197140. [DOI] [PubMed] [Google Scholar]