

Abstract
Fish schools are able to display a rich variety of collective states and behavioural responses when they are confronted by threats. However, a school's response to perturbations may be different depending on the nature of its collective state. Here we use a previously developed data-driven fish school model to investigate how the school responds to perturbations depending on its different collective states, we measure its susceptibility to such perturbations, and exploit its relation with the intrinsic fluctuations in the school. In particular, we study how a single or a small number of perturbing individuals whose attraction and alignment parameters are different from those of the main population affect the long-term behaviour of a school. We find that the responsiveness of the school to the perturbations is maximum near the transition region between milling and schooling states where the school exhibits multistability and regularly shifts between these two states. It is also in this region that the susceptibility, and hence the fluctuations, of the polarization order parameter is maximal. We also find that a significant school's response to a perturbation only happens below a certain threshold of the noise to social interactions ratio.
Keywords: fish school model, collective behaviour, self-organization, computational modelling
1. Introduction
Fish schools behave as coherent entities and display complex emergent properties such as coordinated motion, different ordered collective states and rapid escape manoeuvres when attacked by predators [1–5]. These group-level properties provide evolutionary advantages to fish schools and arise through social interactions by which individuals exchange information and perform specific behavioural responses such as changing their direction and velocity, or avoiding collision with group members or obstacles in the environment [6–12]. These interactions facilitate the transfer of information between fish and their ability to respond quickly to changes in the environment. For instance, when some fish spot a predator they abruptly change their direction of travel. Their close neighbours react in turn by changing their own velocity so that the information gradually propagates through the whole group, allowing all individuals to escape [6]. Experimental and theoretical works have shown that the same interactions can lead a few individuals having salient information, such as knowledge about the location of a food source or of a migration route, to guide other group members and bias the resulting direction of travel of the school [13–15]. In all these situations, a small proportion of individuals deeply influence the collective behaviour of the whole group. This is a direct consequence of the asymmetry of behavioural responses that exist between the perturbing or informed individuals and the other group members. Indeed, the behaviour of a perturbing or an informed fish is weakly influenced by the behaviour of its neighbours. While the decisions of the neighbours are mostly dictated by the behaviour of nearby fish.
Theoretical investigations have shown that not only the features of local interactions among individuals but also the number and position of neighbours to which a fish pays attention determine the patterns of collective motion that emerge at the group level [16–19]. For instance, we have recently shown in a data-driven fish school model that the relative weights of attraction and alignment interactions between fish give rise to a small number of specific collective states [20–22]: a swarming (disordered) state in which fish aggregate without cohesion, with a low level of polarization of their velocity; a schooling (ordered) state in which individuals are aligned with each other; a milling (ordered) state in which individuals constantly rotate around an empty core; and a winding (ordered) state in which the group exhibits an elongated phase characterized by a linear crawling motion. However, according to whether fish pay equal attention to their surrounding neighbours or focus their attention only of those neighbours that are ahead of them [22] (i.e. interactions depend on the neighbour angular position), the number of collective states that can be reached by a school is different. Only the swarming and schooling states survive when the behavioural reactions of fish do not depend on the angular position of their neighbours, while the full repertoire of collective states exists when a front/back asymmetry in fish interaction is introduced. Moreover, the exploration of the model has shown that in the transition region between milling and schooling states, the school exhibits multistability and regularly shifts from schooling to milling for the same combination of individual parameters, a property that was also reported in the model of Couzin et al. [18] and in experimental observations on groups of golden shiners [23].
While the past literature on schooling models has been devoted to a better understanding of the link between interaction rules and collective behaviours, less attention was paid to the group response and to its sensitivity to external perturbations [16,18–20,22,24–27]. In particular, one may wonder if a small number of perturbed or informed individuals could trigger the same response whatever the collective state of the school.
In the present work, we extensively study a previously developed data-driven model [21], which has been validated previously on actual experiments on Khulia mugil and was able to describe quantitatively several properties: individual trajectories of a single fish in a tank (validating in particular the noise and friction terms; see the Model section), interaction between two and 30 fish, diffusion properties, mean distance between fish, mean fish alignment/polarization versus the velocity or the number of fish in the tank. In another work [22], the phase diagram of the model without the tank boundaries (in free space) was studied as a function of the attraction and alignment parameters, reproducing several collective states observed in actual fish schools (see below). The excellent qualitative and quantitative accuracy of the model in describing real fish schools in a tank is a good motivation to consider it as a fair description of fish moving in a free space (in particular, their response to a perturbation) for which relevant experiments would be much harder to implement.
The present work hence addresses the response properties of a fish school in free space, using the model as presented in [22], and that we will briefly review below for completeness, but also to emphasize the crucial role played by the anisotropic angular perception of the fish to their environment. We investigate how a single or a small number of perturbed individuals affect the long-term behaviour of a school. In particular, we study how the school responds to perturbations depending on its different collective states and introduce their susceptibility to such perturbations. We relate the fish school response quantified by these susceptibilities to the fluctuations (for instance, of the polarization order parameter) already existing in the unperturbed fish school. This deep connection between response to a perturbation and intrinsic fluctuations without perturbation, although well understood and studied in the context of physical systems, is less familiar in the present context of fish schools, but equally applies. It is thus also one of the main purposes of the present work to illustrate and study qualitatively and quantitatively this general connection. In addition, we also exploit the fact that fluctuations and hence adequate susceptibilities are maximum (and would diverge in an infinite systems) at the transition between two (collective) states separated by a continuous phase transition.
We first analyse school susceptibility in the absence of any perturbation and its relation to fluctuations and to the identification of transition lines between the different collective states. We also explore the influence on group behaviour of a perturbing fish with an independent set of attraction and alignment parameters, while keeping the main population in the high susceptibility region, in order to determine the perturbations that have the highest impact. We finally explore the model parameter space to determine how the relative weighting of attraction and alignment of fish affects the school's responsiveness to perturbations. Finally, we discuss the implications of our work for real fish schools.
2. Model
The present model was originally proposed by Gautrais et al. [20,21] to describe the coordination of movements in groups of K. mugil through the use of stochastic equations of motion for their angular velocity ωi = dϕi/dt, while fish move with a constant speed along their angular direction given by ϕi. In a previous work [22], we introduced a non-dimensionalized version of the model in which we included an angular modulation of the strength of interactions between a fish and its neighbours according to their angular position, a property also supported by experiments [20,21], but of negligible consequence for fish in a not too large tank. This angular modulation breaks the symmetry of interactions between fish in front and those behind. For completeness, we briefly recall the main components of the model, where the individual angular velocity ωi evolves according to the following non-dimensional stochastic differential equation:
| 2.1 |
where α can be understood as an angular inertia term, 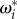 is the response function resulting from the interaction with the neighbouring fish (figure 1a) and dWi(t) refers to a random variable, uncorrelated in time and uniformly distributed in the interval [−1,1] (times
is the response function resulting from the interaction with the neighbouring fish (figure 1a) and dWi(t) refers to a random variable, uncorrelated in time and uniformly distributed in the interval [−1,1] (times 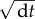 ). As a consequence of the large number theorem, such a uniformly distributed noise has exactly the same effect as the usual Gaussian noise (Wiener process) in the long run and in the limit of a time step dt → 0, with the benefit of being much faster to implement numerically.
). As a consequence of the large number theorem, such a uniformly distributed noise has exactly the same effect as the usual Gaussian noise (Wiener process) in the long run and in the limit of a time step dt → 0, with the benefit of being much faster to implement numerically.
Figure 1.

Graphical representation of fish interactions used in the model. (a) dij is the distance of fish j from fish i; ϕij is the relative orientation of fish j compared to fish i; θij is the angle between the angular position of fish j with respect to fish i. (b) Illustration of the Voronoi neighbourhood; arrows indicate fish headings. A focal fish is displayed in red and its Voronoi neighbours in orange. (c) Snapshots of typical configurations for the four distinct states displayed by the model.
The interaction is described by the normalized linear superposition of pair interactions between the focal fish and the first shell of Voronoi neighbours (figure 1b) as follows:
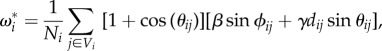 |
2.2 |
where θij is the angular position between the focal fish i and the neighbour j, Ni refers to the number of neighbouring fish in the first shell of the Voronoi tessellation Vi. The following terms correspond respectively to the alignment and attraction ‘forces', with β and γ controlling their intensities. ϕij = ϕj − ϕi is the heading angle difference and dij is the non-dimensional distance between j and the focal fish i. Originally, when investigating fish interactions, Gautrais et al. [21] considered different kinds of neighbourhoods to combine fish interactions. It was shown that besides the Voronoi neighbourhood, other choices were compatible with the experimental data in a tank, in particular the interactions between the k (k ≈ 6–8) nearest neighbours, which is remarkably similar to the average number of neighbours contained in the first shell of the Voronoi tessellation (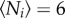 , exactly, in two dimensions).
, exactly, in two dimensions).
In equation (2.2), the prefactor Aij = 1 + cos(θij) modulates the amplitude of the interaction between fish i and j, as a function of the angle of view of the former. It is larger if j is ahead of i rather than behind and is hence maximum for θij = 0 (Aij = 2), minimum for θij = π (Aij = 0), while its angular average is 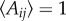 . This term also introduces a strong asymmetry between the force exerted by j on i and the one exerted by i on j, and hence breaks the (Newtonian) action–reaction principle which is most familiar in the context of purely physical force, such as gravitation. Apart from the technical fact that the alignment and attraction forces become non-conservative (i.e. not deriving from a potential energy), it has the practical and important consequence of allowing for the milling and winding phases, both observed in actual fish schools in the ocean (quasi-free space).
. This term also introduces a strong asymmetry between the force exerted by j on i and the one exerted by i on j, and hence breaks the (Newtonian) action–reaction principle which is most familiar in the context of purely physical force, such as gravitation. Apart from the technical fact that the alignment and attraction forces become non-conservative (i.e. not deriving from a potential energy), it has the practical and important consequence of allowing for the milling and winding phases, both observed in actual fish schools in the ocean (quasi-free space).
In [22], we have shown that varying the parameters β and γ of the model strongly affects the school's behaviour, leading to four distinct collective states (see the complete phase diagram in [22]): (I) schooling, (II) milling, (III) elongated winding state, and (IV) swarming, all of which can be visualized in figure 1c.
3. Quantification of collective behaviours and responses to perturbations
We now proceed to describe the tools used to characterize the collective states and to measure the susceptibility and the school's response to perturbations. We also describe the numerical details used for the simulations herein.
3.1. Order parameters
The aforementioned states can be quantified by two order parameters: (i) the polarization order parameter which provides a measure of how aligned the individuals in a group are
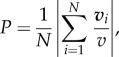 |
3.1 |
where P takes values between 0 (no alignment on average) and 1 (all fish are aligned); and (ii) the rotational or milling order parameter which provides a measure of the milling behaviour. It is the absolute value of the normalized angular momentum
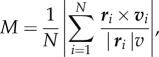 |
3.2 |
where 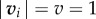 in the non-dimensional version of the model. M takes values between 0 (no collective rotation) and 1 (strong collective rotation of the school). The analysis on the transition line between the schooling and milling phases [22] shows that it obeys a simple functional form
in the non-dimensional version of the model. M takes values between 0 (no collective rotation) and 1 (strong collective rotation of the school). The analysis on the transition line between the schooling and milling phases [22] shows that it obeys a simple functional form 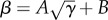 and that it is independent of the angular inertia term α from equation (2.1) considering values of equivalent of constant speeds between 0.4 and 1.2 m s−1.
and that it is independent of the angular inertia term α from equation (2.1) considering values of equivalent of constant speeds between 0.4 and 1.2 m s−1.
3.2. Susceptibilities and fluctuations
In many physical systems, some physical quantity may be coupled linearly and influenced by an external ‘field’. For instance, in a magnetic system, the atomic or electronic spins are coupled and tend to align along the direction of an external magnetic field. Similarly, an elastic medium (a spring or a rubber) can be elongated or compressed by exerting an external force. In general, the linear response of such a quantity P (for instance, the spin/fish polarization—i.e. the average spin/fish direction) to a small change in the associated external field h is quantified by introducing the P-susceptibility
| 3.3 |
In the present context of the dynamics of fish school, the change in the order parameter P (or M) to a small perturbation (of a nature detailed in the two next sections) will give a first characterization of the response of the system which will be presented in §§4.2 and 4.3.
Moreover, for a system at equilibrium or in a stationary state associated with an energy functional (an Hamiltonian), the fluctuation–dissipation theorem (FDT) [28] states that there exists a direct relation, in fact an exact equality up to a constant factor, between the P-susceptibility, as defined above by means of a small perturbing field, and the (thermal) fluctuations of P at equilibrium
| 3.4 |
where N is the number of particles, and the angular brackets refer to the average over time of the corresponding quantity, e.g.
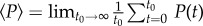 . In practice, in numerical simulations, t0 is of course finite and is taken as large as computation time permits. In addition, the susceptibility is also averaged over as many different samples/initial conditions as computationally possible. Note that the neighbouring spins/fish of a given spin/fish exert an effective magnetic/alignment field on the latter, making the connection between response and fluctuations very natural. In addition, the noise in equation (2.1) formally plays exactly the same role as the thermal noise in physics.
. In practice, in numerical simulations, t0 is of course finite and is taken as large as computation time permits. In addition, the susceptibility is also averaged over as many different samples/initial conditions as computationally possible. Note that the neighbouring spins/fish of a given spin/fish exert an effective magnetic/alignment field on the latter, making the connection between response and fluctuations very natural. In addition, the noise in equation (2.1) formally plays exactly the same role as the thermal noise in physics.
This powerful FDT has several very important implications, apart from the clear physical insight gained on the relations between the fluctuations and the response of a system. For instance, in numerical simulations (molecular dynamics or Monte Carlo), it is much easier and much more precise to measure the susceptibility from the fluctuations of the order parameter P (see the first equality in equation (3.4)), rather than applying a small field h, waiting for equilibrium to settle, measuring the (small) perturbation on P and ultimately trying to extrapolate to h = 0 (see the second quality in equation (3.3) and the original definition of equation (3.3)). In the present study of fish schools, we will first measure the susceptibility via the fluctuations of the system (first equality in equation (3.4)) and will in particular compare this susceptibility to the change of the order parameter under the addition of a few perturbing fish to the school (effectively acting as a small perturbing field).
As already mentioned, the asymmetric forces resulting from the Aij = 1 + cos(θij) term breaks the Newtonian action–reaction principle and forbids the existence of an underlying Hamiltonian. However, the FDT has been generalized in out of equilibrium situations, including in cases where the system, although in a stationary state, is not formally described by an energy functional (see [28] for a review). Hence, the connection between the fluctuations of the school polarization and the polarization response to a perturbation made in the present work appears very natural and will lead to important biological implications.
In the same manner, as we just defined the polarization susceptibility, we can define the milling susceptibility χm associated with the milling order parameter
| 3.5 |
Again, it should be intimately related to the change of M under a small perturbation defined hereafter, a relation that will be illustrated in the Results section and in the electronic supplementary material figures.
Finally, in the physical context, the susceptibility and hence fluctuations are known to diverge (at least in the limit of an infinite system N → ∞) exactly at the critical point between two phases separated by a continuous (second order) phase transition involving the considered order parameter. In the present context of fish schools, we will be naturally interested in the behaviour of the susceptibility and fluctuations near transition lines, in particular near the schooling–milling transition. The maximum of the susceptibility (characterized by fluctuations or response) as a function of the model parameters hence provides an alternative identification of the transition lines which will be illustrated extensively in §4.1 and in the electronic supplementary material figures.
3.3. Quantifying school response to perturbations
To detect behavioural changes, we compare the average values of the polarization and milling order parameters for a given set of alignment and attraction parameters of the unperturbed case, with the new average value given by the simulations with one or more perturbing fish. These results are presented for different combinations of the attraction and alignment parameters: complete parameter space scans, cross sections where the attraction parameter is kept fixed, and lastly, by the set of parameters which describe the transition between the milling and schooling state.
3.4. Simulations
We investigate the long-term consequences on the resulting school behaviour of a small number of perturbing fish that differ from the main population by having a different combination of attraction and alignment parameters. Henceforth, we call Np the number of perturbing fish and Nm = N − Np the main population of an N fish school. Accordingly, (γp, βp) and (γm, βm) are the attraction and alignment parameters, respectively, of the perturbing fish and the main population. All simulations were run in an unbounded space with 400 random initial conditions for 1000 non-dimensional time units, where the first half was discarded to remove transient states. A simple Euler's method integration with a time step of dt = 1.44 × 10−3 proved sufficient to avoid numerical imprecisions. Simulations results shown in the next sections were performed with N = 100 or 200, meanwhile the number of perturbing fish Np may vary from one to nine depending on the studied conditions. The electronic supplementary material, C.1, provides the source code from the algorithms used, together with all datasets from the figures presented.
4. Results
4.1. Susceptibility of a group of fish without perturbations
Having defined the susceptibility previously via the order parameter fluctuations (see equation (3.4)), we have calculated the polarization susceptibility values for different combinations of attraction and alignment parameters in a group of N = Nm = 100 and N = Nm = 200 fish, respectively, shown in figure 2a,b. One can clearly see that the highest values of susceptibility correspond perfectly to the fitted transition line between the schooling and milling regions. This indicates that the transition region is a good candidate to test different types of perturbing fish. Together with the susceptibility, other statistics of the unperturbed simulations were also computed, such as polarization and the milling order parameters, so that we can use some regions of this map as a baseline to measure the impact of perturbing fish on the school's behaviour.
Figure 2.

Susceptibility of unperturbed fish school simulations for (a) 100 and (b) 200 fish. The susceptibility is calculated according to equation (3.4) for different values of the attraction and alignment parameters. Each data point represents an average over 400 simulations with random initial conditions. The white lines following the peak of susceptibility represents the function that fits the schooling/milling transition line as reported in [22]. The circled numbers indicate the four different collective states (I) schooling, (II) milling, (III) winding, (IV) swarming and the transition zone between schooling and milling (I–II).
In §4.3 (see in particular figure 5), and in the supplementary material (by considering the milling susceptibility χm), we will indeed find a very strong correlation between the susceptibility computed from the fluctuations in the unperturbed system, and the response properties of the system under small perturbations.
Figure 5.

Difference in the average polarization (left axis) in groups of 100 (a) and 200 (b) fish, with 1, 3, 5, 7 and 9 (light to dark blue) perturbing fish (γp = 14, βp = 1), as a function of the alignment parameter of the main population (βm[0,16]), keeping the attraction parameter of the main population to a constant value (γm = 10). The black line represents the susceptibility values (right axis) for the unperturbed condition. The insets show the maximum difference in average polarization as a function of the number of perturbing fish.
4.2. Quantification of perturbations
We now proceed to investigate the impact of various combinations of attraction and alignment parameters of a single perturbing fish on the resulting group behaviour. The main population is located in the transition region (γm = 14, βm = 10) while the parameters γp and βp of the perturbing fish both vary in the interval [0,16] with a 0.4 step, forming an uniform 41 × 41 grid in the parameters space. Figure 3 shows the resulting difference in the average polarization P. Note that as the main population is located in the transition region, the values of the polarization P and milling M parameters fluctuate around 0.5. This means that in the case where the group changes to an almost perfect schooling state (P ≈ 1), the maximum difference is 0.5. Figure 3 shows that this change to a schooling state happens for both low attraction and low alignment values (γp and βp < 2).
Figure 3.

Each data point represents the difference in the average polarization caused by a single perturbing fish, while varying its attraction (γp) and alignment (βp) parameters, in comparison to the unperturbed case located in the transition region (γm = 14, βm = 10). The six insets shown here represent the location distribution of the positions of the perturbing fish in relation to the group's centroid (white circle), and reoriented according to the average direction movement of the group (white arrow).
One can easily understand that for low values of γp, corresponding to a weak attraction, the perturbing fish will not remain as close to the group as the others, forcing them to follow it, and in doing so, increasing the polarization of the group. For low values of βp, fish can have the same attraction parameters as the main population, but the weak alignment disrupts the milling intermittence completely. The six insets in figure 3 represent the distribution of perturbing fish locations with respect to the group's centroid (white circle), and reoriented according to the average direction movement of the school (white arrow). Inset 1 shows the unperturbed case (γm = γp and βm = βp), where we can see that the perturbing fish has an equal distribution all around the school. One can see in insets 2–4 (and in electronic supplementary material, videos B.1 and B.2) that when there is a low attraction (γp = 1), the perturbing fish stays ahead of the group most of the time. This ‘leading’ behaviour in which the school is attracted by the perturbing fish, but not the opposite, is a consequence of the smaller attraction of the perturbing fish compared with the main population (γp < γm). Higher values of γp combined with a weak alignment (γp = 7 or 14, and βp = 1) lead the perturbing fish to stay usually behind the group's centroid (insets 5 and 6 and electronic supplementary material, video B.3). It is also important to highlight that in the latter case the perturbing fish is much closer to the group's centroid in comparison to the conditions shown in the insets 2 through 4. The reason for the perturbing fish to remain close to and behind the school's centroid is quite simple. The high attraction ensures that the perturbing fish remains close to the school, but with low alignment, it is unable to cope with the directional changes of the other fish. As a consequence, it remains behind the school.
We also performed the same systematic analysis of the impact of a perturbing fish on the group's behaviour when the main population is in the schooling or the milling state. Electronic supplementary material, figures A.2 and A.3, show the results of these simulations. When the main population is located in the schooling region (γm = 4, βm = 14), a perturbing fish causes almost no change to the group's behaviour. When the population is located in the milling region (γm = 14, βm = 4), one can observe a change from milling to schooling for low parameter values of the perturbing fish (γp and βp < 2) and intermittent transition between schooling and milling appear for low attraction and high alignment values (γp < 2 and βp > 10). As explained previously, these effects in the milling region are only due to a fish that is not able to remain close to the group, forcing its neighbours to follow him, and in doing so, disrupting the mill, as shown in figure 3 (insets 2–4). It is worth noting that: (i) the only observed change in the group's behaviour resulting from the presence of a perturbing fish is a transition to schooling; (ii) only a perturbing fish with very low attraction values (γp ≈ 0) is able to disrupt a group engaged in a milling state; any other behaviour of the perturbing fish has no effective impact on the group tendency to rotate.
4.3. Group response to perturbations
Considering the results shown in figure 3, we have chosen a configuration of parameters for the perturbing fish (γp = 14 and βp = 1) which lies in the winding region (inset 6). The choice is motivated to prevent the perturbing effects to be the simple consequence of non-gregarious fish (γp ≈ 0), like the situations depicted in insets 2–4 in figure 3. We now proceed to analyse the group's response to this perturbation for different configurations of the main population parameter space. We focus on a cross section of the parameter space keeping a fixed value of γm = 10 and varying βm in the interval [0,16] represented by the vertical purple line in figure 4a. Highlighted in figure 4b are typical time series of the polarization and milling order parameters in the schooling region (I), the transition region (I–II) and the milling region (II). One can see that in the transition region, simulations with the perturbing fish display a change to a purely schooling behaviour.
Figure 4.

(a) Transition functions between schooling and milling fitted for simulations with 100 and 200 fish 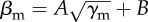 , where [A = 3.22, B = −2.23] for 100 fish and [A = 3.28, B = −1.17] for 200 fish. (b) Time series of polarization P and M milling parameters when the main population is in three different states: schooling (γm = 4, βm = 14), milling (γm = 14, βm = 4) and the transition between both states (γm = 14, βm = 10), for unperturbed condition (red line) and with a single perturbing fish introduced in the group (blue line, γp = 10, βp = 1).
, where [A = 3.22, B = −2.23] for 100 fish and [A = 3.28, B = −1.17] for 200 fish. (b) Time series of polarization P and M milling parameters when the main population is in three different states: schooling (γm = 4, βm = 14), milling (γm = 14, βm = 4) and the transition between both states (γm = 14, βm = 10), for unperturbed condition (red line) and with a single perturbing fish introduced in the group (blue line, γp = 10, βp = 1).
We can now investigate how an increasing number of perturbing fish affect group behaviour. We performed simulations with Np = 1, 3, 5, 7 and 9 perturbing fish (and Nm = 99, 97, 95, 93 and 91, when N = 100 fish, and Nm = 199, 197, 195, 193 and 191, in simulations with groups of N = 200 fish).
Figure 5 shows the resulting difference in the average polarization induced by the perturbation in comparison to the unperturbed condition in groups of 100 and 200 fish, respectively. In both cases, the resulting change in the group polarization and the susceptibility follow a similar pattern reaching a peak in the transition region. A smaller peak can also be seen for values of low βm. This peak is related to the transition from the milling zone to the winding region.
Increasing the number of perturbing fish leads to an increase in the polarization of the group (linear perturbation regime) up to a saturation value (3 < Np < 5). Ultimately, it shows a steady decline with more perturbing fish. This happens due to the fact that the perturbing fish have a lower alignment parameter value, meaning that after the initial perturbing effect, they will have a negative impact (nonlinear perturbation regime) on the average polarization of the school. While 100 and 200 fish simulations show a very similar pattern of change in group polarization for all quantities of perturbing fish, simulations with 200 fish display a systematic lower response due to the perturbing fish.
One might note the difference in two orders of magnitude between the difference in average polarization and susceptibility. This comes from the fact that the susceptibility is proportional to NP2, while the difference in average polarization is only proportional to P. Also, our main concern here is to use the susceptibility as a reference point to which the group responses are compared.
Having established that the transition zone is the region of the parameter space in which a fish group displays the highest responsiveness to perturbations, we have studied the group's response throughout this region. As previously seen [22], the transition between schooling and milling follows the functional form 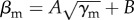 , where A and B fitted the parameter space data in which the school presented both polarization and milling parameter values above 0.8 more than 40% of the time (figure 4). We can systematically vary the attraction parameter γm within the range [0,16] and determine the parameter βm estimated by this procedure. In the following analysis, the parameters of the single perturbing fish are kept unchanged (γp = 14 and βp = 1).
, where A and B fitted the parameter space data in which the school presented both polarization and milling parameter values above 0.8 more than 40% of the time (figure 4). We can systematically vary the attraction parameter γm within the range [0,16] and determine the parameter βm estimated by this procedure. In the following analysis, the parameters of the single perturbing fish are kept unchanged (γp = 14 and βp = 1).
Figure 6 shows that even for a range of parameters where the susceptibility has already reached a maximum value, the difference in average polarization still increases with the attraction parameter γm before it starts oscillating around the values shown at γm = 16. This means that while being in the transition region is a required condition for a group of fish to exhibit sensitivity to perturbations, a minimum level of attraction and alignment between fish is required to significantly alter the group's response to these perturbations. This additional requirement is probably due to the fact that both γm and βm increase while keeping a constant noise, indicating that the main population only reacts to the perturbation when the ratio noise to social interactions is kept below a certain threshold.
Figure 6.

Difference in average polarization for 100 and 200 fish (light blue and orange filled circle, respectively) with a single perturbing fish (γp = 14 and βp = 1), along the transition region between schooling and milling defined by 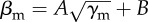 . A and B have been estimated for different group sizes: where A = 3.08, B = −1.97 for N = 100 and A = 3.28, B = −1.17 for N = 200. The corresponding susceptibility for unperturbed fish of 100 and 200 fish is shown on the right axis (blue and red hollow circle, respectively).
. A and B have been estimated for different group sizes: where A = 3.08, B = −1.97 for N = 100 and A = 3.28, B = −1.17 for N = 200. The corresponding susceptibility for unperturbed fish of 100 and 200 fish is shown on the right axis (blue and red hollow circle, respectively).
5. Discussion
How interactions between individuals control the sensitivity to perturbations of the group to which they belong and its ability to respond to threats is an important issue to understand the evolution of collective behaviours in animal swarms. The survival of each individual within the group strongly depends on the capacity of individuals to perform collective adaptive responses to different conditions. Performing such responses not only requires coordination mechanisms but also a high responsiveness to perturbations at the group level, which can be favoured by the presence of highly correlated fluctuations in the unperturbed state (illustrating again the deep connection between response and fluctuation) [29,30].
Here, we have addressed this question through an extensive investigation of the responsiveness of a fish school model to long-term standardized perturbations in the form of a single or a small number of fish that display different interactions than the main population in the school. We show that the school's response depends not only on the characteristics of the perturbing fish, but of the collective state of the school as well. Indeed, in the parameter space defining the way fish interacts with their neighbours, there is a region that maximizes the school response to perturbations. This region is located throughout the transition between schooling and milling states, where the school exhibits multistability and regularly shifts between both states, and where fluctuations are hence maximal.
The perturbing fish consists of agent(s) with intensities of the attraction and alignment behaviours that differ from the rest of the group. Borrowing the concept of susceptibility from magnetic systems and other analyses of collective behaviour in biological systems [31,32], we analysed its equivalent in our simulations to measure the group's behavioural change caused by the perturbing fish. We found that groups of fish display the highest susceptibility in the transition region between the schooling and milling states (figure 2). If one assumes that our fish are indeed in a transition region, the results presented here could be compared to recent works [29,32,33] which have shown that animal swarms are in a critical state to better adapt to various environmental conditions.
Indeed, the evolutionary advantages of social behaviour in animals can easily be reduced if the organisms fail to adapt rapidly and/or efficiently to a new challenging situation, for instance, in case of a predator attack. It has been argued for some time that a more probable solution for this problem is for a biological system to stay in a perpetual state of transition from the most common behaviour available, close to criticality [29,32–35], so that a minimal effort on its part is able to push the collective behaviour into the new and more appropriate one. Despite this, the system should also be steady/robust enough to ignore certain perturbations and avoid unnecessary transitions.
Our systematic study of the impact of perturbations in the parameter space reveals that a group of fish in the transition zone is highly affected by a perturbing fish with low attractions and/or alignment values (figure 3). When choosing which set of parameters to use for the perturbing fish, we avoided the trivial case where low attraction values coupled with the Voronoi neighbourhood causes a following behaviour. For this reason, we used a set of parameters that induced the perturbing fish to be located closer to the group's centroid. We found that the group responds to this perturbation by significantly increasing its level of polarization, shifting from a state in which the group spends half the time in the schooling and milling states to a new state where the group is schooling permanently (figures 4 and 5a,b).
We also checked whether the transition region is the only factor involved in the group's responsiveness. As shown in figure 6, even in the transition region (maximum susceptibility), at lower values of γm (and low βm), the school does not show the same responsiveness. The main difference in this region compared to other transitions points is the value of noise to social interactions ratio. This is in agreement with the observed lower responsiveness of the school as the number of perturbing fish is increased (insets of figure 5a,b). This can be seen as an unnecessarily large amount of noise, which cancels the impact of the perturbation and decreases the noise to social interactions ratio, as if the main population of the school was located in lower values of the transition.
In this context, it is also interesting to mention the work of Ioannou et al. [36], who studied the predatory tactics of a fish towards a virtual school with different attraction and alignment parameters. They found that the most frequently attacked fish (fig. 2 of [36]) have low attraction and/or low alignment parameters like the most influential perturbing fish in our model (figure 3). The fact that this parameter region (low attraction and/or alignment) corresponds to a vulnerability of the species (in Ioannou's work) and to a large capacity to change the behaviour of the whole school, suggests that when such deviant behaviour is detected it automatically triggers the other group members to flee either from the current location and/or from the vulnerable perturbing fish itself.
Previous works [13,26] had already studied the impact of perturbations in fish schools, but they focused on punctual or instantaneous perturbations, while we looked into the long-term changes that result from the perturbation. These analyses were also mainly related to changes in the school trajectory and did not focus on the main behavioural changes undergone by the school. When studying perturbations, one can either study how a punctual change immediately affects the system, and the subsequent recovering of the system to its original state, or one can investigate what is the minimal constant perturbation imposed on the system which is able to completely change its properties.
A similar approach to this continuous perturbation analysis has been presented by Aureli & Porfiri [27], where they used a self-propelled particle model to study the effect of an external leader particle. However, Aureli et al. chose a perturbing agent which is totally independent of the school's reactions. This choice also enabled them to compare their results with experiments performed with a remote-controlled robotic fish able to influence a school of giant danios (Devario aequipinnatus) [37]. Despite these similarities, there are two main differences between the two models: (i) the total independence of the perturbing agent, equivalent here to γp = βp = 0; (ii) the movement of the perturbing agent itself differs completely from the school. More specifically, the perturbing agent and the school can have different speeds. These differences limit the comparisons between the two approaches. Nevertheless, for the case where the perturbing agent has a similar speed to the rest of the particles, they observed an increase in polarization similar to the one we have found in figure 3, and electronic supplementary material, figures A.2 and A.3, for the case γp = βp = 0. Unfortunately, the observed state in their simulations and experiments of agents/fish milling around the perturbing agent cannot be reproduced here due to the fact that all fish have the same speed. Given the model dependence on the fish distance, in the case where the perturbing fish was fixed at some point, the main population would be forced to remain close, either swarming or milling around it, depending on the attraction parameters γm used.
In order to best preserve the data-driven model developed by Gautrais et al. [20,21], certain limitations to the analysis arise. For instance, one can mention the recent work by Couzin and co-workers [12] which has shown the importance of speed variation for the fish collective response. However, in their original work, Gautrais et al. [20] observed that speed variation was minimal and could be neglected for the considered species. Another limitation results from the size of the school, given the dependence on fish distance for their interactions. This term causes a limitation to the size of the school as seen in our previous work [22], limiting simulations around the sizes of 100 and 200 fish presented here.
In conclusion, our work has revealed that the collective states of a school deeply influence its ability to respond to external or internal perturbations. By providing a high responsiveness to perturbations, the transition region between milling and schooling appears to be a highly desired state that optimizes the ability of the fish to react collectively (e.g. to a predator attack), thus increasing the survival of each individual within the school. Our results call for further experimental observations on fish schools in order to measure both their susceptibility and responsiveness to perturbations.
Supplementary Material
Acknowledgements
We are grateful to J. Gautrais for inspiring discussions. We thank the Laboratoire Plasma et Conversion d'Energie (CNRS UMR 5283) and the Laboratoire de Physique Théorique (CNRS UMR 5152) for providing us access to their computing facilities.
Funding statement
D.S.C. was funded by the Conselho Nacional de Desenvolvimento Científico e Tecnológico â Brazil. U.L. was supported by a doctoral fellowship from the scientific council of the University Paul Sabatier. This study was supported by grants from the Centre National de la Recherche Scientifique and University Paul Sabatier (project Dynabanc).
References
- 1.Breder CM. 1951. Studies on the structure of fish schools. Bull. Am. Mus. Nat. Hist. 98, 3–27. [Google Scholar]
- 2.Radakov D. 1973. Schooling in the ecology of fish. New York, NY: J. Wiley. [Google Scholar]
- 3.Partridge BL. 1982. The structure and function of fish schools. Sci. Am. 246, 114–123. ( 10.1038/scientificamerican0682-114) [DOI] [PubMed] [Google Scholar]
- 4.Treherne J, Foster W. 1981. Group transmission of predator avoidance-behavior in a marine insect: the Trafalgar effect. Anim. Behav. 29, 911–917. ( 10.1016/S0003-3472(81)80028-0) [DOI] [Google Scholar]
- 5.Domenici P, Standen E, Levine R. 2004. Escape manoeuvres in the spiny dogfish (Squalus acanthilas). J. Exp. Biol. 207, 2339–2349. ( 10.1242/jeb.01015) [DOI] [PubMed] [Google Scholar]
- 6.Krause J, Ruxton G. 2002. Living in groups, Oxford Series in Ecology and Evolution. Oxford, UK: Oxford University Press. [Google Scholar]
- 7.Treherne JE, Foster WA. 1980. The effects of group size on predator avoidance in a marine insect. Anim. Behav. 28, 1119–1122. ( 10.1016/S0003-3472(80)80100-X) [DOI] [Google Scholar]
- 8.Foster WA, Treherne JE. 1981. Evidence for the dilution effect in the selfish herd from fish predation on a marine insect. Nature 293, 466–467. ( 10.1038/293466a0) [DOI] [Google Scholar]
- 9.Magurran A. 1990. The adaptive significance of schooling as an anti-predator defence in fish. Ann. Zool. Fenn. 27, 51–66. [Google Scholar]
- 10.Domenici P, Batty R. 1997. Escape behaviour of solitary herring (Clupea harengus) and comparisons with schooling individuals. Mar. Biol. 128, 29–38. ( 10.1007/s002270050065) [DOI] [Google Scholar]
- 11.Herbert-Read J, Perna A, Mann R, Schaerf T, Sumpter DJT, Ward AJW. 2011. Inferring the rules of interaction of shoaling fish. Proc. Natl Acad. Sci. USA 108, 18 726–18 731. ( 10.1073/pnas.1109355108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Katz Y, Tunstrøm K, Ioannou C, Huepe C, Couzin ID. 2011. Inferring the structure and dynamics of interactions in schooling fish. Proc. Natl Acad. Sci. USA 108, 18 720–18 725. ( 10.1073/pnas.1107583108) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Romey WL. 1996. Individual differences make a difference in the trajectories of simulated schools of fish. Ecol. Model. 92, 65–77. ( 10.1016/0304-3800(95)00202-2) [DOI] [Google Scholar]
- 14.Huse G, Railsback S, Feronö A. 2002. Modelling changes in migration pattern of herring: collective behaviour and numerical domination. J. Fish Biol. 60, 571–582. ( 10.1111/j.1095-8649.2002.tb01685.x) [DOI] [Google Scholar]
- 15.Couzin I, Krause J, Franks N, Levin S. 2005. Effective leadership and decision-making in animal groups on the move. Nature 433, 513–516. ( 10.1038/nature03236) [DOI] [PubMed] [Google Scholar]
- 16.Aoki I. 1982. A simulation study on the schooling mechanism in fish. Bull. J. Soc. Sci. Fish 48, 1081–1088. ( 10.2331/suisan.48.1081) [DOI] [Google Scholar]
- 17.Huth A, Wissel C. 1992. The simulation of the movement of fish schools. J. Theor. Biol. 156, 365–385. ( 10.1016/S0022-5193(05)80681-2) [DOI] [Google Scholar]
- 18.Couzin I, Krause J, James R, Ruxton G, Franks N. 2002. Collective memory and spatial sorting in animal groups. J. Theor. Biol. 218, 1–11. ( 10.1006/jtbi.2002.3065) [DOI] [PubMed] [Google Scholar]
- 19.Viscido SV, Parrish JK, Grünbaum D. 2005. The effect of population size and number of influential neighbors on the emergent properties of fish schools. Ecol. Model. 183, 347–363. ( 10.1016/j.ecolmodel.2004.08.019) [DOI] [Google Scholar]
- 20.Gautrais J, Jost C, Soria M, Campo A, Motsch S, Fournier R, Blanco S, Theraulaz G. 2009. Analyzing fish movement as a persistent turning walker. J. Math. Biol. 58, 429–445. ( 10.1007/s00285-008-0198-7) [DOI] [PubMed] [Google Scholar]
- 21.Gautrais J, Ginelli F, Fournier R, Blanco S, Soria M, Chaté H, Theraulaz G. 2012. Deciphering interactions in moving animal groups. PLoS Comp. Biol. 8, e1002678 ( 10.1371/journal.pcbi.1002678) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Calovi DS, Lopez U, Ngo S, Sire C, Chaté H, Theraulaz G. 2014. Swarming, schooling, milling: phase diagram of a data-driven fish school model. New J. Phys. 16, 15026 ( 10.1088/1367-2630/16/1/015026) [DOI] [Google Scholar]
- 23.Tunstrøm K, Katz Y, Ioannou C, Huepe C, Lutz M, Couzin I. 2013. Collective states, multistability and transitional behavior in schooling fish. PLoS Comp. Biol. 9, e1002915 ( 10.1371/journal.pcbi.1002915) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hemelrijk CK, Hildenbrandt H. 2008. Self-organized shape and frontal density of fish schools. Ethology 114, 245–254. ( 10.1111/j.1439-0310.2007.01459.x) [DOI] [Google Scholar]
- 25.Hemelrijk CK, Hildenbrandt H, Reinders J, Stamhuis EJ. 2010. Emergence of oblong school shape: models and empirical data of fish. Ethology 116, 1099–1112. ( 10.1111/j.1439-0310.2010.01818.x) [DOI] [Google Scholar]
- 26.Kolpas A, Busch M, Li H, Couzin ID, Petzold L, Moehlis J. 2013. How the spatial position of individuals affects their influence on swarms: a numerical comparison of two popular swarm dynamics models. PLoS ONE 8, e58525 ( 10.1371/journal.pone.0058525) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Aureli M, Porfiri M. 2010. Coordination of self-propelled particles through external leadership. EPL 92, 40004 ( 10.1209/0295-5075/92/40004) [DOI] [Google Scholar]
- 28.Marconi UMB, Puglisi A, Rondoni L, Vulpiani A. 2008. Fluctuation–dissipation: response theory in statistical physics. Phys. Rep. 461, 111–195. ( 10.1016/j.physrep.2008.02.002) [DOI] [Google Scholar]
- 29.Attanasi A, et al. 2014. Finite-size scaling as a way to probe near-criticality in natural swarms. Phys. Rev. Lett. 113, 238102 ( 10.1103/PhysRevLett.113.238102) [DOI] [PubMed] [Google Scholar]
- 30.Chaté H, Muñoz M. 2014. Insect swarms go critical. Physics 7, 120 ( 10.1103/Physics.7.120) [DOI] [Google Scholar]
- 31.Cavagna A, Cimarelli A, Giardina I, Parisi G, Santagati R, Stefanini F, Viale M. 2010. Scale-free correlations in starling flocks. Proc. Natl Acad. Sci. USA 107, 11 865–11 870. ( 10.1073/pnas.1005766107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Attanasi A, et al. 2014. Collective behaviour without collective order in wild swarms of midges. PLoS Comput. Biol. 10, e1003697 ( 10.1371/journal.pcbi.1003697) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bialek W, Cavagna A, Giardina I, Mora T, Pohl O, Silvestri E, Viale M, Walczak AM. 2014. Social interactions dominate speed control in poising natural flocks near criticality. Proc. Natl Acad. Sci. USA 111, 7212–7217. ( 10.1073/pnas.1324045111) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mora T, Bialek W. 2011. Are biological systems poised at criticality? J. Stat. Phys. 144, 268–302. ( 10.1007/s10955-011-0229-4) [DOI] [Google Scholar]
- 35.Boedecker J, Obst O, Lizier J, Mayer N, Asada M. 2012. Information processing in echo state networks at the edge of chaos. Theor. Biosci. 131, 205–213. ( 10.1007/s12064-011-0146-8) [DOI] [PubMed] [Google Scholar]
- 36.Ioannou CC, Guttal V, Couzin ID. 2012. Predatory fish select for coordinated collective motion in virtual prey. Science 337, 1212–1215. ( 10.1126/science.1218919) [DOI] [PubMed] [Google Scholar]
- 37.Aureli M, Fiorilli F, Porfiri M. 2012. Portraits of self-organization in fish schools interacting with robots. Phys. D 241, 908–920. ( 10.1016/j.physd.2012.02.005) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.