

Abstract
Background
Human metapneumovirus (HMPV) is a leading cause of acute respiratory tract infection, with significant morbidity and mortality. No licensed vaccines or therapeutic agents exist. Monoclonal antibodies (mAbs) are effective at preventing other infectious diseases and could be used against HMPV in high-risk hosts.
Methods
In vitro assays were performed to assess the neutralizing activity and affinity kinetics of human mAb 54G10. A new mouse model was developed to assess prophylactic and therapeutic efficacy in vivo. The epitope of 54G10 was identified by generating mAb-resistant mutants (MARMs).
Results
At low concentrations, 54G10 neutralized all 4 subgroups of HMPV in vitro and had subnanomolar affinity for the fusion protein. DBA/2 mice were permissive for all 4 HMPV subgroups, and 54G10 was effective both prophylactically and therapeutically against HMPV in vivo. Sequencing of HMPV MARMs identified the 54G10 epitope, which was similar to an antigenic site on respiratory syncytial virus (RSV). 54G10 also exhibited in vitro neutralizing activity and in vivo protective and therapeutic efficacy against RSV.
Conclusions
Human mAb 54G10 has broad neutralizing activity against HMPV and could have prophylactic and therapeutic utility clinically. The conserved epitope could represent a structural vaccine target for HMPV and RSV.
Keywords: human metapneumovirus, monoclonal antibody, paramyxovirus, respiratory syncytial virus, DBA/2 mouse
Human metapneumovirus (HMPV), a paramyxovirus identified in 2001, is a leading cause of acute respiratory disease worldwide [1, 2]. In high-risk populations, HMPV causes significant morbidity and mortality [3, 4]. No vaccines or therapeutics specifically target HMPV, and nonspecific therapies such as ribavirin and intravenous immunoglobulin have limited efficacy [5]. HMPV has 4 subgroups, which vary in prevalence but cause similar disease [6, 7]. Although the subgroups are relatively conserved, recurrent infection occurs throughout life [8]. Reinfection likely occurs because of limited cross-protective antibodies [9] and T-cell exhaustion causing a poor memory response [10].
All paramyxoviruses have class I viral fusion (F) proteins, which exist in a pre- and postfusion state and are required for entry [11]. HMPV vaccine studies have focused on the F protein, which is immunogenic and protective in animal models [12, 13]. The F protein is highly conserved among HMPV subgroups, with >95% amino acid similarity between A and B groups and >97% similarity within a single group, suggesting that a broadly neutralizing antibody response could be evoked if a shared epitope were targeted [14].
Monoclonal antibodies (mAbs) are effective at preventing infectious diseases in high-risk populations [15]. However, the use of humanized or chimeric mAbs, both partly murine, is associated with the development of host antibodies to foreign antigens and adverse events [16, 17]. Fully human mAbs are less immunogenic [18]. Researchers have described mAbs effective at neutralizing all 4 subgroups of HMPV in vitro [19, 20], but these had limited therapeutic efficacy in rodents [21].
We hypothesized that a human mAb directed at HMPV F protein could be used to abate infection. Here, we describe a fully human mAb, 54G10, which potently neutralized all 4 subgroups of HMPV in vitro and exhibited subnanomolar affinity for the F protein. To test in vivo efficacy, we established a new mouse model permissive for all 4 HMPV subgroups in the upper and lower airways. 54G10 decreased peak HMPV titer below the limit of detection (LOD) in both prophylactic and therapeutic experiments. Sequencing of mAb-resistant mutants (MARMs) identified that 54G10's epitope is a conserved region among paramyxoviruses [22]. Confirming this, 54G10 neutralized respiratory syncytial virus (RSV), another leading cause of respiratory infections. Thus, 54G10 is a highly effective human mAb, which recognizes a similar epitope on HMPV and RSV.
METHODS
Cells and Viruses
LLC-MK2 cells (ATCC CCL-7) were maintained in either OptiMEM (Life Technologies) with 2% fetal bovine serum (FBS) or Dulbecco's modified Eagle's medium (Corning) with 10% FBS, both with amphotericin B, L-glutamine, and gentamicin. HEp-2 cells (kindly provided by Dr Stokes Peebles) were maintained in Eagle's minimal essential medium (EMEM; Corning) with 10% FBS, amphotericin B, and gentamicin (10% EMEM). Suspension 293-F cells were maintained in 293 Freestyle media (Life Technologies). A representative strain of each HMPV subgroup was selected from clinical isolates: TN/96-12 (subgroup A1), TN/94-49 (subgroup A2), TN/98-242 (subgroup B1), and TN/89-515 (subgroup B2) [6]. Stock virus was propagated on LLC-MK2 cells [23]. The A1, B1, and B2 isolates were sucrose purified [24]. RSV A2 (kindly provided by Dr Stokes Peebles) was propagated on HEp-2 cells [25].
Monoclonal and Polyclonal Antibodies
CD22+ B cells were isolated by immunomagnetic positive selection (Miltenyi) from donor peripheral blood mononuclear cells and stimulated for 5 days with CpG and recombinant human interleukin 2 (IL-2). Cells were negatively immunomagnetically selected with antibodies against immunoglobulin M (IgM), immunoglobulin D, and immunoglobulin A, Epstein-Barr virus transformed, and seeded at 20 cells/well onto irradiated, IL-2–supplemented feeder cells. Two weeks later, supernatants were tested by enzyme-linked immunosorbent assay (ELISA) against recombinant HMPV B2FΔTM protein. Variable-heavy and variable-light chain genes were cloned from oligoclonal B-cell cultures corresponding to antigen-reactive wells, and specificity of the cloned antibodies was confirmed by ELISA [12]. Palivizumab (MedImmune) was obtained from the Monroe Carell Jr. Children's Hospital at Vanderbilt pharmacy. DS7 is a human mAb against HMPV F [20]. Expression plasmids encoding 54G10 or DS7 were transfected as described previously [12].
Neutralization Assays
Virus was incubated at room temperature (HMPV) or on ice (RSV) with 100 µL of antibody diluted in OptiMEM (HMPV) or EMEM (RSV) for 1 hour. Assays were performed using 2-fold (for HMPV) or 4-fold (for RSV) dilutions of mAb. Virus-antibody complexes and an untreated control (virus plus medium) were inoculated onto LLC-MK2 cells (HMPV) or HEp-2 cells (RSV), adsorbed for 1 hour at room temperature, overlaid with 0.75% methylcellulose in either OptiMEM with 5 µg/mL trypsin–ethylenediaminetetraacetic acid (HMPV) or 10% EMEM (RSV), incubated at 37°C with 5% CO2 for 4 days (HMPV) or 5 days (RSV), and titered [20, 25]. Serum neutralization assays were similarly performed using 4-fold dilutions.
Immunofluorescent Assay (IFA)
LLC-MK2 or Hep-2 cells were infected with HMPV A2 or RSV A2, respectively, at a multiplicity of infection of 1. After 24 hours, cells were fixed and blocked [20]. Cells were incubated with 54G10 and secondary AlexaFluor 546-conjugated goat anti-human IgG, both diluted 1:1000 in 5% powdered milk in PBS-0.05% Tween (PBS-T-milk) for 1 hour at 37°C. Images were captured using an Axiovert 200 fluorescence microscope and converted to black and white in Adobe Illustrator CS4.
ELISA
Immulon II-HB plates were coated with 100 ng/well of B2FΔTM in PBS. Wells were blocked with PBS-T-milk and then incubated with sera in 2-fold dilutions in PBS-T-milk followed by horseradish peroxidase–conjugated goat anti-human immunoglobulin diluted 1:1000 in PBS-T-milk. All incubations were performed at room temperature for 1 hour and washed with PBS-T between steps. Pierce Ultra-TMB was added for 15 minutes, the reaction was stopped with 2 M sulfuric acid, and absorbance was measured at 450 nm. Two-fold dilutions of 54G10 were used to create a linear regression formula by which the 54G10 concentration in animal sera was determined.
Kinetic Analysis
Equilibrium dissociation constant (KD) values were measured with bio-layer interferometry, using an Octet Red96 instrument (ForteBio). Streptavidin biosensors were loaded with biotinylated B2FΔTM at a concentration of 20 ng/µL in PBS for 5 minutes. Coated biosensors were blocked for 5 minutes in a 25-ng/µL solution of biocytin and washed for 5 minutes in PBS. Biosensors were incubated with varying concentrations of 54G10 (0.12 nM–500 nM) for 5 minutes to measure association (Kon) and were subsequently moved to a buffer-only well for 5 minutes to measure dissociation (Koff). The KD was calculated by the ForteBio software.
Animal Experiments
Six-week-old female DBA/2 and BALB/c mice were purchased from Jackson Laboratories and fed a standard diet and water ad libitum. Animals were housed in specific-pathogen-free conditions. The Vanderbilt Animal Care Committee approved the protocol. Animals were anesthetized intraperitoneally with ketamine/xylazine before any intranasal administration. Virus (105 plaque-forming units [PFU] for all animal experiments), drugs, and controls were given in 100-µL volumes [23]. Sera were collected by submandibular venipuncture before and 24 hours after 54G10 administration. Animals were euthanized with CO2, and nasal turbinates (NTs) and lungs were harvested [23]. The LOD was determined by the following formula: minimum detectable PFU/mL × [1/(average weight of processed specimens/2)]. For histopathologic analysis, the left lower lobe of the lung was removed and inflated with 10% formalin. Specimens were fixed, paraffin embedded, stained, and analyzed [10].
Generation of MARMs
TN/94-49 was adsorbed onto LLC-MK2 cells in the presence of 5 times the 50% inhibitory concentration (IC50) of 54G10. After 1 hour, fresh medium containing 10 times the IC50 of 54G10 was added. Cells were harvested when syncytia appeared (4–7 days). The supernatant was passed to a fresh monolayer, and the process was repeated with increasing concentrations of 54G10 (maximum, 15 times the IC50). An in vitro neutralization assay was performed to confirm 54G10 resistance, and the MARM was thrice plaque purified. Last, plaque picks were placed on a monolayer to grow a stock of purified MARM. Plaque pick resistance was confirmed.
Reverse-Transcription Polymerase Chain Reaction (RT-PCR) Amplification and Sequencing of MARMs
The F genes of 4 MARMs plaque purified from different parent wells were amplified by RT-PCR as described elsewhere [24]. PCR reactions were run on a 1% agarose gel, purified using the Qiaquick Gel Extraction Kit (Qiagen), and sequenced directly, as well as cloned into pGEM (Promega) and sequenced. Sequences were analyzed in MacVector12.0.6 (MacVector).
Generation of Mutant F Protein
Soluble B2F protein lacking the transmembrane region and cytoplasmic domain (B2FΔTM) was generated [12, 20, 26]. Site-directed mutagenesis using the QuikChange II Kit (Agilent Technologies) was performed to create 3 mutated B2FΔTM proteins containing either a valine to glycine substitution at amino acid 397, a glutamine to histidine substitution at amino acid 434, or both mutations. Mutations were confirmed by sequencing. Mutant F proteins were generated and purified from 293-F cells [12, 20, 26].
Statistical Analysis
For in vitro neutralization assays, the percentage neutralization for each antibody dilution was calculated with respect to findings for the untreated control. Dilutions were log10 transformed and analyzed by nonlinear regression with a sigmoidal dose-response curve to determine the IC50. Virus titer from animal experiments was log10 transformed. If no virus was detected by plaque assay, the number representing the LOD was used. Mean values from each group were analyzed by 1-way analysis of variance (ANOVA) with post-hoc analysis. P values of <.05 were considered statistically significant. All statistics were performed using Prism 6 (GraphPad).
RESULTS
54G10 Neutralizes HMPV In Vitro
ELISA revealed that human mAb 54G10 bound to the HMPV B2FΔTM protein (data not shown). To assess neutralization, the IC50 of 54G10 against HMPV was calculated. 54G10 neutralized all 4 subgroups of HMPV, with an IC50 of 90 ng/mL for A1, 400 ng/mL for A2, 210 ng/mL for B1, and 60 ng/mL for B2.
54G10 Binds to HMPV F Protein With High Affinity
The affinity of 54G10 to HMPV F protein was determined (Supplementary Figure 1A). 54G10 had a Kon of 1.3 × 106 M−1 s−1 and a Koff of 3.6 × 10−4 s−1, for a KD of 2.7 × 10−10 M. The subnanomolar KD indicates high-affinity binding. Palivizumab, an RSV F protein–specific mAb, did not bind (Supplementary Figure 1B).
DBA/2 Mice Are Permissive for HMPV
To establish the kinetics of HMPV infection in DBA/2 mice, animals were infected intranasally with HMPV A2 and euthanized on multiple days after infection. The titer of HMPV in the NTs and lungs was determined. In the NTs, replication peaked on day 2, with a mean titer of 1.8 × 104 PFU/g, and most mice cleared the virus by day 10 (Figure 1A). In the lungs, virus titer peaked on day 5, at a mean of 5 × 104 PFU/g (Figure 1B), and was cleared by day 10. No significant weight change was noted (data not shown).
Figure 1.

Replication kinetics of human metapneumovirus (HMPV) in the DBA/2 mouse. HMPV A2 virus titer peaks on day 2 in the nasal turbinates (NTs; A) and on day 5 in the lungs (B). DBA/2 mice are permissive for all 4 HMPV subgroups in the NTs (C) and lungs (D). Dashed lower lines represent the limit of detection. Solid lines within groups represent mean values. Abbreviation: PFU, plaque-forming unit.
To assess the replication of all HMPV subgroups, mice were infected with each HMPV subgroup and euthanized on day 5. For all subgroups, virus titer was at least 10-fold above the LOD in the NTs (Figure 1C) and >100-fold higher than the LOD in the lungs (Figure 1D), demonstrating that DBA/2 mice are permissive for all 4 HMPV subgroups. Increasing virus inoculum did not increase virus titer or cause weight loss (data not shown). Histopathologic analysis of lungs obtained from HMPV-infected mice at multiple time points revealed a mononuclear infiltrate with peribronchiolitis and perivasculitis, compared with PBS-inoculated mice (Figure 2A). T-cell infiltrates peaked on day 7, with a perivascular distribution (Figure 2B).
Figure 2.
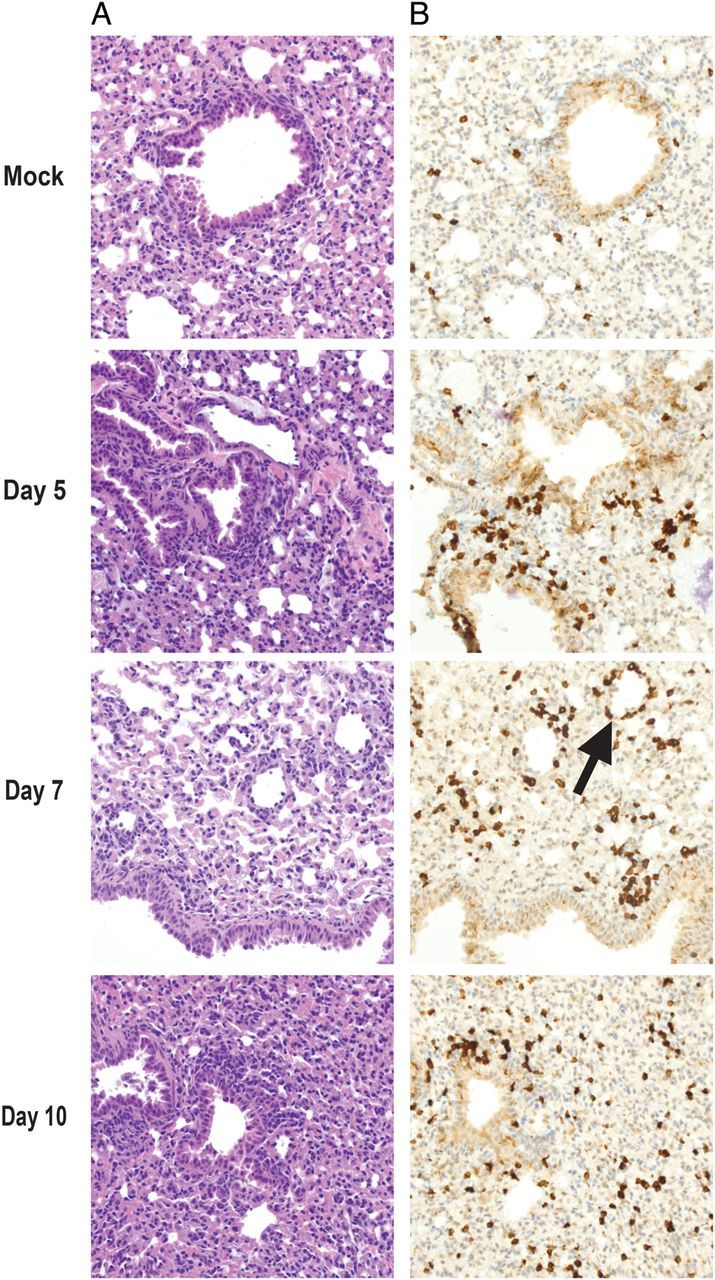
Pulmonary histopathologic findings from DBA/2 mice infected with human metapneumovirus. A, Hematoxylin-eosin–stained specimens depict a mononuclear infiltrate with bronchiolitis and perivascular inflammation. B, CD3+ stained specimens demonstrate peak T-cell infiltrates 7 days after infection (arrow). Images are 40× the original magnification.
54G10 Is Effective Prophylactically Against HMPV
DBA/2 mice were inoculated intraperitoneally with 54G10, PBS, or palivizumab, an RSV F protein–specific isotype control mAb. Twenty-four hours later, mice were infected with HMPV A2 and euthanized on day 5 to measure peak virus titer. The NTs of mice that received prophylaxis with 54G10 had no significant decrease in titer, compared with mice that received PBS or palivizumab (Figure 3A). The lungs of mice that received 54G10 prophylaxis had a significant reduction in titer, compared with control mice. Mice that received 0.2 mg/kg of 54G10 had >2-log10 decrease in virus, and at doses of 0.6 mg/kg 54G10 decreased the lung titer >1000-fold (P < .001; Figure 3B). This dose yielded a mean serum 54G10 concentration (±standard error of the mean) of 12.2 ± 0.1 µg/mL at the time of infection. Sera from treated mice also neutralized HMPV A2 in vitro at a similar IC50 (550 ng/mL) to purified 54G10. In mice infected with HMPV 1 week after receiving prophylaxis, a 54G10 dose of 0.6 mg/kg retained protective efficacy (Supplementary Figure 2A and 2B).
Figure 3.
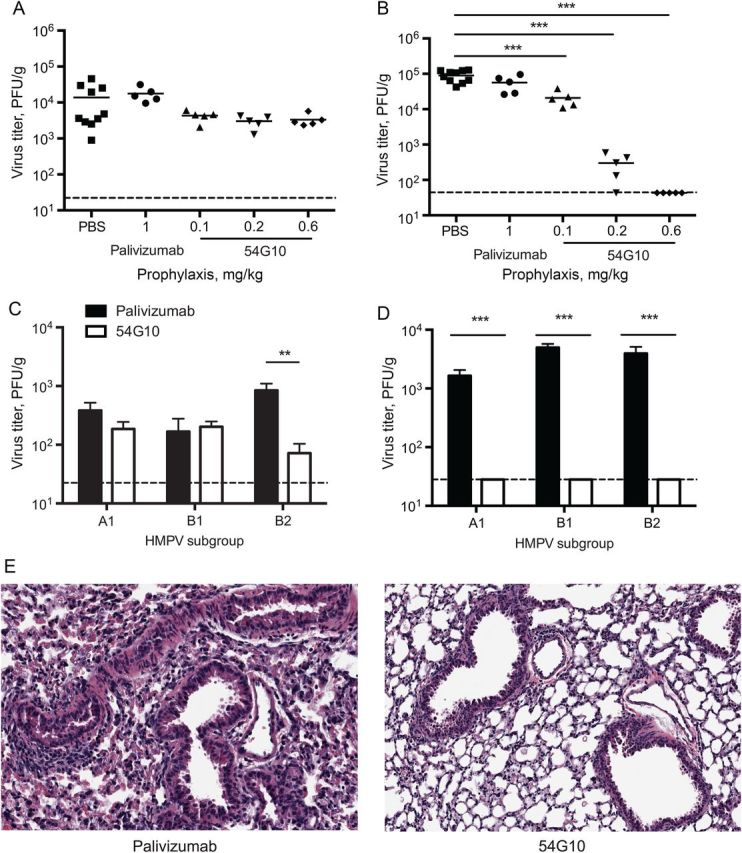
Prophylactic efficacy of 54G10 in mice. A, Prophylactic 54G10 does not decrease virus titer in the nasal turbinates (NTs) of human metapneumovirus (HMPV) A2–infected mice, compared with control. B, Prophylactic 54G10 decreases peak virus titer in the lungs of HMPV A2–infected mice (***P < .001). C, 54G10 prophylaxis decreases virus titer in the NTs of mice infected with HMPV B2 (**P = .01). D, All mice that received 54G10 prophylaxis had a decrease in lung titer, regardless of the HMPV subgroup (***P < .001). Virus titer was log10 transformed and analyzed by the Student t test, with 3–5 mice per group. E, Pulmonary histopathologic findings by hematoxylin-eosin staining on day 7 of mice that received 54G10 prophylaxis and were infected with HMPV A2 demonstrate preservation of airway spaces. Images are 100× the original magnification. Abbreviations: PBS, phosphate-buffered saline; PFU, plaque-forming unit.
To test the efficacy of 54G10 against the remaining HMPV subgroups, mice were inoculated intraperitoneally with either 0.6 mg/kg of 54G10 or isotype control mAb and infected with HMPV A1, B1, or B2 1 day later. Mice that received 54G10 prophylaxis had a 1-log10 decrease in B2 virus titer in the NTs, compared with control mice (P = .01). Mice infected with A1 and B1 did not have a significant change in the NT virus titer (Figure 3C). In the lungs, virus was at or below the LOD in mice that received 54G10 prophylaxis, a significant decrease from findings for control mice (P < .001; Figure 3D). 54G10 also diminished lung inflammation. Mice that received 54G10 prophylaxis had decreased airway disease with preservation of the airway spaces on day 7, compared with control mice (Figure 3E). CD3+ staining demonstrated no significant difference in T-cell infiltrates (data not shown).
54G10 Is Effective Therapeutically Against HMPV
Mice were infected with HMPV A2 and treated intranasally with PBS, palivizumab, or 54G10 on day 3 after infection. Animals were euthanized on day 5 to determine the therapeutic efficacy of 54G10 on peak virus titer. Mice treated with a high concentration of 54G10 or palivizumab had a minimal nonspecific decrease in virus titer in NTs (Figure 4A). The lungs of mice that received 0.01 mg/kg of 54G10 had a modest decrease in virus titer, and at doses of ≥0.5 mg/kg, virus was below the LOD (P < .001; Figure 4B). Since mAbs are administered to humans parenterally rather than via the respiratory tract, we determined the therapeutic efficacy of intraperitoneally administered 54G10. When 54G10 was administered intraperitoneally in the therapeutic model, NT virus titer was unchanged (Figure 4C), but lung virus titer was significantly decreased (P < .001; Figure 4D).
Figure 4.

Therapeutic efficacy of 54G10 in mice. A, Mice treated with inhaled 54G10 on day 3 after infection with human metapneumovirus (HMPV) A2 did not have a significant decrease in virus titer, compared with those that received isotype control antibody. A nonspecific decrease in virus titer was noted after treatment using any monoclonal antibody (*P < .05). Virus titer was log10 transformed and analyzed by analysis of variance with the post-hoc Tukey multiple comparisons test. B, In the lungs, virus titer decreases below the limit of detection in mice treated with 54G10 (***P < .001 for all 54G10 groups, compared with the group that received phosphate-buffered saline [PBS]). Intraperitoneal 54G10 yields a similar reduction in viral titer in the nasal turbinates (C) and lungs (D; *P < .05 and ***P < .001). Virus titer was log10 transformed and analyzed by analysis of variance with the post-hoc Dunnett multiple comparisons test. Abbreviations: NS, not significant; PFU, plaque-forming unit.
MARMs Identify the Epitope of 54G10
MARMs were generated by passage of HMPV A2 in increasing 54G10 concentrations. The MARM was not neutralized by 10 times the IC50 of 54G10 for the parent strain; the IC50 of 54G10 for the MARM was >100 µg/mL (Figure 5A). The F proteins of 4 plaque-purified clones were sequenced and compared to that of the parent virus. All 4 MARMs contained 2 mutations: a valine to glycine substitution at amino acid 397 and a glutamine to histidine substitution at amino acid 434 (Figure 5B). These mutations were not present in the HMPV subgroups used in these experiments.
Figure 5.

Human metapneumovirus (HMPV) 54G10 monoclonal antibody–resistant mutant (MARM). A, The 50% inhibitory concentration of 54G10 for the HMPV MARM is >100 µg/mL. Bars represent the standard error of the mean from 2 independent experiments performed in triplicate. B, The escape mutant sequence has a valine to glycine mutation at amino acid 397 and a glutamine to histidine mutation at amino acid 434. Mutations are not present in other HMPV sequences.
We generated F proteins containing V397G, Q434H, and V397G/Q434H mutations and assessed the binding affinity of 54G10 to these mutated proteins (Supplementary Table 1). The Koff of 54G10 to B2FΔTM-V397G was decreased, compared with wild type, yielding a 1-log10 decrease in the KD (4.1 × 10−9 M). B2FΔTM-V397G/Q434H had similar characteristics to B2FΔTM-V397G, while B2FΔTM-Q434H had rates that closely approximated that of the wild-type protein. These data suggest that the primary binding epitope of 54G10 is near amino acid 397.
54G10 Exhibits Neutralizing Activity Against RSV
The putative 54G10 epitope on HMPV F is similar to antigenic site IV on RSV F protein [22], suggesting that 54G10 may also neutralize RSV. IFA revealed that 54G10 bound to RSV A2 (Figure 6A). DS7, a human mAb against HMPV F, did not bind to RSV A2 (data not shown). We performed in vitro neutralization assays to determine the IC50 of 54G10 for RSV A2 and compared the efficacy with palivizumab, a potent RSV mAb, and DS7 (Figure 6B). 54G10 neutralized RSV A2, with an IC50 of 14.2 µg/mL. The IC50 of palivizumab was 2.4 µg/mL, and DS7 did not neutralize RSV A2.
Figure 6.

54G10 neutralization of respiratory syncytial virus (RSV). A, 54G10 binds to RSV, as revealed by immunofluorescent assay. B, 54G10 neutralizes RSV in vitro. Brackets represent the standard error of the mean. Palivizumab neutralizes RSV, but DS7 does not. C, Prophylactic 54G10 decreases virus titer in the lungs, compared with control (***P < .001). D, Therapeutic 54G10 decreases lung virus titer, compared with control (***P < .001). Virus titer was log10 transformed and analyzed by analysis of variance with the post-hoc Dunnett multiple comparisons test. Abbreviations: mAb, monoclonal antibody; PFU, plaque-forming unit.
To assess the in vivo efficacy of 54G10 against RSV, BALB/c mice received intraperitoneal prophylaxis with PBS, 5 mg/kg DS7, or 1 mg/kg 54G10. Mice were infected with RSV A2 24 hours later and were euthanized on day 5, when the virus titer in BALB/c mice peaked [25]. Virus titer in the NTs was unchanged (data not shown). Lung virus titer was significantly decreased in mice that received 54G10 prophylaxis, compared with control mice (P < .001; Figure 6C). 54G10 also demonstrated therapeutic efficacy against RSV. When administered intraperitoneally 2 days after RSV infection, 54G10 demonstrated therapeutic efficacy in the lungs, reducing the RSV titer on day 5 >100-fold (Figure 6D).
We tested whether 54G10's efficacy could be due to ex vivo neutralization during tissue processing by combining equal amounts of lung homogenate from 54G10-treated and mock-treated mice in both HMPV and RSV models. Plaque assay demonstrated a dilutional effect without evidence of further neutralization.
DISCUSSION
We describe a highly efficacious HMPV mAb, with activity against RSV, in a new mouse model. HMPV A subgroups are used in established murine models [10, 21, 23, 27], while the B subgroups replicate poorly in both murine and larger animal models [28, 29]. Thus, the capacity to test prophylactic or therapeutic interventions against B subgroup viruses has been lacking, even though B subgroup human infections are common [6]. A limitation of our model is that mice did not demonstrate overt signs of illness, but this is common in other HMPV rodent models [23, 30]. Pulmonary histopathologic findings approximate those in animal models [10, 31] and humans [32]. The DBA/2 mouse is a cost-effective, permissive small animal model for all HMPV subgroups and has significant utility in vaccine and therapeutic studies.
54G10 exhibited potent neutralization, with an IC50 in the nanograms/milliliter range against all 4 HMPV subgroups. The IC50 is lower than that of previously reported human mAbs against HMPV F protein and comparable to that of murine mAbs [19, 20]. Although the F protein is highly conserved, mAbs targeting HMPV F protein do not always neutralize all subgroups [19, 20]. Given the variability in subgroup circulation, an effective mAb must have broad neutralizing properties [6, 33, 34]. 54G10's efficacy is likely related to its high affinity to HMPV F protein.
Prophylactic 54G10 was highly effective at decreasing viral replication at doses of 0.6 mg/kg, a dose lower than other prophylactic models [21, 35, 36]. Although 54G10 prophylaxis led to preservation of the alveolar spaces, CD3+ cells were observed, indicating that a T-cell response and, likely, development of memory CD8+ cells occurred. Previous studies have demonstrated that T-cell immunity develops in the presence of neutralizing antibodies [37, 38]. Therefore, passive immunization would not be expected to prevent the development of immune memory, which is also important for protection.
The utility of mAbs for the treatment of infectious diseases has been less successful than prophylaxis. Palivizumab was ineffective at treating RSV in a mouse model, although motavizumab, an RSV F protein mAb, reduced virus titer at high doses [36]. Similarly, high doses of HMPV mAb have been used in mice, with limited success [21]. Smaller doses were effective when administered intranasally; however, intranasal administration is not practical in humans [20]. Our data demonstrates that 54G10 has therapeutic efficacy both intranasally and systemically.
Development of MARMs identified 2 mutations. Although mutation Q434H did not significantly change 54G10 affinity, mutation V397G resulted in a 1-log10 decrease in the KD, suggesting that this is the primary epitope. This mutation maps to HMPV antigenic site 5-6 [39]. Previous mAbs binding in this region had limited efficacy in vitro, including the inability to neutralize all 4 subgroups [19]. Therefore, 54G10 binds to HMPV F protein at a unique epitope. Neither mutation was present in 81 other HMPV isolates worldwide, suggesting that the epitope is conserved among HMPV strains [40]. 54G10 did not inhibit HMPV binding or hemi-fusion (data not shown).
The F proteins of HMPV and RSV are immunogenic [12, 41], and broadly neutralizing antibodies specific for each virus have been identified [19, 42]. Despite 33% amino acid similarity between proteins [43], cross-neutralization of both viruses was thought to not occur [44]. Recently, human mAb MPE8, with in vitro efficacy against all subgroups of RSV and HMPV, was identified from screening 114 000 B cells [45]. The putative epitope of MPE8 is accessible only on the prefusion RSV F protein. This region is not near the mutations in 54G10 MARMs, suggesting multiple conserved epitopes among pneumoviruses. We postulate that the 54G10 epitope is present in both the pre- and postfusion states, based on the analogous RSV epitope [46]. Analysis of 63 RSV strains demonstrate that the epitope is a conserved region.
Monoclonal antibodies have great potential for clinical use in infectious diseases. Palivizumab decreases hospitalization rates from RSV in high-risk infants [15]; however, few data support its therapeutic use [35, 47]. Our data suggest that 54G10 may have therapeutic utility. One concern in using chimeric and humanized mAbs is immunogenicity and development of side effects [16, 48]. 54G10 is a fully human mAb, which likely would induce less host immunogenicity than mAbs with murine regions.
In addition to passive immunity and therapy, mAbs have other medical implications. Structural vaccine design based on a potent epitope has been used to elicit broadly neutralizing antibodies for other viruses [49, 50]. Structural characterization of 54G10's epitope could guide HMPV vaccine development. Moreover, a structurally conserved epitope between HMPV and RSV would be a highly effective vaccine target against leading causes of pediatric and adult respiratory disease.
In summary, 54G10 is a human mAb with subnanomolar affinity for the HMPV F protein, broad neutralizing capacity, and in vivo prophylactic and therapeutic efficacy against HMPV and RSV. Further studies are needed in other animal models and to define this conserved epitope among pneumoviruses, which should be considered as a target for both passive immunity, as well as vaccine design.
Supplementary Data
Supplementary materials are available at The Journal of Infectious Diseases online (http://jid.oxfordjournals.org). Supplementary materials consist of data provided by the author that are published to benefit the reader. The posted materials are not copyedited. The contents of all supplementary data are the sole responsibility of the authors. Questions or messages regarding errors should be addressed to the author.
Notes
Acknowledgments. We thank Jessica Gillon, for assistance in obtaining palivizumab; and the Vanderbilt Antibody and Protein Resource (which is supported by the Vanderbilt Institute of Chemical Biology and the Vanderbilt Ingram Cancer Center [P30 CA68485]), for performing interaction assays by means of the ForteBio Octet Red96 system.
Disclaimer. The contents of this article are solely the responsibility of the authors and do not necessarily represent official views of the National Center for Advancing Translational Sciences or the National Institutes of Health.
Financial support. This work was supported by the National Institutes of Health (grants R01 AI085062 to J. V. W., R01AI072414 to D. R. B., and 5T32HD060554 to J. E. S.) and the National Center for Advancing Translational Sciences (CTSA award UL1TR000445).
Potential conflicts of interest. J. V. W. serves on the scientific advisory board of Quidel. All other authors report no potential conflicts.
All authors have submitted the ICMJE Form for Disclosure of Potential Conflicts of Interest. Conflicts that the editors consider relevant to the content of the manuscript have been disclosed.
References
- 1.Williams JV, Harris PA, Tollefson SJ, et al. Human metapneumovirus and lower respiratory tract disease in otherwise healthy infants and children. N Engl J Med. 2004;350:443–50. doi: 10.1056/NEJMoa025472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.van den Hoogen BG, de Jong JC, Groen J, et al. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat Med. 2001;7:719–24. doi: 10.1038/89098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Edwards KM, Zhu Y, Griffin MR, et al. Burden of human metapneumovirus infection in young children. N Engl J Med. 2013;368:633–43. doi: 10.1056/NEJMoa1204630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Englund JA, Boeckh M, Kuypers J, et al. Brief communication: fatal human metapneumovirus infection in stem-cell transplant recipients. Ann Intern Med. 2006;144:344–9. doi: 10.7326/0003-4819-144-5-200603070-00010. [DOI] [PubMed] [Google Scholar]
- 5.Shahda S, Carlos WG, Kiel PJ, Khan BA, Hage CA. The human metapneumovirus: a case series and review of the literature. Transpl Infect Dis. 2011;13:324–8. doi: 10.1111/j.1399-3062.2010.00575.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Williams JV, Wang CK, Yang CF, et al. The role of human metapneumovirus in upper respiratory tract infections in children: a 20-year experience. J Infect Dis. 2006;193:387–95. doi: 10.1086/499274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Agapov E, Sumino KC, Gaudreault-Keener M, Storch GA, Holtzman MJ. Genetic variability of human metapneumovirus infection: evidence of a shift in viral genotype without a change in illness. J Infect Dis. 2006;193:396–403. doi: 10.1086/499310. [DOI] [PubMed] [Google Scholar]
- 8.Falsey AR, Erdman D, Anderson LJ, Walsh EE. Human metapneumovirus infections in young and elderly adults. J Infect Dis. 2003;187:785–90. doi: 10.1086/367901. [DOI] [PubMed] [Google Scholar]
- 9.van den Hoogen BG, Herfst S, de Graaf M, et al. Experimental infection of macaques with human metapneumovirus induces transient protective immunity. J Gen Virol. 2007;88:1251–9. doi: 10.1099/vir.0.82663-0. [DOI] [PubMed] [Google Scholar]
- 10.Erickson JJ, Gilchuk P, Hastings AK, et al. Viral acute lower respiratory infections impair CD8+ T cells through PD-1. J Clin Invest. 2012;122:2967–82. doi: 10.1172/JCI62860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lamb RA, Jardetzky TS. Structural basis of viral invasion: lessons from paramyxovirus F. Curr Opin Struct Biol. 2007;17:427–36. doi: 10.1016/j.sbi.2007.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cseke G, Wright DW, Tollefson SJ, Johnson JE, Crowe JE, Jr., Williams JV. Human metapneumovirus fusion protein vaccines that are immunogenic and protective in cotton rats. J Virol. 2007;81:698–707. doi: 10.1128/JVI.00844-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Herfst S, de Graaf M, Schrauwen EJ, et al. Immunization of Syrian golden hamsters with F subunit vaccine of human metapneumovirus induces protection against challenge with homologous or heterologous strains. J Gen Virol. 2007;88:2702–9. doi: 10.1099/vir.0.83084-0. [DOI] [PubMed] [Google Scholar]
- 14.Peret TC, Boivin G, Li Y, et al. Characterization of human metapneumoviruses isolated from patients in North America. J Infect Dis. 2002;185:1660–3. doi: 10.1086/340518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Palivizumab, a humanized respiratory syncytial virus monoclonal antibody, reduces hospitalization from respiratory syncytial virus infection in high-risk infants. Pediatrics. 1998;102:531–7. [PubMed] [Google Scholar]
- 16.Carbonell-Estrany X, Simoes EA, Dagan R, et al. Motavizumab for prophylaxis of respiratory syncytial virus in high-risk children: a noninferiority trial. Pediatrics. 2010;125:e35–51. doi: 10.1542/peds.2008-1036. [DOI] [PubMed] [Google Scholar]
- 17.Maini R, St Clair EW, Breedveld F, et al. Infliximab (chimeric anti-tumour necrosis factor alpha monoclonal antibody) versus placebo in rheumatoid arthritis patients receiving concomitant methotrexate: a randomised phase III trial. ATTRACT Study Group. Lancet. 1999;354:1932–9. doi: 10.1016/s0140-6736(99)05246-0. [DOI] [PubMed] [Google Scholar]
- 18.den Broeder A, van de Putte L, Rau R, et al. A single dose, placebo controlled study of the fully human anti-tumor necrosis factor-alpha antibody adalimumab (D2E7) in patients with rheumatoid arthritis. J Rheumatol. 2002;29:2288–98. [PubMed] [Google Scholar]
- 19.Ulbrandt ND, Ji H, Patel NK, et al. Isolation and characterization of monoclonal antibodies which neutralize human metapneumovirus in vitro and in vivo. J Virol. 2006;80:7799–806. doi: 10.1128/JVI.00318-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Williams JV, Chen Z, Cseke G, et al. A recombinant human monoclonal antibody to human metapneumovirus fusion protein that neutralizes virus in vitro and is effective therapeutically in vivo. J Virol. 2007;81:8315–24. doi: 10.1128/JVI.00106-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hamelin ME, Gagnon C, Prince GA, et al. Prophylactic and therapeutic benefits of a monoclonal antibody against the fusion protein of human metapneumovirus in a mouse model. Antiviral Res. 2010;88:31–7. doi: 10.1016/j.antiviral.2010.07.001. [DOI] [PubMed] [Google Scholar]
- 22.McLellan JS, Chen M, Chang JS, et al. Structure of a major antigenic site on the respiratory syncytial virus fusion glycoprotein in complex with neutralizing antibody 101F. J Virol. 2010;84:12236–44. doi: 10.1128/JVI.01579-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Williams JV, Tollefson SJ, Johnson JE, Crowe JE., Jr The cotton rat (Sigmodon hispidus) is a permissive small animal model of human metapneumovirus infection, pathogenesis, and protective immunity. J Virol. 2005;79:10944–51. doi: 10.1128/JVI.79.17.10944-10951.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cox RG, Livesay SB, Johnson M, Ohi MD, Williams JV. The human metapneumovirus fusion protein mediates entry via an interaction with RGD-binding integrins. J Virol. 2012;86:12148–60. doi: 10.1128/JVI.01133-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Graham BS, Perkins MD, Wright PF, Karzon DT. Primary respiratory syncytial virus infection in mice. J Med Virol. 1988;26:153–62. doi: 10.1002/jmv.1890260207. [DOI] [PubMed] [Google Scholar]
- 26.Wen X, Krause JC, Leser GP, et al. Structure of the human metapneumovirus fusion protein with neutralizing antibody identifies a pneumovirus antigenic site. Nat Struct Mol Biol. 2012;19:461–3. doi: 10.1038/nsmb.2250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hamelin ME, Yim K, Kuhn KH, et al. Pathogenesis of human metapneumovirus lung infection in BALB/c mice and cotton rats. J Virol. 2005;79:8894–903. doi: 10.1128/JVI.79.14.8894-8903.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Wyde PR, Chetty SN, Jewell AM, Schoonover SL, Piedra PA. Development of a cotton rat-human metapneumovirus (hMPV) model for identifying and evaluating potential hMPV antivirals and vaccines. Antiviral Res. 2005;66:57–66. doi: 10.1016/j.antiviral.2004.12.009. [DOI] [PubMed] [Google Scholar]
- 29.Skiadopoulos MH, Biacchesi S, Buchholz UJ, et al. The two major human metapneumovirus genetic lineages are highly related antigenically, and the fusion (F) protein is a major contributor to this antigenic relatedness. J Virol. 2004;78:6927–37. doi: 10.1128/JVI.78.13.6927-6937.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.MacPhail M, Schickli JH, Tang RS, et al. Identification of small-animal and primate models for evaluation of vaccine candidates for human metapneumovirus (hMPV) and implications for hMPV vaccine design. J Gen Virol. 2004;85:1655–63. doi: 10.1099/vir.0.79805-0. [DOI] [PubMed] [Google Scholar]
- 31.Kuiken T, van den Hoogen BG, van Riel DA, et al. Experimental human metapneumovirus infection of cynomolgus macaques (Macaca fascicularis) results in virus replication in ciliated epithelial cells and pneumocytes with associated lesions throughout the respiratory tract. Am J Pathol. 2004;164:1893–900. doi: 10.1016/S0002-9440(10)63750-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Vargas SO, Kozakewich HP, Perez-Atayde AR, McAdam AJ. Pathology of human metapneumovirus infection: insights into the pathogenesis of a newly identified respiratory virus. Pediatr Dev Pathol. 2004;7:478–86. doi: 10.1007/s10024-004-1011-2. discussion 21. [DOI] [PubMed] [Google Scholar]
- 33.Papenburg J, Hamelin ME, Ouhoummane N, et al. Comparison of risk factors for human metapneumovirus and respiratory syncytial virus disease severity in young children. J Infect Dis. 2012;206:178–89. doi: 10.1093/infdis/jis333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Vicente D, Montes M, Cilla G, Perez-Yarza EG, Perez-Trallero E. Differences in clinical severity between genotype A and genotype B human metapneumovirus infection in children. Clin Infect Dis. 2006;42:e111–3. doi: 10.1086/504378. [DOI] [PubMed] [Google Scholar]
- 35.Mejias A, Chavez-Bueno S, Rios AM, et al. Anti-respiratory syncytial virus (RSV) neutralizing antibody decreases lung inflammation, airway obstruction, and airway hyperresponsiveness in a murine RSV model. Antimicrob Agents Chemother. 2004;48:1811–22. doi: 10.1128/AAC.48.5.1811-1822.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mejias A, Chavez-Bueno S, Rios AM, et al. Comparative effects of two neutralizing anti-respiratory syncytial virus (RSV) monoclonal antibodies in the RSV murine model: time versus potency. Antimicrob Agents Chemother. 2005;49:4700–7. doi: 10.1128/AAC.49.11.4700-4707.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Crowe JE, Jr., Bui PT, Siber GR, Elkins WR, Chanock RM, Murphy BR. Cold-passaged, temperature-sensitive mutants of human respiratory syncytial virus (RSV) are highly attenuated, immunogenic, and protective in seronegative chimpanzees, even when RSV antibodies are infused shortly before immunization. Vaccine. 1995;13:847–55. doi: 10.1016/0264-410x(94)00074-w. [DOI] [PubMed] [Google Scholar]
- 38.Crowe JE, Jr., Firestone CY, Murphy BR. Passively acquired antibodies suppress humoral but not cell-mediated immunity in mice immunized with live attenuated respiratory syncytial virus vaccines. J Immunol. 2001;167:3910–8. doi: 10.4049/jimmunol.167.7.3910. [DOI] [PubMed] [Google Scholar]
- 39.Ulbrandt ND, Ji H, Patel NK, et al. Identification of antibody neutralization epitopes on the fusion protein of human metapneumovirus. J Gen Virol. 2008;89:3113–8. doi: 10.1099/vir.0.2008/005199-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yang CF, Wang CK, Tollefson SJ, et al. Genetic diversity and evolution of human metapneumovirus fusion protein over twenty years. Virol J. 2009;6:138. doi: 10.1186/1743-422X-6-138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Groothuis JR, King SJ, Hogerman DA, Paradiso PR, Simoes EA. Safety and immunogenicity of a purified F protein respiratory syncytial virus (PFP-2) vaccine in seropositive children with bronchopulmonary dysplasia. J Infect Dis. 1998;177:467–9. doi: 10.1086/517377. [DOI] [PubMed] [Google Scholar]
- 42.Kwakkenbos MJ, Diehl SA, Yasuda E, et al. Generation of stable monoclonal antibody-producing B cell receptor-positive human memory B cells by genetic programming. Nat Med. 2010;16:123–8. doi: 10.1038/nm.2071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.van den Hoogen BG, Bestebroer TM, Osterhaus AD, Fouchier RA. Analysis of the genomic sequence of a human metapneumovirus. Virology. 2002;295:119–32. doi: 10.1006/viro.2001.1355. [DOI] [PubMed] [Google Scholar]
- 44.Wyde PR, Chetty SN, Jewell AM, Boivin G, Piedra PA. Comparison of the inhibition of human metapneumovirus and respiratory syncytial virus by ribavirin and immune serum globulin in vitro. Antiviral Res. 2003;60:51–9. doi: 10.1016/s0166-3542(03)00153-0. [DOI] [PubMed] [Google Scholar]
- 45.Corti D, Bianchi S, Vanzetta F, et al. Cross-neutralization of four paramyxoviruses by a human monoclonal antibody. Nature. 2013;501:439–43. doi: 10.1038/nature12442. [DOI] [PubMed] [Google Scholar]
- 46.McLellan JS, Yang Y, Graham BS, Kwong PD. Structure of respiratory syncytial virus fusion glycoprotein in the postfusion conformation reveals preservation of neutralizing epitopes. J Virol. 2011;85:7788–96. doi: 10.1128/JVI.00555-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.de Fontbrune FS, Robin M, Porcher R, et al. Palivizumab treatment of respiratory syncytial virus infection after allogeneic hematopoietic stem cell transplantation. Clin Infect Dis. 2007;45:1019–24. doi: 10.1086/521912. [DOI] [PubMed] [Google Scholar]
- 48.Feltes TF, Sondheimer HM, Tulloh RM, et al. A randomized controlled trial of motavizumab versus palivizumab for the prophylaxis of serious respiratory syncytial virus disease in children with hemodynamically significant congenital heart disease. Pediatr Res. 2011;70:186–91. doi: 10.1203/PDR.0b013e318220a553. [DOI] [PubMed] [Google Scholar]
- 49.Kanekiyo M, Wei CJ, Yassine HM, et al. Self-assembling influenza nanoparticle vaccines elicit broadly neutralizing H1N1 antibodies. Nature. 2013;499:102–6. doi: 10.1038/nature12202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.McLellan JS, Chen M, Joyce MG, et al. Structure-based design of a fusion glycoprotein vaccine for respiratory syncytial virus. Science. 2013;342:592–8. doi: 10.1126/science.1243283. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.