

Abstract
Helicobacter pylori is a helical-shaped, gram negative bacterium that colonizes the human gastric niche of half of the human population1,2. H. pylori is the primary cause of gastric cancer, the second leading cause of cancer-related deaths worldwide3. One virulence factor that has been associated with increased risk of gastric disease is the Cag-pathogenicity island, a 40-kb region within the chromosome of H. pylori that encodes a type IV secretion system and the cognate effector molecule, CagA4,5. The Cag-T4SS is responsible for translocating CagA and peptidoglycan into host epithelial cells5,6. The activity of the Cag-T4SS results in numerous changes in host cell biology including upregulation of cytokine expression, activation of proinflammatory pathways, cytoskeletal remodeling, and induction of oncogenic cell-signaling networks5-8. The Cag-T4SS is a macromolecular machine comprised of sub-assembly components spanning the inner and outer membrane and extending outward from the cell into the extracellular space. The extracellular portion of the Cag-T4SS is referred to as the “pilus”5. Numerous studies have demonstrated that the Cag-T4SS pili are formed at the host-pathogen interface9,10. However, the environmental features that regulate the biogenesis of this important organelle remain largely obscure. Recently, we reported that conditions of low iron availability increased the Cag-T4SS activity and pilus biogenesis. Here we present an optimized protocol to grow H. pylori in varying conditions of iron availability prior to co-culture with human gastric epithelial cells. Further, we present the comprehensive protocol for visualization of the hyper-piliated phenotype exhibited in iron restricted conditions by high resolution scanning electron microscopy analyses.
Keywords: Infection, Issue 93, Helicobacter pylori, iron acquisition, cag pathogenicity island, type IV secretion, pili
Introduction
H. pylori infection is a significant risk factor for gastric cancer1. However, disease outcomes vary and depend on numerous factors such as host genetics, genetic diversity of H. pylori strains, and environmental elements such as host diet11. Previous reports have established that a correlation exists between H. pylori infection, iron deficiency (as measured by decreased blood ferritin and hemoglobin concentrations), and increased proinflammatory cytokine production, including IL-8 secretion, which ultimately leads to increased gastric disease progression12. Acute H. pylori infection is also associated with hypochlorhydria which impairs the host’s ability to absorb nutrient iron, and ultimately leads to changes in iron homeostasis13. These clinical findings suggest that iron availability within the gastic niche could be an important factor in disease outcome. In fact, animal models of H. pylori infection have demonstrated that low dietary iron consumption exacerbates gastric disease14. The reduced iron levels in these animals necessitate that H. pylori induce an iron-acquisition response in order to obtain the iron needed for bacterial replication. H. pylori has the capacity to perturb iron trafficking within host cells to facilitate bacterial replication in a CagA-dependent fashion15. Interestingly, the cag-pathogenicity island has been shown to be regulated by the iron-responsive transcription factor Fur16,17. Furthermore, Cag+ strains are associated with increased inflammation and gastric diseases such as cancer1. These findings support a model whereby H. pylori alters Cag-T4SS expression in an effort to obtain iron from host cells that reside in an iron deplete environment resulting in exacerbated disease outcomes.
Two factors that increase inflammation and morbidity are Cag expression and low dietary iron intake. These facts support the hypothesis that reduced iron availability increases the production of Cag-T4SS pili at the host pathogen interface resulting in worse gastric disease11-14. The goal of the method provided in this manuscript is to establish the role of the micronutrient iron in the regulation of the Cag-T4SS pilus biogenesis. In previous work, we utilized two approaches to observe an iron-dependent increase in Cag-T4SS expression. First, output strains from animals maintained on high and low iron diets were analyzed and revealed that low-iron diet output strains produced more Cag-T4SS pili than high-iron diet strains14. Second, growing the H. pylori 7.13 strain in vitro in iron replete conditions resulted in reduced pili formation while cells grown in the presence of an iron chelator produced significantly more pili.
We have continued to investigate the iron-dependent regulation of Cag-T4SS pili phenotype and offer the following optimized protocol and representative results performed with an additional Helicobacter pylori strain, PMSS1. The rationale behind the development of this technique was to correlate increased Cag-T4SS activity in conditions of iron-limitation with increased Cag-T4SS pilus formation. The broader implication and use of this technique will provide optimized culture conditions that result in elevated production of the Cag-T4SS pili. This assay will be useful to researchers seeking to determine the composition and architecture of the Cag-T4SS by enriching for this important bacterial surface feature. The sample preparation and visualization by field-emission gun electron microscopy has numerous advantages over alternative techniques such as light-microscopy methods to visualize the Cag-T4SS and will be appropriate to investigators interested in studying the regulation of this organelle10.
Protocol
1. H. pylori Growth in Various Conditions of Iron Availability and Co-culture with Human Gastric Epithelial Cells
Select H. pylori strain PMSS1 for these studies because it has an intact cag pathogenicity island and expresses a functioning type IV secretion system. Also utilize an isogenic cagE mutant (PMSSI ΔcagE) as a negative control. Grow bacteria on TSA plates supplemented with 5% sheep blood (blood agar plates) for 24 hr at 37 °C in the presence of 5% CO2. NOTE: Prepare reagents and assemble materials as outlined in the Materials and Equipment Table. Final concentrations for each reagent is listed in parentheses.
Scrape bacteria from the plate using a sterile cotton-tipped applicator and inoculate into modified brucella broth prepared from components (see Materials and Equipment Table) and supplemented with cholesterol. Incubate cultures overnight at 37 °C in room air supplemented with 5% CO2 with shaking at 200 rotations per minute (rpm). NOTE: Cholesterol is used in place of fetal bovine serum due to the fact that serum is complex; containing numerous sources of nutrient iron such as heme which causes variability in results.
On the day of bacterial culturing, seed AGS human gastric epithelial cells (ATCC CRL-1739) onto poly-D-lysine treated coverslips in 12-well plates (about 1 x 105 cells per well).
The following day, dilute bacterial cultures to an OD600 of 0.3 in modified brucella broth alone or supplemented with 100 µM FeCl3, 200 µM dipyridyl (a synthetic iron chelator), or 200 µM dipyridyl plus 250 µM FeCl3. Incubate these cultures for 4 hours at 37 °C in room air supplemented with 5% CO2 with shaking at 200 rpm.
Centrifuge bacteria at 1,000 x g to collect cells and remove supernatants before resuspending in an equal volume of fresh modified brucella broth supplemented with cholesterol. Measure OD600 of the culture (OD600 = 1.0 = 5.5 x 108 cells/ml) and add bacteria to the epithelial cells in a multiplicity of infection (MOI) of 20:1. Perform serial dilutions and plate bacterial cells onto blood agar plates to assess bacterial cell viability.
Incubate H. pylori and AGS human gastric epithelial cells co-cultures for 4 hr at 37 °C in the presence of 5% CO2 in static conditions before performing scanning electron microscopy (SEM) sample preparation.
2. SEM Sample Preparation to Visualize H. pylori Cag-T4SS Pili
Remove H. pylori and AGS co-cultures from the incubator and decant culture supernatant (save for IL-8 ELISA). Wash gently three times with 0.05 M sodium cacodylate buffer (pH 7.4). Fix samples for 2-4 hours at room temperature in a solution of 2.0% paraformaldehyde, 2.5% gluteraldehyde, and 0.05 M sodium cacodylate buffer. After primary fixation, wash samples three times with 0.05 M sodium cacodylate buffer.
Perform secondary fixation using 0.1% osmium tetroxide in 0.05 M sodium cacodylate buffer in two sequential 10 min fixation steps. After secondary fixation, wash samples three times with 0.05 M sodium cacodylate buffer.
After primary and secondary fixation, dehydrate samples by sequential washing with increasing concentrations of ethanol as per Table 1.
After ethanol dehydration, perform three sequential washes of liquid carbon dioxide. Then dry samples at a critical point using a critical point dryer machine at a temperature of 31.1 ºC and pressure of 1073 PSI.
Mount coverslips onto aluminum SEM sample stubs and paint with a thin line of colloidal silver at the sample edge to achieve appropriate grounding and avoid charging during SEM imaging.
Coat samples with 5 nm of gold-palladium using a sputter coat machine to increase sample secondary electron signal and edge effect.
3. Imaging Parameters for High Resolution SEM Analyses
View samples with a Field-Emission Gun Scanning Electron Microscope (FEG-SEM).
Image in a high vacuum mode at a working distance of 5-10 mm.
Set the accelerating voltage to 5 kV.
Set spot size to 2-2.5.
Tilt the sample between 15-25 degrees to achieve a better view of the host-pathogen interface.
Prioritize imaging bacteria adherent to the edges of the epithelial cells for high-magnification viewing, as these areas of host-pathogen interaction are enriched for pili-forming bacteria.
4. Statistical Analyses to Evaluate Pili Quantifications
Analyze H. pylori Cag-T4SS pili using ImageJ software to quantify number of pili/cell as well as percent of cells exhibiting the piliated phenotype per instructions below.
Open micrograph files in ImageJ software. Identify pili as structures formed between the bacterial cell and host cell, with uniform width (10-13 nm) and length (60-150 nm). NOTE: The pilus structure is present on WT bacterial strains, but absent on isogenic strains such as ΔcagE mutant strains, which harbor an inactivation in the gene encoding the major ATPase that powers type IV secretion pilus assembly.
Calibrate the measurement tool by drawing a line with the straight line tool and then clicking on the “Analyze” tab. Select “Set Scale” from the dropdown menu and enter the magnification bar value into the box labeled “Known Distance” and set the unit of length to the appropriate setting (nm). Click “OK”.
Measure Cag-T4SS pili using the straight line tool to draw over the length or width of the pilus and then press “Ctrl-M” to measure each pilus. Observe a dialogue box with the measured values. Record these values or copy and paste into a spreadsheet program.
Use the length and width parameters previously established24, disregard features that do not fit the profile of Cag-T4SS. Then quantify the number of pili based on values that are within the 10-13 nm width and 60-150 nm length cutoffs.
Analyze the quantification of pili statistically by two-tailed Student’s t test using GraphPad Prism software.
5. Optional: Evaluation of IL-8 Secretion to Correlate with Pilus Production
Using supernatants from Step 2.1, perform an IL-8 ELISA per manufacturer’s instructions with the kit (See Materials and Equipment Table).
Analyze the IL-8 secreted by host cells and calculate the percent IL-8 induction with respect to WT PMSS1 grown in medium alone. Perform statistical analysis using two-tailed Student’s t test using GraphPad Prism software.
Representative Results
In this report, we have demonstrated that conditions of varying iron availability have the capacity to modulate H. pylori Cag-T4SS pilus biogenesis at the host pathogen interface. When cultured in medium alone, H. pylori forms an average of 3 pili/cell. When H. pylori is grown in iron deplete conditions (using the synthetic chelator dipyridyl) that are sub-inhibitory to bacterial growth (Figure 1), the bacteria produce numerous Cag-T4SS pili (~7 pili/cell) when co-cultured with human gastric cells (Figure 2). Conversely, when an exogenous source of nutrient iron is present, the formation of Cag-T4SS pili is repressed (less than 1 pilus/cell in FE sample, less than 1 pilus/cell in FE+DIP sample) (Figure 2). Quantification of the pili at the host-pathogen interface reveals that in conditions of iron restriction, H. pylori exhibits an 2-fold increase in Cag-T4SS pili (p<0.0001) and the percent piliated cells increases by 11% (p<0.05) compared to cells grown in medium alone (Figure 3). Interestingly, the activity of the Cag-T4SS, as measured by host IL-8 secretion, is enhanced 107% under conditions of iron restriction and repressed 49% under conditions of excess iron availability compared to medium alone (p=0.03, and p<0.001, respectively) a result that supports the SEM data. However, pili dimensions remain consistent among all conditions of iron availability (Figure 4). Furthermore, iron availability does not change the biogenesis of pili on the surface of a cagE mutant, nor does it change this isogenic derivative’s ability to induce an IL-8 response from host cells, supporting the hypothesis that the structures demonstrated are, in fact, Cag-T4SS and not an additional unrelated surface feature.
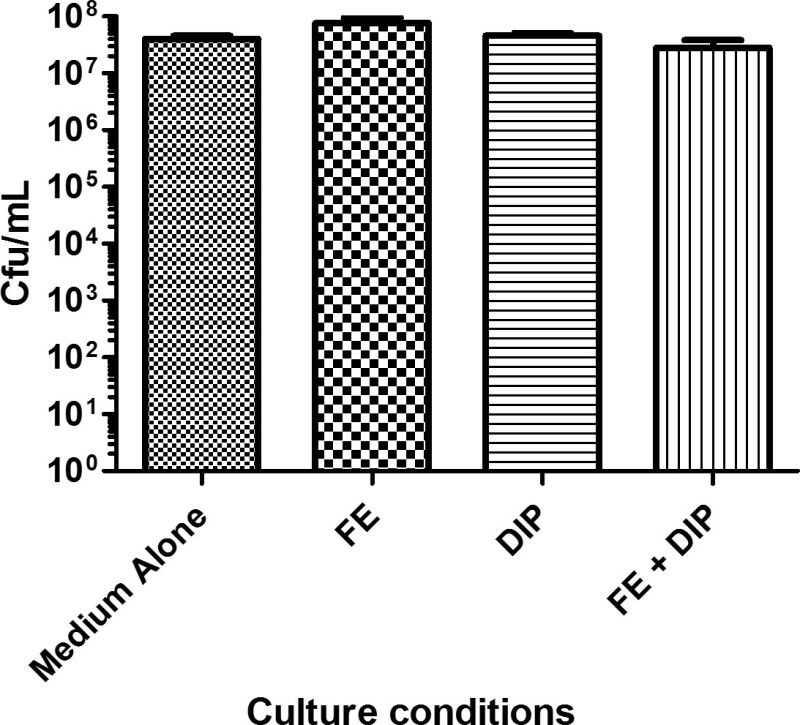 Figure 1: Bacterial viability determined in various conditions of iron availability. H. pylori was cultured in modified brucella broth alone (Medium Alone), or supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 µM FeCl3 (FE+DIP). Bacteria were diluted serially and plated onto bacteriological medium prior to incubation for 2 days at 37 °C in the presence of 5% CO2. Colonies were counted and colony forming units/mL (Cfu/ml) were calculated. Bars indicate mean of three independent experiments +/- SEM. Treatment with DIP, FE, or FE+DIP does not significantly alter bacterial viability.
Figure 1: Bacterial viability determined in various conditions of iron availability. H. pylori was cultured in modified brucella broth alone (Medium Alone), or supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 µM FeCl3 (FE+DIP). Bacteria were diluted serially and plated onto bacteriological medium prior to incubation for 2 days at 37 °C in the presence of 5% CO2. Colonies were counted and colony forming units/mL (Cfu/ml) were calculated. Bars indicate mean of three independent experiments +/- SEM. Treatment with DIP, FE, or FE+DIP does not significantly alter bacterial viability.
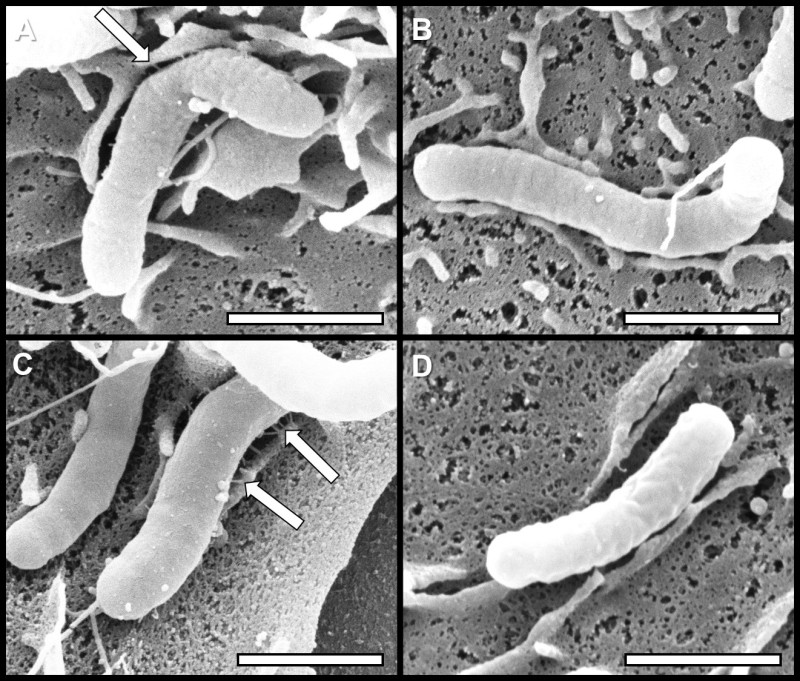 Figure 2: High resolution scanning electron microscopy analyses of H. pylori Cag-T4SS pili. Bacteria were grown in A) medium alone, B) medium supplemented with 100 µM FeCl3, C) medium supplemented with 200 µM dipyridyl, or D) medium supplemented with 200 µM dipyridyl plus 250 µM FeCl3 prior to co-culture with AGS human gastric cells. Arrows indicate Cag-T4SS pili formed at the host-pathogen interface. Samples were analyzed by high-resolution scanning electron microscopy to evaluate Cag-T4SS biogenesis. Conditions of low iron availability increase the prevalence of Cag-T4SS pilus formation, while conditions of excess available nutrient iron repress Cag-T4SS pilus formation.
Figure 2: High resolution scanning electron microscopy analyses of H. pylori Cag-T4SS pili. Bacteria were grown in A) medium alone, B) medium supplemented with 100 µM FeCl3, C) medium supplemented with 200 µM dipyridyl, or D) medium supplemented with 200 µM dipyridyl plus 250 µM FeCl3 prior to co-culture with AGS human gastric cells. Arrows indicate Cag-T4SS pili formed at the host-pathogen interface. Samples were analyzed by high-resolution scanning electron microscopy to evaluate Cag-T4SS biogenesis. Conditions of low iron availability increase the prevalence of Cag-T4SS pilus formation, while conditions of excess available nutrient iron repress Cag-T4SS pilus formation.
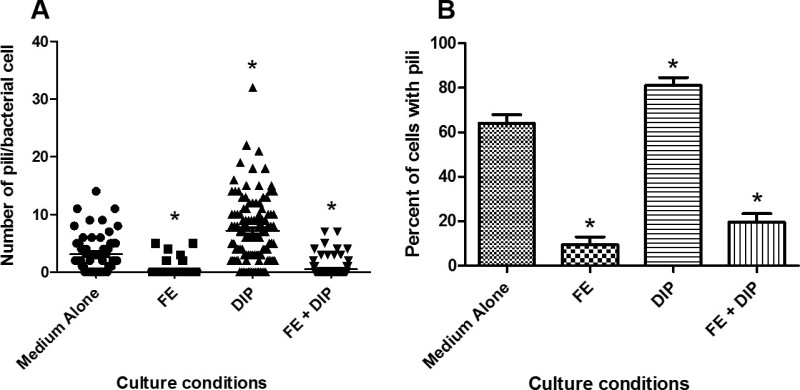 Figure 3: Quantification of pili/cell and percent piliated cells cultured in various conditions of iron availability. Bacteria were grown in modified brucella broth alone (Medium Alone), or medium supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 µM FeCl3 (FE+DIP) prior to co-culture with human gastric epithelial cells. Samples were analyzed by high-resolution scanning electron microscopy and pili were enumerated using ImageJ software. A) Scatterplot of pili per cell (*p<0.0001 compared to medium alone) and B) Bar graph depicting percent of piliated cells (*p<0.05 compared to medium alone) derived from 60-112 cells per culture condition derived from 3 separate biological experiments.
Figure 3: Quantification of pili/cell and percent piliated cells cultured in various conditions of iron availability. Bacteria were grown in modified brucella broth alone (Medium Alone), or medium supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 µM FeCl3 (FE+DIP) prior to co-culture with human gastric epithelial cells. Samples were analyzed by high-resolution scanning electron microscopy and pili were enumerated using ImageJ software. A) Scatterplot of pili per cell (*p<0.0001 compared to medium alone) and B) Bar graph depicting percent of piliated cells (*p<0.05 compared to medium alone) derived from 60-112 cells per culture condition derived from 3 separate biological experiments.
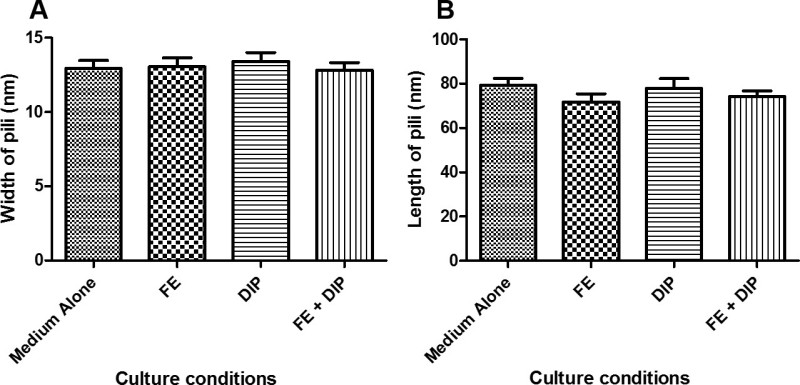 Figure 4: Pilus dimensions measured from cells cultured in various conditions of iron availability. Bacteria were grown in modified brucella broth alone (Medium Alone), or medium supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 µM FeCl3 (FE+DIP) prior to co-culture with human gastric epithelial cells. Samples were analyzed by high-resolution scanning electron microscopy and pili dimensions were measured using ImageJ software. A) Pilus width is an average of 13.1+/- 2.4 nm among all conditions. B) Pilus length is an average of 75.8 +/- 16.0 nm among all conditions. No significant difference in pilus dimensions are seen in the various conditions of iron availability.
Figure 4: Pilus dimensions measured from cells cultured in various conditions of iron availability. Bacteria were grown in modified brucella broth alone (Medium Alone), or medium supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 µM FeCl3 (FE+DIP) prior to co-culture with human gastric epithelial cells. Samples were analyzed by high-resolution scanning electron microscopy and pili dimensions were measured using ImageJ software. A) Pilus width is an average of 13.1+/- 2.4 nm among all conditions. B) Pilus length is an average of 75.8 +/- 16.0 nm among all conditions. No significant difference in pilus dimensions are seen in the various conditions of iron availability.
Supplemental Figure 1: High resolution scanning electron microscopy analyses of H. pylori cagE mutant in various conditions of iron availability. Bacteria were grown in A) medium supplemented with 100 µM FeCl3, B) medium supplemented with 200 µM dipyridyl (DIP) or C) medium supplemented with 200 µM dipyridyl (DIP) plus 250 µM FeCl3 prior to co-culture with AGS human gastric cells. Samples were analyzed by high-resolution scanning electron microscopy to evaluate Cag-T4SS biogenesis. Conditions of low or high iron do not modulate any Cag-independent pilus biogenesis, supporting the hypothesis that the structures visualized in Figure 2 are Cag-T4SS pili.
Supplemental Figure 2: ELISA analysis of host IL-8 secretion in response to bacteria grown in varying conditions of iron availability. Wild-type (white bars) or cagE mutant (grey bars) bacteria were grown in modified brucella broth alone (Medium Alone), or medium supplemented with 100 µM FeCl3 (FE), 200 µM dipyridyl (DIP), or 200 µM dipyridyl plus 250 uM FeCl3 (FE+DIP) prior to co-culture with human gastric epithelial cells. Iron restriction results in increased IL-8, a host response that is dependent upon Cag-T4SS activity. Bars indicate mean +/- SEM, derived from 3 separate biological experiments. (*p<0.05 compared to PMSS1 grown in medium alone).
Discussion
Iron is an essential micronutrient for most forms of life, including bacterial pathogens. In an effort to restrict the viability of invading microorganisms, vertebrate hosts sequester nutrient iron in a process known as “nutritional immunity”18. In response to this, bacterial pathogens have evolved to use iron as a global signaling molecule to sense their surroundings and regulate the elaboration of virulence features such as iron acquisition systems, toxins, and toxin secretion machinery19,20.
H. pylori requires micronutrients such as iron, zinc, and magnesium to grow22 and can use alternate iron sources such as heme, hemoglobin, transferrin, and lactoferrin23. H. pylori, like most successful pathogens, has evolved a wide repertoire of virulence factors to help it scavenge nutrient iron from the host21. One virulence factor, CagA, the effector molecule of the Cag-T4SS, is implicated in rerouting transferrin from the basolateral side of polarized cells to the apical surface where it can be readily used by the bacterial cells as a nutrient iron source and promote replication15. Thus, it seems an intuitive evolutionary strategy to promote the secretion of CagA in conditions of low iron availability.
The H. pylori Cag-T4SS is an important bacterial organelle, due to the proinflammatory host cell responses it elicits. The effects of the Cag-T4SS have been well studied in both co-cultures and animal models of infection. However, studies of the macromolecular structure and composition of this toxin secretion system have been hindered by the lack of high-resolution imaging protocols. Our previously published reports studying the Cag-T4SS have reported an average of four Cag-T4SS pili per bacterial cell by high-resolution electron microscopy analyses24. Our improved method provides a robust imaging technique to visualize the Cag-T4SS by using a culture technique that promotes the production of this feature on the surface of the bacterial cell. Utilization of culture techniques in conjunction with high-resolution field-emission gun scanning electron microscope is critical for this imaging protocol. Two-fold enrichment of the Cag-T4SS surface feature is a significant increase for investigators interested in performing immunohistochemical analyses such as immunofluorescence or immunoprecipitation techniques to further characterize the subcellular location or composition of the Cag-T4SS. Once the high-resolution electron microscopy imaging of the Cag-T4SS has been mastered, it is also possible to develop other applications, such as immunogold-labeling, to determine the location of bacterial proteins with respect to the Cag-T4SS pilus25.
One limitation of our technique is the fact that the investigator is still restricted to using a co-culture system for visualization of the Cag-T4SS. In a single field, only a subset of H. pylori under normal culture conditions form these structures (about 50-70 percent, see Figure 3 panel B). So numerous cells in a field will be non-piliated. It is unknown why these subpopulations exist in the same location, but these cells are always accounted within both the quantifications of pili/cell and percent of cells with pili on the surface. Because the Cag-T4SS is only observed at the host-bacterial interface, the primary and secondary fixation steps within the SEM sample preparation are critical for the preservation of both proteinaceous structures and the membranes of the eukaryotic and prokaryotic cells. However, over-fixation can occur if fixation steps are carried out longer than outlined, or if the solutions are not freshly prepared. If over-fixation occurs, samples will have a stony appearance when viewed under SEM and the Cag-T4SS will be masked by this. To troubleshoot, repeat the SEM sample preparation using less concentrated fixatives and/or less time incubated in the presence of the fixative.
The co-culture system also renders biochemical characterization (such as mass spectrometry analyses) cumbersome due to the contamination of large quantities of host proteins. However, understanding the molecular mechanisms that regulate the production of this bacterial secretion system could ultimately lead to new techniques that will trigger Cag-T4SS pilus formation in the absence of host cells. This would greatly improve the isolation and purification techniques for biochemical assays.
In conclusion, we report that conditions of restricted iron availability result in increased production of H. pylori Cag-T4SS that can be visualized by high resolution scanning electron microscopy techniques. We also report the iron-dependent repression of this surface feature at the host-pathogen interface. This assay may be optimized and utilized for numerous bacterial pathogens that form iron-regulated toxin-secretions and is broadly applicable to a variety of host-microbe interactions.
Disclosures
The authors declare that they have no competing financial interests.
Acknowledgments
This research was supported by the Department of Veterans Affairs Career Development Award 1IK2BX001701 and the CTSA award UL1TR000445 from the National Center for Advancing Translational Sciences. Its contents are solely the responsibility of the authors and do not necessarily represent official views of the National Center for Advancing Translational Sciences or the National Institutes of Health. Scanning electron microscopy experiments were performed in part through the use of the VUMC Cell Imaging Shared Resource, supported by NIH grants CA68485, DK20593, DK58404, DK59637 and EY08126.
References
- Cover TL, Blaser MJ. Helicobacter pylori in health and disease. Gastroenterology. 2009. pp. 1863–1873. [DOI] [PMC free article] [PubMed]
- Sycuro LK, et al. Peptidoglycan crosslinking relaxation promotes Helicobacter pylori's helical shape and stomach colonization. Cell. 2010;141(5):822–833. doi: 10.1016/j.cell.2010.03.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kodaman N, et al. Human and Helicobacter pylori coevolution shapes the risk of gastric disease. Proc. Natl. Acad. Sci. U.S.A. 2014;111(4):1455–1460. doi: 10.1073/pnas.1318093111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tegtmeyer N, Wessler S, Backert S. Role of the cag-pathogenicity island encoded type IV secretion system in Helicobacter pylori pathogenesis. FEBS J. 2011;278(8):1190–1202. doi: 10.1111/j.1742-4658.2011.08035.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer W. Assembly and molecular mode of action of the Helicobacter pylori Cag type IV secretion apparatus. FEBS J. 2011;278(8):1203–1212. doi: 10.1111/j.1742-4658.2011.08036.x. [DOI] [PubMed] [Google Scholar]
- Odenbreit S, Puls J, Sedlmaier B, Gerland E, Fischer W, Haas R. Translocation of Helicobacter pylori CagA into gastric epithelial cells by type IV secretion. Science. 2000;287(5457):1497–1500. doi: 10.1126/science.287.5457.1497. [DOI] [PubMed] [Google Scholar]
- Bourzac K, Guillemin K. Helicobacter pylori-host cell interactions mediated by type IV secretion. Cell Microbiol. 2005;7(7):911–919. doi: 10.1111/j.1462-5822.2005.00541.x. [DOI] [PubMed] [Google Scholar]
- Bronte-Tinkew DM, et al. Helicobacter pylori cytoxin-associated gene A activates the signal transducer and activator of transcription 3 pathway in vitro and in vivo. Cancer Res. 2009;69(2):632–639. doi: 10.1158/0008-5472.CAN-08-1191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson EM, Gaddy JA, Cover TL. Alterations in Helicobacter pylori triggered by contact with gastric epithelial cells. Front. Cell. Infect. Microbiol. 2012. p. 17. [DOI] [PMC free article] [PubMed]
- Rohde M, Puls J, Buhrdorf R, Fischer W, Haas R. A novel sheathed surface organelle of the Helicobacter pylori cag type IV secretion system. Mol. Microbiol. 2003;49(1):219–234. doi: 10.1046/j.1365-2958.2003.03549.x. [DOI] [PubMed] [Google Scholar]
- Cover TL, Peek RM., Jr Diet, microbial virulence and Helicobacter pylori induced gastric cancer. Gut Microbes. 2013;4(6):482–493. doi: 10.4161/gmic.26262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Queiroz DM, et al. Increased gastric IL-1β concentration and iron deficiency parameters in Helicobacter pylori infected children. PLoS One. 2013;8(2):57420. doi: 10.1371/journal.pone.0057420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harris PR, et al. Helicobacter pylori-associated hypochlorhydria and development of iron deficiency. J. Clin. Pathol. 2013;66(4):343–347. doi: 10.1136/jclinpath-2012-201243. [DOI] [PubMed] [Google Scholar]
- Noto JM, et al. Iron deficiency accelerates Helicobacter pylori-induced carcinogenesis in rodents and humans. J. Clin. Invest. 2013;123(1):479–492. doi: 10.1172/JCI64373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan S, Noto JM, Romero-Gallo J, Peek RM, Jr, Amieva MR. Helicobacter pylori perturbs iron trafficking in the epithelium to grow on the cell surface. PLoS Pathog. 2011;7(5) doi: 10.1371/journal.ppat.1002050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Danielli A, Roncarati D, Delany I, Chiarini V, Rappouli R, Scarlato V. In vivo dissection of the Helicobacter pylori Fur regulatory circuit by genome-wide location analysis. J. Bacteriol. 2006;188(13):4654–4662. doi: 10.1128/JB.00120-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pich OQ, Carpenter BM, Gilbreath JJ, Merrell DS. Detailed analysis of Helicobacter pylori Fur-regulated promoters reveals a Fur box core sequence and novel Fur-regulated genes. Mol. Microbiol. 2012;84(5):921–941. doi: 10.1111/j.1365-2958.2012.08066.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cassat JE, Skaar EP. Iron in infection and immunity. Cell Host Microbe. 2013;13(5):509–519. doi: 10.1016/j.chom.2013.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielubowicz GR, Mobley HL. Host-pathogen interactions in urinary tract infection. Nat. Rev. Urol. 2010;7(8):430–441. doi: 10.1038/nrurol.2010.101. [DOI] [PubMed] [Google Scholar]
- Brickman TJ, Cummings CA, Liew SY, Relman DA, Armstrong SK. Transcriptional profiling of the iron starvation response in Bordatella pertussis provides new insights into siderophore utilization and virulence gene expression. J. Bacteriol. 2011;193(18):4798–4812. doi: 10.1128/JB.05136-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mobley HL. Helicobacter pylori factors associated with disease development. Gastroenterology. 1997;113(6):21–28. doi: 10.1016/s0016-5085(97)80006-6. [DOI] [PubMed] [Google Scholar]
- Testerman TL, Conn PB, Mobley HL, McGee DL. Nutritional requirements and antibiotic resistance patterns of Helicobacter species in chemically defined. J. Clin. Microbiol. 2006;44(5):1650–1658. doi: 10.1128/JCM.44.5.1650-1658.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Senkovich O, Ceaser S, McGee DJ, Testerman TL. Unique host iron utilization mechanisms of Helicobacter pylori revealed with iron-deficient chemically defined media. Infect. Immun. 2010;78(5):1841–1849. doi: 10.1128/IAI.01258-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaffer CL, et al. Helicobacter pylori exploits a unique repertoire of type IV secretion system components for pilus assembly at the bacteria-host cell interface. PLoS Pathog. 2011;7(9) doi: 10.1371/journal.ppat.1002237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrozo RM, et al. Functional plasticity in the type IV secretion system of Helicobacter pylori. PLoS Pathog. 2013;9(2) doi: 10.1371/journal.ppat.1003189. [DOI] [PMC free article] [PubMed] [Google Scholar]