

Abstract
Much hypoxia research has been carried out at high altitude in a hypobaric hypoxia (HH) environment. Many research teams seek to replicate high-altitude conditions at lower altitudes in either hypobaric hypoxic conditions or normobaric hypoxic (NH) laboratories. Implicit in this approach is the assumption that the only relevant condition that differs between these settings is the partial pressure of oxygen (PO2), which is commonly presumed to be the principal physiological stimulus to adaptation at high altitude. This systematic review is the first to present an overview of the current available literature regarding crossover studies relating to the different effects of HH and NH on human physiology. After applying our inclusion and exclusion criteria, 13 studies were deemed eligible for inclusion. Several studies reported a number of variables (e.g. minute ventilation and NO levels) that were different between the two conditions, lending support to the notion that true physiological difference is indeed present. However, the presence of confounding factors such as time spent in hypoxia, temperature, and humidity, and the limited statistical power due to small sample sizes, limit the conclusions that can be drawn from these findings. Standardisation of the study methods and reporting may aid interpretation of future studies and thereby improve the quality of data in this area. This is important to improve the quality of data that is used for improving the understanding of hypoxia tolerance, both at altitude and in the clinical setting.
Keywords: Normobaric hypoxia, Hypobaric hypoxia, Altitude
Background
Hypoxia research has numerous applications. These include investigating the pathogenesis and developing treatments for medical conditions characterised by hypoxia [1] and acute high altitude illness [2], as well as setting optimum training regimes for athletes [3].
Much hypoxia research has been carried out at high altitude in a hypobaric hypoxia (HH) environment. Such ‘field’ studies present practical and logistical challenges including safety concerns about carrying out invasive procedures in a remote setting. For these reasons, many research teams seek to replicate high-altitude conditions at lower altitudes in either hypobaric hypoxic conditions or normobaric hypoxic (NH) laboratories. In these two conditions, the hypoxic dose is calculated by the combination of the various barometric pressures × inspired fraction of oxygen [4]. As emphasised in Conkin's ‘Critique of the equivalent air altitude model’ [5], implicit in this approach is the assumption that the only relevant condition that differs between these settings is the partial pressure of oxygen (PO2), which is commonly presumed to be the principal physiological stimulus to adaptation at high altitude [6]. Although this assumption underpins the interpretation of many studies that form the basis of hypoxia physiology, it remains open to question as recently highlighted by Millet et al. [7] and controversy exists relating to the sporadic data in this area with various opinions on the matter as discussed in a recent series of ‘point-counterpoints’ [8]. The notion that HH and NH environments are interchangeable in terms of their effect on physiological responses is not proven.
The practical outcomes of this debate affect a variety of fields. Many national teams in various sports incorporate altitude or hypoxic training into their programmes to aid haematological adaptations [3]. Additionally, armies across the world employ pre-acclimatisation strategies to train troops for deployment at high altitude [9]. This issue also applies to medical research such as therapeutic intermittent hypoxic methods [10] or critical care research into tissue hypoxia [1]. Thus, understanding the different impacts of NH and HH on physiology is important.
The aim of this systematic review is to conduct a comprehensive systematic literature search to address the questions: do humans react differently to HH when compared with NH (when evaluated in studies with a crossover design)?
Review: methods
Criteria for considering studies for this review
Candidate studies were identified using the following criteria (Figure 1).
Figure 1.
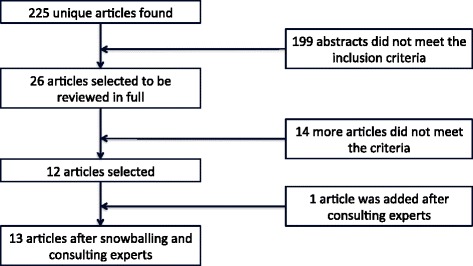
Methodology flow chart. This flow chart shows search methodology and results.
Types of studies
We searched for primary research articles describing crossover trials comparing physiological responses to NH and HH. Only crossover trials were considered due to the large inter-subject variation in their response to hypoxia.
Types of participants
We included studies involving lowland (defined as permanently living in locations <2,000 m) human subjects of any age who were not acclimatised to high altitude.
Types of interventions
We compared NH and HH. NH and HH must be calculated to be equivalent to the same altitude. We included studies investigating any duration of exposure, and the HH may have been performed either at sea level in a hypobaric chamber or at high altitude.
Types of outcome measures
Outcome variables were any human physiological response to atmospheric hypoxia. These responses included common phenotypes of interest in high altitude literature including (but not limited to) ventilation, hypoxaemia, exercise metabolism, nitric oxide (NO) production, osmotic balance, erythropoiesis and high-altitude illness.
Search methods for identification of studies
Search strategy
We attempted to identify all relevant trials regardless of language or publication status (published, unpublished, in press, and in progress). A literature search was carried out using the search engines Embase (all to date), Medline (performed on 15 October 2013) and Web of Science (performed on 15 October 2013). Snowballing was carried out; thus, the reference lists of all the shortlisted studies were checked for possible eligible studies.
Search terms
Search terms include (‘Hypobari* hypoxia’ OR ‘simulated altitude’ OR ‘hypobari* anoxia’) AND (‘normbari* hypoxia’ OR ‘sea level hypoxia’ OR ‘sea-level hypoxia’ OR ‘normobari* anoxia’).
Data collection and analysis
Selection of studies
Titles and abstracts of candidate studies were screened for eligibility and duplicate references independently by two authors (JC and EG). The reasons for study exclusion were independently documented. For those papers that could not be excluded based on their titles and abstracts, the full paper was read to confirm eligibility. We resolved disagreements by consulting a third author (PH) who arbitrated on inclusion. We obtained the assistance of translators when abstracts were not available in English.
Data extraction and management
Using data extraction forms, JC extracted information from each study and EG crosschecked the data. Data fields within the data extraction forms were directly linked to the formulated review question and planned assessment of included studies.
The data extraction forms contained the following information: study reference and reviewer identity, verification of study eligibility, study characteristics, study quality (see Quality of data section below), research methods, participants, intervention, outcome measures, results, and additional information.
Assessment of risk of bias and study quality in included studies
The risk of bias was independently assessed by JC and EG. Studies that do not report statistical significance (P values) for reported results were included, but their results were considered either as high risk of bias or unclear. In terms of study quality, we assessed the following: randomisation of subjects for the order of the crossover and whether they were blinded, length of washout period, presence/absence of sample size calculations, whether the statistical analyses accounted for the increased risk of type I errors when analysing large numbers of variables (adjustment for multiple comparisons), test-retest reliability, normalisation of testing environments for humidity (pH2O) as this can impact hypoxic dose calculations [11], and control of carbon dioxide (CO2) in the testing environments.
Data synthesis
The results were tabulated and compared. No statistical analysis was carried out because the heterogeneity of the studies was such that the data could not be pooled (see below). Studies were categorised according to duration of hypoxic exposure. One hour was used as the cut off between ‘long’ and ‘short’ studies. All variables identified through our search strategy are highlighted in Table 1 (List of variables) but due to the number of variables reported, not all are considered in the written results and discussion. Emphasis is placed on the major physiological variables and those that are reported in more than one study. This was because a consistent result across multiple independent studies suggests validity of the finding. Additionally, when study characteristics were being determined, if a study did not mention a design feature, such as randomization of exposure order, it was assumed that the feature was not present. The results for each variable and time point can be found in Tables 2, 3, 4, 5, 6.
Table 1.
List of variables
| Oxygenation and ventilation | Cardiovascular | AMS (acute mountain sickness) and neurology | Other |
|---|---|---|---|
| Ventilation (VE) | Hypoxic cardiac response (HCR) | Postural stability | Sweat rates |
| Tidal volume (VT) | Forced vascular conductance | ||
| Lake Louise AMS scores | Oesophageal temperature thresholds for increasing forearm skin vascular conductance | ||
| Respiratory rate (Bf) | Heart rate variability (LH/HF) | ||
| Exhaled nitric oxide (NO) levels (exNO) | Oesophageal temperature thresholds for increased sweat rate | ||
| Heart rate (HR) | |||
| Oesophageal temperature | |||
| End tidal partial pressure of oxygen (PetO2) | Stroke volume (SV) | Skin temperature | |
| Urine volume | |||
| Cardiac output (CO) | Plasma volume | ||
| End tidal partial pressure of carbon dioxide (PETCO2) | Glomerular filtration rate (GFR) | ||
| Plasma potassium concentration (plasma K+) | |||
| Plasma sodium concentration (plasma Na+) | |||
| Blood pressure (BP) | |||
| Plasma renin activity | |||
| Plasma aldosterone | |||
| Alveolar ventilation (VA) | Free water clearance | ||
| Adrenocorticotropic hormone (ACTH) | |||
| Volume of CO2 produced (VCO2) | |||
| Anti-diuretic hormone (ADH) | |||
| Anti-naturetic protein (ANP) | |||
| Blood base excess | |||
| Urine sodium-potassium ratio (urine Na+/K+) | |||
| Volume of oxygen consumed (VO2) | |||
| Catecholamines, plasma osmolarity | |||
| PH | |||
| End tidal fraction of oxygen (FetO2) | Urine osmolarity | ||
| Plasma lactate levels | |||
| Blood NO metabolites | |||
| Glutathione peroxidase (GPX) | |||
| MDA | |||
| Nitrotyrosine | |||
| End tidal fraction of oxygen (FetCO2) | Plasma advanced oxidation protein products and superoxide dismutase | ||
| Duration of inspiration and expiration | Haematocrit (Hct) | ||
| Haemoglobin concentration (Hb conc) | |||
| Hypoxic ventilatory response (HVR) | |||
| Respiratory quotient (RQ) | |||
| Peripheral oxygen saturations (SpO2) | |||
| Arterial oxygen saturations (SaO2) | |||
| Arterial oxygen and carbon dioxide partial pressure (PaO2) (PaCO2) | |||
| Arterial oxygen content | |||
| Alveolar-arterial PO2 difference |
All the variables measured in the 13 accepted studies are listed. These have been subdivided into physiological systems.
Table 2.
Study design
| Author and year | Type of outcome variable | Population | PiO 2 of exposure (calculated by PiO 2 = (Pb-47) × FiO 2 [4]) (mmHg) | Duration of exposure (h:min) | Randomised (Y, N) | Washout period between trials (days:h) | |
|---|---|---|---|---|---|---|---|
| NH HH | |||||||
| Basualto-Alarcon 2012 | Ventilatory and cardiovascular + exercise | 7 men | 3,000 mc | 3,000 mc | 00:15 builda, 00:10 | N | 7 |
| Degache 2012 | Postural stability | 12 men | 118 and 102 | 121 and 103 | 00:20–25 builda, 00:30b | Nil stated | <00:24 |
| Hemmingsson 2009 | Exhaled NO | 6 men, 2 women | 103 and 81 | 99 and 75 | 00:10 at each altitude | Y | <00:12 |
| Loeppky 2005 | Fluid balance | 9 men | 81 | 80 | 00:05 builda 10:00 | Y | 7 |
| Loeppky 1997* | Ventilatory and cardiovascular | 9 men | 81 | 80 | 00:05 builda 10:00 | Y | 7 |
| Miyagawa 2010 | Ventilatory, cardiovascular and sweat + exercise | 7 men | 93 | 97 | 00:30 builda only for HH 01:40 | Y | >6 |
| Naughton 1995 | Haematological + exercise | 9 subjects with chronic airflow limitation (CAL) with 6 controls | 1,829 and 2,438 mc | 117 and 108 | 00:12 builda included 00:52:00 | Y | 00:02 |
| Roach 1996 | AMS and cardiovascular | 9 men | 4,564 mc | 80 | 09:00 | Y | 7 |
| Savourey 2003 | Ventilatory and cardiovascular | 18 men | 4,500 mc | 4,500 mc | 00:10 builda, 00:30 | Y | 7 |
| Savourey 2007 | Ventilatory and cardiovascular | 17 men, 1 woman | 4,500 mc | 4,500 mc | 00:10 builda, 00:30 | Y | 14 |
| Self 2011 | AMS and cardiovascular | 17 men and 3 women | 7,620 mc | 7,620 mc | 00:05 | N | <00:24 |
| Tucker 1983 | Mixed | 11 men | 82 | 80 | 00:15 builda included 02:20 | N | Several weeks |
| Faiss 2013 | Exhaled NO, ventilatory and cardiovascular + exercise | 10 men | 99 | 101 | 24:00:00 | Y | 23 |
This table describes all the features of the accepted studies.
aWhen the subjects entered the chamber, the environmental conditions were that of normal sea level but then were gradually made more hypoxic over the specified amount of time until the target hypoxic dose was reached.
bThe different altitudes were tested consecutively. So sometimes the exposure was 1 h at 3,000 or 1,700 m.
cWhen PiO2 could not be calculated due to lack of information, the equivalent attitude estimated by the authors was given.
*P values were for three conditions; in recovery no P values unless stated in the discussion.
Table 3.
Oxygenation and ventilation variables
| Outcome | Author and year | Duration of exposure (h:min) | Hypobaric hypoxia result [mean (SD)] | Normobaric hypoxia result [mean (SD)] | Difference (HH − NH) | P value | Direction of difference NH compared to HH |
|---|---|---|---|---|---|---|---|
| VE (L/min) | |||||||
| Loeppky 1997 | 00:00 | 12.9 | 13.6 | - | - | - | |
| Savourey 2007 | 00:10 build, 00:05 | 10.49 (2.59) | 10.14 (1.51) | - | >0.05 | NS | |
| Basualto-Alarcon 2012 | 00:15 build, 00:05 acclimatisation | 10.5 (4.9) | 10.3 (1.8) | 0.2 | - | - | |
| Basualto-Alarcon 2012 | 00:15 build, 00:10 acclimatisation | 35.7 (5.9) | 39.7 (6.7) | -4 | <0.05 | NH > HH | |
| Savourey 2003 | 00:10 build, 00:30 | - | - | - | <0.02 | NH > HH | |
| Savourey 2007 | 00:10 build, 00:30 | 10.70 (1.93) | 10.78 (1.93) | - | >0.05 | NS | |
| Faiss 2013 | 01:00 | 13.6 (1.8) | 13.3 (3.3) | 0.3 | >0.05 | NS | |
| Miyagawa 2010a | 01:05 | 69.2 | 65.6 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:10 | 70.5 | 65.7 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:20 | 73.9 | 70.9 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:40 | 75.3 | 77 | - | >0.05 | NS | |
| Tucker 1983b | 02:20 including 00:15 build | 2.07 | 4.82 | −2.75 | - | - | |
| Loeppky 1997 | 03:00 | 10.3 | 14.3 | −4.0 | <0.01 | NH > HH | |
| Loeppky 1997 | 06:00 | 10.6 | 12.7 | −2.1 | <0.05 | NH > HH | |
| Faiss 2013 | 08:00 | 11.8 (1.9) | 14.9 (3.5) | −3.1 | <0.1 | NH > HH | |
| - | 10.7 (1.8) | 12.2 (1.6) | −1.5 | <0.05 | NH > HH | ||
| - | 12.7 (2.3) | 14.2 (1.5) | −1.5 | <0.1 | NH > HH | ||
| Loeppky 1997 | 09:00 | 10.2 | 12.2 | - | <0.05 | NH > HH | |
| Loeppky 1997 | Recovery 12:00:00 | 9.2 | 10.1 | - | >0.05 | NS | |
| VT (L) | |||||||
| Savourey 2007 | 00:10 buildc, 00:05 | 0.72 (0.25) | 0.88 (0.22) | - | 0.03 | NH > HH | |
| Basualto-Alarcon 2012 | 00:15 buildc, 00:05 acclimatisation | 0.81 (0.36) | 0.82 (0.21) | −0.01 | - | - | |
| Basualto-Alarcon 2012 | 00:15 buildc, 00:10 acclimatisation | 1.85 (0.56) | 1.91 (0.53) | −0.06 | - | NH > HH | |
| Savourey 2003 | 00:10 buildc, 00:30 | - | - | - | <0.001 | NH > HH | |
| Savourey 2007 | 00:10 buildc, 00:30 | 0.83 (0.37) | 0.86 (0.34) | - | >0.05 | NS | |
| Faiss 2013 | 01:00 | 0.88 (0.21) | 0.89 (0.26) | −0.01 | >0.05 | NS | |
| Tucker 1983b | 02:20 including 00:15 buildc | 106 | 152 | -46 | - | NH > HH | |
| Faiss 2013 | 08:00 | 0.75 (0.21) | 0.94 (0.3) | −0.19 | <0.05 | NH > HH | |
| Faiss 2013 | 16:00 | 0.75 (0.23) | 0.84 (0.24) | −0.9 | <0.1 | NH > HH | |
| Loeppky 1997 | 10:00 | - | - | - | >0.05 | NS | |
| Faiss 2013 | 24:00:00 | 0.86 (0.25) | 0.95 (0.23) | −0.09 | <0.05 | NH > HH | |
| Bf (cycles/min) | |||||||
| Savourey 2007 | 00:10 buildc, 00:05 | 15.73 (4.64) | 12.24 (3.80) | - | 0.03 | HH > NH | |
| Basualto-Alarcon 2012 | 00:15 buildc, 00:05 acclimatisation | 13.3 (4.0) | 13.4 (4.7) | −0.1 | - | - | |
| Basualto-Alarcon 2012 | 00:15 buildc, 00:10 acclimatisation | 20.4 (5.4) | 22.3 (7.4) | −1.9 | <0.05 | NH > HH | |
| Savourey 2003 | 00:10 buildc, 00:30 | - | - | - | <0.001 | HH > NH | |
| Savourey 2007 | 00:10 buildc, 00:30 | 14.77 (4.17) | 13.76 (4.47) | - | >0.05 | NS | |
| Faiss 2013 | 01:00 | 16.8 (3.4) | 15.9 (4.2) | −0.9 | >0.05 | NS | |
| Miyagawa 2010 | 01:05 | 31 | 31 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:10 | 32 | 34 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:20 | 34 | 36 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:40 | 38 | 42 | - | >0.05 | NS | |
| Tucker 1983b | 02:20 including 00:15 buildc | 0.9 | 3.8 | −2.9 | - | NH > HH | |
| Faiss 2013 | 08:00 | 16.8 (2.7) | 17.1 (4.4) | −0.3 | >0.05 | NS | |
| Loeppky 1997 | 10:00 | - | - | - | >0.05 | NS | |
| Faiss 2013 | 16:00 | 16.1 (3) | 15.8 (3.7) | −0.3 | >0.05 | NS | |
| Faiss 2013 | 24:00:00 | 16.8 (3.8) | 16.2 (3.8) | 0.6 | >0.05 | NS | |
| PetO2 (mmHg) | |||||||
| Savourey 2007 | 00:10 buildc, 00:05 | 72.5 (6.58) | 79.56 (11.94) | - | 0.08 | Borderline NH > HH | |
| Savourey 2003 | 00:10 buildc, 00:30 | - | - | - | >0.05 | NS | |
| Savourey 2007 | 00:10 buildc, 00:30 | 73.15 (7.16) | 76.09 (11.61) | - | 0.08 | Borderline NH > HH | |
| Faiss 2013 | 01:00 | 66.4 (4.1) | 62.3 (2.8) | - | 4.1 | NS | |
| Tucker 1983b | 02:20 including 00:15 buildc | −36.1 | −32.6 | −3.5 | - | - | |
| Faiss 2013 | 08:00 | 61.9 (6.0) | 61.6 (2.2) | 0.3 | >0.05 | NS | |
| Loeppky 1997 | 10:00 | - | - | - | >0.05 | NS | |
| Faiss 2013 | 16:00 | 65.0 (5.4) | 62.7 (2.6) | 2.3 | >0.05 | NS | |
| Faiss 2013 | 24:00:00 | 65.6 (5.5) | 65.6 (2.8) | 0 | >0.05 | NS | |
| PetCO2 (mmHg) | |||||||
| Savourey 2007 | 00:10 build, 00:05 | 44.09 (6.38) | 48.87 (5.53) | - | 0.05 | Borderline NH > HH | |
| Savourey 2003 | 00:10 build, 00:30 | - | - | - | >0.05 | NS | |
| Savourey 2007 | 00:10 build, 00:30 | 43.43 (6.02) | 46.13 (6.61) | - | >0.05 | NS | |
| Faiss 2013 | 01:00 | 33.4 (2.5) | 29.4 (2.4) | 4 | <0.1 | HH > NH | |
| Tucker 1983c | 02:20 including 00:15 build | −2.8 | −3.6 | 0.8 | >0.05 | NS | |
| Faiss 2013 | 08:00 | 33.8 (2.1) | 27.5 (1.3) | 6.3 | <0.01 | HH > NH | |
| Loeppky 1997 | 10:00 | - | - | −1.6 | <0.02 | NH > HH | |
| Faiss 2013 | 16:00 | 33.1 (1.3) | 27.9 (0.9) | 5.2 | <0.01 | HH > NH | |
| Faiss 2013 | 24:00:00 | 30.8 (1.4) | 26.5 (1.5) | 4.3 | <0.01 | HH > NH | |
| VA (alveolar ventilation L/min) | |||||||
| Loeppky 1997 | 00:00 | 9.4 | 10.1 | - | - | - | |
| Loeppky 1997 | 03:00 | 7.2 | 10.5 | −46% | <0.05 | NH > HH | |
| Loeppky 1997 | 06:00 | 7.6 | 9.1 | - | <0.05 | NH > HH | |
| Loeppky 1997 | 09:00 | 7.6 | 9.2 | - | <0.05 | NH > HH | |
| Loeppky 1997 | Recovery 12:00:00 | 6.7 | 7.2 | - | - | - | |
| VCO2 ml/min | |||||||
| Loeppky 1997 | 00:00 | 295 | 333 | - | - | - | |
| Miyagawa 2010 | 01:05 | 2188 | 2108 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:10 | 2121 | 2007 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:20 | 2078 | 2060 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:40 | 2021 | 2082 | - | >0.05 | NS | |
| Loeppky 1997 | 03:00 | 216 | 330 | - | <0.05 | NH > HH | |
| Loeppky 1997 | 06:00 | 227 | 296 | - | <0.05 | NH > HH | |
| Loeppky 1997 | 09:00 | 235 | 302 | −67 | <0.05 | NH > HH | |
| Loeppky 1997 | Recovery 12:00 | 241 | 267 | - | - | - | |
| VO2 consumed | |||||||
| Loeppky 1997 | 00:00 | 329 | 340 | - | - | - | |
| Miyagawa 2010 | 01:05 | 1709 | 1611 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:10 | 1783 | 1637 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:20 | 1826 | 1748 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:40 | 1836 | 1840 | - | >0.05 | NS | |
| Loeppky 1997 | 03:00 | 250 | 361 | - | <0.05 | NH > HH | |
| Loeppky 1997 | 06:00 | 262 | 319 | - | <0.05 | NH > HH | |
| Loeppky 1997 | 09:00 | 278 | 326 | - | <0.05 | NH > HH | |
| Loeppky 1997 | 12:00 | 301 | 291 | - | - | - | |
| FetO2 | |||||||
| Savourey 2007 | 00:10 buildc, 00:05 | - | - | - | >0.05 | NS | |
| Savourey 2003 | 00:10 buildc, 00:30 | - | - | - | <0.00001 | HH > NH | |
| Savourey 2007 | 00:10 buildc, 00:30 | - | - | - | >0.05 | NS | |
| FetCO2 | |||||||
| Savourey 2007 | 00:10 build, 00:05 | - | - | - | >0.05 | NS | |
| Savourey 2003 | 00:10 build, 00:30 | - | - | - | <0.00001 | HH > NH | |
| Savourey 2007 | 00:10 build, 00:30 | - | - | - | >0.05 | NS | |
| Duration of inspiration/s | |||||||
| Savourey 2007 | 00:10 buildc, 00:05 | 1.94 (0.65) | 2.99 (0.98) | - | 0.01 | NH > HH | |
| Savourey 2007 | 00:10 buildc, 00:30 | 2.40 (1.25) | 3.00 (1.16) | - | >0.05 | NS | |
| Duration of expiration/s | |||||||
| Savourey 2007 | 00:10 buildc, 00:05 | 2.09 (0.87) | 1.98 (0.84) | - | >0.05 | NS | |
| Savourey 2007 | 00:10 buildc, 00:30 | - | - | - | >0.05 | NS | |
| Hypoxic ventilatory response (HVR) 1%−1 | |||||||
| Savourey 2007 | 00:10 buildc, 00:05 | −0.05 | 0.03 | - | >0.05 | NS | |
| Savourey 2007 | 00:10 buildc, 00:30 | −0.09 | −0.07 | - | >0.05 | NS | |
| SpO2 (%) | |||||||
| Savourey 2007 | 00:10 buildc, 00:05 | 83.03 (4.49) | 87.11 (4.81) | −4.08 | <0.05 | NH > HH | |
| Basualto-Alarcon 2012 | 00:15 buildc, 00:05 acclimatisation | 91.6 (4.2) | 89.1 (3.8) | 2.5 | <0.05 | HH > NH | |
| Basualto-Alarcon 2012 | 00:15 buildc, 00:10 acclimatisation | 85.3 (3.8) | 86.0 (1.7) | −0.7 | - | - | |
| Savourey 2003 | 00:10 buildc, 00:30 | - | - | - | <0.05 | NH > HH | |
| Savourey 2007 | 00:10 buildc, 00:30 | 82.49 (4.39) | 85.50 (4.84) | −2.99 | 0.04 | NH > HH | |
| Faiss 2013 | 01:00 | 93 (1) | 90 (3) | 3 | >0.05 | NS | |
| Tucker 1983b | 02:20 including 00:15 buildc | −13.2 | −13.5 | 0.3 | >0.05 | NS | |
| Faiss 2013 | 08:00 | 91 (3) | 91 (3) | 0 | >0.05 | NS | |
| Roach 1996 | 09:00 | 83% (1%) | 83% (0.7%) | 0 | >0.05 | NS | |
| Loeppky 2005 | 10:00 | 82% | 83% | −1% | >0.05 | NS | |
| Loeppky 1997 | 10:00 | - | - | - | >0.05 | NS | |
| Faiss 2013 | 16:00 | 92 (2) | 91 (2) | 1 | >0.05 | NS | |
| Faiss 2013 | 24:00:00 | 93 (2) | 92 (1) | 1 | >0.05 | NS | |
| SaO2 (%) | |||||||
| Self 2011 | 00:05 | - | - | - | 0.005 | NH > HH | |
| Savourey 2007 | 00:10 buildc, 00:05 | - | - | - | >0.05 | NS | |
| Savourey 2003 | 00:10 buildc, 00:30 | 85% (4) | 88% (3) | −3 | <0.05 | NH > HH | |
| Savourey 2007 | 00:10 buildc, 00:30 | 81.09% (7.76) | 85.48% (5.63) | −4.39 | 0.07 | Borderline NH > HH | |
| Miyagawa 2010 | 01:05 | 82 | 83 | −1 | >0.05 | NS | |
| Miyagawa 2010 | 01:10 | 81 | 82 | −1 | >0.05 | NS | |
| Miyagawa 2010 | 01:20 | 81 | 82 | −1 | >0.05 | NS | |
| Miyagawa 2010 | 01:40 | 82 | 81 | 1 | >0.05 | NS | |
| Roach 1996 | 09:00 | 83% (1%) | 83% (0.7%) | 0 | >0.05 | NS | |
| PaO2 | |||||||
| Self 2011 | 00:05 | - | - | - | 0.004 | HH > NH | |
| Savourey 2007 | 00:10 buildc, 00:05 | - | - | - | >0.05 | NS | |
| Savourey 2003 | 00:10 buildc, 00:30 | 6.38 (0.60) | 6.90 (0.86) | −0.52 | ≤0.05 | Borderline NH > HH | |
| Savourey 2007 | 00:10 buildc, 00:30 | - | - | - | >0.05 | NS | |
| CAL subjects | Naughton 1995 | 00:52 | - | - | −1.1 | >0.05 | NS |
| Control subjects | Naughton 1995 | 00:52 | - | - | 0.7 | >0.05 | NS |
| PaCO2 | |||||||
| Self 2011 | 00:05 | - | - | - | 0.005 | NH > HH | |
| Savourey 2007 | 00:10 build, 00:05 | - | - | - | >0.05 | NS | |
| Savourey 2003 | 00:10 build, 00:30 | 4.65 (0.54) | 5.06 (0.46) | −0.41 | ≤0.05 | Borderline NH > HH | |
| Savourey 2007 | 00:10 build, 00:30 | 46.3 (6.5) | 52.2 (4.2) | −5.9 | 0.005 | NH > HH | |
| CAL subjects | Naughton 1995 | 00:52 | - | - | 0.3 | >0.05 | NS |
| Control subjects | Naughton 1995 | 00:52 | - | - | −0.8 | >0.05 | NS |
| Tucker 1983b | 02:20 including 00:15 build | −3.7 | −5.6 | 1.9 | - | - | |
| Alveolar-arterial PO2 difference | |||||||
| CAL subjects | Naughton 1995 | 00:52 | - | - | 0.7 | >0.05 | NS |
| Control subjects | Naughton 1995 | 00:52 | - | - | 0.2 | >0.05 | NS |
| Arterial O2 content | |||||||
| Savourey 2007 | 00:10 buildc, 00:05 | - | - | - | >0.05 | NS | |
| Savourey 2007 | 00:10 buildc, 00:30 | - | - | - | >0.05 | NS |
This table lists all the values of the measured variables that relate to oxygenation and ventilation.
NS insignificant, hyphen no values given, CAL Chronic Airflow Limitation.
aExercise started at 01:00.
bDifference from controls.
cWhen the subjects entered the chamber, the environmental conditions were that of normal sea level but then were gradually made more hypoxic over the specified amount of time until the target hypoxic dose was reached.
Table 4.
Cardiovascular variables
| Outcomes | Author and year | Duration of exposure (h:min) | Hypobaric hypoxia result [mean (SD)] | Normobaric hypoxia result [mean (SD)] | Difference (HH − NH) | P value | Direction of difference NH compared to HH |
|---|---|---|---|---|---|---|---|
| Hypoxic cardiac response (HCR) bpm %−1 | |||||||
| Savourey 2007 | 00:10 builda, 00:05 | −0.61 | −0.63 | 0.02 | >0.05 | NS | |
| Savourey 2007 | 00:10 builda, 00:30 | −0.52 | −0.79 | 0.27 | >0.05 | NS | |
| LF/HF% | |||||||
| Basualto-Alarcon 2012 | 00:15 builda, 00:10 acclimatisation | 1.96 (2.6) | 1.28 (0.92) | 0.68 | <0.05 | HH > NH | |
| HR | |||||||
| Self 2011 | 00:01 | 104.9 (14.3) | 96.6 (14.6) | 8.3 | <0.05 | HH > NH | |
| Self 2011 | 00:04 | - | - | - | >0.05 | NS | |
| Basualto-Alarcon 2012 | 00:15 builda, 00:05 acclimatisation | 61 (9) | 62 (6) | −1 | - | - | |
| Basualto-Alarcon 2012 | 00:15 builda, 00:10 acclimatisation | 129 (23) | 134 (16) | −5 | <0.05 | NH > HH | |
| Savourey 2007 | 00:10 builda, 00:05 | 70.32 (9.91) | 69.62 (9.95) | >0.05 | NS | ||
| Savourey 2003 | 00:10 builda, 00:30 | - | - | - | <0.05 | HH > NH | |
| Savourey 2007 | 00:10 builda, 00:30 | 69.50 (12.07) | 70.67 (12.07) | - | >0.05 | NS | |
| Faiss 2013 | 01:00 | 62 (8) | 63 (10) | −1 | >0.05 | NS | |
| Miyagawa 2010 | 01:05 | 144 | 137 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:10 | 150 | 146 | - | >0.05 | NS | |
| Miyagawa 2010 | 01:40 | 166 | 164 | - | >0.05 | NS | |
| Tucker 1983a | 02:20 including 00:15 builda | 10.4 | 3.6 | 6.8 | - | HH > NH | |
| Faiss 2013 | 08:00 | 68 (13) | 69 (13) | −1 | >0.05 | NS | |
| Faiss 2013 | 16:00 | 61 (10) | 66 (7) | −5 | >0.05 | NS | |
| Faiss 2013 | 24:00 | 65 (9) | 71 (10) | −6 | >0.05 | NS | |
| Stroke volume | |||||||
| Miyagawa 2010 | 01:05 | 105 | 107 | −2 | >0.05 | NS | |
| Miyagawa 2010 | 01:10 | 113 | 126 | −13 | >0.05 | NS | |
| Miyagawa 2010 | 01:40 | 116 | 124 | −8 | >0.05 | NS | |
| Cardiac output | |||||||
| Miyagawa 2010 | 01:05 | 15.3 | 14.7 | >0.05 | NS | ||
| Miyagawa 2010 | 01:10 | 17.1 | 18.2 | >0.05 | NS | ||
| Miyagawa 2010 | 01:40 | 19.4 | 20.1 | >0.05 | NS | ||
| Mean BP | |||||||
| Miyagawa 2010 | 01:05 | 112 | 107 | >0.05 | NS | ||
| Miyagawa 2010 | 01:10 | 108 | 107 | >0.05 | NS | ||
| Miyagawa 2010 | 01:40 | 100 | 99 | >0.05 | NS | ||
| BP systolic (Torr) | |||||||
| Faiss 2013 | 01:00 | 124 (9) | 129 (13) | −5 | >0.05 | NS | |
| Tucker 1983b | 02:20 including 00:15 builda | −1 | 5 | −6 | NH > HH | ||
| Faiss 2013 | 08:00 | 124 (9) | 123 (7) | 1 | >0.05 | NS | |
| Faiss 2013 | 16:00 | 121 (9) | 118 (9) | 3 | >0.05 | NS | |
| Faiss 2013 | 24:00:00 | 131 (10) | 129 (9) | 2 | >0.05 | NS |
This table lists all the values of the measured variables that relate to the cardiovascular system.
NS insignificant.
aWhen the subjects entered the chamber, the environmental conditions were that of normal sea level but then were gradually made more hypoxic over the specified amount of time until the target hypoxic dose was reached.
bDifference from controls.
Table 5.
AMS and neurology variables
| Outcome | Author and year | Duration of exposure (h:min) | Hypobaric hypoxia result [mean (SD)] | Normobaric hypoxia result [mean (SD)] | Difference (HH − NH) | P value | Direction of difference NH compared to HH |
|---|---|---|---|---|---|---|---|
| Length of centre of pressure trajectory in Y axis | |||||||
| Eyes open 1,700 m | Degache 2012 | 00:30 | 114.2 (38.8) | 129.5 (53.3) | -15.3 | - | NH > HH |
| Eyes closed 1,700 m | Degache 2012 | 00:30 | 127.2 (54.9) | 87.7 (44.8) | 39.5 | - | HH > NH |
| Dual task 1,700 m | Degache 2012 | 00:30 | 128.7 (87.1) | 79.9 (30.3) | 48.8 | - | HH > NH |
| Romberg’s index 1,700 m | Degache 2012 | 00:30 | 1.35 (0.19) | 1.42 (0.34) | -0.07 | - | - |
| Eyes open 3,000 m | Degache 2012 | 00:30 | 123.1 (22.6) | 127.2 (41.5) | -4.1 | - | - |
| Eyes closed 3,000 m | Degache 2012 | 00:30 | 104.7 (27.0) | 89.1 (39.9) | 15.6 | - | HH > NH |
| Dual task 3,000 m | Degache 2012 | 00:30 | 91.9 (22.4) | 82.4 (30.4) | 9.5 | - | HH > NH |
| Romberg’s index 3,000 m | Degache 2012 | 00:30 | 1.33 (0.22) | 1.39 (0.29) | -0.06 | - | - |
| Variance of speed of CoP | |||||||
| Eyes open 1,700 m | Degache 2012 | 00:30 | 111.0 (56.2) | 151.4 (30.2) | -40.4 | - | NH > NH |
| Eyes closed 1,700 m | Degache 2012 | 00:30 | 111.0 (58.8) | 149.9 (31.5) | -38.9 | - | NH > HH |
| Dual task 1,700 m | Degache 2012 | 00:30 | 112.1 (57.7) | 151.1 (31.7) | -39 | - | NH > HH |
| Romberg’s index 1,700 m | Degache 2012 | 00:30 | 0.98 (0.09) | 0.99 (0.03) | -0.01 | - | - |
| Eyes open 3,000 m | Degache 2012 | 00:30 | 150.5 (42.3) | 160.8 (14.0) | -10.3 | - | NH > HH |
| Eyes closed 3,000 m | Degache 2012 | 00:30 | 142.9 (40.8) | 158.6 (13.7) | -15.7 | - | NH > HH |
| Dual task 3,000 m | Degache 2012 | 00:30 | 143.4 (39.2) | 160.1 (15.0) | -16.7 | - | NH > HH |
| Romberg’s index 3,000 m | Degache 2012 | 00:30 | 0.95 (0.11) | 0.99 (0.02) | -0.04 | - | - |
| Lake Louise AMS scores | Self 2011 | 00:01 | - | - | 2.36 | >0.05 | NS |
| Self 2011 | 00:04 | - | - | -4.89 | >0.05 | NS | |
| Roach 1996 | 09:00 | - | - | - | <0.01 | HH > NH | |
| Loeppky 2005* | 10:00 | - | - | - | <0.001 | HH > NH |
This table lists all the values of the measured variables that relate to AMS and neurology.
NS insignificant.
*P value calculated including hypobaric normoxia.
Table 6.
Additional physiological variables
| Outcome | Author and year | Duration of exposure (h:min) | Hypobaric hypoxia result [mean (SD)] | Normobaric hypoxia result [mean (SD)] | Difference (HH − NH) | P value | Direction of difference NH compared to HH |
|---|---|---|---|---|---|---|---|
| Exhaled NO (PE NO) | |||||||
| Hemmingsson 2009 | 00:10 at each ascending altitude | - | - | 33% mean reduction (at 5,000 m) | 0.002 | NH > HH | |
| Faiss 2013 | 01:00 | 9.5 (5.0) | 14.9 (9.2) | −5.4 | <0.01 | NH > HH | |
| Faiss 2013 | 08:00 | 8.8 (5.3) | 14.1 (7.4) | −5.3 | <0.01 | NH > HH | |
| Faiss 2013 | 16:00 | 7.9 (4.5) | 14.7 (8.6) | −6.8 | <0.01 | NH > HH | |
| Faiss 2013 | 24:00:00 | 8.9 (5.4) | 15.7 (8.7) | −5.8 | <0.01 | NH > HH | |
| RQ | |||||||
| Self 2011 | 00:05 | 2.37 (0.53) | 1.41 (0.15) | 0.96 | 0.005 | HH > NH | |
| Forced vascular conductance | |||||||
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Sweat rate | |||||||
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Oesophageal temperature thresholds for increasing forearm skin vascular conductance | |||||||
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Oesophageal temperature thresholds for increasing sweat rate | |||||||
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Oesophageal temperature | |||||||
| Miyagawa 2010 | 01:05 | 36.63 | 36.61 | 0.02 | >0.05 | NS | |
| Miyagawa 2010 | 01:10 | 37.12 | 37.11 | 0.01 | >0.05 | NS | |
| Miyagawa 2010 | 01:40 | 37.95 | 37.96 | −0.01 | >0.05 | NS | |
| Skin temperature | |||||||
| Miyagawa 2010 | 01:05 | 33.37 | 33.47 | −0.1 | >0.05 | NS | |
| Miyagawa 2010 | 01:10 | 33.35 | 33.43 | −0.08 | >0.05 | NS | |
| Miyagawa 2010 | 01:40 | 34.44 | 34.59 | −0.15 | >0.05 | NS | |
| Urine vol (ml) | |||||||
| Loeppky 2005a | 10:00 | - | - | - | 0.005 | HH > NH | |
| Tucker 1983b | 02:20 including 00:15 buildd | −1.6 | 0.1 | −1.7 | - | NH > HH | |
| Plasma volume | |||||||
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Loeppky 2005 | 10:00 | - | - | −6% | 0.002** | HH > NH | |
| GFR | |||||||
| Loeppky 2005 | 10:00 | - | - | - | >0.05 | NS | |
| Plasma K+ | |||||||
| Loeppky 2005 | 10:00 | - | - | - | 0.003 | NH > HH | |
| Plasma Na+ | |||||||
| Loeppky 2005 | 10:00 | - | - | - | 0.006 | NH > HH | |
| Plasma renin activity (PRA) | |||||||
| Loeppky 2005a | 10:00 | - | - | - | <0.05 | HH > NH | |
| Plasma aldosterone | |||||||
| Loeppky 2005a | 10:00 | - | - | - | <0.001 | NH > HH | |
| Free water clearance (CH2O) | |||||||
| Loeppky 2005a | 10:00 | - | - | - | <0.05 | HH > NH | |
| ACTH | |||||||
| Loeppky 2005a | 10:00 | - | - | - | 0.18 | NS | |
| ADH | |||||||
| Loeppky 2005 | 10:00 | - | - | - | >0.05 | NS | |
| ANP | |||||||
| Loeppky 2005* | 10:00 | - | - | - | 0.97 | NS | |
| Blood base excess | |||||||
| Loeppky 2005 | 10:00 | - | - | - | >0.05 | NS | |
| Urine Na+/K+ | |||||||
| Loeppky 2005 | 10:00 | - | - | - | 0.7 | NS | |
| Catecholamines | |||||||
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Loeppky 2005 | 10:00 | - | - | - | 0.43 | NS | |
| Haematocrit | |||||||
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Tucker 1983b | 02:20 including 00:15 buildd | 0.7 | 0.7 | 0 | NS | ||
| Haemoglobin concentration | |||||||
| Savourey 2007 | 00:10 buildd, 00:05 | - | - | - | >0.05 | NS | |
| Savourey 2007 | 00:10 buildd, 00:30 | - | - | - | >0.05 | NS | |
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Plasma osmolarity (mOsm) | |||||||
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Tucker 1983b | 02:20 including 00:15 buildd | −0.4 | −1 | 0.6 | - | HH > NH | |
| pH | |||||||
| Savourey 2003 | 00:10 buildd, 00:30 | 7.46 (SEM 0.03) | 7.44 (SEM 0.02) | 0.02 | ≤0.05 | Borderline HH > NH | |
| Savourey 2007 | 00:10 buildd, 00:30 | 7.45 (0.04) | 7.44 (0.04) | 0.01 | 0.02 | HH > NH | |
| CAL subjects | |||||||
| Naughton 1995 | 00:52 | - | - | −0.02 | >0.05 | NS | |
| Control subjects | |||||||
| Naughton 1995 | 00:52 | - | - | 0.01 | >0.05 | NS | |
| Tucker 1983d | 02:20 including 00:15 buildd | 0.047 | 0.015 | 0.032 | - | HH > NH | |
| Faiss 2013 | 24:00:00 | - | - | - | <0.01 | NH > HH | |
| Urine osmolarity (mOsm) | |||||||
| Tucker 1983c | 02:20 including 00:15 buildb | 15 | 0.1 | 14.9 | - | HH > NH | |
| Lactate mmol/kgH2O | |||||||
| Miyagawa 2010 | 01:00-01:40 | - | - | - | >0.05 | NS | |
| Blood NO metabolites | |||||||
| Faiss 2013 | 01:00 | 31.6 (19.6) | 27.7 (7.3) | 3.9 | <0.01 | HH > NH | |
| Faiss 2013 | 08:00 | 28.1 (18.9) | 32.7 (9.7) | −4.6 | <0.01 | NH > HH | |
| Faiss 2013 | 16:00 | 24.2 (16.3) | 30.2 (7.1) | −6 | <0.01 | NH > HH | |
| Faiss 2013 | 24:00 | 22.85 (16.2) | 28.9 (6.9) | −6.05 | <0.01 | NH > HH | |
| GPX (% baseline) | |||||||
| Faiss 2013 | 01:00 | 114 (26) | 111 (30) | 3 | >0.05 | NS | |
| Faiss 2013 | 08:00 | 85 (27) | 123 (23) | −37 | >0.05 | NS | |
| Faiss 2013 | 16:00 | 105 (43) | 107 (21) | −2 | >0.05 | NS | |
| Faiss 2013 | 24:00 | 103 (43) | 107 (21) | −4 | >0.05 | NS | |
| MDA (% baseline) | |||||||
| Faiss 2013 | 01:00 | 117 (40) | 92 (36) | 25 | >0.05 | NS | |
| Faiss 2013 | 08:00 | 103 (62) | 111 (35) | −8 | >0.05 | NS | |
| Faiss 2013 | 16:00 | 111 (56) | 116 (55) | −5 | >0.05 | NS | |
| Faiss 2013 | 24:00 | 108 (52) | 97 (51) | 11 | >0.05 | NS | |
| Nitrotyrosine (% baseline) | |||||||
| Faiss 2013 | 01:00 | 86 (16) | 105 (26) | −19 | >0.05 | NS | |
| Faiss 2013 | 08:00 | 77 (35) | 75 (37) | 2 | >0.05 | NS | |
| Faiss 2013 | 16:00 | 91 (20) | 98 (16) | −7 | >0.05 | NS | |
| Faiss 2013 | 24:00 | 75 (40) | 87 (25) | −12 | >0.05 | NS | |
| Plasma advanced oxidation protein products | |||||||
| Faiss 2013 | 01:00 | 120% | 13% | 107% | - | NH > HH | |
| Faiss 2013 | 24:00 | 260% | 88% | 172% | - | NH > HH | |
| Superoxide dismutase | Faiss 2013 | 24:00 | - | - | 37% | - | NH > HH |
This table lists all the values of all other the measured physiological variables.
NS insignificant, SEM Standard Error of the Mean.
aMeasured 2 h after exposure.
bDifference from controls.
cSubjects were gradually exposed increasing levels of hypoxia over the stated time until the target hypoxic dose was reached.
dWhen the subjects entered the chamber, the environmental conditions were that of normal sea level but then were gradually made more hypoxic over the specified amount of time until the target hypoxic dose was reached.
*P value calculated including hypobaric normoxia.
**P value calculated including hypobaric normoxia and after 3 h.
Review: results
Studies
A total of 225 unique articles were identified in the EMBASE, MEDLINE and Web of Science searches. After applying our inclusion and exclusion criteria, 13 studies were deemed eligible for inclusion (Table 2: Study design). A total of 153 subjects were included in our review. Of these, six subjects were women and nine had chronic airflow limitations. One study was added after snowballing [12].
The studies investigated simulated altitudes from 1,700 m to 7,620 m, and exposure to the hypoxia lasted between 5 min to 24 h. The 13 studies were carried out in seven different countries: Australia (1), Japan (1), Spain (1), Sweden (1), Switzerland (2), France (2), and America (5).
Variables measured
Quality of data
Study design
Nine studies [12-20] randomised the order of the crossover. Three studies [6,21,22] did not, and one [23] was ambiguous as to whether randomization was used or not. Three studies [12,18,22] had a washout period of at least 14 days, six studies [13-17,21] used 7 days, and four studies [6,19,20,23] used less than 24 h.
The largest study involved 20 people, the smallest 7, and the mean was 12. None of the studies stated they had conducted a sample size calculation to justify their chosen number. Only two studies [12,23] mentioned accounting for the inflated risk of type I errors that arises when multiple comparisons are made, and both of these performed Bonferroni adjustments. Only one study measured the test-retest reliability of their outcome variable [23]. They performed an intra-class coefficient correlation and showed a good reliability of the postural stability measurements.
The methods used to control the degree of hypoxia administered in each study varied. Five studies mentioned controlling the relative humidity between HH and NH. Of these, three [15,17,18] maintained 50% humidity (±1%) and two [13,14] maintained it between 45% and 55%. One paper [6] specifically mentioned the measurement and control of CO2 levels in the chambers using CO2 scrubbers.
Ventilation
Eight studies were identified that reported ventilation and oxygenation. Five of these lasted ≥1 h (long studies) [12,13,15,22,16], and three lasted <1 h (short studies) [17,18,21]. Five out of seven studies reporting minute ventilation reported values that were significantly lower in HH [12,13,17,21,22] (by up to 4 L/min) [13] (Figure 2: Graph of minute ventilation), whereas two identified no difference between conditions [15,18]. Consistent with this, the tidal volume was lower in HH in five out of six studies where this was reported (Figure 3: Graph of tidal volume) [12,17,18,21,22]. The largest difference in tidal volume found in a study was 0.9 L [12]. Two of the seven studies reporting breathing frequency found it to be higher in HH [17,18], whilst two others reported lower values in HH [21,22] and there was no difference in the remainder [12,13,15] (Figure 4: Graph of breathing frequency). The only study that reported alveolar ventilation found that it was higher in NH than HH [13].
Figure 2.
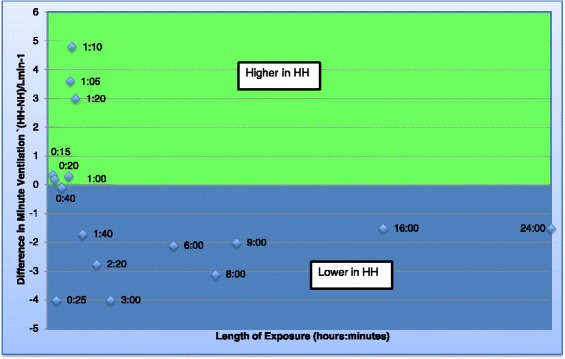
Graph of minute ventilation. Graph to show the difference in minute ventilation between the two environments over time. Each data point represents data obtained from a study and the number refers to the time point. If the data point is in the green area, the minute ventilation was found to be higher in HH but if in the blue area, the minute ventilation was found to be lower in HH.
Figure 3.
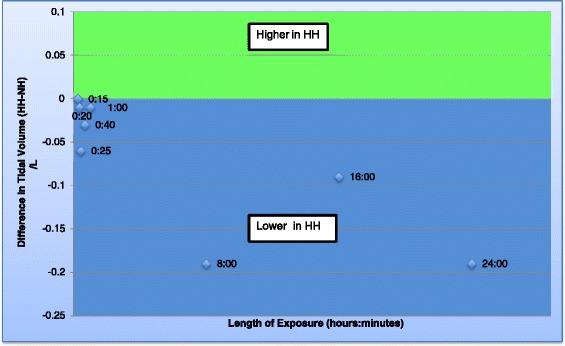
Graph of tidal volume. Graph to show the difference in tidal volume between the two environments over time. Each data point represents data obtained from a study and the number refers to the time point. If the data point is in the green area, the tidal volume was found to be higher in HH but if in the blue area, the tidal volume was found to be lower in HH.
Figure 4.
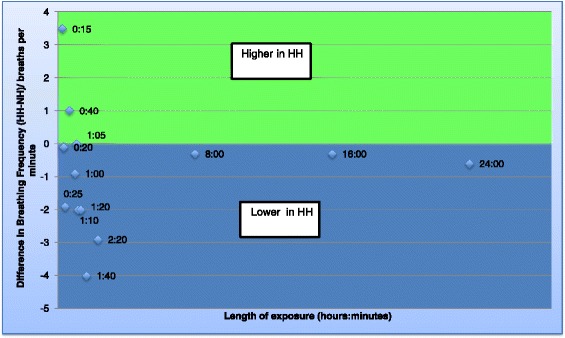
Graph of breathing frequency. Graph to show the difference in breathing frequency between the two environments over time. Each data point represents data obtained from a study and the number refers to the time point. If the data point is in the green area, the breathing frequency was found to be higher in HH but if in the blue area, the breathing frequency was found to be lower in HH.
Oxygenation
The peripheral oxygen saturations measured by pulse oximetry (SpO2) were significantly lower in HH in two out of three short studies [17,18]. One study found that the saturations were 4.08% lower in HH [18]. However, no differences were found in all five of the long studies [12-14,16,22]. The arterial blood saturations (SaO2) were lower in HH in all three short studies [6,17,18] but not in the two longer studies [15,16]. Arterial partial pressures of oxygen (PaO2) was lower in NH in one study [6], higher in NH in one study [17], and no different in two studies [18,20] (Table 3: Oxygenation and ventilation). Only the two studies by Savourey et al. [17,18] measured the end tidal fractions of O2 and these report discordant results. In 2003, Savourey et al. [17] found the end tidal fractions of O2 to be higher (P < 0.00001) in HH than NH; however, in their 2007 [18] study following the same protocols, no difference was demonstrated (P > 0.05).
Carbon dioxide (CO2) clearance
In three out of five studies, there was no difference in the end tidal partial pressure of CO2 (PETCO2) between HH and NH [17,18,22]; however, one study [12] found it to be higher in HH and one found it to be lower in HH [13]. In four out of five studies, the PaCO2 levels were lower [11,17,13] or the same [20] in HH compared to NH. For example, one study found the difference in PaCO2 to be as large as 5.9 hPa lower in HH [18]. Two studies measured the end tidal fractions of CO2 [17,18]. In 2003, Savourey et al. [17] found the end tidal fractions of CO2 to be higher (P < 0.00001) in HH than NH, whereas in the same group's 2007 study [18] (following the same protocols), no difference was demonstrated (P > 0.05).
Cardiovascular variables
Seven studies were identified that reported physiological variables relating to the cardiovascular system. Three of these were long studies [12,15,22] and four were short studies [6,17,18,21] (Table 4: Cardiovascular variables). All seven studies measured heart rate (HR). Three found HR to be higher in HH [6,17,22], one found it to be lower [21], and the others found no differences [12,15,18] (Figure 5: Graph of heart rates). Three studies measured blood pressure. Two found no difference in blood pressure between environments [12,15]; however, one other found it to be lower in HH than in NH [22]. Sympathetic drive, measured by a specific electrocardiogram (ECG) recorder of low- and high-frequency components of heart rate, was only investigated in one study [21] and was found to be higher in HH than in NH.
Figure 5.
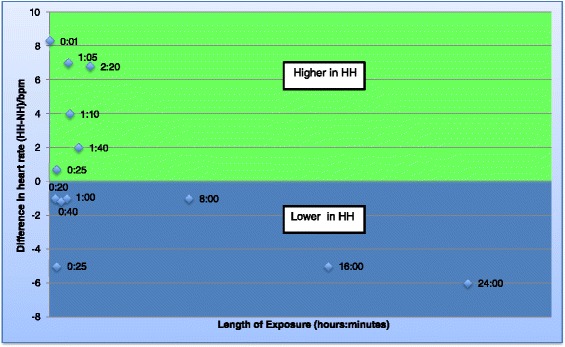
Graph of heart rates. Graph to show the difference in heart rate between the two environments over time. Each data point represents data obtained from a study and the number refers to the time point. If the data point is in the green area, the heart rate was found to be higher in HH but if in the blue area, the heart rate was found to be lower in HH.
Acute mountain sickness and neurology
Four studies were found relating to acute mountain sickness (AMS) and neurological symptoms and signs, two of which were long studies [14,16] and two of which were short studies [6,23] (Table 5: AMS and neurology). Two out of three studies [14,16] that measured AMS scores found that AMS was significantly worse in HH than in NH. Only one study [23] measured postural stability, which was significantly reduced in HH compared to NH. Subjects deviated from the midline more in HH than NH, and the speed with which movements occurred to correct their posture was slower.
Additional physiological variables
Six studies [6,12,14,15,19,22] researched a variety of other physiological variables (Table 6: Additional physiological variables). In three out of five of these studies [17,18,22], the plasma pH was higher in HH than NH; however, one of the other papers found the pH to be higher in NH [12] (Figure 6: Graph of pH). The greatest difference in pH found in a study was 0.032 [22]. Urine osmolarity was measured in only one study and was significantly higher in HH [22]. Additionally, the same study was the only one to measure the volume of urine produced and found it to be lower during HH exposure [22]. One study also measured K+, Na+ and aldosterone concentrations in plasma and found they were higher in NH [14]. In recovery post-hypoxic exposure, authors in [14] found that the urine volume, plasma renin activity and free water clearance were higher whilst aldosterone was lower after HH than NH. This was the only study to measure these variables.
Figure 6.

Graph of pHs. Graph to show the difference in pH between the two environments over time. Each data point represents data obtained from a study and the number refers to the time point. If the data point is in the green area, the pH was found to be higher in HH but if in the blue area, the pH was found to be lower in HH.
Only two studies [12,19] measured exhaled NO levels and both found that they were lower in HH than in NH. The greatest difference in exhaled NO found in these papers was 6.8 mmHg [12]. Additionally, only one study measured systemic NO and the authors found it was lower in HH [12]. The same study was the only study to measure plasma-advanced oxidation protein products and superoxide dismutase. These were higher in HH [12]. Only one study [6] measured the respiratory quotient (RQ) and found it was higher in HH than NH.
Review: discussion
Summary of physiological findings
We believe this is the first systematic review comparing the effect of NH and HH on human physiology. Significant differences were demonstrated in variables relating to ventilation, NO, fluid retention, and in factors relating to AMS. For other variables, there was no consistent pattern across the reviewed studies.
Oxygenation and ventilation parameters
The majority of studies included data relating to oxygenation and ventilation. Oxygen saturations measured from pulse oximetry (SpO2) and arterial blood gases correlated with each other. During short-term exposure, oxygen saturations were lower in HH [6,17]. This was not the case for long-term exposure, perhaps as the initial process of acclimatisation to hypoxia had occurred [18,20]. PaO2 did not differ at any stage.
A decreased minute ventilation and alveolar ventilation was seen in HH. This finding is in accord with the smaller tidal volumes demonstrated in HH; however, breathing frequencies varied [12,13,17,18,21,22]. Despite the lower SpO2 in HH initially, oxygen saturations were maintained at the same level in both environments over longer time periods. Interestingly, despite the reduction in ventilation, PETCO2 levels did not change significantly [12,13,17,18,22].
Notably, Savourey et al. found different results in end tidal fractions of both O2 and CO2 between their two studies in 2003 [17] and 2007 [18]. This not only makes interpretation of their results very challenging but also highlights issues concerning reliability of studies (as discussed in Study quality section).
Cardiovascular variables
The majority of these variables were similar for HH and NH. Heart rate and blood pressure results were inconsistent, although some data suggests that heart rate may be raised initially in HH [6,12,15,17,18,21,22].
AMS and neurology variables
Mountain sickness is an area of research that has attracted much interest and consequently the multiple variables that combine to make the Lake Louise AMS scoring system are commonly collected in these studies. In two of four studies, AMS severity was found to increase in HH compared with NH [14,16]. Furthermore, in the one study that found no differences between environments [6], participants only had a 5-min exposure time, as opposed to 9- and 10-h exposures in other studies. Additionally, postural stability was significantly worse in HH [23]. The authors suggest that visual contrast sensitivity is lower in HH than in NH and that this may contribute to the postural stability [23].
Other homeostatic variables
Plasma pH appears to be higher in HH than NH [17,18,22]. The study [20] that did not find significant differences in pH between the two conditions was conducted at 1,829 and 2,438 m whereas the elevated pH values were from 4,500 m, suggesting that the differences between the two conditions may be more pronounced at higher altitudes. Elevated pH in HH is surprising in the context of the finding that ventilation is reduced under these conditions.
Exhaled and systemic NO levels were lower in HH [12,19]. Faiss et al. [12] found increased oxidative stress in HH and attributed the systemic differences in NO bioavailability to this.
Many other physiological variables were measured; however, most of these were reported in only a single study. Thus, it is difficult to make conclusions without verification from other studies, and we have not considered these further.
Study quality
The search results reveal several issues relating to study design. Very few studies state the reliability of their measurements or performed a sample size calculation. It is therefore difficult to evaluate if they are adequately powered to identify a real difference between conditions, should such a difference exist. Given that differences in physiological responses between NH and HH conditions are likely quite small, large sample sizes would likely be required to avoid type II (false negative) errors. Additionally, by performing statistical analysis on a large number of variables over many time points, the risk of type I (false positive) errors increases.
Failure to account for the PH2O leads to an overestimate of the hypoxic dose in NH, such that incorrect partial pressure of inspired oxygen (PiO2) may be attained [11]. In one study [6], a NH exposure equivalent to 7,620 m was described; however, the conditions were in fact closer to 7,010 m once pH2O was accounted for [11]. We have emphasised these differences by calculating, where possible, the PiO2 in the different hypoxic conditions (Table 2: Study design). We found the differences in PiO2 to be as much as 4 mmHg. The level of CO2 in the test environment was a potential source of error. Basualto-Alarcon et al. [21] highlight this issue in stating that different gas inflow rates into each hypoxic system allow different levels of CO2 accumulation. Additionally, they state that their NH environment may have been more hypercapnic because it had half the total volume of the hypobaric chamber. These control group contrast issues will either enhance or diminish the effect size and therefore the difference between HH and NH. This may be of particular relevance to ventilator variables.
Mechanisms for results
Many hypothetical mechanisms have been proposed for the effect of low barometric pressure on physiology. These include intravascular bubble formation, increased alveolar deadspace, altered fluid permeability, changes in chemosensitivity, and a mismatch in ventilation and perfusion [13,16,17]. Although pressure may be the principle confounder between the two scenarios, we must also reflect that other factors may differ between HH and NH, thus impacting participant's physiology. For example, the laboratory-based components of the studies reviewed were performed between 22°C and 25°C, a temperature likely to be far warmer that experiences at 4,000 m in a field laboratory. Such differences in ambient temperature may alter physiological mechanisms such as the degree of peripheral vasoconstriction, NO metabolism or the production of reactive oxidative species [24].
As highlighted, the duration of the hypoxic exposure impacts on the results obtained. Different physiological systems will have different response rates for adaptation to hypoxia [25]. For some physiological parameters, the short study durations may not be long enough for differences between NH and HH to be elicited. Studies reporting repeated measures over time provide a window on this phenomenon. For example, in the 1997 study by Loeppky et al. [13], where no differences in minute ventilation were reported after 30 or 60 min of hypoxic exposure, significant differences were evident after 3 h of exposure. Additionally, Savourey et al. [17,18] initially found lower PETO2 and PETCO2 in HH than NH but then no difference in prolonged exposure. This may be because during HH exposure, the ambient partial pressure of nitrogen (PN2) is initially lower than the body's and therefore nitrogen (N2) initially diffuses from the tissues to the alveoli [5]. Until this equilibrium is achieved, the arterial oxygen content, PAO2, and the arterial carbon dioxide content, PACO2, are lowered as a result of the relatively higher PAN2 in HH than NH.
Loeppky et al. [13] also suggests that an initial increase in CO2 produced in HH compared to NH might be due to microbubble formation similar to the nitrogen bends in divers. This emphasises the importance of study duration on physiological response and the problems inherent in comparing studies of different hypoxia exposure times.
If there are indeed differences between HH and NH, at what equivalent altitude do they become apparent? Most of the studies have been carried out at 4,500 m (or equivalent), but Naughton et al.'s study [20] performed at 1,829 and 2,438 m was unable to find any significant differences between HH and NH. These altitudes correspond to PO2 values of 118 mmHg (15.7 kPa) and 108 mmHg (14.4 kPa) [25] respectively, and it is possible that these altitudes were not high enough to elicit differences in the measured variables. Significant differences between the effects of NH and HH may impact the interpretation and application of results from studies at high altitude where the change in pressure may be a confounding influence in the evaluation of physiological responses to high altitude.
Strengths and limitations of this study
Although this is the first systematic review to summarise crossover studies comparing physiological responses to hypobaric and normobaric hypoxia, other publications have come to similar conclusions on the topic. Millet et al. [10] stress the importance of disentangling hypoxia and hypobaria and Fulco et al. [26] highlight the need for further investigations into NH versus HH, for particular application to pre-acclimatisation strategies.
The strengths of this systematic review include the clear research question, comprehensive search strategy and consistent methods used for identifying eligible manuscripts and extracting data. Limitations of this review include the focus on crossover studies but are predominantly related to the quantity and quality of the underlying literature. There are few studies that compare HH and NH and the number of participants in each study is small. Whilst several of these studies report interesting differences between HH and NH, there is marked inconsistency in the reported results. This may be due to a number of other factors including heterogeneity of study design, duration and magnitude of hypoxic dose and outcome reporting. Furthermore, the reporting of multiple phenotypes in each study without correction for repeat testing may be associated with an increased likelihood of type 1 errors. Conversely, the small sample sizes may be associated with an increased likelihood of type 2 errors.
As mentioned, the studies were heterogeneous by design. For example, they differed in regard to the subjects' prior exposure to altitude. In two of the studies reviewed [16,22], the subjects lived between 1,500 and 1,600 m and so may have been partially acclimatised to high altitude. It is not clear whether the same effects would be seen in partially and not acclimatised subjects.
Finally, the self-reported nature of AMS scores could be associated with inconsistent responses from participants. In the study by Self et al. [6], there was a disparity between post-hypoxia interview responses and the responses during hypoxic exposure. There is no gold standard method for these types of studies and so there is much variability due to the methodology employed.
Conclusions
We present an overview of the current available literature regarding crossover studies relating to the different effects of HH and NH on human physiology. This systematic review is the first to compare the effects of a NH and HH environment on human physiology. Several studies reported a number of variables (e.g. minute ventilation and NO levels) that were different between the two conditions, lending support to the notion that true physiological difference are indeed present. However, the presence of confounding factors such as time spent in hypoxia, temperature, and humidity, and the limited statistical power due to small sample sizes, limit the conclusions that can be drawn from these findings.
Standardisation of study methods and reporting may aid interpretation of future studies and thereby improve the quality of data in this area. This is important to improve the quality of data that is used both for improving understanding of hypoxia tolerance, both at altitude and in the clinical setting.
Acknowledgements
We would like to thank Caroline De Brun.
Abbreviations
- ACTH
Adrenocorticotropic hormone
- ADH
Anti-diuretic hormone
- AMS
Acute mountain sickness
- ANP
Anti-naturetic protein
- Bf
Respiratory rate
- BP
Blood pressure
- CO2
Carbon dioxide
- CO
Cardiac output
- ECG
Electrocardiogram
- FetCO2
End tidal fraction of carbon dioxide
- FetO2
End tidal fraction of oxygen
- exNO
Exhaled nitric oxide levels
- GFR
Glomerular filtration rate
- GPX
Glutathione peroxidase
- Hb conc
Haemoglobin concentration
- Hct
Haematocrit
- HR
Heart rate
- LF/HF
Heart rate variability
- pH2O
Humidity
- HH
Hypobaric hypoxia
- HCR
Hypoxic cardiac response
- HVR
Hypoxic ventilatory response
- NO
Nitric oxide
- N2
Nitrogen
- NH
Normobaric hypoxic
- SpO2
Pulse oximetry
- PN2
Partial pressure of nitrogen
- PO2
Partial pressure of oxygen
- PaCO2
Arterial partial pressures of carbon dioxide
- PaO2
Arterial partial pressures of oxygen
- PetO2
End tidal partial pressure of oxygen
- PETCO2
End tidal partial pressure of CO2
- Plasma K+
Plasma potassium concentration
- plasma Na+
Plasma sodium concentration
- RQ
Respiratory quotient
- SaO2
Arterial blood saturations
- SV
Stroke volume
- urine Na+/K+
Urine sodium-potassium ratio
- VA
Alveolar ventilation
- VCO2
Volume of CO2 produced
- VE
Ventilation
- VO2
Volume of oxygen consumed
- VT
Tidal volume
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
EGK, PH and MG conceived the idea of the review. JC, EGK and PH planned the methodology and JC carried out the systematic review of the literature. The results were then checked by EGK and all authors discussed them. The review was drafted by JC and then commented on by all authors. All authors read and approved the final manuscript.
Contributor Information
Jonny Coppel, Email: jonny.coppel@gmail.com.
Philip Hennis, Email: p.hennis@ucl.ac.uk.
Edward Gilbert-Kawai, Email: edgilbert82@hotmail.com.
Michael PW Grocott, Email: mike.grocott@soton.ac.uk.
References
- 1.Grocott M, Montgomery H, Vercueil A. High-altitude physiology and pathophysiology: implications and relevance for intensive care medicine. Crit Care. 2007;11:203. doi: 10.1186/cc5142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Grant S, MacLeod N, Kay JW, Watt M, Patel S, Paterson A, et al. Sea level and acute responses to hypoxia: do they predict physiological responses and acute mountain sickness at altitude? Brit J Sport Med. 2002;36:141–146. doi: 10.1136/bjsm.36.2.141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Millet GP, Faiss R, Brocherie F, Girard O. Hypoxic training and team sports: a challenge to traditional methods? Brit J Sport Med. 2013;47:6–7. doi: 10.1136/bjsports-2013-092793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.DeHart RLE. Fundamentals of aerospace medicine. 2. Baltimore: Williams and Wilkins; 1996. pp. 90–91. [Google Scholar]
- 5.Conkin J, Wessel JH. Critique of the equivalent air altitude model. Aviat Space Envir Md. 2008;79:975–982. doi: 10.3357/ASEM.2331.2008. [DOI] [PubMed] [Google Scholar]
- 6.Self DA, Mandella JG, Prinzo OV, Forster EM, Shaffstall RM. Physiological equivalence of normobaric and hypobaric exposures of humans to 25,000 feet (7620 m) Aviat Space Envir Md. 2011;82:97–103. doi: 10.3357/ASEM.2908.2011. [DOI] [PubMed] [Google Scholar]
- 7.Debevec T, Millet GP. Discerning normobaric and hypobaric hypoxia: significance of exposure duration. J Appl Physiol. 2014;116:1255–5. doi: 10.1152/japplphysiol.00873.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Olivier G, Koehle MS, Guenette JA, Verges S, Chapman RF, Conkin J, et al. Comments on point: counterpoint: hypobaric hypoxia induces/does not induce different responses from normobaric hypoxia. J Appl Physiol. 2012;112:1788–1794. doi: 10.1152/japplphysiol.00356.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Muza SR. Military applications of hypoxic training for high-altitude operations. Med Sci Sports Exerc. 2007;39:1625–1631. doi: 10.1249/mss.0b013e3180de49fe. [DOI] [PubMed] [Google Scholar]
- 10.Millet GP, Faiss R, Pialoux V. Evidence for differences between hypobaric and normobaric hypoxia is conclusive. Exerc Sport Sci Rev. 2013;41:133. doi: 10.1097/JES.0b013e318271a5e1. [DOI] [PubMed] [Google Scholar]
- 11.Conkin J. PH2O and simulated hypobaric hypoxia. Aviat Space Envir Md. 2011;82:1157–1158. doi: 10.3357/ASEM.3145.2011. [DOI] [PubMed] [Google Scholar]
- 12.Faiss R, Pialoux V, Sartori C, Faes C, Deriaz O, Millet GP. Ventilation, oxidative stress, and nitric oxide in hypobaric versus normobaric hypoxia. Med Sci Sports Exerc. 2013;45:253–260. doi: 10.1249/MSS.0b013e31826d5aa2. [DOI] [PubMed] [Google Scholar]
- 13.Loeppky JA, Icenogle M, Scotto P, Robergs R, HinghoferSzalkay H, Roach RC. Ventilation during simulated altitude, normobaric hypoxia and normoxic hypobaria. Resp Physiol. 1997;107:231–239. doi: 10.1016/S0034-5687(97)02523-1. [DOI] [PubMed] [Google Scholar]
- 14.Loeppky JA, Roach RC, Maes D, Hinghofer-Szalkay H, Roessler A, Gates L, et al. Role of hypobaria in fluid balance response to hypoxia. High Alt Med Biol. 2005;6:60–71. doi: 10.1089/ham.2005.6.60. [DOI] [PubMed] [Google Scholar]
- 15.Miyagawa K, Kamijo Y, Ikegawa S, Goto M, Nose H. Reduced hyperthermia-induced cutaneous vasodilation and enhanced exercise-induced plasma water loss at simulated high altitude (3,200 m) in humans. J Appl Physiol. 2011;110:157–165. doi: 10.1152/japplphysiol.00950.2010. [DOI] [PubMed] [Google Scholar]
- 16.Roach RC, Loeppky JA, Icenogle MV. Acute mountain sickness: increased severity during simulated altitude compared with normobaric hypoxia. J Appl Physiol. 1996;81:1908–1910. doi: 10.1152/jappl.1996.81.5.1908. [DOI] [PubMed] [Google Scholar]
- 17.Savourey G, Launay JC, Besnard Y, Guinet A, Travers S. Normo- and hypobaric hypoxia: are there any physiological differences? Eur J Appl Physiol. 2003;89:122–126. doi: 10.1007/s00421-002-0789-8. [DOI] [PubMed] [Google Scholar]
- 18.Savourey G, Launay JC, Besnard Y, Guinet-Lebreton A, Alonso A, Sauvet F, et al. Normo or hypobaric hypoxic tests: propositions for the determination of the individual susceptibility to altitude illnesses. Eur J Appl Physiol. 2007;100:193–205. doi: 10.1007/s00421-007-0417-8. [DOI] [PubMed] [Google Scholar]
- 19.Hemmingsson T, Linnarsson D. Lower exhaled nitric oxide in hypobaric than in normobaric acute hypoxia. Resp Physiol Neurobi. 2009;169:74–77. doi: 10.1016/j.resp.2009.08.006. [DOI] [PubMed] [Google Scholar]
- 20.Naughton MT, Rochford PD, Pretto JJ, Pierce RJ, Cain NF, Irving LB. Is normobaric simulation of hypobaric hypoxia accurate in chronic air-flow limitation. Am J Resp Crit Care. 1995;152:1956–1960. doi: 10.1164/ajrccm.152.6.8520762. [DOI] [PubMed] [Google Scholar]
- 21.Basualto-Alarcon C, Rodas G, et al. Cardiorespiratory parameters during submaximal exercise under acute exposure to normobaric and hypobaric hypoxia. Apunts Med Esport. 2012;47:65–72. doi: 10.1016/j.apunts.2011.11.005. [DOI] [Google Scholar]
- 22.Tucker A, Reeves JT, Robertshaw D, Grover RF. Cardiopulmonary response to acute altitude exposure—water loading and denitrogenation. Resp Physiol. 1983;54:363–380. doi: 10.1016/0034-5687(83)90079-8. [DOI] [PubMed] [Google Scholar]
- 23.Degache F, Larghi G, Faiss R, Deriaz O, Millet G. Hypobaric versus normobaric hypoxia: same effects on postural stability? High Alt Med Biol. 2012;13:40–45. doi: 10.1089/ham.2011.1042. [DOI] [PubMed] [Google Scholar]
- 24.Isa KBM, Kawasaki N, Ueyama K, Sumii T, Kudo S. Effect of cold exposure and shear stree on endothelial nitric oxide synthase activation. Biochem Biophys Res Commun. 2011;412:318–322. doi: 10.1016/j.bbrc.2011.07.092. [DOI] [PubMed] [Google Scholar]
- 25.West JB, Milledge JS, Schoene RB, Luks A. High altitude medicine and physiology. 5. Boca Raton: CRC Press/Taylor & Francis Group; 2013. [Google Scholar]
- 26.Fulco CS, Beidleman BA, Muza SR. Effectiveness of preacclimatization strategies for high-altitude exposure. Exerc Sport Sci Rev. 2013;41:55–63. doi: 10.1097/JES.0b013e31825eaa33. [DOI] [PubMed] [Google Scholar]