

Abstract
B lymphocyte immunoglobulin heavy chain (IgH) class switch recombination (CSR) is a process wherein initially expressed IgM switches to other IgH isotypes, such as IgA, IgE and IgG. Measurement of IgH CSR in vitro is a key method for the study of a number of biologic processes ranging from DNA recombination and repair to aspects of molecular and cellular immunology. In vitro CSR assay involves the flow cytometric measurement surface Ig expression on activated B cells. While measurement of IgA and IgG subclasses is straightforward, measurement of IgE by this method is problematic due to soluble IgE binding to FcεRII/CD23 expressed on the surface of activated B cells. Here we describe a unique procedure for accurate measurement of IgE-producing mouse B cells that have undergone CSR in culture. The method is based on trypsin-mediated cleavage of IgE-CD23 complexes on cell surfaces, allowing for detection of IgE-producing B lineage cells by cytoplasmic staining. This procedure offers a convenient solution for flow cytometric analysis of CSR to IgE.
Keywords: Immunology, Issue 94, Class switch recombination, AID, B cell activation, IgE, IgG1, CD23/FcεRII, flow cytometry, trypsin, cytosolic staining
Introduction
During immunoglobulin heavy chain (IgH) class switch recombination (CSR) in mice and humans, the IgH μ constant region exons (Cμ) are deleted and replaced by one of several sets of downstream constant region exons (CHs) (e.g. Cγ, Cε, and Cα), resulting in a switch from the production of IgM to the production of other Ig classes (e.g. IgG, IgE, or IgA). CSR occurs within switch (S) regions, which are 1-10 kb sequences located 5’ to each set of CHs1. The Activation-Induced Cytidine Deaminase (AID) enzyme initiates CSR via cytidine deamination activity.
IgE is a key mediator of allergic disease2 and a greater understanding of how IgE is produced and regulated may open doors to new therapeutic approaches to atopic disease. In mice and humans, IgE is the most tightly regulated Ig isotype. IgE is normally detected at levels thousands of times less than other IgH isotypes3, but can be highly elevated in disease states4. However, CSR to IgE is incompletely understood. In vitro activation of B cells using IL-4 in combination with either anti-CD40 or LPS, induces CSR to both IgG1 and IgE5. Activated B cells express the low affinity IgE receptor FcεRII/CD232,6, which binds soluble IgE secreted in culture after CSR. Therefore when analyzing with flow cytometry, B cells with receptor-bound IgE stain similarly to B cells endogenously expressing IgE7. While it is known that mouse B cells express the low affinity IgG receptor FcγRIIB1 (CD32)8, in our experience it does not appear to interfere with measurement of class switching to IgG1. However, when measuring IgE switching after B cell activation in culture, nonspecific surface-bound IgE may obscure the analysis. With common staining methods, non-IgE-expressing cells stain positively for IgE.
Outlined here is a strategy that has been utilized to conduct CSR assays and detect true IgE-expressing B cells from mice9–11. Treatment of activated B cells with trypsin, a common lab reagent to digest protein, removes both cytophylic and membrane bound surface IgE. Subsequent permeabilization and staining for cytoplasmic IgE thus reveals the true IgE-producing cells.
Protocol
NOTE: All experiments described here were in accordance with The Institutional Animal Care and Use Committee (IACUC) guidelines and approved by Animal Research Children’s Hospital (ARCH), Boston, Mass.
1. Preparation of Reagents
1,000x IL-4 Stock: Dilute IL-4 cytokine to 20 µg/ml in H2O or 0.1% bovine serum albumin (BSA). Dispense in 200 µl aliquots to 1.5 ml microcentrifuge tubes and store at -20 °C.
1,000x anti-CD40: Obtain from the manufacturer at 1.0 mg/ml, store at 4 °C.
B cell stimulation media: To 425 ml of RPMI-1640 medium, add 75 ml of freshly thawed fetal calf serum (FCS), 10 ml of 1.0 M HEPES buffer, 5 ml L-glutamine stock, 5 ml Penicillin:Streptomycin stock, 5 ml Minimum Essential Medium Non-Essential Amino Acids (MEM-NEAA), and 3.5 µl of 2-mercaptoethanol. Keep media at 4 ˚C for up to a week. Add IL-4 (20 ng/ml) and anti-CD40 (1.0 µg/ml) immediately before use only to the volume of media required.
2% FCS-FACS Buffer: Add 10 ml of FCS to 490 ml of 1x PBS. Keep at 4 °C.
2x trypsin-EDTA solution: Dilute 5 ml of 10x trypsin-EDTA stock solution (5.0 g trypsin, 2.0 g EDTA per liter) in 20 ml of 1x PBS. Store trypsin-EDTA in 2x working aliquots at 4 °C. Allow to warm to room temperature before use.
Fetal calf serum: Keep thawed FCS at 4 ˚C for up to two weeks. Place on ice 15 min before the cytoplasmic staining procedure.
Neutral-buffered formalin: Prepare a 10% formalin solution (3.7% paraformaldehyde). Formalin is toxic and therefore handle it under a laboratory fume hood while wearing appropriate PPE. Store formalin at room temperature in a chemical storage cabinet.
Methanol: Store absolute methanol (100%) at -20 °C until the cells are ready to be permeabilized. Note that methanol is flammable and should be stored in an appropriate freezer.
2. Purification and Activation of B Cells
From a freshly-harvested mouse spleen, prepare a single cell suspension by gently pressing the organ through a 70 µm cell strainer with the plunger of a 3 ml syringe. In a 15 ml conical centrifuge tube, collect cells in 10 ml of 1x PBS and centrifuge at 300 x g for 5 min at 4 °C.
Decant the supernatant and resuspend pellet in 1 ml of Red Blood Cell Lysing Buffer for 5 min. Neutralize with 13.5 ml of 1x PBS and centrifuge at 300 x g for 5 min.
(Optional) Magnetically separate the B cells using anti-CD45R(B220)-labeled beads, following the MACS purification protocol according to manufacturer’s instructions.
Stimulate purified B cells at 1.0 x 106 cells/ml in warmed B cell stimulation media containing IL-4 (20 ng/ml) and anti-CD40 (1.0 µg/ml) for 5 days at 37 °C. Start with 8 ml of culture divided into 2 wells of a 6-well culture plate (4 ml each).
Check culture daily to maintain at 1.0 x 106 cells/ml. Adjust the concentration by splitting into other wells and diluting the culture with B cell stimulation media containing fresh IL-4 and anti-CD40.
3. Trypsinization, Fixation, and Permeabilization
- Trypsinization
- Collect up to 1.0 x 107 of cultured cells in a 15 ml conical tube, centrifuge at 300 x g for 5 min, and decant the supernatant.
- Wash with 10 ml of PBS to remove the residual protein from the cell collection media. Repeat the centrifugation step and decant the supernatant.
- Resuspend the cell pellet in the residual volume after decanting (about 100 μl).
- Carefully add 800 μl of room temperature 0.1% (2x) trypsin-EDTA to the cells. Dispense slowly at a 45° angle down the side of the conical tube.
- Close the tube and mix gently by tilting the tube so the liquid runs along the length of the tube 5-6 times. A stringy, white precipitate may form in the solution. Limit the trypsinization time to under a min.
- Quench the trypsinization reaction with 1 ml of cold FCS. Dilute the FCS immediately by adding 10 ml of cold PBS.
- Centrifuge at 300 x g for 5 min at 4 °C, then decant the supernatant and return the tubes to ice.
- Fixation
- Resuspend the cells in the residual volume after decanting. If necessary, bring volume up to 100 μl with cold PBS.
- In a fume hood, add 800 μl of 10% neutral buffered formalin solution to the cells and pipet up and down. The tube will warm upon the addition of formalin. Incubate at 37 °C for 10 min.
- Permeabilization
- Set up a benchtop vortex mixer at a gentle shaking speed.
- Draw 9 ml of cold (-20 °C) methanol into a 10 ml serological pipet.
- Hold the tube containing the cells over the vortex mixer and gently start shaking the cells.
- Add cold methanol to the tube so it falls dropwise onto the cells while the tube is still shaking so that the methanol is continuously mixing. After the first 2 ml of methanol the dispensing speed can be increased to a slow stream.
- Let the tube sit on ice for 30 min to complete the permeabilization. Alternatively, the cells can be stored at -20 °C for up to a month.
4. Fluorescence Staining for IgG1 and IgE Class Switching
Centrifuge sample at 300 x g for 5 min and decant the methanol solution. A white precipitate may pellet with the cells.
Add 10 ml of room temperature 1x PBS and centrifuge at 300 x g for 5 min to wash cells. Repeat once. Note: Washing the pellet with room temperature PBS dissolves extra precipitate that may form after step 4.1.
Perform an additional wash with 10 ml of 2% FCS-FACS buffer.
Distribute samples to a 96-well round bottom plate and centrifuge at 300 x g for 5 min. Decant supernatant and put the plate on ice.
Stain in 60 µl of 2% FCS-FACS buffer per well with the following antibody dilutions: Anti-IgM RPE/Cy7, 1:200, Anti-IgG1 PE, 1:600, Anti-IgE FITC, 1:100, Anti-CD45R(B220) PerCP-Cy5.5, 1:200
After 5 min of staining, wash the plate with 150 µl of 2% FCS-FACS buffer and centrifuge for 5 min at 300 x g.
Resuspend in 400 µl of 2% FCS-FACS buffer and pipet through a 40 µm nylon cell strainer into a 5 ml FACS tube.
5. Use the Following Gating Strategy:
Isolate CD45R(B220) positive cells from the blasting lymphocyte population located on the FSC/SSC plot (Figure 1).
From the B220 positive cells, use the IgE and IgG1 plot to reveal the corresponding populations.
Representative Results
This procedure has been successfully implemented to study CSR to IgE in mouse B cells. To demonstrate the efficiency in measuring CSR, we stimulated mouse splenic B cells with anti-CD40 and IL-4 as described previously11. After five days of stimulation, cells were collected and processed using the protocol described above and stained with fluorescently-labeled IgM, IgG1, IgE, and B220 (CD45R) antibodies. Gated on the blasting lymphocytes (Figure 1), a clear population of IgE+ cells cannot be cleanly discriminated without trypsin treatment (Figure 2a). However, trypsinization of cells prior to fixation and permeabilization enables separation of the IgE-producing cells (Figure 2b) due to the removal of surface-bound IgE.
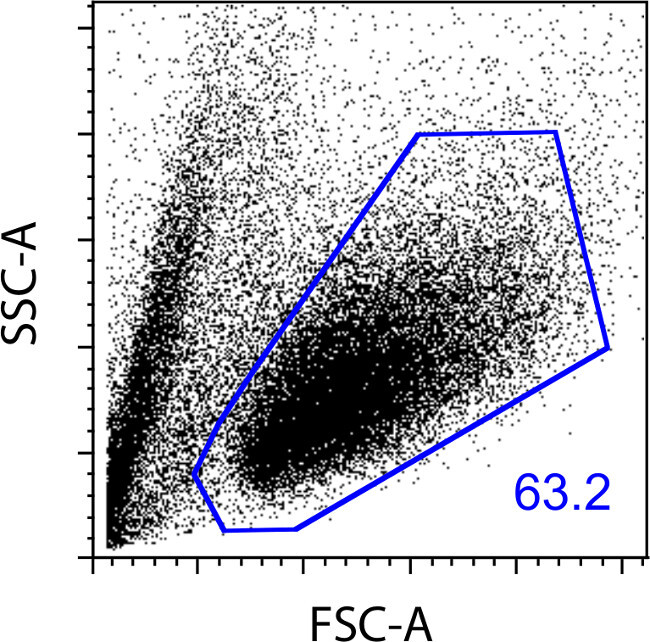 Figure 1:Forward vs. side scatter FACS plot of mouse splenic B cells after 5 days of stimulation in culture with IL-4 and anti-CD40. Blasting lymphocytes make up 63% of the total acquired events.
Figure 1:Forward vs. side scatter FACS plot of mouse splenic B cells after 5 days of stimulation in culture with IL-4 and anti-CD40. Blasting lymphocytes make up 63% of the total acquired events.
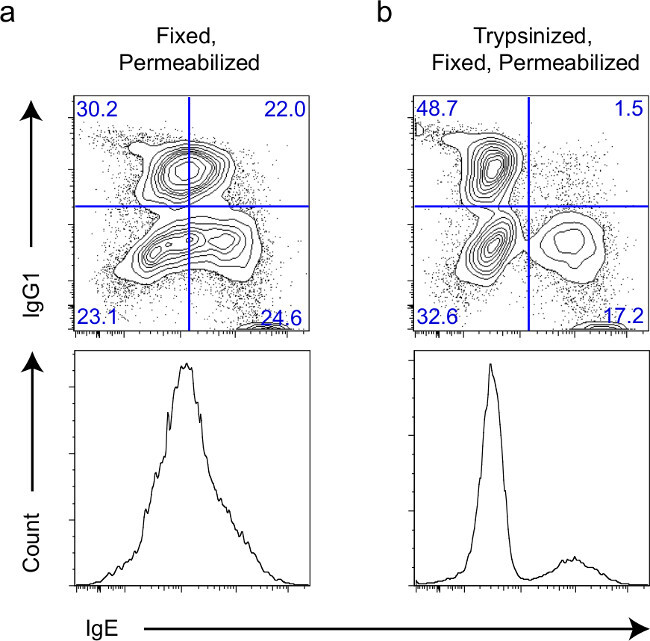 Figure 2:Effect of trypsin on cytoplasmic staining of IgE and IgG1 in activated B cells.(a, b) FACS plots (above) and histograms (below) gated on blasting splenic B cells from 129/B6 mixed background mice after activation in culture for 5 days with anti-CD40 and IL-4. Fixation and methanol permeabilization alone (a) and with addition of trypsin (b) are shown.
Figure 2:Effect of trypsin on cytoplasmic staining of IgE and IgG1 in activated B cells.(a, b) FACS plots (above) and histograms (below) gated on blasting splenic B cells from 129/B6 mixed background mice after activation in culture for 5 days with anti-CD40 and IL-4. Fixation and methanol permeabilization alone (a) and with addition of trypsin (b) are shown.
Discussion
Stimulation of mouse splenic B cells in culture with anti-CD40 and IL-4 will simulate T helper type 2 (TH2) interactions, encouraging class switching to IgG1 and IgE5. B cells can be activated for CSR in the context of total splenocytes12 or as purified splenic B cells11. As noted in the protocol (step 2.3), B cell enrichment is optional, and is at the discretion of the experimenter to determine if it would beneficial. Positive magnetic separation of B cells using CD45R(B220)-labeled beads is utilized here. After separation, it is recommended to check the cell purity by FACS so that the B cell concentration can be adjusted to 1.0 x 106 cells/ml for proper cytokine and antibody stimulation. Also it should be noted that IL-4 protein that has undergone freeze/thaw cycles might have decreased activity.
Trypsinization of cells prior to fixation and permeabilization allows for the accurate measurement of class switching via flow cytometry (Figure 2). Proper trypsinization of the cells is vital to the successful application of this procedure. As FCS can inhibit trypsin, a 1x PBS wash is performed prior to the trypsinization step. We found a 30 sec incubation at room temperature is sufficient for complete digestion. Longer durations can noticeably affect cell viability.
Prior to analysis, it is essential to wash the cells multiple times with PBS to remove traces methanol. Methanol carryover may alter antibody binding13 and can precipitate proteins from FCS used in downstream flow cytometric analysis14. In this regard, other permeabilization methods (i.e. saponin) may be an alternative to using methanol. Methanol is an organic solvent that dissolves membrane lipids, while detergents disrupt membrane integrity15. A benefit to methanol permeabilization is the ability of extended storage of fixed cells at -20 °C14.
There are some limitations to keep in mind when using this procedure. Because trypsin cleaves protein residues, some FACS stains may appear different after treatment, or may lose the ability to bind to target epitopes altogether. Additionally, a loss in cell number after trypsin treatment can occur. However, this does not appear to affect the accuracy of IgE CSR measurements as we have found this method to be clearly comparable to class switching data collected from measuring the antibody secretions of individual hybridoma clones11.
Other methods have been reported to detect IgE-producing mouse B cells. One method uses an acid wash procedure to remove receptor-bound IgE prior to staining for membrane-bound IgE on IgE-expressing cells16–18. Another method uses a monoclonal anti-IgE antibody to block all surface IgE before cell permeabilization and staining for intracellular IgE of the IgE-expressing cells19. Furthermore, a recent study profiled an eight-color FACS panel to identify IgE-switched B cell populations in humans by excluding IgM+, IgD+, IgG+ and IgA+ B cell subsets20. Compared to these other methods, the method described here avoids the use of denaturing acid washes, and excess antibodies are not necessary.
A benefit to this procedure is that it does not prescribe specific monoclonal antibodies for measurement of CSR to IgE. Alternative anti-IgE monoclonal antibodies have been shown to detect only endogenous IgE molecules. The hybridoma clone R1E4 produces a rat-anti-mouse monoclonal IgE antibody specific only for endogenous surface molecules, not cytophilic IgE bound to CD2321,22, but is not commercially available. The protocol described above is convenient and economical because it uses common lab reagents. The anti-IgE antibody used here likely binds to receptor-bound IgE at a site sensitive to trypsin-mediated cleavage, because the IgE signal due to cytophilic IgE is reduced after treatment (Figure 2). However researchers executing this protocol have the freedom to select from a range of commercially available clones for experiments. Furthermore, the concept of using trypsin to remove cell surface proteins can be extended to the detection of other cell markers by FACS analysis.
Disclosures
The authors have no disclosures.
Acknowledgments
D.R.W. is supported by NIH grants AI089972 and AI113217, and by the Mucosal Immunology Studies Team, and holds a Career Award for Medical Scientists from the Burroughs Wellcome Fund.
References
- Chaudhuri J, et al. Evolution of the immunoglobulin heavy chain class switch recombination mechanism. Advances in immunology. 2007;94:157–214. doi: 10.1016/S0065-2776(06)94006-1. [DOI] [PubMed] [Google Scholar]
- Gould HJ, Sutton BJ. IgE in allergy and asthma today. Nature reviews. Immunology. 2008;8:205–217. doi: 10.1038/nri2273. [DOI] [PubMed] [Google Scholar]
- Winter WE, Hardt NS, Fuhrman S. Immunoglobulin E. Archives of Pathology & Laboratory Medicine. 2000;124(9):1382–1385. doi: 10.5858/2000-124-1382-IE. [DOI] [PubMed] [Google Scholar]
- Ozcan E, Notarangelo LD, Geha RS. Primary immune deficiencies with aberrant IgE production. Journal of Allergy and Clinical Immunology. 2008;122:1054–1062. doi: 10.1016/j.jaci.2008.10.023. [DOI] [PubMed] [Google Scholar]
- Bacharier LB, Geha RS. Molecular mechanisms of IgE regulation. The Journal of allergy and clinical immunology. 2000;105:547–558. doi: 10.1016/s0091-6749(00)90059-9. [DOI] [PubMed] [Google Scholar]
- Yokota A, et al. Two species of human Fc epsilon receptor II (Fc epsilon RII/CD23): tissue-specific and IL-4-specific regulation of gene expression. Cell. 1988;55(4):611–618. doi: 10.1016/0092-8674(88)90219-x. [DOI] [PubMed] [Google Scholar]
- Boboila C, et al. Alternative end-joining catalyzes class switch recombination in the absence of both Ku70 and DNA ligase 4. The Journal of experimental medicine. 2010;207:417–427. doi: 10.1084/jem.20092449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nimmerjahn F, Ravetch J. Fcgamma receptors as regulators of immune responses. Nature reviews. Immunology. 2008;8(1):34–47. doi: 10.1038/nri2206. [DOI] [PubMed] [Google Scholar]
- Callen E, et al. 53BP1 mediates productive and mutagenic DNA repair through distinct phosphoprotein interactions. Cell. 2013;153(6):153–1280. doi: 10.1016/j.cell.2013.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wesemann DR, et al. Reprogramming IgH isotype-switched B cells to functional-grade induced pluripotent stem cells. Proceedings of the National Academy of Sciences of the United States of America. 2012;109(34):13745–13750. doi: 10.1073/pnas.1210286109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wesemann D, et al. Immature B cells preferentially switch to IgE with increased direct Sµ to Sε recombination. The Journal of experimental medicine. 2011;208(13):2733–2746. doi: 10.1084/jem.20111155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zarrin AA, et al. An evolutionarily conserved target motif for immunoglobulin class-switch recombination. Nature immunology. 2004;5:1275–1281. doi: 10.1038/ni1137. [DOI] [PubMed] [Google Scholar]
- Fan G, et al. Development of a class-specific polyclonal antibody-based indirect competitive ELISA for detecting fluoroquinolone residues in milk. Journal of Zhejiang University. Science. B. 2012;13(7):545–554. doi: 10.1631/jzus.B1200001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perez O, Krutzik P, Nolan G. Methods in molecular biology (Clifton, N.J.) Vol. 263. Clifton, NJ: 2004. Flow cytometric analysis of kinase signaling cascades; pp. 67–94. [DOI] [PubMed] [Google Scholar]
- Jamur M, Oliver C. Cell fixatives for immunostaining. Methods in molecular biology (Clifton, N.J.) 2010;588:55–61. doi: 10.1007/978-1-59745-324-0_8. [DOI] [PubMed] [Google Scholar]
- Ishizaka T, Ishizaka K. Mechanisms of passive sensitization. IV. Dissociation of IgE molecules from basophil receptors at acid pH. Journal of immunology (Baltimore, MD. : 1950) 1950;112(3):1078–1084. [PubMed] [Google Scholar]
- Erazo A, et al. Unique Maturation Program of the IgE Response In Vivo. Immunity. 2007;26:191–203. doi: 10.1016/j.immuni.2006.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiong H, Dolpady J, Wabl M, Curotto de Lafaille MA, Lafaille JJ. Sequential class switching is required for the generation of high affinity IgE antibodies. Journal of Experimental Medicine. 2012;209:353–364. doi: 10.1084/jem.20111941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Z, Sullivan B, Allen C. Fluorescent in vivo detection reveals that IgE(+) B cells are restrained by an intrinsic cell fate predisposition. Immunity. 2012;36(5):857–872. doi: 10.1016/j.immuni.2012.02.009. [DOI] [PubMed] [Google Scholar]
- Berkowska MA, et al. Human IgE(+) B cells are derived from T cell-dependent and T cell-independent pathways. The Journal of allergy and clinical immunology. 2014. [DOI] [PubMed]
- He J-S, et al. The distinctive germinal center phase of IgE+ B lymphocytes limits their contribution to the classical memory response. The Journal of experimental medicine. 2013;210(12):2755–2771. doi: 10.1084/jem.20131539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keegan A, Fratazzi C, Shopes B, Baird B, Conrad D. Characterization of new rat anti-mouse IgE monoclonals and their use along with chimeric IgE to further define the site that interacts with Fc epsilon RII and Fc epsilon RI. Molecular immunology. 1991;28(10):1149–1154. doi: 10.1016/0161-5890(91)90030-n. [DOI] [PubMed] [Google Scholar]