

Abstract
A previous study showed that ammonia oxidation by the Thaumarchaeota Nitrosopumilus maritimus (group 1.1a) was resistant to concentrations of the C8 1-alkyne, octyne, which completely inhibits activity by ammonia-oxidizing bacteria. In this study, the inhibitory effects of octyne and other C2 to C10 1-alkynes were evaluated on the nitrite production activity of two pure culture isolates from Thaumarchaeota group 1.1b, Nitrososphaera viennensis strain EN76 and Nitrososphaera gargensis. Both N. viennensis and N. gargensis were insensitive to concentrations of octyne that cause complete and irreversible inactivation of nitrite production by ammonia-oxidizing bacteria. However, octyne concentrations (≥20 μM) that did not inhibit N. maritimus partially inhibited nitrite production in N. viennensis and N. gargensis in a manner that did not show the characteristics of irreversible inactivation. In contrast to previous studies with an ammonia-oxidizing bacterium, Nitrosomonas europaea, octyne inhibition of N. viennensis was: (i) fully and immediately reversible, (ii) not competitive with NH4+, and (iii) without effect on the competitive interaction between NH4+ and acetylene. Both N. viennensis and N. gargensis demonstrated the same overall trend in regard to 1-alkyne inhibition as previously observed for N. maritimus, being highly sensitive to ≤C5 alkynes and more resistant to longer-chain length alkynes. Reproducible differences were observed among N. maritimus, N. viennensis, and N. gargensis in regard to the extent of their resistance/sensitivity to C6 and C7 1-alkynes, which may indicate differences in the ammonia monooxygenase binding and catalytic site(s) among the Thaumarchaeota.
INTRODUCTION
With only a few isolates of ammonia-oxidizing archaea (AOA) available in pure/axenic culture, very little is known about the physiological and biochemical differences that might exist across the broad genotypic range of AOA (1–3) or how AOA physiology differs from that of ammonia-oxidizing bacteria (AOB). Although it is generally believed that both AOA and AOB initiate the oxidation of NH3 with ammonia monooxygenase (AMO), there is evidence that suggests some aspects of NH3 oxidation are probably different between them. For example, the NO scavenger 2-phenyl-4,4,5,5-tetramethylimidazoline-1-oxide-3-oxyl (PTIO) inhibits NH3 oxidation by AOA and has little effect on AOB activity (4–6), suggesting that the intermediary products of NH3 oxidation might be fundamentally different in AOA and AOB. Furthermore, whereas NH3-oxidizing activity in AOB is inhibited by low concentrations (0.4 to 10 μM) of the Cu chelator, allylthiourea (ATU) (4, 7–9), the ATU sensitivity of NH3 oxidation by AOA is more variable. For example, NH3-oxidizing activity of Nitrososphaera gargensis is only partially inhibited by 100 μM ATU (10), and 500 μM ATU is required to completely stop NH3-oxidizing activity in Nitrososphaera viennensis strain EN76 (4). In contrast, NH3 oxidation by an obligate acidophilic AOA, Nitrosotalea devanaterra, is completely inhibited by 50 to 100 μM ATU (11).
Recently, Taylor et al. (12) showed that NO2− production by the marine AOA, Nitrosopumilus maritimus, was unaffected by 20 μM aqueous concentration (Caq) of C8 and C9 1-alkynes, whereas NH3-oxidizing activity of AOB is completely and quickly (≤30 min) inactivated by 1 to 5 μM Caq of 1-alkynes with a chain length of C2 to C10 (13). A short-term assay, based upon the difference in sensitivity of AOA and AOB to 1-octyne, was subsequently developed to measure the contributions of AOA and AOB to soil nitrifying activity. With the availability of more AOA isolates in pure culture, we deemed it important to confirm if the octyne resistance properties of N. maritimus were similar among other AOA isolates, and particularly among the group 1.1b lineage of Thaumarchaeota, whose members are widely distributed in terrestrial environments (2, 14). We report here the results of studies carried out to assess the effects of 1-alkynes of various chain lengths on the NH3-oxidizing activity of two axenic culture isolates of AOA from group 1.1b, namely, Nitrososphaera viennensis strain EN76 (15, 16) and Nitrososphaera gargensis (1, 10). Both strains showed similar, albeit not identical 1-alkyne chain length inhibition profiles to N. maritimus. However, both strains were partially inhibited by ≥20 μM octyne, which led to additional studies to compare the inhibitory properties of octyne with those of the more extensively studied alkyne, acetylene.
MATERIALS AND METHODS
Chemicals.
NH4Cl and linear aliphatic 1-alkynes (C3 to C10, ≥97%) were obtained from Sigma-Aldrich (St. Louis, MO), and acetylene (C2H2) was obtained from Airgas (Radnor, PA).
Cultures.
Nitrososphaera viennensis strain EN76, here referred to as N. viennensis, was cultivated at its optimal growth temperature of 42°C in a freshwater medium (FWM) (16, 17), supplemented with 2 mM NH4+ and 1 mM pyruvate. Growth was routinely followed by measuring NO2− production as described previously (12). Nitrososphaera gargensis has now been obtained into pure culture (M. Palatinszky et al., unpublished data) and was cultivated at its optimal growth temperature of 46°C in a carbonate media (18) supplemented with 2 mM NH4+. During routine growth of N. gargensis, further additions of 2 mM NH4+ were made after NH4+ was consumed. Cultures that had consumed a total of 8 to 10 mM NH4+ were used for experimental purposes. The excess insoluble carbonate in the N. gargensis medium provides: (i) buffering against the acidification that occurs with 10 mM NO2− production and (ii) is thought to provide a growth-promoting surface for N. gargensis. The presence of the carbonate provides challenges during experimental manipulations. For example, harvesting and concentration of the culture leads to a dense suspension of carbonate that settles immediately upon onset of static conditions. This phenomenon makes it difficult to dispense uniform aliquots of cells to experimental replicates. Because of the challenges posed by the excess carbonate in the media, only a subset of the experiments were performed with N. gargensis. As expected, the N. gargensis cultures grown with 8 to 10 mM NH4+ yielded higher protein concentrations (132.6 ± 21.0 μg of protein/ml of culture) than the N. viennensis cultures grown with 2 mM NH4+ (9.0 ± 0.3 μg of protein/ml of culture). However, the N. gargensis cultures had much lower rates of NO2− production when expressed on a milligram protein basis. Protein concentrations were determined by using a Pierce BCA protein assay kit (Thermo Scientific, Waltham, MA) as described by the manufacturer.
Octyne sensitivity.
Octyne stocks were prepared as described previously (12). N. viennensis was cultivated at 42°C in 200-ml volumes of FWM. After accumulation of 1.8 to 2 mM NO2−, the cultures were harvested by centrifugation (10,000 × g, 15 min) and concentrated 100-fold by resuspension in 2 ml of fresh FWM supplemented with 1 mM pyruvate but minus NH4+. N. gargensis was cultivated at 46°C in 200 ml of carbonate medium supplemented with a total addition of 8 to 10 mM NH4+, harvested by centrifugation (10,000 × g, 15 min), and concentrated 100-fold by resuspension in 2 ml of fresh carbonate medium minus NH4+. Aliquots (100 μl) of N. viennensis or N. gargensis cell suspensions were added to 10-ml portions of their respective growth media supplemented with 1 mM NH4+ that had been pre-equilibrated with octyne (0, 10, 20, or 40 μM) for 30 min at either 42 or 46°C for N. viennensis and N. gargensis, respectively. Bottles were incubated as described above and sampled periodically over 6 h to quantify the rate of accumulation of NO2−. Additional incubations were carried out with N. viennensis at two different temperatures (35 and 46°C) to evaluate the sensitivity of NO2− production to 10 μM octyne. An experiment was performed at 42°C in the presence or absence of 1 mM pyruvate to determine whether the latter influenced the rate and sensitivity of NO2− production to 10 μM octyne.
Sensitivity of AOA isolates to C2 to C10 alkynes.
A preliminary experiment was conducted in bottles containing alkynes pre-equilibrated at 30, 42, and 46°C. Headspace alkyne concentrations were monitored by using gas chromatography. There was no significant difference in headspace alkyne concentrations between bottles incubated across this temperature range, indicating that temperature shifts over this magnitude did not change the gas/aqueous phase-partitioning in the assay bottles sufficient to modify the aqueous concentration (Caq) of alkynes. N. viennensis and N. gargensis were grown and harvested as described above, and 100-fold-concentrated cell aliquots (100 μl) were added to 10-ml portions of their respective growth media supplemented with 1 mM NH4+ that had been pre-equilibrated for 30 min with 10 μM Caq of each alkyne (C2 to C10) in 125-ml bottles sealed with gray butyl stoppers (12). This specific alkyne concentration was chosen after it had been determined that 10 μM octyne had no effect on NH3 oxidation by either N. viennensis or N. gargensis but was previously shown to be more than sufficient to inactivate AOB (12). Bottles were incubated at 42 or 46°C for N. viennensis and N. gargensis, respectively, and NO2− accumulation monitored over 8 h.
Characteristics of acetylene and octyne inhibition of NO2− production by N. viennensis. (i) Kinetic relationships between NH4+ and alkyne inhibition of N. viennensis.
Aliquots of a N. viennensis cell suspension were exposed to different concentrations of either octyne (30 or 40 μM) or acetylene (1, 3, or 6 μM) in the presence of various concentrations of NH4+ (0.01 to 2 mM) at 42°C. Initial rates of NO2− production were determined by sampling every 15 min for a total of 90 to 120 min, and the data were plotted in double-reciprocal Lineweaver-Burk plots.
(ii) Examining the influence of octyne on the relationship between NH4+ concentration and acetylene on the inhibition of NO2− production by N. viennensis.
Aliquots of N. viennensis were exposed to 30 μM octyne with increasing concentrations of NH4+ (0.25 to 2 mM) for 15 min, followed by the addition of 1 μM acetylene. NO2− accumulation was evaluated at 30-min intervals for 120 min at 42°C, and analysis of variance (ANOVA) of rates determined differences in response to acetylene, octyne, or the combination of acetylene and octyne. Controls included treatments without alkyne and a treatment containing 1 μM acetylene minus octyne.
(iii) Time course of recovery of NO2− production in N. viennensis following inhibition by acetylene or octyne.
Aliquots of a N. viennensis cell suspension were exposed to either 40 μM octyne or 6 μM acetylene with an initial NH4+ concentration of 0.5 mM for 2 h, along with a minus-alkyne control also incubated for 2 h. This concentration of NH4+ was chosen to minimize the potential for competition with acetylene. After the alkyne exposure period, bottles were degassed to remove the alkynes (12). Additional NH4+ was added to increase the concentration to 2 mM for optimal growth conditions, and the recovery of NO2− production was monitored by sampling at 60-min intervals over 4 h.
(iv) Hydroxylamine-dependent NO2− production by acetylene- and octyne-treated N. viennensis.
It is well established that hydroxylamine-dependent NO2− production is unaffected by alkyne inactivation of AMO in AOB and has been used on several occasions to infer that alkynes specifically target AMO and have no effect on downstream enzymes in the NO2− production pathway (19, 20). Aliquots of N. viennensis cell suspensions were exposed to 18 μM acetylene or 40 μM octyne, with 2 mM NH4+, and a range of hydroxylamine concentrations (20 to 400 μM), and evidence for hydroxylamine-dependent NO2− accumulation was sought by incubation at 42°C and sampling at 15-min intervals for 2 h. Abiotic controls containing only growth medium plus hydroxylamine were included. We were unable to measure significant hydroxylamine-dependent NO2− production in octyne-inhibited or acetylene-inactivated cultures (P > 0.3). Furthermore, hydroxylamine-dependent NO2− production could not be measured in N. viennensis cultures immediately after total depletion of NH4+ and untreated with either acetylene or octyne.
Statistics.
SigmaPlot 12.5 (Systat Software, Inc., San Jose, CA) was used to perform statistical analyses. ANOVA was used to determine whether the differences in the mean values among the treatment groups were greater than would be expected by chance. If there was a statistically significant difference the Holm-Sidak method of all pairwise multiple-comparison procedure was used to isolate the group or groups that differed from the others. A nonlinear regression procedure was used to estimate N. viennensis Km for NH4+ in the presence of 1 μM acetylene. The best-fit slopes of the relationship between NO2− production and NH4+ concentration in response to the presence of acetylene or octyne were estimated by linear regression of double-reciprocal plots.
RESULTS
Sensitivity of N. viennensis and N. gargensis to octyne.
Initial work focused on the response of NO2− production by N. viennensis and N. gargensis to different concentrations of octyne (C8). The rate of NO2− production by N. viennensis in the presence of 10 μM octyne was not significantly different (P > 0.05) than the rate in the no-alkyne control (175 ± 20 nmol/min/mg of protein, Fig. 1A) at the optimal growth temperature of 42°C. However, 20 and 40 μM octyne partially inhibited NO2− accumulation to 58 and 33%, respectively, of the activity of the no-alkyne control. Initial rates of NO2− production (0 to 6 h) by N. viennensis were also insensitive to 10 μM octyne at 35 or 46°C (P ≥ 0.1), and the presence or absence of 1 mM pyruvate had no effect on the inhibitory effects of octyne (P ≥ 0.1). The rate of NO2− production by N. gargensis was not affected by the presence of 10 μM octyne (P > 0.1) compared to the rate in the no-alkyne control (3.3 ± 1.1 nmol/min/mg of protein, Fig. 1B). As with N. viennensis, NO2− accumulation was partially inhibited by 20 and 40 μM octyne to 56 and 29%, respectively, of the activity of the no-alkyne control. In the case of both AOA isolates, NO2− production in the presence of 20 and 40 μM octyne continued at a constant rate for at least 24 h (data not shown), suggesting that octyne was not behaving as a turnover-dependent irreversible inactivator of AMO. Octyne concentrations in the headspace of the biological treatments at 24 h were no different than in abiotic controls (P > 0.17), providing no evidence of substantial consumption by the AOA. In addition, gas chromatographic analysis provided no evidence of trace contamination of octyne by other volatile compounds that might explain partial inhibition by ≥20 μM octyne.
FIG 1.
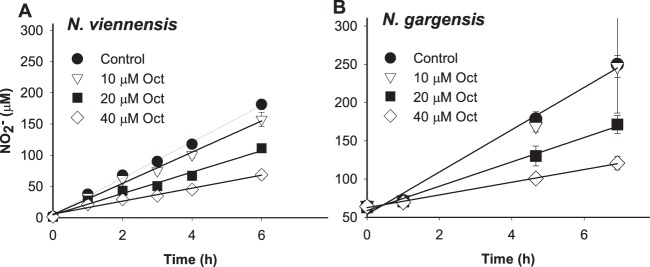
Response of NO2− production by N. viennensis (A) and N. gargensis (B) to different aqueous concentrations of octyne (Oct). Cell suspensions were added to their respective media pre-equilibrated with Oct in the presence of 1 mM NH4+. Bottles were incubated at 42 or 46°C for N. viennensis and N. gargensis, respectively, and the accumulation of NO2− was monitored. Experiments were repeated three times (in triplicate) with similar results. Error bars represent the standard deviations of three replicate bottles of each treatment.
Sensitivity of AOA isolates to C2 to C10 alkynes.
To determine whether there was differential sensitivity to alkynes of various chain lengths, we compared the NO2− production response of N. viennensis and N. gargensis to C2 to C10 1-alkynes in the presence of 1 mM NH4+ (Fig. 2). There was a significant decrease in NO2− production compared to the no-alkyne control when N. viennensis was exposed to 10 μM C2 to C5 and C7 1-alkynes (P < 0.05), but there was no significant difference (P > 0.05) in NO2− accumulation when exposed to C6, and C8 to C10 alkynes. The reduced rate of NO2− production by N. viennensis in the presence of 10 μM C7 (69% ± 1% of control) continued at the same rate for at least 24 h, showing no sign of a time-dependent reduction of the rate as would be anticipated if C7 was behaving as an inactivator of NO2− production (data not shown). Although 10 μM C7 was inhibitory, 2.5 and 5.0 μM C7 did not have a significant effect on the rates of NO2− accumulation (P > 0.07). Furthermore, as described above for octyne, there was no evidence for substantial consumption of C7 in the N. viennensis treatments after 24 h inhibition (P > 0.15), and no evidence of contamination of C7 by other alkynes that might explain partial inhibition. In the case of N. gargensis, there was a significant inhibitory effect of 10 μM C2 to C5 but no significant effect of C6 to C10 alkynes on NO2− accumulation (P > 0.05). In comparison, our previous work had also shown a significant inhibitory effect of C2 to C5 1-alkynes on NO2− production by the marine AOA N. maritimus whereas 20 μM C6 and C7 1-alkynes were partially inhibitory (56% ± 2% and 58% ± 4%, respectively), and 20 μM C8 and C9 1-alkynes had no effect (12).
FIG 2.
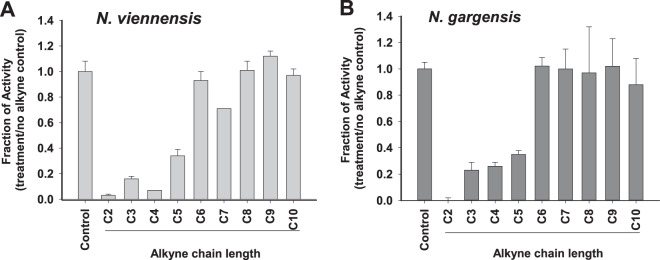
Response of NO2− production by N. viennensis and N. gargensis when exposed to C2 to C10 1-alkynes. Cultures were harvested and aliquots of cell suspensions were added to treatments containing their respective media with 10 μM Caq of C2 to C10 alkyne and 1 mM NH4+. N. viennensis and N. gargensis were incubated at 42 and 46°C, respectively, and NO2− accumulation was measured. Experiments were repeated at least three times with similar results. Bars represent the average activity in three replicate treatments compared to the no-alkyne control. Error bars represent the standard deviations of the average.
Characteristics of acetylene and octyne inhibition of NO2− production by N. viennensis.
Because NO2− production rates in both strains were partially inhibited by ≥20 μM octyne but showed no sign of time-dependent rate reduction, several experiments were conducted to compare in more detail the characteristics of acetylene and octyne inhibition in N. viennensis.
(i) Kinetic relationships between NH4+ and alkyne inhibition of N. viennensis.
The first experiment compared the concentration-dependent inhibition of NO2− production by acetylene and octyne. In the absence of acetylene or octyne the rate of NO2− production by N. viennensis was saturated with NH4+ concentrations from 0.01 to 2.0 mM (Fig. 3A). In the presence of 1 μM acetylene and 0.25 mM NH4+, the initial rate of NO2− production (Vi) was inhibited to 47% of the control, but the inhibition could be completely overcome by increasing the NH4+ concentration to 1 mM (Fig. 3A). When acetylene concentrations were increased to 3 and 6 μM, inhibition could not be completely overcome by increasing the NH4+ concentration to 2 mM. These results suggest that acetylene and NH4+ are competitive substrates for binding at the active site. Although Vmax was achieved in the presence of 1 μM acetylene by raising the NH4+ concentration, the apparent Km increased from ≤0.01 mM to 0.55 ± 0.08 mM NH4+. Although there are no published values of the Ks for NH4+/NH3 in these particular AOA isolates, a maximum velocity of NO2− production was achieved with 10 μM NH4+/NH3 in the absence of acetylene (Fig. 3A). When the responses of the rates of NO2− production to a range of acetylene and NH4+ concentrations were plotted in a double-reciprocal plot (Fig. 3B), the lines representing each inhibitor concentration had a different slope, also diagnostic of competitive inhibition. In contrast, there was no significant effect of NH4+ concentration (P > 0.07) on Vi at any inhibitory octyne concentration, indicating that octyne inhibition could not be overcome with the addition of higher concentrations of NH4+ (Fig. 3C). When the responses of NO2− production to a range of octyne and NH4+ concentrations were plotted in a double-reciprocal plot the intercepts on the y axis are distinctively different (Fig. 3D). This result would be expected if inhibition by octyne was noncompetitive with respect to NH4+.
FIG 3.

Response of initial velocities of NO2− production by N. viennensis to acetylene (A and B) and octyne (C and D) at increasing concentrations of NH4+. (A) Inhibition by 1 μM acetylene is overcome by increasing the NH4+ concentration. (B) Lineweaver-Burk double-reciprocal plot shows that the slopes for each acetylene concentration differ, indicating competitive inhibition. (C) Vi decreases with higher octyne concentrations, but octyne inhibition cannot be overcome by increasing NH4+. (D) Lineweaver-Burk double-reciprocal plots intercept the 1/V0 axis at different values indicating noncompetitive inhibition. Error bars represent the standard deviations of three replicates.
(ii) Examining the influence of octyne on the relationship between NH4+ concentration and acetylene on the inhibition of NO2− production by N. viennensis.
An additional experiment was conducted to confirm that the mechanisms of octyne and acetylene inhibition differed. We reasoned that if octyne does not interact at the same site as acetylene it should not affect the competitive behavior between acetylene and NH4+. N. viennensis was treated with octyne in some treatments before adding acetylene (Table 1). In the no-alkyne control Vmax was saturated by all NH4+ concentrations (0.25 to 2 mM NH4+, P > 0.3). When N. viennensis was treated with 1 μM acetylene, there was a statistically significant effect of NH4+ concentration on Vmax (P ≤ 0.005), as shown earlier (Fig. 3). In contrast, the addition of 30 μM octyne decreased Vmax (P < 0.01), but there was no significant protective effect of increasing NH4+ concentration on the rate of NO2− production (0.25 to 2 mM NH4+, P > 0.3). When N. viennensis was treated with octyne prior to acetylene, there was a statistically significant protective effect of increasing NH4+ concentrations on the rate of octyne-insensitive, acetylene-sensitive NO2− production (P ≤ 0.005), indicating that the competitive behavior between NH4+ and acetylene was not modified by the presence of octyne. This result gives additional support, suggesting that octyne is not functioning akin to acetylene as a competitive inhibitor of NH4+/NH3 oxidation.
TABLE 1.
Comparison of the response of rates of NO2− production by N. viennensis in a minus-alkyne control to treatments exposed to 1 μM acetylene, 30 μM octyne, or a combination of acetylene and octyne at different concentrations of NH4+a
| NH4+ concn (mM) | NO2− production (nmol/ml/h) after exposure to various alkyne treatments |
|||
|---|---|---|---|---|
| Control | Acetylene | Octyne | Acetylene + octyne | |
| 2.0 | 13.1 (1.9)A | 11.5 (2.5)A | 6.5 (2.6)A | 6.7 (1.1)A |
| 1.0 | 15.5 (3.8)A | 11.7 (2.4)A | 4.9 (1.3)A | 4.9 (0.6)A |
| 0.5 | 14.0 (2.3)A | 7.2 (1.7)AB | 5.1 (1.2)A | 1.6 (0.7)B |
| 0.25 | 16.9 (0.8)A | 4.1 (1.3)B | 4.9 (1.5)A | 1.0 (0.3)B |
The acetylene-octyne samples were exposed to 30 μM octyne for 15 min before the addition of 1 μM acetylene. Values in parentheses represent the standard deviations of three replicates. Different superscript letters indicate significant differences (P ≤ 0.03) between different NH4+ concentrations within a specific alkyne treatment. Values with superscript letters in common are not significantly different.
(iii) Time course of recovery of NO2− production in N. viennensis following inhibition by acetylene or octyne.
It is known that acetylene is an irreversible inactivator of AMO of AOA (12, 21) and AOB (22). Recovery from inactivation requires new AMO protein synthesis resulting in a lag of several hours for resumption of maximum NO2− production. Previously, Vajrala et al. (21) showed that the eukaryotic protein synthesis inhibitor, cycloheximide, prevented recovery of NO2− production in N. maritimus after acetylene inhibition. In the case of N. viennensis, the same concentration range of cycloheximide slowed but did not prevent recovery of NO2− production after inactivation with acetylene (data not shown). The lack of an unambiguous protein synthesis inhibitor for N. viennensis made us reliant upon the comparative time course of recovery of NO2− production after exposure to acetylene or octyne. After exposure of N. viennensis to either 6 μM acetylene or 40 μM octyne, NO2− production was inhibited to 0 or 21%, respectively, of the control (Fig. 4). After the alkynes were removed by degassing, the initial rates of NO2− production during the first hour of the recovery period averaged 32% (acetylene) and 96% (octyne) of the no-alkyne control, respectively, indicating that the majority of octyne inhibition was quickly reversible. In the case of acetylene, 3 h were required for the NO2− production rate to reach the rate of the control treatment. A “starved” control incubated without NH4+ for 2 h immediately accumulated NO2− at the same rate as the minus alkyne control upon the addition of NH4+ (data not shown).
FIG 4.
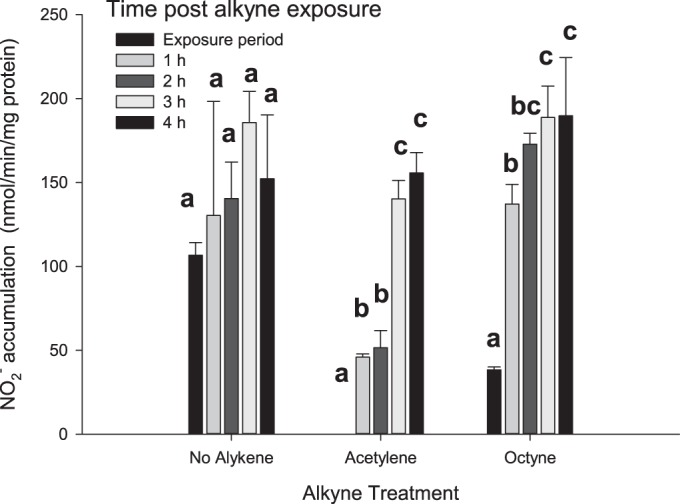
Response of NO2− production by N. viennensis after acetylene or octyne exposure. Cells were exposed for 2 h to either 6 μM acetylene or 40 μM octyne in media with 0.5 mM NH4+. Alkyne and no-alkyne treatments were then degassed and NH4+ increased to 2 mM. NO2− accumulation was monitored after alkyne exposure for 4 h. Bars represent the average activity in three replicate treatments. Error bars represent the standard deviations of the average. Lowercase letters indicate the significance (P ≤ 0.02) of sampling time within a specific treatment. Values that have lowercase letters in common are not significantly different.
DISCUSSION
In an earlier study, we showed that NO2− production by the marine AOA, N. maritimus (Thaumarchaeota group 1.1a) was very resistant to inhibition by ≥C7 1-alkynes in comparison to the AOB, N. europaea and Nitrosospira multiformis (12). In the present study, we confirm that NO2− production by pure culture isolates of N. viennensis strain EN76 (15, 16) and N. gargensis (10), both members of Thaumarchaeota group 1.1b, show the same overall trend, being highly sensitive to ≤C5 and much more resistant to longer-chain-length alkynes. They are also insensitive to concentrations of the C8 alkyne, octyne, that cause complete and irreversible inactivation of NH3 oxidation in AOB. These observations provide direct evidence that octyne-resistant NO2− production is a property of members of the phylogenetically distinct Thaumarchaeota group 1.1b and strengthens the idea that octyne might be exploited to differentiate between the relative activities of AOA and AOB in different natural environments where groups 1.1a and 1.1b are found.
Our observations that higher concentrations of octyne partially inhibit AOA activity in a manner that is (i) time independent, (ii) fully reversible, (iii) not competitive with NH4+, and (iv) without effect on the competitive interaction between NH4+ and acetylene raises the possibility that kinetic and catalytic properties of AMO from AOA and AOB might differ. For example, the observation that octyne inhibition of NO2− production in N. viennensis is noncompetitive with respect to NH4+ concentration (23), suggests that octyne binds to an alternate (non-NH3) binding site on AMO. This observation relates to earlier kinetic work of Keener et al. (24), who showed that, whereas NH4+ and acetylene competed for the same binding site on AMO of N. europaea, NH4+ concentration did not affect the rate of inactivation of N. europaea AMO by either 1- or 3-hexyne. These authors suggested that N. europaea AMO contained different binding sites or different access channels to the active site for small substrates (e.g., NH3, CH4, and acetylene) versus larger substrates (e.g., 1- and 3-hexyne). Indeed, Keener and Arp (25) also showed that most alternate substrates for N. europaea AMO are noncompetitive inhibitors of NH3 oxidation. The specific reason(s) why octyne inhibits rather than inactivates AOA AMO are unknown and require further study.
Our data on the inhibitory properties of alkynes of different chain length on AOA AMO are worthy of discussion in context with the topic of substrate range of the membrane-bound, Cu-containing methane monooxygenase (pMMO). Whereas it makes intuitive sense to compare characteristics of AMO from AOA and AOB, a suggestion has been made recently that AOA amo sequences are more closely related to the particulate methane monooxygenase (pmo) of the Verrucomicrobia phylum than to AOB amo sequences (26). It has been known for many years that pMMOs of the well-studied methanotrophs, Methylosinus trichosporium OB3b and Methylococcus capsulatus (Bath), exhibit a hydrocarbon substrate range that extends beyond CH4 and yet, are restricted to short-chain alkanes (C2 to C5) and alkenes (C2 to C4) (27–29). It has been suggested that the chain-length-limited substrate range of pMMO is due to the active site of the enzyme being located in a hydrophobic pocket that cannot accommodate alkanes of >C5, and there is speculation that the pocket might have separate binding sites for CH4 versus the larger substrates (28–31). Perhaps, it is not unreasonable to link our own data with the above-mentioned properties of pMMO to suggest that AOA AMO might also contain a binding pocket resembling pMMO that is either too small to freely accommodate ≥C6 1-alkynes or constrained sufficiently to allow binding but does not allow turnover of the longer 1-alkynes. Clearly, since AMO of N. europaea oxidizes C2 to C8 alkanes and is also inactivated by C2 to C8 1-alkynes, it is reasonable to conclude that the substrate range and catalytic properties of the two AMOs are different.
Furthermore, the reproducible differences in alkyne-chain-length sensitivity among the three isolates of Thaumarchaeota may also indicate differences in the properties of the AMO among the AOA. Whether such differences have ecological significance awaits further study.
Acetylene specifically and irreversibly inactivates AMO of N. europaea and under that inactivated condition N. europaea demonstrates hydroxylamine-dependent NO2− production at a rate identical to NH3-driven NO2− production (19, 20). N. maritimus also demonstrates hydroxylamine-dependent NO2− production when inactivated with acetylene (32), suggesting that acetylene inhibition is AMO specific, and led those authors to suggest that hydroxylamine is an intermediate in NH4+ oxidation in AOA, as well as AOB. However, we cannot rule out that partial inhibition by octyne occurs at a site other than AMO since we were unable to measure hydroxylamine-dependent NO2− production in cultures of N. viennensis either partially inhibited by octyne, inactivated with acetylene, or under NH4+-depleted conditions. Much remains to be resolved regarding the mechanism of NH3/NH4+ oxidation in AOA (33), and further work is needed to determine whether there are fundamental differences among the AOA.
ACKNOWLEDGMENTS
This research was supported by USDA NIFA award 2012-67019-3028, an Oregon Agricultural Research Foundation competitive grant, and the Agricultural Experiment Station. M.P. and M.W. were supported by a grant from the European Research Council (advanced grant NITRICARE 294343).
We thank N. Vajrala for valuable discussion on hydroxylamine experiments.
REFERENCES
- 1.Spang A, Poehlein A, Offre P, Zumbragel S, Haider S, Rychlik N, Nowka B, Schmeisser C, Lebedeva EV, Rattei T, Bohm C, Schmid M, Galushko A, Hatzenpichler R, Weinmaier T, Daniel R, Schleper C, Spieck E, Streit W, Wagner M. 2012. The genome of the ammonia-oxidizing Candidatus Nitrososphaera gargensis: insights into metabolic versatility and environmental adaptations. Environ Microbiol 14:3122–3145. doi: 10.1111/j.1462-2920.2012.02893.x. [DOI] [PubMed] [Google Scholar]
- 2.Pester M, Rattei T, Flechl S, Grongroft A, Richter A, Overmann J, Reinhold-Hurek B, Loy A, Wagner M. 2012. amoA-based consensus phylogeny of ammonia-oxidizing archaea and deep sequencing of amoA genes from soils of four different geographic regions. Environ Microbiol 14:525–539. doi: 10.1111/j.1462-2920.2011.02666.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Walker CB, de la Torre JR, Klotz MG, Urakawa H, Pinel N, Arp DJ, Brochier-Armanet C, Chain PSG, Chan PP, Gollabgir A, Hemp J, Hugler M, Karr EA, Konneke M, Shin M, Lawton TJ, Lowe T, Martens-Habbena W, Sayavedra-Soto LA, Lang D, Sievert SM, Rosenzweig AC, Manning G, Stahl DA. 2010. Nitrosopumilus maritimus genome reveals unique mechanisms for nitrification and autotrophy in globally distributed marine Crenarchaea. Proc Natl Acad Sci U S A 107:8818–8823. doi: 10.1073/pnas.0913533107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Shen T, Stieglmeier M, Dai J, Urich T, Schleper C. 2013. Responses of the terrestrial ammonia-oxidizing archaeon Ca. Nitrososphaera viennensis and the ammonia-oxidizing bacterium Nitrosospira multiformis to nitrification inhibitors. FEMS Microbiol Lett 344:121–129. doi: 10.1111/1574-6968.12164. [DOI] [PubMed] [Google Scholar]
- 5.Jung MY, Well R, Min D, Giesemann A, Park SJ, Kim JG, Kim SJ, Rhee SK. 2014. Isotopic signatures of N2O produced by ammonia-oxidizing archaea from soils. ISME J 8:1115–1125. doi: 10.1038/ismej.2013.205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yan J, Haaijer SCM, den Camp HJMO, van Niftrik L, Stahl DA, Konneke M, Rush D, Damste JSS, Hu YY, Jetten MSM. 2012. Mimicking the oxygen minimum zones: stimulating interaction of aerobic archaeal and anaerobic bacterial ammonia oxidizers in a laboratory-scale model system. Environ Microbiol 14:3146–3158. doi: 10.1111/j.1462-2920.2012.02894.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ginestet P, Audic JM, Urbain V, Block JC. 1998. Estimation of nitrifying bacterial activities by measuring oxygen uptake in the presence of the metabolic inhibitors allylthiourea and azide. Appl Environ Microbiol 64:2266–2268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hooper AB, Terry KR. 1973. Specific inhibitors of ammonia oxidation in Nitrosomonas. J Bacteriol 115:480–485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bédard C, Knowles R. 1989. Physiology, biochemistry, and specific inhibitors of CH4, NH4+, and CO oxidation by methanotrophs and nitrifiers. Microbiol Rev 53:68–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hatzenpichler R, Lebedeva EV, Spieck E, Stoecker K, Richter A, Daims H, Wagner M. 2008. A moderately thermophilic ammonia-oxidizing crenarchaeote from a hot spring. Proc Natl Acad Sci U S A 105:2134–2139. doi: 10.1073/pnas.0708857105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lehtovirta-Morley LE, Verhamme DT, Nicol GW, Prosser JI. 2013. Effect of nitrification inhibitors on the growth and activity of Nitrosotalea devanaterra in culture and soil. Soil Biol Biochem 62:129–133. doi: 10.1016/j.soilbio.2013.01.020. [DOI] [Google Scholar]
- 12.Taylor AE, Vajrala N, Giguere AT, Gitelman AI, Arp DJ, Myrold DD, Sayavedra-Soto L, Bottomley PJ. 2013. Use of aliphatic n-alkynes to discriminate soil nitrification activities of ammonia-oxidizing thaumarchaea and bacteria. Appl Environ Microbiol 79:6544–6551. doi: 10.1128/AEM.01928-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hyman MR, Murton IB, Arp DJ. 1988. Interaction of ammonia monooxygenase from Nitrosomonas europaea with alkanes, alkenes, and alkynes. Appl Environ Microbiol 54:3187–3190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gubry-Rangin C, Hai B, Quince C, Engel M, Thomson BC, James P, Schloter M, Griffiths RI, Prosser JI, Nicol GW. 2011. Niche specialization of terrestrial archaeal ammonia oxidizers. Proc Natl Acad Sci U S A 108:21206–21211. doi: 10.1073/pnas.1109000108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Tourna M, Stieglmeier M, Spang A, Konneke M, Schintlmeister A, Urich T, Engel M, Schloter M, Wagner M, Richter A, Schleper C. 2011. Nitrososphaera viennensis, an ammonia oxidizing archaeon from soil. Proc Natl Acad Sci U S A 108:8420–8425. doi: 10.1073/pnas.1013488108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Stieglmeier M, Klingl A, Alves RJE, Rittmann SK-MR, Melcher M, Leisch N, Schleper C. 2014. Nitrososphaera viennensis gen. nov., sp. nov., an aerobic and mesophilic, ammonia-oxidizing archaeon from soil and a member of the archaeal phylum Thaumarchaeota. Int J Syst Evol Microbiol 64:2738–2752. doi: 10.1099/ijs.0.063172-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Tourna M, Freitag TE, Nicol GW, Prosser JI. 2008. Growth, activity and temperature responses of ammonia-oxidizing archaea and bacteria in soil microcosms. Environ Microbiol 10:1357–1364. doi: 10.1111/j.1462-2920.2007.01563.x. [DOI] [PubMed] [Google Scholar]
- 18.Lebedeva EV, Hatzenpichler R, Pelletier E, Schuster N, Hauzmayer S, Bulaev A, Grigor'eva NV, Galushko A, Schmid M, Palatinszky M, Le Paslier D, Daims H, Wagner M. 2013. Enrichment and genome sequence of the group I.1a ammonia-oxidizing archaeon “Ca. Nitrosotenuis uzonensis” representing a clade globally distributed in thermal habitats. PLoS One 8:e80835. doi: 10.1371/journal.pone.0080835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hyman MR, Arp DJ. 1992. 14C2H2- and 14CO2-labeling studies of the de novo synthesis of polypeptides by Nitrosomonas europaea during recovery from acetylene and light inactivation of ammonia monooxygenase. J Biol Chem 267:1524–1545. [PubMed] [Google Scholar]
- 20.Hynes RK, Knowles R. 1978. Inhibition by acetylene of ammonia oxidation in Nitrosomonas europaea. FEMS Microbiol Lett 4:319–321. [DOI] [PubMed] [Google Scholar]
- 21.Vajrala N, Bottomley PJ, Stahl DA, Arp DJ, Sayavedra-Soto LA. 2014. Cycloheximide prevents the de novo polypeptide synthesis required to recover from acetylene inhibition in Nitrosopumilus maritimus. FEMS Microbiol Ecol 88:495–502. doi: 10.1111/1574-6941.12316. [DOI] [PubMed] [Google Scholar]
- 22.Hyman MR, Wood PM. 1985. Suicidal inactivation and labeling of ammonia monooxygenase by acetylene. Biochem J 227:719–725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Mathews C, van Holde KE, Ahern K. 2000. Biochemistry. Benjamin/Cummings, San Francisco, CA. [Google Scholar]
- 24.Keener WK, Russell SA, Arp DJ. 1998. Kinetic characterization of the inactivation of ammonia monooxygenase in Nitrosomonas europaea by alkyne, aniline, and cyclopropane derivatives. Biochim Biophys Acta 1388:373–385. [DOI] [PubMed] [Google Scholar]
- 25.Keener WK, Arp DJ. 1993. Kinetic studies of ammonia monooxygenase inhibition in Nitrosomonas europaea by hydrocarbons and halogenated hydrocarbons in an optimized whole-cell assay. Appl Environ Microbiol 59:2501–2510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tavormina P, Orphan V, Kalyuzhnaya M, Jetten MS, Klotz M. 2011. A novel family of functional operons encoding methane/ammonia monooxygenase-related proteins in gammaproteobacterial methanotrophs. Environ Microbiol Rep 3:91–100. doi: 10.1111/j.1758-2229.2010.00192.x. [DOI] [PubMed] [Google Scholar]
- 27.Burrows KJ, Cornish A, Scott D, Higgins IJ. 1984. Substrate specificities of the soluble and particulate methane mono-oxygenases of Methylosinus trichosporium OB3b. J Gen Microbiol 130:3327–3333. [Google Scholar]
- 28.Elliott SJ, Zhu M, Tso L, Nguyen HHT, Yip JHK, Chan SI. 1997. Regio- and stereoselectivity of particulate methane monooxygenase from Methylococcus capsulatus (Bath). J Am Chem Soc 119:9949–9955. [Google Scholar]
- 29.Culpepper MA, Rosenzweig AC. 2012. Architecture and active site of particulate methane monooxygenase. Crit Rev Biochem Mol Biol 47:483–492. doi: 10.3109/10409238.2012.697865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ng KY, Tu LC, Wang YS, Chan SI, Yu SSF. 2008. Probing the hydrophobic pocket of the active site in the particulate methane monooxygenase (pMMO) from Methylococcus capsulatus (Bath) by variable stereoselective alkane hydroxylation and olefin epoxidation. Chembiochem 9:1116–1123. doi: 10.1002/cbic.200700628. [DOI] [PubMed] [Google Scholar]
- 31.Miyaji A, Miyoshi T, Motokura K, Baba T. 2011. The substrate binding cavity of particulate methane monooxygenase from Methylosinus trichosporium OB3b expresses high enantioselectivity for n-butane and n-pentane oxidation to 2-alcohol. Biotechnol Lett 33:2241–2246. doi: 10.1007/s10529-011-0688-3. [DOI] [PubMed] [Google Scholar]
- 32.Vajrala N, Martens-Habbena W, Sayavedra-Soto L, Schauer A, Bottomley PJ, Stahl DA, Arp D. 2013. Hydroxylamine as an intermediate in ammonia oxidation by globally abundant marine archaea. Proc Natl Acad Sci U S A 110:1006–1011. doi: 10.1073/pnas.1214272110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Urakawa H, Martens-Habbena W, Stahl DA. 2011. Physiology and genomics of ammonia-oxidizing Archaea, p 117–155. In Ward BB, Arp D, Klotz M (ed), Nitrification. ASM Press, Washington, DC. [Google Scholar]