

Abstract
Fluorescent nanoparticles (NPs) with desirable chemical, optical and mechanical properties are promising tools to label intracellular organelles. Here, we introduce a method using gold-BSA-rhodamine NPs to label the endo-lysosomal system of eukaryotic cells and monitor manipulations of host cellular pathways by the intracellular pathogen Salmonella enterica. The NPs were readily internalized by HeLa cells and localized in late endosomes/lysosomes. Salmonella infection induced rearrangement of the vesicles and accumulation of NPs in Salmonella-induced membrane structures. We deployed the Imaris software package for quantitative analyses of confocal microscopy images. The number of objects and their size distribution in non-infected cells were distinct from the ones in Salmonella-infected cells, indicating extremely remodeling of the endo-lysosomal system by WT Salmonella.
Keywords: Immunology, Issue 95, fluorescent nanoparticles, endo-lysosomal system, labeling, intracellular bacteria, quantitative image analysis, tubular compartments
Introduction
Fluorescent nanoparticles (NPs), including metal NPs, quantum dots, polymer NPs, silica NPs, carbon dots, etc., have attracted considerable attention during the past decades1,2. Compared to traditional organic dyes, fluorescent NPs show desirable chemical, optical and mechanical properties, such as strong signal strength, resistance to photobleaching and high biocompatibility3,4. These advantages make them the method of choice for intracellular sensing and live cell imaging. Furthermore, a variety of electron-dense NPs are visible by electron microscopy (EM), facilitating their use for correlated microscopic analysis, which allows combination of live cell tracking with light microscopy (LM) and higher resolution at ultrastructural level with EM5. For example, gold NPs have been long time efficiently used as biosensors in living cells for sensitive diagnosis as well as in the field of immuno-labeling6. Recent studies indicate that gold NPs with different size and shape can be readily uptake by a large variety of cell lines and routinely transport through the endosomal pathway, therefore have great potential being applied for intracellular vesicle transportation tracking and the endo-lysosomal system labeling7,8.
Microbial pathogens, such as Salmonella enterica, Shigella flexneri and Listeria monocytogenes, have developed different mechanisms to invade non-phagocytic host cells9. After being internalized, the pathogens, either localized in the cytosol or sequestered in membrane-bound compartments, interact extensively with their host environments and modulate these to favor their own survival10. For instance, Salmonella enterica resides and replicates within an intracellular phagosomal compartment termed the Salmonella-containing vacuole (SCV) upon infection11. The maturing SCV traffics towards the Golgi apparatus, undergoing continuous interactions with the endocytic pathway, and induces formation of extensive tubular structures, such as Salmonella-induced filaments (SIF), sorting nexin tubules, Salmonella-induced secretory carrier membrane protein 3 (SCAMP3) tubules, etc.12-14. Studying how these bacterial pathogens manipulate host-cell pathways is essential to understanding infectious disease.
Here, gold-BSA-rhodamine NPs were used as fluid tracers to label the host cellular endo-lysosomal system, and the human gastrointestinal pathogen Salmonella enterica serovar Typhimurium (Salmonella) was used as a model bacterium to study the interactions of the pathogen with the host endocytic pathway. Intracellular gold-BSA-rhodamine NPs in non-infected cells and cells infected with WT Salmonella or mutant strains were imaged by a confocal laser-scanning microscope (CLSM). Then Imaris software was used to quantify the distribution of NPs, indicating that Salmonella infection induced extreme rearrangement of the endosomes/lysosomes. Following the description of this method, analogous experiments can be designed to track long-term fate of the internalized NPs and to investigate the influence of various exogenous substances or endogenous factors on the endocytic pathway of eukaryotic cells.
Protocol
1. Synthesis of 10 nm Gold Nanoparticles (Gold NPs)15
Prepare solution A: add 2 ml 1% aqueous gold chloride into 160 ml Milli-Q, or double distilled, water.
Prepare solution B: add 8 ml 1% tri-sodium citrate x 2 H2O and 160 µl 1% Tannic acid into 32 ml Milli-Q, or double distilled, water.
Warm up solution A and B to 60 °C and mix them while stirring. Observe a dark blue color immediately. Observe red color after about 15 min. Then heat up to 95°C, keep 5 min and cool the solution to RT.
Add a drop of the NP suspension onto a carbon coated grid and allow to dry in air. Check the size and morphology of NPs by transmission electron microscopy.
2. Coating of Gold NPs with BSA and Labeling with Rhodamine N-hydroxysuccinimidyl Ester (NHS)16
Add 900 μl gold NPs into a 1.5 ml Eppendorf tube, centrifuge at 15,000 x g for 30 min. Optionally, prepare multiple tubes may at once.
Discard the supernatant, re-suspend the pellet in 900 μl sterilized Milli-Q water and add 100 μl 2 mg/ml BSA/PBS, mix on a Vortex at 800 rpm for 30 min.
To remove excess BSA, centrifuge the preparation at 15,000 x g for 60 min.
Discard the supernatant, and re-suspend the gold-BSA NPs in 125 μl PBS, add 12.5 µl of 1 M bicarbonate.
Immediately before use, prepare a 10 mg/ml solution of rhodamine NHS/DMSO, add 30 µl of rhodamine NHS/DMSO solution to 1ml of the gold-BSA NPs suspension. Incubate the reaction for 2 hr (or O/N) at RT during 800 rpm mixing, avoiding exposure to light.
Purify the gold-BSA-rhodamine NPs through dialysis against PBS at 4 ºC with 5 buffer changes over the period of 36 - 48 hr.
To stabilize NPs, add 10 μl of 200 mg/ml BSA/PBS into each 1 ml gold-BSA-rhodamine NPs. To remove the free or released BSA-rhodamine, centrifuge at 15,000 x g for 60 min. Resuspend the pellet in 2 mg/ml BSA/PBS, measure OD520, and store at 4 °C, avoiding exposure to light.
Dilute the NPs 10 times with Milli-Q or double distilled water, add a drop onto a carbon-coated grid, and allow to dry in air. Check size and morphology of NPs by transmission electron microscopy (TEM). NOTE: All materials used for NP preparation were sterilized and the operation was conducted in a cell culture hood or on a bench beside a flame.
3. Culture of HeLa Cells
HeLa cells permanently expressing LAMP1-GFP are cultured in DMEM with 10% fetal calf serum (FCS) and grown at 37 °C in an atmosphere containing 5% CO2.
Seed the cells at a density of 50,000 per well in an 8-well chamber slide (Ibidi) and incubate O/N.
4. Culture of Bacteria
Use Salmonella enterica serovar Typhimurium NCTC 12023 wild-type (WT) strain. For comparison, use mutant strains ΔssaV defective in the SPI2-T3SS and ΔsifA lacking key effector SifA for SIF. Use strains harboring plasmid pFPV25.1 for constitutive expression of enhanced GFP. NOTE: Bacterial strains are routinely cultured in Luria-Bertani broth (LB) with addition of 50 µg/ml carbenicillin (WT) and LB with addition of 50 µg/ml carbenicillin and/or 50 µg/ml kanamycin (ΔssaV, ΔsifA) of required to maintain plasmids.
Inoculate a single colony of bacteria in 3 ml LB with appropriate antibiotics and grow O/N at 37 ºC under shaking conditions for aeration, then dilute 1:31 in fresh LB and continue growth for 3.5 hr. At this time-point, the cultures reach the late log phase and bacteria are highly invasive. A ‘roller drum’ is convenient to incubate test tube cultures with aeration.
5. Infection of HeLa cells by Salmonella and Gold-BSA-Rhodamine NPs Pulse Chase-labeling (see Figure 1 for scheme)
Measure OD600 of the sub-cultured bacteria and dilute to OD600 = 0.2 in 1 ml PBS (~3 × 108 cfu/ml), add appropriate amounts of bacteria to HeLa cells in 8-well chamber slides to achieve a multiplicity of infection (MOI) of 100.
Incubate for 30 min in the cell incubator, wash 3 times with PBS to remove non-internalized bacteria (this time point was set as 0 hr post-infection, or 0 hr p.i.). Add 300 µl of fresh culture medium containing 100 µg/ml gentamicin and maintain for 1 hr. Then replace the medium with fresh medium containing 10 µg/ml gentamicin for the rest of the incubation time.
After incubation with culture medium containing 100 µg/ml gentamicin for 1 hr, the medium is replaced by imaging medium (Eagles MEM without FCS, L-glutamine, phenol red and sodium bicarbonate, with 30 mM HEPES, pH 7.4) containing 10 µg/ml gentamicin. Gold-BSA-rhodamine NPs are added to HeLa cells to obtain a final concentration of OD520 = 0.1. NOTE: Gold-BSA-rhodamine NPs may also be added to cells before infection, or at various time-points p.i.
After 1 hr incubation, remove the medium, wash 3 times with PBS, and add 300 µl of fresh imaging medium containing 10% FCS and 10 µg/ml gentamicin for the rest of the incubation time. NOTE: Duration of incubation may vary depending on concentration of NPs and cell lines used. For RAW264.7 macrophages, we observed that 30 min incubation with NPs at a concentration of OD520 = 0.05 allowed sufficient internalization.
6. Imaging
Use a confocal imaging system such as a confocal laser-scanning (CLSM) or spinning disc (SD) microscope with a humidified environment chamber for high resolution imaging at different time-points.
Switch on the temperature control system and wait until it is stable. Optimize the imaging settings such as magnification, scanning speed, resolution, Z-stack etc. Use appropriate excitation/emission settings for GFP and gold-BSA-rhodamine NPs. For this protocol, use a 400 Hz scanning speed, resolution of 512 x 512 pixels and Z-step size of 0.25 µm. Excite GFP and gold-BSA-rhodamine using an Ar laser (488 nm) and a HeNe laser (543 nm), respectively. Include a bright-field (BF) channel for observing the shape of the cells. For other microscopy systems and infection conditions, the setting have to be adjusted accordingly. NOTE: Use the same Ar laser as the light source of BF channel and signal was detected with a photo multiplier (PMT) detector.
At indicated time-points p.i., mount the 8-well chamber slides containing infected cells on the microscope stage and record images.
7. Analysis of Images
Use microscopy image analysis software (see Materials and Equipment’s Table) to analyze the remodeling of the endo-lysosomal system by intracellular Salmonella. Open the data by clicking ‘Open’ and choosing the file. NOTE: Alternatively, open source software packages such as ImageJ or FIJI may be used to quantitative image analyses.
In the objects toolbar of the Surpass view click on the icon
 to add a new surface item. Click on
to add a new surface item. Click on  (Next).
(Next).To analyze a Region of Interest (ROI), select segment a ROI. In a viewing area, a rectangle-bordered section overlaid on image is representing the ROI. Enter the values in the corresponding x-, y-, and z- fields, or directly click on the arrows in the preview rectangle to modify size and position of the ROI. Then click
(Next).As a source channel, select Channel 3 (signal for Gold-BSA-Rhodamine NPs). Check the smooth option to set up the smoothness of the resulting area. Define a value manually or accept the automatically generated value. For Threshold, select the Absolute Intensity option.
For the threshold adjustment, select the manual option and set a value. In the viewing area, a surface threshold preview is displayed in gray.
On the tab Classify Surfaces the resulting surface can be filtered by various criteria. A default filter is ‘number of voxels >10’, and other filters can also be included by clicking ‘add’. If a filtering is not necessary, then delete all filters by clicking on the delete button. Click
(Next).To complete Surface creation, click on
 (Finish). In the Object list, now un-check the box for the item Volume and new created surface is displayed in viewing area.
(Finish). In the Object list, now un-check the box for the item Volume and new created surface is displayed in viewing area.Export the statistics. Now, in the Surpass view the color of the subjects vary from purple to red according to their area. And the ‘Plot Numbers Area’ table shows the statistics information (e.g., ID number and area of objects). Export all statistics of surface 1 to an Excel file by click
 (Save).
(Save).
Representative Results
Gold NPs were generated through a well-established method via reduction of chloroauric acid by citrate and tannic acid. As shown in Figure 2A, the synthesized gold NPs were quasi-spherical in shape with a size of approximately 10 nm. BSA-coating and rhodamine-labeling did not influence their morphology or size (Figure 2B).
It has been reported that gold NPs can be readily taken up by various mammalian cells and ended up in the endocytic systems7. In accordance with previous works, bright red fluorescence signals were observed in HeLa cells after 1 hr incubation with gold-BSA-rhodamine NPs (Figure 3, WT 5 hr p.i.), indicating effective internalization of the NPs by cells. Most of the red signals were found colocalized with green signals, showing that the NPs were confined in late endosomes or lysosomes. However, unlike uniform distribution of NPs in late endosomes/lysosomes in non-infected cells (Figure 3, mock), their locations were largely rearranged in WT Salmonella-infected cells. In the early phase of infection (Figure 3, WT 5 hr p.i.), Salmonella replicates within the SCV and SIF undergo fast extension or contraction movement. At this time point, a fraction of the NPs was found in SCV and SIF, while another fraction was still located in free late endosomes/lysosomes. At 8 hr p.i. (Figure 3, WT 8 hr p.i.), a stabilized network of SIF was formed, and most NPs were found accumulated within the tubular structures, while only a very small portion remained located in free late endosomes/ lysosomes. The mutant strains ΔssaV and ΔsifA exhibited distinct behaviors. As shown in Figure 3 (ΔssaV), at 8 hr p.i., ΔssaV was confined inside SCV while no SIF structures were formed. Some NPs were observed surrounding Salmonella inside the SCV, but the majority of NPs were still located in free late endosomes/lysosomes. For the ΔsifA strain (Figure 3, ΔsifA), escape of Salmonella into cytoplasm occurred, and no association between NPs and bacteria was observed.
In our previous study it was found that SIF display highly dynamic properties in the early phase of infection (3 - 5 hr p.i.), but become stabilized in the later period (>8 hr p.i.)12. Therefore, we are wondering whether Salmonella in early and late phases of infection have comparable ability to rearrange distribution of intracellular NPs. As shown in Figure 4, gold NPs were incubated with HeLa cells O/N before infection, or at different time-points after infection (1 - 7 hr p.i.) for 1 hr, in all cases most of the internalized NPs were observed accumulated in SCV or SIF.
Microscopic observations revealed that infection by Salmonella results in massive rearrangement of the endo-lysosomal system of host cells. As a next step, we used the Imaris software package to analyze Salmonella-induced host cell phenotypes in a quantitative manner. Using the image of one infected HeLa cell at 8 hr p.i. as an example, a step-by-step instruction for quantitative analysis is depicted in Figure 5. Through an intensity threshold of 15 and a ‘number of voxels >10’ filter, 3D objects were extracted from the original image file. The objects were supposed to be intracellular structures in which the gold-BSA-rhodamine NPs located with. In this example, 67 objects were extracted in total, with a summed area as 1,974.64 µm2. The largest object, which colocalized with the SIF network, has an area of 1,836.23 µm2, while the others are smaller than 50 µm2. For comparison, one example showing the quantitative analysis result of a non-infected cell was given in. Here, 431 objects were extracted in total, within which the average value of area is 5 µm2 and the largest object has an area of 34 µm2. The summed area of the objects is 2,252 µm2, which is comparable to the infected cell in Figure 4 (1,975 µm2). However, the number of objects is much larger than the other one (431 and 67 objects, respectively), which indicates large extent fusion of vesicles with the Salmonella-induced tubular structures.
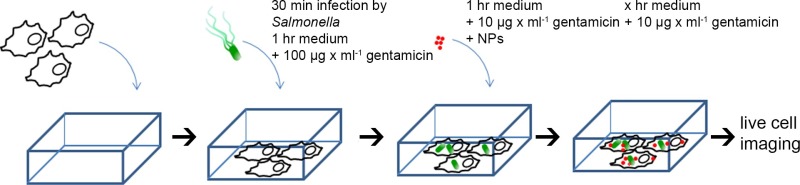 Figure 1. Scheme for preparation of HeLa cells for live cell imaging. HeLa cells were seeded in chamber slides, mock-infected, or infected with various Salmonella strains for 30 min, washed 3 times with PBS, then incubated with medium containing 100 µg/ml gentamicin for 1 hr. Subsequently, the medium was replaced by imaging medium containing 10 nm gold-BSA-rhodamine NPs (OD520 = 0.1) and 10 µg/ml gentamicin, incubated for 1 hr, and then chased by NP-free medium for the rest of the experiment. Live cell imaging was performed at different time points post infection (p.i.). Please click here to view a larger version of this figure.
Figure 1. Scheme for preparation of HeLa cells for live cell imaging. HeLa cells were seeded in chamber slides, mock-infected, or infected with various Salmonella strains for 30 min, washed 3 times with PBS, then incubated with medium containing 100 µg/ml gentamicin for 1 hr. Subsequently, the medium was replaced by imaging medium containing 10 nm gold-BSA-rhodamine NPs (OD520 = 0.1) and 10 µg/ml gentamicin, incubated for 1 hr, and then chased by NP-free medium for the rest of the experiment. Live cell imaging was performed at different time points post infection (p.i.). Please click here to view a larger version of this figure.
 Figure 2. TEM images of colloidal Gold NPs before (a) and after labeling (b).
Please click here to view a larger version of this figure.
Figure 2. TEM images of colloidal Gold NPs before (a) and after labeling (b).
Please click here to view a larger version of this figure.
 Figure 3. Remodeling of the host cellular endo-lysosomal system by intracellular Salmonella. HeLa cells were mock-infected, or infected with Salmonella WT, ΔssaV or ΔsifA mutant strains and pulse/chase with 10 nm gold-BSA-rhodamine NPs (OD520 = 0.1) was performed as described in Figure 1. Maximal intensity projections of CLSM Z-stacks are shown. Scale bars, 10 µm. Please click here to view a larger version of this figure.
Figure 3. Remodeling of the host cellular endo-lysosomal system by intracellular Salmonella. HeLa cells were mock-infected, or infected with Salmonella WT, ΔssaV or ΔsifA mutant strains and pulse/chase with 10 nm gold-BSA-rhodamine NPs (OD520 = 0.1) was performed as described in Figure 1. Maximal intensity projections of CLSM Z-stacks are shown. Scale bars, 10 µm. Please click here to view a larger version of this figure.
 Figure 4. Accessibility of the endo-lysosomal system at various phases of Salmonella infection. HeLa cells were pulsed with 10 nm gold-BSA-rhodamine NPs (OD520 = 0.1) O/N prior infection, or for 1 hr at various time-points p.i. as indicated. Live cell imaging was performed at 8 - 9 hr p.i. Maximal intensity projections of CLSM Z-stacks are shown. Scale bars, 10 µm. Please click here to view a larger version of this figure.
Figure 4. Accessibility of the endo-lysosomal system at various phases of Salmonella infection. HeLa cells were pulsed with 10 nm gold-BSA-rhodamine NPs (OD520 = 0.1) O/N prior infection, or for 1 hr at various time-points p.i. as indicated. Live cell imaging was performed at 8 - 9 hr p.i. Maximal intensity projections of CLSM Z-stacks are shown. Scale bars, 10 µm. Please click here to view a larger version of this figure.
 Figure 5. Step by step instruction for quantitative analysis by Imaris using an image of an infected HeLa cell at 8 hr p.i. as an example. (A) Open the data by clicking ‘Open’ and choosing the file. (B) In the objects toolbar of the Surpass view add a new surface item. (C) To analyze a ROI, then select segment a ROI. (D) Enter the values in the corresponding x-, y-, and z-fields or (E) directly click on the arrows in the preview rectangle to modify the size and the position of the ROI. In this example, a ROI was not need and the whole image was analyzed. (F) As a source channel select the Channel 3 (signal of Gold-BSA-Rhodamine NPs). Check the smooth option to set up the smoothness of the resulting area. Define a value manually or accept the automatically generated value. Here, 0.1 µm was used. For ‘Threshold’ select the ‘Absolute Intensity’ option. (G) For the threshold adjustment select the manual option and set a value (in this example 15 was used). In the viewing area a surface threshold preview is displayed in gray. (H) On the tab ‘Classify Surfaces’ the resulting surface can be filtered by various criteria. A default filter is ‘number of voxels >10’. The filter can be adjusted by enter a new value and other filters can be added. In this example, we kept the filter ‘number of voxels >10’. (I) To complete Surface creation click on ‘Finish’. In the Object list now un-check the box for the item Volume and new created surface is displayed in viewing area in red. (J) In the Surpass view the color of the subjects vary from purple to red according to their area. (K) The ‘Plot Numbers Area’ table shows the statistics information. Export statistics to an Excel file. Please click here to view a larger version of this figure.
Figure 5. Step by step instruction for quantitative analysis by Imaris using an image of an infected HeLa cell at 8 hr p.i. as an example. (A) Open the data by clicking ‘Open’ and choosing the file. (B) In the objects toolbar of the Surpass view add a new surface item. (C) To analyze a ROI, then select segment a ROI. (D) Enter the values in the corresponding x-, y-, and z-fields or (E) directly click on the arrows in the preview rectangle to modify the size and the position of the ROI. In this example, a ROI was not need and the whole image was analyzed. (F) As a source channel select the Channel 3 (signal of Gold-BSA-Rhodamine NPs). Check the smooth option to set up the smoothness of the resulting area. Define a value manually or accept the automatically generated value. Here, 0.1 µm was used. For ‘Threshold’ select the ‘Absolute Intensity’ option. (G) For the threshold adjustment select the manual option and set a value (in this example 15 was used). In the viewing area a surface threshold preview is displayed in gray. (H) On the tab ‘Classify Surfaces’ the resulting surface can be filtered by various criteria. A default filter is ‘number of voxels >10’. The filter can be adjusted by enter a new value and other filters can be added. In this example, we kept the filter ‘number of voxels >10’. (I) To complete Surface creation click on ‘Finish’. In the Object list now un-check the box for the item Volume and new created surface is displayed in viewing area in red. (J) In the Surpass view the color of the subjects vary from purple to red according to their area. (K) The ‘Plot Numbers Area’ table shows the statistics information. Export statistics to an Excel file. Please click here to view a larger version of this figure.
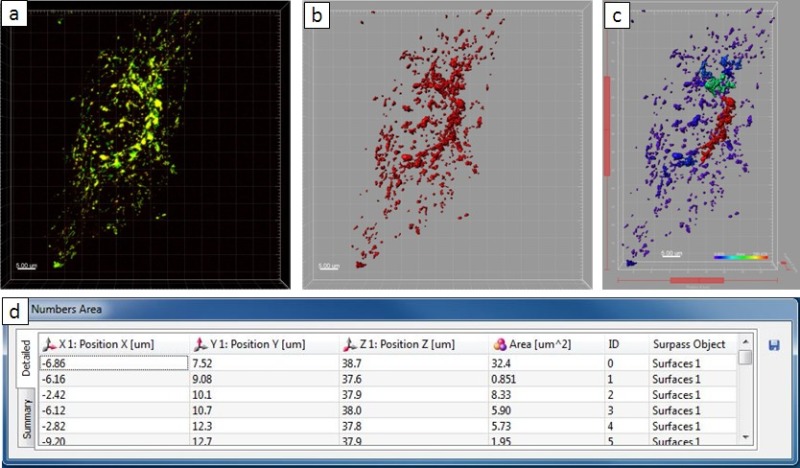 Figure 6. Quantitative analysis result of a non-infected cell. (A) Confocal microscope image. (B) The new created surface. (C) The new surface in the Surpass view. (D) The ‘Plot Numbers Area’ table shows the statistics information. Please click here to view a larger version of this figure.
Figure 6. Quantitative analysis result of a non-infected cell. (A) Confocal microscope image. (B) The new created surface. (C) The new surface in the Surpass view. (D) The ‘Plot Numbers Area’ table shows the statistics information. Please click here to view a larger version of this figure.
Discussion
The endo-lysosomal system of mammalian cells controls important physiologic processes, including nutrient absorption, hormone-mediated signal transduction, immune surveillance, and antigen presentation17. Up to now, a variety of markers have been used for labeling of the endocytic pathway and tracking studies. For example, LysoTracker probes are fluorescent acidotropic probes developed by Molecular Probes (Life Technologies, USA) for lysosome labeling, which can selectively accumulate in cellular compartments with low internal pH and effectively label living cells at nanomolar concentrations18. However, long time incubation of LysoTracker probes with cells may induce an increase in lysosomal pH and lead to potential physiological changes of lysosomes19. Some large biomolecules, such as fluorophore-labeled dextrans, are also frequently used for endosome/lysosomes labeling. However, low-photostability, short-circulating life, and poor retention during fixation hinder their applications in long-term live cell imaging and correlative microscope studies16,19. Here, gold-BSA-rhodamine NPs were used as fluid tracers to label the host cellular endo-lysosomal system. High uptake efficiency and strong intracellular fluorescence signals were observed. The almost complete colocalization of NPs with lysosomal glycoprotein LAMP1, which is highly enriched on the membrane of late endosomes/lysosomes, verified proper labeling of the endo-lysosomal system by the NPs. Combining the use of fluorescent NPs with the other markers such as Lysotracker may provide complementary information about host cell vesicular trafficking and maturation.
It is worth to mention that cellular internalization of NPs is highly dependent on their physical dimensions and chemical components. We have tested gold-BSA-rhodamine NPs with a size of 5 nm, 10 nm, 15 nm and 30 nm, and similar intracellular distribution inside HeLa cells were observed (results not shown). We used HeLa cells as convenient cell line for Salmonella infection studies. However, gold-BSA-rhodamine NPs can also be readily internalized by various other cell lines and primary cells are amenable. For example, we observed Salmonella-induced tubular structures and labeling by NP in CHO, COS-7, CaCo2, or 3T3 cells, as well as in Interferon-γ-stimulated RAW264.7 macrophage-like cell line cells or primary murine macrophages and dendritic cells. However, each cell line will require adjustment of the infection and pulse/chase protocols.
It has been reported that dye-entrapped silica NPs can be used as a biocompatible, long-living, and highly photostable lysosome marker19, and we also compared the behavior of rhodamine-doped silica NPs with varying size (30 - 1,000 nm). Proper labeling of late endosomes/lysosomes by silica NPs in non-infected cells and accumulation of NPs in SCV and SIF in Salmonella-infected cells was observed. However, due to the formation of aggregates of NPs or the large size of single silica NPs, distribution of NPs along with Salmonella-induced tubular structures was not uniform (data not shown).
A characteristic feature of SCV as well as SIF is the presence of highly abundant lysosomal glycoproteins such as LAMP112. Here, a HeLa cell line stably expressing LAMP1-GFP was used for the convenience of live cell imaging of the endo-lysosomal system and the SCV and SIF structures. Salmonella strains constitutively expressing enhanced GFP were used in the infection experiments to indicate location of bacteria. Due to the uniform size and shape of the intracellular bacteria, the bacterial GFP fluorescence in general is easily distinguishable from the GFP label on endo-lysosomal membranes. However, for future experiments in which fluorescent signal from bacteria and membrane proteins need to be precisely differentiate, other fluorescent fusion proteins, e.g., mTurquoise, are recommended to be integrated. This, however, results in additional rounds of illumination and image acquisition, bearing the risk of higher photo-damage in live cell experiments.
To find out essential genes and mechanisms for bacterial pathogens to manipulate host-cell pathways, numerous mutants need to be created and their performances need to be carefully compared. For example, Salmonella mutant strains ΔssaV and ΔsifA are highly attenuated in systemic virulence and intracellular replication20,21. Both ΔssaV and ΔsifA strains fail to induce SIF, and additionally ΔsifA Salmonella lose the SCV membrane during the course of infection12. Comparing phenotypes of WT and mutant strains is instrumental in understanding the ability of Salmonella to remodel host cell vesicular traffic. Based on microscopic examination, it is obvious that WT Salmonella subvert host cellular endocytic pathways to a much higher extent compared to ΔssaV and ΔsifA mutant strains. This function might enable intracellular Salmonella to obtain more nutrients for their intracellular survival and replication.
Confocal microscopy images clearly demonstrate the remodeling of host cellular endo-lysosomal system by intracellular bacteria. However, quantitative analysis of the images is required for precise comparison. In the method described here, Imaris software is used for extracting 3D objects which have fluorescence intensity in Channel 3 > 15 and number of voxels > 10. These objects are supposed to be intracellular membrane structures containing Gold-BSA-Rhodamine NPs. The fluorescence intensity threshold and filters can be defined according to the quality of images, but should be kept constant for comparing different conditions or strains. Shown here are examples of a non-infected cell and a WT Salmonella-infected cell at 8 hr p.i., with 431 and 67 objects extracted, respectively. Despite the apparent difference, it is not rational to compare only the number of the objects, since the size of the cells are not the same. One solution is to normalize the number of the objects to the summed area (in this example, 2,252 µm2 and 1,975 µm2, respectively) or the sum of fluorescent intensities of the objects inside cells. Alternatively, it is also possible to track one cell before infection and at different time-points p.i. in order to compare the distribution of NPs at different stages of infection.
In conclusion, in this method gold-BSA-rhodamine NPs were applied to label the endo-lysosomal system of eukaryotic cells. The manipulation of the host cellular endo-lysosomal system by an intracellular pathogen was observed by live cell CLSM and quantitative analysis results indicated extreme rearrangements of late endosomes/lysosomes by WT Salmonella. Salmonella was used as a model pathogen in this study, but intracellular behavior of other bacterial pathogens, such as Shigella flexneri and Listeria monocytogenes could also be investigated using this approach. In principle, gold-BSA-rhodamine NPs could be internalized by a large variety of cell lines, therefore are promising being broadly applied in studies investigating interaction of the endocytic pathway with divergent endogenous or exogenous substances.
Disclosures
No conflicts of interest declared.
Acknowledgments
This work was supported by the Deutsche Forschungsgemeinschaft by grant Z within Sonderforschungsbereich 944 ‘Physiology and Dynamics of Cellular Microcompartments’ and HE1964/18 within priority program 1580.
References
- Coto-Garcia AM. Nanoparticles as fluorescent labels for optical imaging and sensing in genomics and proteomics. Anal. Bioanal. Chem. 2011;399:29–42. doi: 10.1007/s00216-010-4330-3. [DOI] [PubMed] [Google Scholar]
- Xie J, Lee S, Chen X. Nanoparticle-based theranostic agents. Adv. Drug Deliv. Rev. 2010;62:1064–1079. doi: 10.1016/j.addr.2010.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruedas-Rama MJ, Walters JD, Orte A, Hall EA. Fluorescent nanoparticles for intracellular sensing: a review. Anal. Chim. Acta. 2012;751:1–23. doi: 10.1016/j.aca.2012.09.025. [DOI] [PubMed] [Google Scholar]
- Wu C, Chiu DT. Highly fluorescent semiconducting polymer dots for biology and medicine. Angew. Chem. Int. Ed. Engl. 2013;52:3086–3109. doi: 10.1002/anie.201205133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giepmans BN, Deerinck TJ, Smarr BL, Jones YZ, Ellisman MH. Correlated light and electron microscopic imaging of multiple endogenous proteins using Quantum dots. Nat. Methods. 2005;2:743–749. doi: 10.1038/nmeth791. [DOI] [PubMed] [Google Scholar]
- Kumar D, Saini N, Jain N, Sareen R, Pandit V. Gold nanoparticles: an era in bionanotechnology. Expert Opin. Drug Deliv. 2013;10:397–409. doi: 10.1517/17425247.2013.749854. [DOI] [PubMed] [Google Scholar]
- Dykman LA, Khlebtsov NG. Uptake of engineered gold nanoparticles into mammalian cells. Chem. Rev. 2014;114:1258–1288. doi: 10.1021/cr300441a. [DOI] [PubMed] [Google Scholar]
- Chithrani BD, Ghazani AA, Chan WC. Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells. Nano Lett. 2006;6:662–668. doi: 10.1021/nl052396o. [DOI] [PubMed] [Google Scholar]
- Finlay BB, Cossart P. Exploitation of mammalian host cell functions by bacterial pathogens. Science. 1997;276:718–725. doi: 10.1126/science.276.5313.718. [DOI] [PubMed] [Google Scholar]
- Bhavsar AP, Guttman JA, Finlay BB. Manipulation of host-cell pathways by bacterial pathogens. Nature. 2007;449:827–834. doi: 10.1038/nature06247. [DOI] [PubMed] [Google Scholar]
- Malik-Kale P, et al. Salmonella - at home in the host cell. Front. Microbiol. 2011;2:125. doi: 10.3389/fmicb.2011.00125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajashekar R, Liebl D, Seitz A, Hensel M. Dynamic remodeling of the endosomal system during formation of Salmonella-induced filaments by intracellular Salmonella enterica. Traffic. 2008;9:2100–2116. doi: 10.1111/j.1600-0854.2008.00821.x. [DOI] [PubMed] [Google Scholar]
- Schroeder N, Mota LJ, Meresse S. Salmonella-induced tubular networks. Trends Microbiol. 2011;19:268–277. doi: 10.1016/j.tim.2011.01.006. [DOI] [PubMed] [Google Scholar]
- Drecktrah D, Knodler LA, Howe D, Steele-Mortimer O. Salmonella trafficking is defined by continuous dynamic interactions with the endolysosomal system. Traffic. 2007;8:212–225. doi: 10.1111/j.1600-0854.2006.00529.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slot JW, Geuze HJ. A new method of preparing gold probes for multiple-labeling cytochemistry. Eur. J. Cell Biol. 1985;38:87–93. [PubMed] [Google Scholar]
- Zhang Y, Hensel M. Evaluation of nanoparticles as endocytic tracers in cellular microbiology. Nanoscale. 2013;5:9296–9309. doi: 10.1039/c3nr01550e. [DOI] [PubMed] [Google Scholar]
- Pollard TD, Earnshaw WC, Lippincott-Schwartz J. Chapter 22. Cell Biology. 2007.
- LysoTracker and LysoSensor Probes. Carlsbad, CA: Life Technologies Corporation; 2013. [Google Scholar]
- Shi H, He X, Yuan Y, Wang K, Liu D. Nanoparticle-based biocompatible and long-life marker for lysosome labeling and tracking. Anal. Chem. 2010;82:2213–2220. doi: 10.1021/ac902417s. [DOI] [PubMed] [Google Scholar]
- Hensel M. Genes encoding putative effector proteins of the type III secretion system of Salmonella pathogenicity island 2 are required for bacterial virulence and proliferation in macrophages. Mol. Microbiol. 1998;30:163–174. doi: 10.1046/j.1365-2958.1998.01047.x. [DOI] [PubMed] [Google Scholar]
- Beuzon CR, et al. Salmonella maintains the integrity of its intracellular vacuole through the action of SifA. EMBO J. 2000;19:3235–3249. doi: 10.1093/emboj/19.13.3235. [DOI] [PMC free article] [PubMed] [Google Scholar]