

Abstract
Leishmaniasis is considered by the World Health Organization as one of the infectious parasitic diseases endemic of great relevance and a global public health problem. Pentavalent antimonials used for treatment of this disease are limited and new phytochemicals emerge as an alternative to existing treatments, due to the low toxicity and cost reduction. Usnic acid is uniquely found in lichens and is especially abundant in genera such as Alectoria, Cladonia, Evernia, Lecanora, Ramalina, and Usnea. Usnic acid has been shown to exhibit antiviral, antiprotozoal, antiproliferative, anti-inflammatory, and analgesic activity. The aim of this study was to evaluate the antileishmanial activity of usnic acid on Leishmania infantum chagasi promastigotes and the occurrence of drug-induced ultrastructural damage in the parasite. Usnic acid was effective against the promastigote forms (IC50 = 18.30 ± 2.00 µg/mL). Structural and ultrastructural aspects of parasite were analyzed. Morphological alterations were observed as blebs in cell membrane and shapes given off, increasing the number of cytoplasmic vacuoles, and cellular and mitochondrial swelling, with loss of cell polarity. We concluded that the usnic acid presented antileishmanial activity against promastigote forms of Leishmania infantum chagasi and structural and ultrastructural analysis reinforces its cytotoxicity. Further, in vitro studies are warranted to further evaluate this potential.
1. Introduction
Leishmaniasis is one of the most neglected diseases that remain a public health problem worldwide, affecting approximately 12 million people in more than 98 countries, with more than 350 million people at risk and when not treated, death is expected to occur after a period of four months to one year [1, 2].
Leishmaniasis is endemic in areas of tropics and subtropics, including southern Europe, Asia, Africa, and the Americas [3–5].
Leishmania infantum chagasi, a trypanosomatid parasite, is an etiological agent of visceral leishmaniasis (VL) in the American continent and it is now admitted to be the same species causing visceral leishmaniasis in Europe and certain parts of Africa [6].
Pentavalent antimonials such as sodium stibogluconate and meglumine antimoniate have been used in the treatment of all forms of leishmaniasis for more than half a century; although the mechanism of action of pentavalent antimonial is not fully understood, it is generally accepted that the active form of the metal is the reduced form Sb (III), which can lead to serious limitations due to high toxicity and resistance to drugs [2, 7]. In addition, a major problem in antimonial chemotherapy is the emergence of clinical resistance against pentavalent antimonial drugs that has reached epidemic proportions in parts of India [8–10]. Newer drugs such as miltefosine and amphotericin B have shown promising results, but they also cause some side effects that may limit their use. In this context, the discovery of new active and promising compounds with antileishmanial potential remains essential for control and prevention of leishmaniasis [11, 12].
Several natural products with antileishmanial activity have been recently reported, including naphthoquinones, lignans, neolignans, alkaloids, flavanol, and terpenoids [13, 14], but none has reached clinical use.
Lichens synthesize over eight hundred types of metabolites [15] and depsides, depsidones, dibenzofurans, xanthenes, anthraquinones, and usnic acids are amongst the more extensively studied lichen metabolites [16]. The search for new phytochemicals with antiparasitic action emerges as an alternative to existing treatments, due to the low toxicity and cost reduction.
Some studies showed that usnic acid, secondary metabolite of the lichen Cladonia substellata Vainio, was initially used in the treatment of pulmonary tuberculosis [17], and there are data regarding its biological activities as antibiotic [18], antiproliferative [19], analgesic and antipyretic [20], anti-inflammatory [21], antiviral [22, 23], antifungal [24], against the parasite Trypanosoma cruzi [25], and an immunologic modulator [26]. In addition, its mutagenic and cytotoxic activities have been determined against normal and malignant human cells lines [27–30].
Taking into account the side effects and the resistance that pathogenic protozoan parasites develop against drugs currently used in the treatment of leishmaniasis, more attention should be given to extracts and biologically active compounds isolated from plant species commonly used in herbal medicine. Thus, in this work, we have analyzed the in vitro effect of usnic acid on growth and ultrastructure of Leishmania infantum chagasi promastigote forms.
2. Materials and Methods
2.1. Extraction of Usnic Acid
Usnic acid was isolated from the crude extract of the lichen Cladonia substellata Vainio, and was obtained from Dr. Nicácio Henrique da Silva at Department of Biochemistry, Federal University of Pernambuco. Fractionation and purification of this compound were performed as previously described [31]. A stock solution was prepared at 10 mg/mL in DMSO and stored at 4°C until use.
2.2. Parasites Culture
The strain of Leishmania infantum chagasi (MHOM/BR2000/Merivaldo2) was kindly provided by Dr. Osvaldo Pompílio de Melo Neto at Department of Microbiology of Research Center Aggeu Magalhães, FIOCRUZ. The promastigotes were routinely grown in Liver Infusion Tryptose medium (LIT) at 26°C, supplemented with 10% heat-inactivated fetal bovine serum (FBS) (LGC Biotechnology), 0.1% penicillin and streptomycin, and 0.2% hemin (Sigma).
2.3. In Vitro Antileishmanial Activity
The antileishmanial activity was assessed by the colorimetric method MTT [3-{4.5-dimethylthiazol-2-yl}-2.5-diphenyl-tetrazolium] (SIGMA) that is based on the conversion of the tetrazolium salt into the colored formazan product, the concentration of which can be determined spectrophotometrically. The promastigotes were seeded (2 × 106 cells/mL) with LIT medium in 96-well microplates and incubated with 50 μL of different concentrations of usnic acid (100 to 0.195 μg/mL) and amphotericin B (positive control). LIT medium (50 μL) (negative control) was also added. After 72 h of incubation with usnic acid and amphotericin B 25 μL of MTT solution was added (5 mg/mL). After 3 h of incubation at 37°C, MTT solution was aspirated and 100 μL DMSO was added for the solubilization of the formazan crystals. After solubilization, the absorbance was determined in a spectrophotometer at 585 nm wavelength. The results were expressed as percentage of relative viability of cells to the negative control group [32]. All the experiments were performed in triplicate.
2.4. Scanning Electron Microscopy
To evaluate parasite ultrastructural alterations by scanning electron microscopy, Leishmania infantum chagasi promastigotes were grown for 72 h as described in LIT-DMSO or the same medium containing 25 μg/mL usnic acid; they were subsequently collected by centrifugation at 1500 g, washed twice in 0.1 M Phosphate buffer (pH 7.2), and fixed in a solution containing 2.5% glutaraldehyde, 4% paraformaldehyde, and 0.1 M T phosphate buffer. After washing twice in the same buffer, the parasites were adhered to glass slides previously coated with 0.1% poly-I-lysine for 30 min at 37°C. Subsequently, the slides were washed twice with 0.1 M phosphate buffer, postfixed in solution of OsO4 for 1 h at room temperature, and washed twice again with 0.1 M phosphate buffer. All samples were dehydrated gradually increasing the ethanol concentrations (30–100%) and were critical point dried using CO2, mounted on metal stubs, and coated with gold (5–30 nm) for observation in a scanning electron microscope (JEOL T-200).
2.5. Transmission Electron Microscopy
Control and treated culture promastigotes with 25 μg/mL were harvested, at room temperature by centrifugation at 1500 g, washed twice in PBS, and fixed for 2 h at 4°C in a solution containing 2.5% glutaraldehyde, 4% paraformaldehyde, and 0.1 M cacodylate buffer, pH 7.2. After washing in this similar buffer, cells were postfixed for 1 h in a solution containing 1% OsO4, 0.8% potassium ferricyanide, and 5 mM CaCl2 in 0.1 M cacodylate buffer, pH 7.2. The cells were then dehydrated in acetone and embedded in epoxy resin. Ultrathin sections were stained with uranyl acetate and lead citrate and examined in a Zeiss EM 109 transmission electron microscope.
2.6. Statistical Analysis
Statistical analysis of growth differences between treated and control cultures (IC50) was performed using the Anova with Tukey test, with P < 0.05. The results were expressed as mean values ± standard deviation (S.D.).
3. Results
The antileishmanial activity was directly proportional to usnic acid and amphotericin B concentrations. Usnic acid antileishmanial activity was estimated by the IC50 concentration at 72 h after incubation. Usnic acid was found to exhibit a lower inhibitory activity against Leishmania infantum chagasi (IC50 = 18.30 ± 2.00 μg/mL) than the reference drug amphotericin B (1.36 ± 0.067 μg/mL) (Figure 1).
Figure 1.
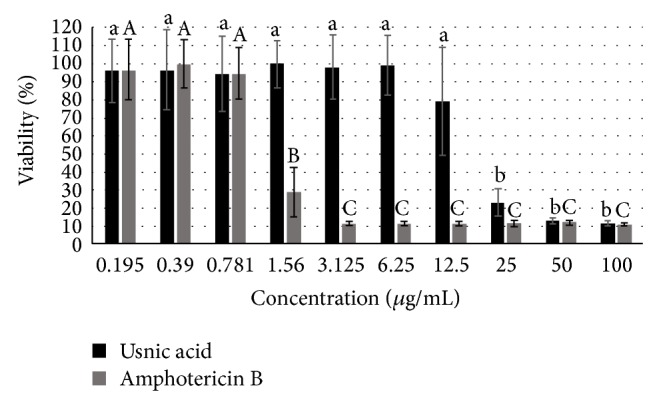
In vitro effects of different concentrations of usnic acid and amphotericin B on the growth of Leishmania infantum chagasi promastigote forms. Means followed by different letters indicating statistically significant differences according to the Anova-Tukey test (P < 0.05, compared to controls).
The analysis of scanning electron micrographs of treated parasites demonstrated that usnic acid affected the parasite surface. Morphological alterations were observed as the presence of blebs in the cell membrane and of shapes given off. No ultrastructural change was observed in promastigotes grown with LIT-DMSO for 72 h, showing the elongated normal morphology (Figures 2(a)–2(f)).
Figure 2.

Scanning electron micrographs of Leishmania infantum chagasi promastigotes exposed to usnic acid. (a) Control group (no treatment); (b, c, d, e, and f) treated group usnic acid at a concentration of 25 μg/mL for 72 hours. Blebs on the surface of promastigotes (thin arrows), membrane rupture (full arrow), swelling (asterisk), and the formation pore (arrowhead). Bars: 1 μm (a, d); 2 μm (c, e and f). Magnifications: (a) 11.000x, (b) 5.000x, (c) 8.000x, (d) 13.000x, (e) 5.500x, and (f) 8.500x.
Morphological alterations of Leishmania infantum chagasi promastigotes treated with 25 μg/mL usnic acid were observed by transmission electron microscopy. The compound caused changes in the cytoplasm density, cell swelling, and loss of cell polarity. Changes in mitochondrial morphology were characterized by marked swelling. We also noticed an increased number of intracellular vacuoles. Blebs were observed in the plasma membrane and detached from the membrane. It was also observed that the membrane was separated from the cytoplasm and there was a higher accumulation of fat compared to controls. Nucleus was observed with presence of an electron-lucent space, when compared to the control (Figures 3(a)–3(f)).
Figure 3.

Transmission electron micrographs promastigote forms of Leishmania infantum chagasi. (a) Control group; (b), (c), (d), (e), and (f) treated groups (AU) at a concentration of 25 μg/mL for 72 hours. Vacuoles (V), acidocalcisomes (arrows), nucleus (N), lipid (L), kinetoplast (Ct), flagellar pocket (Bf), mitochondrial swelling (Mt), detachment of the plasma membrane (dM), blebs (Bl), detached blebs (Bd), and electron-lucent space (Ep). Bars 10 μm (a, c, d, e, and f), Bars 20 μm (b).
4. Discussion
Usnic acid is the most abundant constituent of various species of lichens, including those belonging to the genus Cladonia and Usnea [33]. This compound is known to present several biological activities, acting as an antiprotozoan [34, 35] and antiparasitic [25]. Research with usnic acid in vitro against promastigote forms of Leishmania sp. demonstrated the effectiveness of usnic acid in their antileishmanial activity [35].
The concentration which inhibited 50% growth of a culture of Leishmania infantum chagasi promastigote forms obtained in our experiments (18.30 μg/mL) was close to that obtained by de Carvalho et al. [25] who detected an IC50 around 20 μg/mL for another trypanosomatid parasite (T. cruzi). Our results demonstrated a promising leishmanicidal activity of usnic acid against the Leishmania infantum chagasi promastigote forms in vitro when confronted with the study of Jota et al. [36] who obtained for other depsides from lichens (atranorin and norstictic acid), IC50 values for Leishmania infantum chagasi at concentrations of 30 and 40 µg/mL, respectively.
In a literature review conducted until October 2006 few references regarding the leishmanicidal activity of compounds isolated from lichens were found. Fournet et al. [35] revealed that among the lichen substances tested in vitro against promastigote forms of L. braziliensis, L. amazonensis and L. donovani, the usnic acid presented total lysis of parasites in the concentration of 25 µg/mL. The promastigote forms of L. amazonensis and L. braziliensis under the effect of usnic acid exhibited lysis of parasites at concentration of 10 µg/mL (80–90%) and against L. donovani the activity was total. Already, depsidones (panarin and 1-cloropanarin) were active only in concentrations equal to or greater than 50 μg/mL.
Scanning electron micrographs reveal the formation of blebs in the plasma membrane, membrane rupture, and loss of polarity. da Silva et al. [6] when using warifteine drug against promastigotes of Leishmania infantum chagasi found similar results in scanning microscopy, such as blebs, suggesting that usnic acid metabolism operates through a change in intracellular calcium concentration.
Comparing the ultrastructural effects of usnic acid with those produced by other drug (amiodarone), against the promastigotes and amastigotes forms of L. amazonensis evaluated by da Silva et al. [6], was observed similar effects on the membranes of parasites, such as lipid accumulation and loss of integrity of nuclear envelope and plasma membrane, leading to cell death by necrosis.
According to Abo-Khatwa et al. [37] usnic acid would be similar to the classical uncoupler 2, 4-dinitrophenol (DNP) oxidative phosphorylation, and thereby promote ATP depletion. Furthermore, usnic acid has lipophilic characteristics and ionizing could alter the mitochondrial membrane, causing damage of this organelle [15].
Einarsdóttir et al. [38] showed that mitochondrial membrane potential loss appears to be dose-dependent with respect to the effect of usnic acid in the two cell lines such as breast cancer T-47D and pancreatic cancer Capan-2. These data demonstrate and confirm one of the mechanisms of action of usnic acid previously cited by Müller [15], where changes in membrane lipid composition alter the physical properties with loss of mitochondrial function. We suggest, based on prior observations, that the usnic acid destabilized membranes allowing the leishmanicidal effect.
Adade and Souto-Padrón [39] studying the ultrastructures of the genera Trypanosoma and Leishmania regarding the antiparasitic drugs observed that the ultrastructural effects are similar among the various drugs used as well as between herbal medicines, including usnic acid.
The study in question was relevant to effectiveness of usnic acid in the face of Leishmania infantum chagasi promastigote forms, which leads us to suggest it as a possible phytotherapic agent in the treatment of visceral leishmaniasis. In this context, further studies are needed to elucidate the mechanisms of action of usnic acid and its physiological effects in vivo.
Acknowledgments
The authors like to thank the PROPESQ/UFPE, the Coordination for the Graduated Personnel (CAPES), and Aggeu Magalhães Institute/Oswaldo Cruz Foundation (CPqAM-FIOCRUZ).
Conflict of Interests
The authors declare that there is no conflict of interests that could affect the impartiality of the research reported.
References
- 1.World Health Organization. Report of a Meeting of the WHO Expert Committee on the Control of Leishmaniases. Geneva, Switzerland: WHO; 2010. Technical report series: control of leishmaniases. http://whqlibdoc.who.int/ trs/WHO TRS 949 eng.pdf. [Google Scholar]
- 2.Dos Santos Ferreira C., Silveira Martins P., Demicheli C., Brochu C., Ouellette M., Frézard F. Thiol-induced reduction of antimony(V) into antimony(III): a comparative study with trypanothione, cysteinyl-glycine, cysteine and glutathione. BioMetals. 2003;16(3):441–446. doi: 10.1023/a:1022823605068. [DOI] [PubMed] [Google Scholar]
- 3.Dujardin J.-C., Campino L., Cañavate C., et al. Spread of vector-borne diseases and neglect of leishmaniasis, Europe. Emerging Infectious Diseases. 2008;14(7):1013–1018. doi: 10.3201/eid1407.071589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Machado C. M., Martins T. C., Colturato I., et al. Epidemiology of neglected tropical diseases in transplant recipients. Review of the literature and experience of a Brazilian HSCT center. Revista do Instituto de Medicina Tropical de São Paulo. 2009;51(6):309–324. doi: 10.1590/s0036-46652009000600003. [DOI] [PubMed] [Google Scholar]
- 5.Pavli A., Maltezou H. C. Leishmaniasis, an emerging infection in travelers. International Journal of Infectious Diseases. 2010;14(12):e1032–e1039. doi: 10.1016/j.ijid.2010.06.019. [DOI] [PubMed] [Google Scholar]
- 6.da Silva E. C., Rayol C. D., Medeiros P. L., et al. Antileishmanial activity of warifteine: a bisbenzylisoquinoline alkaloid isolated from Cissampelos sympodialis Eichl. (Menispermaceae) The Scientific World Journal. 2012;2012:5. doi: 10.1100/2012/516408.516408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chakravarty J., Sundar S. Drug resistance in leishmaniasis. Journal of Global Infectious Diseases. 2010;2(2):167–176. doi: 10.4103/0974-777x.62887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Olliaro P. L., Guerin P. J., Gerstl S., Haaskjold A. A., Rottingen J. A., Sundar S. Treatment options for visceral leishmaniasis: a systematic review of clinical studies done in India, 1980-2004. The Lancet Infectious Diseases. 2005;5(12):763–774. doi: 10.1016/s1473-3099(05)70296-6. [DOI] [PubMed] [Google Scholar]
- 9.Croft S. L., Seifert K., Yardley V. Current scenario of drug development for leishmaniasis. Indian Journal of Medical Research. 2006;123(3):399–410. [PubMed] [Google Scholar]
- 10.Sundar S., More D. K., Singh M. K., et al. Failure of pentavalent antimony in visceral leishmaniasis in India: report from the center of the Indian epidemic. Clinical Infectious Diseases. 2000;31(4):1104–1107. doi: 10.1086/318121. [DOI] [PubMed] [Google Scholar]
- 11.Chappuis F., Sundar S., Hailu A., et al. Visceral leishmaniasis: what are the needs for diagnosis, treatment and control? Nature Reviews Microbiology. 2007;5(11):873–882. doi: 10.1038/nrmicro1748. [DOI] [PubMed] [Google Scholar]
- 12.Dorlo T. P. C., Balasegaram M., Beijnen J. H., de vries P. J. Miltefosine: a review of its pharmacology and therapeutic efficacy in the treatment of leishmaniasis. Journal of Antimicrobial Chemotherapy. 2012;67(11):2576–2597. doi: 10.1093/jac/dks275. [DOI] [PubMed] [Google Scholar]
- 13.Fournet A., Muñoz V. Natural products as trypanocidal, antileishmanial and antimalarial drugs. Current Topics in Medicinal Chemistry. 2002;2(11):1215–1237. doi: 10.2174/1568026023393011. [DOI] [PubMed] [Google Scholar]
- 14.Inacio J. D. F., Canto-Cavalheiro M. M., Almeida-Amaral E. E. In vitro and in vivo effects of (−)-epigallocatechin 3-O-gallate on Leishmania amazonensis . Journal of Natural Products. 2013;76(10):1993–1996. doi: 10.1021/np400624d. [DOI] [PubMed] [Google Scholar]
- 15.Müller K. Pharmaceutically relevant metabolites from lichens. Applied Microbiology and Biotechnology. 2001;56(1-2):9–16. doi: 10.1007/s002530100684. [DOI] [PubMed] [Google Scholar]
- 16.Honda N. K., Vilegas W. A química dos liquens. Química Nova. 1998;6(21):110–125. [Google Scholar]
- 17.Plichet A. Usnic acid from lichens in therapy of tuberculosis. La Presse Médicale. 1955;63(17, article 350) [PubMed] [Google Scholar]
- 18.Cocchietto M., Skert N., Nimis P., Sava G. A review on usnic acid, an interesting natural compound. Naturwissenschaften. 2002;89(4):137–146. doi: 10.1007/s00114-002-0305-3. [DOI] [PubMed] [Google Scholar]
- 19.Cardarelli M., Serino G., Campanella L., et al. Antimitotic effects of usnic acid on different biological systems. Cellular and Molecular Life Sciences. 1997;53(8):667–672. doi: 10.1007/s000180050086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Okuyama E., Umeyama K., Yamazaki M., Kinoshita Y., Yamamoto Y. Usnic acid and diffractaic acid as analgesic and antipyretic components of Usnea diffracta. Planta Medica. 1995;61(2):113–115. doi: 10.1055/s-2006-958027. [DOI] [PubMed] [Google Scholar]
- 21.Vijayakumar C. S., Viswanathan S., Kannappa Reddy M., Parvathavarthini S., Kundu A. B., Sukumar E. Anti-inflammatory activity of (+)-usnic acid. Fitoterapia. 2000;71(5):564–566. doi: 10.1016/S0367-326X(00)00206-9. [DOI] [PubMed] [Google Scholar]
- 22.Campanella L., Delfini M., Ercole P., Iacoangeli A., Risuleo G. Molecular characterization and action of usnic acid: a drug that inhibits proliferation of mouse polyomavirus in vitro and whose main target is RNA transcription. Biochimie. 2002;84(4):329–334. doi: 10.1016/s0300-9084(02)01386-x. [DOI] [PubMed] [Google Scholar]
- 23.Scirpa P., Scambia G., Masciullo V., et al. Adjuvant treatment of HPV infection. A perspective and controlled study on the efficacy and safety of intravaginal administration of usnic acid and zinc sulphate compound. Minerva Ginecologica. 1999;51(6):255–260. [PubMed] [Google Scholar]
- 24.Halama P., van Haluwin C. Antifungal activity of lichen extracts and lichenic acids. BioControl. 2004;49(1):95–107. doi: 10.1023/b:bico.0000009378.31023.ba. [DOI] [Google Scholar]
- 25.de Carvalho E. A. B., Andrade P. P., Silva N. H., Pereira E. C., Figueiredo R. C. B. Q. Effect of usnic acid from the lichen Cladonia substellata on Trypanosoma cruzi in vitro: an ultrastructural study. Micron. 2005;36(2):155–161. doi: 10.1016/j.micron.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 26.Santos L. C., Honda N. K., Carlos I. Z., Vilegas W. Intermediate reactive oxygen and nitrogen from macrophages induced by Brazilian lichens. Fitoterapia. 2004;75(5):473–479. doi: 10.1016/j.fitote.2004.04.002. [DOI] [PubMed] [Google Scholar]
- 27.Bucar F., Schneider I., Ögmundsdóttir H., Ingólfsdóttir K. Anti-proliferative lichen compounds with inhibitory activity on 12(S)-HETE production in human platelets. Phytomedicine. 2004;11(7-8):602–606. doi: 10.1016/j.phymed.2004.03.004. [DOI] [PubMed] [Google Scholar]
- 28.Mayer M., O'Neill M. A., Murray K. E., et al. Usnic acid: a non-genotoxic compound with anti-cancer properties. Anti-Cancer Drugs. 2005;16(8):805–809. doi: 10.1097/01.cad.0000175588.09070.77. [DOI] [PubMed] [Google Scholar]
- 29.da Silva Santos N. P., Nascimento S. C., Wanderley M. S. O., et al. Nanoencapsulation of usnic acid: improvement on the antitumour activity and reduction of hepatotoxicity. European Journal of Pharmaceutics and Biopharmaceutics. 2006;64(2):154–160. doi: 10.1016/j.ejpb.2006.05.018. [DOI] [PubMed] [Google Scholar]
- 30.Bazin M.-A., Le-Lamer A.-C., Delcros J.-G., et al. Synthesis and cytotoxic activities of usnic acid derivatives. Bioorganic & Medicinal Chemistry. 2008;16(14):6860–6866. doi: 10.1016/j.bmc.2008.05.069. [DOI] [PubMed] [Google Scholar]
- 31.Odabasoglu F., Cakir A., Suleyman H., et al. Gastroprotective and antioxidant effects of usnic acid on indomethacin-induced gastric ulcer in rats. Journal of Ethnopharmacology. 2006;103(1):59–65. doi: 10.1016/j.jep.2005.06.043. [DOI] [PubMed] [Google Scholar]
- 32.Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. Journal of Immunological Methods. 1983;65(1-2):55–63. doi: 10.1016/0022-1759(83)90303-4. [DOI] [PubMed] [Google Scholar]
- 33.Nishitoba Y., Nishimura I., Nishiyama T., Mizutani J. Lichen acids, plant growth inhibitors from Usnea longissima . Phytochemistry. 1987;26(12):3181–3185. doi: 10.1016/s0031-9422(00)82466-8. [DOI] [Google Scholar]
- 34.Wu J., Zhang M., Ding D., Tan T., Yan B. Effect of Cladonia alpestris on Trichomonas vaginalis in vitro . Chinese Journal of Parasitology & Parasitic Disease. 1995;13(2):126–129. [PubMed] [Google Scholar]
- 35.Fournet A., Ferreira M. E., de Arias A. R., et al. Activity of compounds isolated from Chilean lichens against experimental cutaneous leishmaniasis. Comparative Biochemistry and Physiology C: Pharmacology, Toxicology and Endocrinology. 1997;116(1):51–54. doi: 10.1016/S0742-8413(96)00127-2. [DOI] [PubMed] [Google Scholar]
- 36.Jota V. C. S., Andrade P. P., Silva N. H., Pereira E. C., Figueiredo R., Honda N. K. Análise in vitro de substâncias liquênicas sobre Leishmania chagasi . II Encontro do Grupo Brasileiro de Liquenólogos (EGBL-2); 1998; Campo Grande, Brazil. [Google Scholar]
- 37.Abo-Khatwa A. N., Al-Robai A. A., Al-Jawhari D. A. Lichen acids as uncouplers of oxidative phosphorylation of mouse-liver mitochondria. Natural Toxins. 1996;4(2):96–102. doi: 10.1002/19960402nt7. [DOI] [PubMed] [Google Scholar]
- 38.Einarsdóttir E., Groeneweg J., Björnsdóttir G. G., et al. Cellular mechanisms of the anticancer effects of the lichen compound usnic acid. Planta Medica. 2010;76(10):969–974. doi: 10.1055/s-0029-1240851. [DOI] [PubMed] [Google Scholar]
- 39.Adade C. M., Souto-Padrón T. Contributions of ultrastructural studies to the cell biology of trypanosmatids: targets for anti-parasitic drugs. The Open Parasitology Journal. 2010;4(1):178–187. doi: 10.2174/1874421401004010178. [DOI] [Google Scholar]