

Abstract
Background
Psychological stressors may cause affective disorders, such as depression and anxiety, by altering expressions of corticotropin releasing factor (CRF), serotonin (5-HT), and tyrosine hydroxylase (TH) in the brain. This study investigated the effects of essential oil from Asarum heterotropoides (EOAH) on depression-like behaviors and brain expressions of CRF, 5-HT, and TH in mice challenged with stress.
Methods
Male ICR mice received fragrance inhalation of EOAH (0.25, 0.5, 1.0, and 2.0 g) for 3 h in the special cage capped with a filter paper before start of the forced swimming test (FST) and tail suspension test (TST). The duration of immobility was measured for the determination of depression-like behavior in the FST and TST. The selective serotonin reuptake inhibitor fluoxetine as positive control was administered at a dose of 15 mg/kg (i.p.) 30 min before start of behavioral testing. Immunoreactivities of CRF, 5-HT, and TH in the brain were also measured using separate groups of mice subjected to the FST.
Results
EOAH at higher doses (1.0 and 2.0 g) reduced immobility time in the FST and TST. In addition, EOAH at a dose of 1.0 g significantly reduced the expected increases in the expression of CRF positive neurons in the paraventricular nucleus and the expression of TH positive neurons in the locus coeruleus, and the expected decreases of the 5-HT positive neurons in the dorsal raphe nucleus.
Conclusion
These results provide strong evidence that EOAH effectively inhibits depression-like behavioral responses, brain CRF and TH expression increases, and brain 5-HT expression decreases in mice challenged with stress.
Keywords: Essential oil, Asarum heterotropoides, Depression, Inhalation
Background
Physical and psychological stressors are thought to cause affective disorders including depression and anxiety [1,2]. The neurobiological substrate for depression-like behaviors induced by stress is believed to involve corticotropinergic neurons in the paraventricular nucleus, noradrenergic neurons in the locus coeruleus, and serotonergic system in the dorsal raphe nucleus [3,4]. The involvement of these substrates is supported by the report that selective serotonin reuptake inhibitor (SSRI), serotonin-norepinephrine reuptake inhibitor (SNRI), norepinephrine-dopamine reuptake inhibitor, and inhibitor of hypothalamic-pituitary-adrenal (HPA) axis are used for the treatment of depression [5]. However, the clinical satisfaction is still under debating due to unwanted side effects and genetic variation. Therefore, the alternative medication is necessary to provide effective approaches for reducing the depression.
Studies in laboratory animals and humans have demonstrated that inhalation of essential oils can produce antidepressant and anxiolytic effects exposed to stress by modulating the central nervous system. For examples, inhalation of lemon oil reduced anxiogenic and depressant effects in an elevated plus-maze test and a forced swimming test (FST) by modulating serotonergic and dopaminergic pathways in mice brain [6]. In addition, lavender oil inhalation decreased anxiety and depression-like behaviors of rats in an elevated plus-maze test and a FST [7]. Lavender oil inhalation is also effective in reducing stress-related symptoms in nurses [8].
Asarum heterotropoides Fr. Mandshuricum has been shown to be effective for reducing anxiety and for the relief of pain [9,10]. Furthermore, biological and pharmacological studies have demonstrated that Asarum heterotropoides can produce wide spectrums of action including anti-inflammatory effect [11,12] and anti-allergic [13] and antioxidant activity [14].
The present study was carried out to investigate the effects of inhalation of essential oil from Asarum heteropoides (EOAH) on depression–like behaviors in the FST and tail suspension test (TST). A role for brain corticotropin releasing factor (CRF), serotonin (5-HT), and tyrosine hydroxylase (TH) expression in EOAH effects is also explored.
Methods
Animals
Male mice of ICR strain (Orient Inc., South Korea) at 5 weeks after the birth were housed under a controlled temperature (23 ± 2°C) and humidity (50 ± 10%) with a 12 h light–dark cycle (lights on at 8:00 h). Food and water were made available ad libitum for the study. All animal experiments were carried out in accordance with the animal care guidelines of the National Institute of Health (NIH) and approved by the Institutional Animal Care and Use Committee of Daegu Hanny University.
Preparation of EOAH and chemical analysis
The dried and chipped roots of Asarum heterotropoides (1.2 kg) were purchased from Daegu Yangnyeongsi (a traditional herb market) in Daegu, South Korea. The pulverized Asarum heterotropoides were used to extract essential oil at the room temperature with n-hexane (5 L x 2) for 24 h. The extracts (EOAH) were filtered and evaporated in the vacuum at 40°C to remove n-hexane. The final concentration of essential oil was 22.5 g. The specimen of this plant material (RCBROM-AEM2012) is deposited at the Research Center for Biomedical Resources of Oriental Medicine, Daegu Haany University. EOAH was analyzed for the determination of chemical composition using GC-MS (Agilent Technologies 7890A/5975C insert XL Mass Selective detector) with injection volume 1 μl (Column: DB-5MS; 250 μm i.d.; 30 m length; 0.25 μm film thickness; split ratio 1:50. Carrier: Helium. Injector 250°C, Detector 280°C, Column 70°C, 1 min; 4°C/min to 300°C for 20 min). The identification of EOAH was carried out by comparing their relative retention time and mass spectra to those generated by chromatographic analysis of standards and W9N08.L library and literature.
Inhalation of EOAH
To evaluate the effects of EOAH on depression-like behaviors, the mice were randomly divided into the following 5 groups: Control (n = 5); EOAH 0.25 g (n = 5); EOAH 0.5 g (n = 5); EOAH 1.0 g (n = 5); EOAH 2.0 g (n = 5). Mice weighing between 25 and 30 g at the start of the experiment were used. The mice were pretreated with inhalation of EOAH (0.25, 0.5, 1.0, and 2.0 g) or saline (control group). The inhalation dose of essential oil was chosen based on previous studies [15,16]. To induce saturation of the fragrance by EOAH in the transparent special cage (W 26 X L 22 X H 20 cm), EOAH put 30 min before the individual inhalation of EOAH for 3 h in the special cage under standardized condition (room temperature: 23 ± 2°C, relative humidity: 50 ± 10%). The special cage was capped with a filter paper, which allowed minimum breathing air to pass. In order to vaporize essential oil efficiently, EOAH was put in uncapped eppendorf tube set on the upper side of an inhalation cage. After inhalation of EOAH, no mice have shown abnormal behaviors including enhanced locomotion, loss of body postures, sweats, head turns and vocalization. The selective serotonin reuptake inhibitor fluoxetine (Sigma-Aldrich, St. Louis, MO, U.S.A.) was dissolved in saline. Mice were treated with 15 mg/kg of fluoxetine (n = 7) as positive control or saline (control group, n = 7) by intraperitoneal injection 30 min before the start of behavioral testing. An injection volume was 0.3 ml/30 g body weight.
FST and TST
Following inhalation of EOAH, the mice were subjected to the FST and TST, respectively. The duration of immobility induced by forced swimming was measured as described previously with slight modification [17]. The mice were individually exposed to forced swimming in a transparent acrylic cylinder (25 cm in height and 10 cm in diameter) containing 20 cm height of water at 23°C. The immobility was defined as floating motionless in the water [18]. The duration of immobility was measured during the last 4 min of total 6 min test using a video based Ethovision System (Noldus, Wageningen, Netherlands). TST was carried out according to the method described previously with a slight modification [19]. In brief, the mice were individually hung 5 cm above the floor in clear black acrylic boxes (30 cm × 30 cm × 50 cm) by tail attachment using an adhesive tape to a hook. The mice were individually allowed to hang for 6 min and the duration of immobility was recorded during the last 4 min of total 6 min test. The immobility was defined as absence of body movements [19].
Immunohistochemistry
The expressions of 5-HT, TH, and CRF were measured using different groups of mice. Mice were divided 4 groups: Normal (n = 8); Control (n = 9); EOAH 0.5 g (n = 9); EOAH 1.0 g (n = 9). At the end of the FST, mice were sacrificed and brains were prepared to measure the expressions of 5-HT, TH, and CRF, respectively. Mice in normal group were not allowed to have the FST. Immunohistological study began with transcardial perfusion of heparinized phosphate-buffered saline (PBS; pH 7.4) for 30 s followed by perfusion of 4% paraformaldehyde in 0.1 M phosphate buffered saline (pH 7.4) for 10–15 min. Brains were removed and allowed to stand in the same fixative by overnight, followed by cryoprotected in 30% sucrose solution in PBS. The individual brain was embedded in OCT compound and serially sectioned on a cryostat (Leica, Nussloch, Germany) at 30 μm thickness by the coronal plane, and then collected in PBS using free floating methods. The individual primary antibody includes anti-5HT (rabbit monoclonal, 1:200; Abcam, Cambridge, U.S.A.), anti-CRF (rabbit monoclonal, 1:200; Abcam, Cambridge, U.S.A.), and anti-TH (rabbit monoclonal, 1:2000; Abcam, Cambridge, U.S.A.) was incubated in a cocktail solution (0.3% PBST, 2% blocking serum and 0.001% keyhole limpet hemocyanin) for 72 h at 4°C, respectively. After three time rinses in PBS, the sections were placed in Vectastain Elite ABC reagent (Vector laboratories, Burlingame, CA, U.S.A.) for 2 h at room temperature. Following a further rinsing in PBS, the tissue was developed using diaminobenzadine (Sigma-Aldrich, St. Louis, MO, U.S.A.) as the chromogen. The images of 5-HT-, CRF-, and TH-immunoreactive neurons were captured using a DP2-BSW imaging system (Olympus, CA, USA) and measured as described by others [20]. In brief, the grid was placed on the target area in the brain and the number of cells was counted at 100 x magnification using a microscope rectangle grid. The five sections were collected at the levels of the dorsal raphe nucleus, the paraventricular nucleus and the locus coeruleus in each brain for 5-HT, CRF and TH immunoreaction. The cells were counted and averaged from five sections.
Statistical analysis
Statistical analysis of data was carried out using SPSS 15.0 for Windows. Depression-like behaviors and brain 5-HT-, TH-, and CRF-immunoreactivity were statistically analyzed by one-way ANOVA and post-hoc Newman-Keuls test to compare the experimental and control groups. An unpaired t-test was performed to determine statistical significance for fluoxetine versus saline comparison. The significance level was set at p < 0.05, p < 0.01, and p < 0.001.
Results
Major compounds of EOAH
The total 78 peaks were detected in EOAH. The main compounds in analyzed samples were methyl eugenol (22.58%), pentadecane (6.78%), 2,3,5-trimethoxytoluene (5.54%), 4-(chloromethyl) cyclohexene (3.36%), myristicine (3.27%), sesamin (3.24%), and kakoul (2.63%).
The effect of EOAH on the depression-like behaviors
Inhalation of EOAH at higher doses (1.0 and 2.0 g) significantly reduced immobility time in the FST (Figure 1A). A one-way ANOVA identified a significant main effect (F4,20 = 3.19, p < 0.05). Post hoc tests revealed that mice receiving saline (control) had significantly greater immobility time than those treated with EOAH during the FST (p < 0.05). When mice were exposed to the TST, inhalation of EOAH (0.25, 0.5, 1.0, and 2.0 g) significantly decreased immobility time (Figure 1C). Interestingly, an antidepressant effect of EOAH at lower doses was more apparent in the TST. A one-way ANOVA identified a significant main effect (F4,20 = 3.63, p < 0.05). Post hoc tests revealed that mice receiving saline had significantly greater immobility time than those treated with EOAH during the TST (p < 0.05). Similarly, administration of fluoxetine markedly reduced immobility time at both the FST and TST compared to saline group (p < 0.01, p < 0.001, respectively, Figure 1B and D). This provides additional evidence that EOAH effectively reduces depression-like behaviors. In our study, EOAH at higher doses significantly suppressed depression-like behaviors in the FST. Thus, EOAH at a dose of 1.0 g was exposed to mice subjected to the FST in order to identify EOAH effects on brain levels of 5-HT, CRF, and TH.
Figure 1.
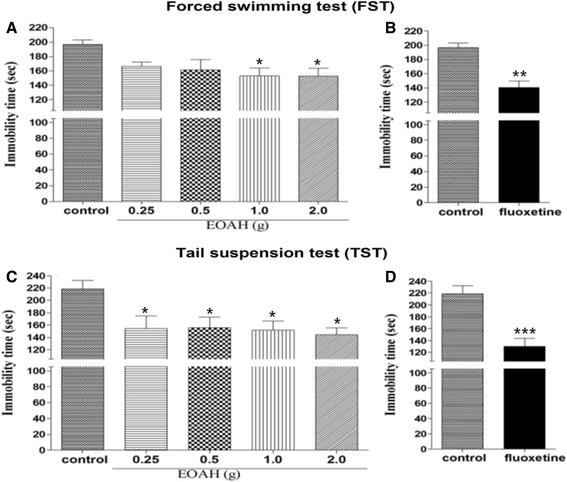
Effects of inhalation of EOAH on the FST and TST in mice. The mice were pretreated with inhalation of EOAH (0.25, 0.5, 1.0, and 2.0 g; EOAH groups) or saline (control group) prior to FST or TST. The graph (A and C) represents the immobility time during a 4 min period in all groups (n = 5 per each group). Data were analyzed by one-way ANOVA and post-hoc Newman-Keuls test. For comparative purpose, mice were administered with fluoxetine or saline. The graph (B and D) represents the immobility time during a 4 min period in all groups (n = 7 per each group). Data were analyzed by t-test. Each value represents the mean ± S.E.M. *p < 0.05 compared to the control group.
5-HT-immunoreactive neurons in the dorsal raphe nucleus
Changes in the expression of 5-HT-immunoreactive neurons were evaluated in mice exposed to the FST following inhalation of EOAH (Figure 2A-D). As shown in Figure 2E, a much larger decrease in the mean number of 5-HT-immunoreactive neurons in the dorsal raphe nucleus was produced in mice exposed to the FST compared with normal mice. While the FST markedly decreased the expression of 5-HT in the dorsal raphe nucleus in mice receiving inhalation of saline (p < 0.01), inhalation of EOAH (1.0 g) increased 5-HT expression in the dorsal raphe nucleus. A one-way ANOVA analysis revealed a significant main effect of EOAH inhalation (F3,31 = 8.339, p < 0.001). Post-hoc comparisons indicated that there was a significant enhancement in the 5-HT activity in the dorsal raphe nucleus of the 1.0 g EOAH group compared with the control group (p < 0.01). Group 0.5 g EOAH mice tended to have greater 5-HT activity in the dorsal raphe nucleus than the control group, but these differences were not statistically significant.
Figure 2.
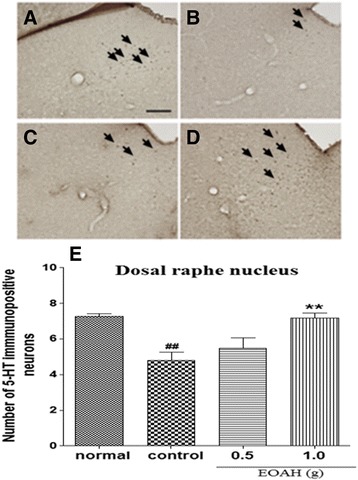
Effect of inhalation of EOAH on 5-HT expression in the dorsal raphe nucleus in mice subjected to the FST. Photographs represent the distribution of 5-HT-immunoreactive neurons in the dorsal raphe nucleus of normal (A, n = 8), control (B, n = 9), EOAH 0.5 g (C, n = 9) and EOAH 1.0 g (D, n = 9) groups. The number of 5-HT immunostained neurons among the groups (E) was analyzed by one-way ANOVA post-hoc Newman-Keuls test. Each value represents the mean ± S.E.M. ## p < 0.01 compared to the normal group, **p < 0.01 compared to the control group. Sections were cut coronally at 30 μm and the scale bar represents 200 μm. Arrowheads indicate 5-HT immunopositive neurons.
CRF-immunoreactive neurons in the paraventricular nucleus
The expression of CRF in the paraventricular nucleus was shown after inhalation of EOAH (Figure 3A-D). Control mice exposed to the FST had significantly higher mean number of CRF-immunoreactive neurons than normal mice (Figure 3E). While the FST markedly increased the expression of CRF in the paraventricular nucleus in mice receiving inhalation of saline (p < 0.001), there was an increase in 5-HT expression in those receiving inhalation of EOAH (0.5 and 1.0 g). A one-way ANOVA revealed a significant main effect of the treatment on the expression of CRF-immunoreactive neurons in the paraventricular nucleus (F3,31 = 11.060, p < 0.001). Post hoc tests indicated that inhalation of EOAH (0.5 and 1.0 g) induced marked increases in the expression of CRF-immunoreactive neurons in the paraventricular nucleus compared with saline-treated mice (p < 0.05, p < 0.001, Figure 3E).
Figure 3.
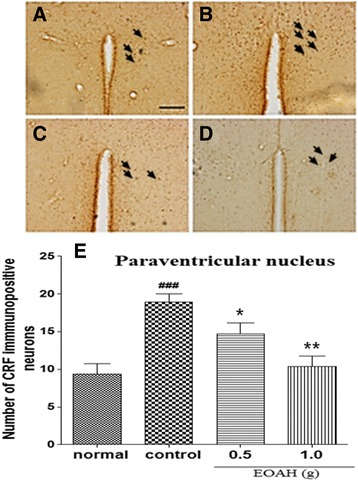
Effect of inhalation of EOAH on CRF expression in the paraventricular nucleus in mice subjected to the FST. Photographs represent the distribution of CRF-immunoreactive cells in the paraventricular nucleus of normal (A, n = 8), control (B, n = 9), EOAH 0.5 g (C, n = 9) and EOAH 1.0 g (D, n = 9) groups. The number of the CRF immunostained neurons among the groups (E) was analyzed by one-way ANOVA post-hoc Newman-Keuls test. Each value represents the mean ± S.E.M. ### p < 0.001 compared to the normal group. *p < 0.05 and **p < 0.01 compared to the control group. Sections were cut coronally at 30 μm and the scale bar represents 200 μm. Arrowheads indicate CRF immunopositive neurons.
TH-immunoreactive neurons in the locus coeruleus
The expression of TH in the paraventricular nucleus was shown after inhalation of EOAH (Figure 4A-D). Similar to the CRF data, mice exposed to the FST showed a significant difference in the mean number of TH-immunoreactive neurons in the locus coeruleus from normal group (p < 0.001, Figure 4E). However, inhalation of EOAH significantly reduced the expression of TH in the locus coeruleus compared to saline inhalation. A one-way ANOVA revealed a significant main effect of the treatment on the expression of TH-immunoreactive neurons in the LC (F3,31 = 36.040, p < 0.001). Post hoc tests indicated that inhalation of EOAH (0.5 and 1.0 g) induced marked increases in the expression of TH-immunoreactive neurons in the locus coeruleus compared with treatment of saline (p < 0.001, Figure 4E).
Figure 4.
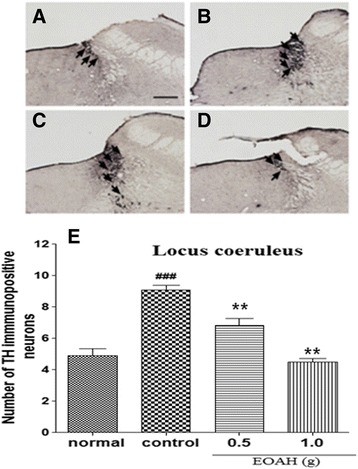
Effect of inhalation of EOAH on TH expression in the locus coeruleus in mice subjected to the FST. Photographs represent the distribution of TH-immunoreactive cells in the locus coerleus of normal (A, n = 8), control (B, n = 9), EOAH 0.5 g (C, n = 9) and EOAH 1.0 g (D, n = 9) groups. The number of the TH-immunostained neurons among the groups (E) was analyzed by one-way ANOVA post-hoc Newman-Keuls test. Each value represents the mean ± S.E.M. ### p < 0.001 compared to the normal group, **p < 0.01 compared to the control group. Sections were cut coronally at 30 μm and the scale bar represents 200 μm. Arrowheads indicate TH immunopositive neurons.
Discussion
The present results demonstrated that the inhalation of EOAH effectively attenuated depression-like behavior, increases in brain expressions of CRF and TH, a decrease in brain expressions of 5-HT in response to 6 min of forced swimming or immobilization stress. Behavioral responses to the FST and TST are consistent with previous findings [3,4]. The FST and TST are useful tests for screening antidepressant drugs and exploring their mechanisms of action, because of its good reliability and predictive validity [21]. The FST and TST commonly trigger psychological despairs with no escapable hope from immobilization by learning that escape from the current situation is impossible. Thus, results in no hope or no trial to escape and consequently develop depression status [17].
Pharmacologically, a variety of anti-depressants and related-compounds with potential anti-depressant activity reduce the duration of immobility in the FST and TST [22,23]. Therefore, the reduction of immobility is the important factor to develop the pharmacological approaches to the treatment of depression-like behaviors in experimental designs. Our results suggest that inhalation of EOAH reduces depression-like behaviors in the FST and TST. However, our results showed that lower doses of EOAH did not produce a significant effect on depression-like behaviors in the FST. This differs with effects of EOAH at lower doses in the TST as demonstrated by a significant decrease in depression-like behaviors. Such inconsistencies may be at least in part related to differences in response to EOAH between the FST and TST. In support of this, one study has shown that TST is more responsive to neurochemical changes than the FST [24]. The present results are similar to others who demonstrated that pure fragrance of aroma oil and essential oil from plants decreased depression-like behaviors in the FST and TST [25,26]. Further support for a role of EOAH is the observation that systemic administration of methyl eugenol, which is known to be one of volatile components of Asarum heterotropoides [27], produced anti-anxiety and anti-depression effects in the FST [28].
Depression-like behaviors induced by stressors have been linked to endogenous systems of CRF and monoamines including 5-HT and NE [29,30]. Pharmacological studies have shown that CRF1 receptor antagonist, 5-HT1 receptor agonist, and adrenoreceptor antagonist can decrease depression-like behaviors, indicating that endogenous systems of CRF and monoamines including 5-HT and NE in the brain contribute to development of depression [31-34]. Thus, it is not unexpected that normalization of these systems attenuates depression-like behaviors [35-37]. In our study, mice subjected to forced swimming stress showed significant increases in brain expressions of CRF and NE, and significant decreases in brain expression of 5-HT. These results are consistent with previous findings, suggesting that endogenous systems of CRF, 5-HT, and NE in the brain plays an important role in modulating depression-like behaviors. For example, the expression of CRF mRNA or CRF immunoreactivity in the hypothalamic neurons is known to be up-regulated by stressor including forced swimming stress [38,39]. Similarly, CRF 2 mutant mice showed increased immobility time in forced swimming test [40]. It is noteworthy that one particular stressor, forced swimming, was associated with decreases of extracellular 5-HT in several brain regions including the dorsal raphe nucleus, lateral septum, and amygdala [41,42]. Also, serotonin receptor 1A knockout (KO) mice or serotonin transporter KO mice exhibited a decreased immobility time in the forced swim test [43,44]. In contrast, exposure to forced swimming considerably enhanced the release of norepinephrine in the locus coeruleus [45,46]. In addition, norepinephrine transporter KO mice have been shown to reduce immobility in the FST and TST [44]. Therefore, it is highly likely that depression-like behaviors are due to activation of CRF neurons in the hypothalamus and noradrenergic neurons in the locus coeruleus and suppression of serotonergic neurons in the dorsal raphe nucleus.
Most importantly, results showed that EOAH at the dose of 1.0 g significantly reduced the expected increases in the expression of CRF positive neurons in the paraventricular nucleus, the expected decreases in the expression of TH in the locus coeruleus and the 5-HT positive neurons in the dorsal raphe nucleus in mice subjected to forced swimming stress. Also, EOAH at the dose of 1.0 g reduced a significant increase in immobility time in the FST and TST. Based on a role for endogenous systems of CRF, 5-HT, and NE in the brain, one possible mechanism where EOAH could diminish depression-like behaviors is by affecting brain levels of 5-HT, CRF, and TH. Recently, it has been proposed a hypothetical model that might explain how inhalation of volatile essential oils activates brain areas [33]. In brief, volatile molecules of essential oil may diffuse into the systemic circulation through the lung, which inhalation takes them to. Subsequently, they are transported to brain areas. In addition, the molecules activate the olfactory system connected to the limbic system via binding to olfactory receptors. Determination of the specific mechanisms involved in anti-depressant effects induced by EOAH will require additional study.
Conclusion
In conclusion, our results suggest that the reduced stress response produced by inhalation of EOAH is most likely mediated via an activation of serotonergic system and an inhibition of corticotropinergic and catecholaminergic system in the brain.
Acknowledgments
This work was supported by the Regional Innovation Center Program (Research Center for Biomedical Resources of Oriental Medicine at Daegu Haany University) of the Ministry of Trade, Industry, and Energy and a grant (14182MFDS979) from Ministry of Food and Drug Safety in 2015.
Abbreviations
- FST
Forced swimming test
- TST
Tail suspension test
- CRF
Corticotropin releasing factor
- 5-HT
Serotonin
- TH
Tyrosine hydroxylase
Footnotes
Hyun-Jung Park and Eun-Ju Lim contributed equally.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
HJP and EJL conducted the animal experiment and analyzed the data. CHY participated in design of the study and preparation of the manuscript. All the authors read and approved the final manuscript.
Contributor Information
Hyun-Jung Park, Email: lauren@khu.ac.kr.
Eun-Ju Lim, Email: ejlim@dhu.ac.kr.
Rong Jie Zhao, Email: zhao_rongjie@yahoo.com.
Sa Rang Oh, Email: sroh@dhu.ac.kr.
Ji Wook Jung, Email: jwjung@dhu.ac.kr.
Eun-Mi Ahn, Email: AhnEM@dhu.ac.kr.
Eun Sook Lee, Email: eslee@dhu.ac.kr.
Jin Suk Koo, Email: kimkoo1114@andong.ac.kr.
Hee Young Kim, Email: hykim@dhu.ac.kr.
Suchan Chang, Email: scchang@dhu.ac.kr.
Hyun Soo Shim, Email: alfm38@hanmail.net.
Kwang Joong Kim, Email: kwangj@dhu.ac.kr.
Young Seob Gwak, Email: ysgwak@dhu.ac.kr.
Chae Ha Yang, Email: chyang@dhu.ac.kr.
References
- 1.Karam EG, Fayyad J, Karam AN, Melhem N, Mneimneh Z, Dimassi H, et al. Outcome of depression and anxiety after war: a prospective epidemiologic study of children and adolescents. J Trauma Stress. 2014;27:192–9. doi: 10.1002/jts.21895. [DOI] [PubMed] [Google Scholar]
- 2.Saravanan C, Wilks R. Medical students’ experience of and reaction to stress: the role of depression and anxiety. ScientificWorldJournal. 2014;2014:737–382. doi: 10.1155/2014/737382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ong LK, Guan L, Damanhuri H, Goodchild AK, Bobrovskaya L, Dickson PW, et al. Neurobiological consequences of acute footshock stress: effects on tyrosine hydroxylase phosphorylation and activation in the rat brain and adrenal medulla. J Neurochem. 2014;128:547–60. doi: 10.1111/jnc.12482. [DOI] [PubMed] [Google Scholar]
- 4.Cryan JF, Valentino RJ, Lucki I. Assessing substrates underlying the behavioral effects of antidepressants using the modified rat forced swimming test. Neurosci Biobehav Rev. 2005;29:547–69. doi: 10.1016/j.neubiorev.2005.03.008. [DOI] [PubMed] [Google Scholar]
- 5.Breitenstein B, Scheuer S, Holsboer F. Are there meaningful biomarkers of treatment response for depression? Drug Discov Today. 2014;16:539–61. doi: 10.1016/j.drudis.2014.02.002. [DOI] [PubMed] [Google Scholar]
- 6.Komiya M, Takeuchi T, Harada E. Lemon oil vapor causes an anti-stress effect via modulating the 5-HT and DA activities in mice. Behav Brain Res. 2006;172:240–9. doi: 10.1016/j.bbr.2006.05.006. [DOI] [PubMed] [Google Scholar]
- 7.Hritcu L, Cioanca O, Hancianu M. Effects of lavender oil inhalation on improving scopolamine-induced spatial memory impairment in laboratory rats. Phytomedicine. 2012;19:529–34. doi: 10.1016/j.phymed.2012.02.002. [DOI] [PubMed] [Google Scholar]
- 8.Chen MC, Fang SH, Fang L: The effects of aromatherapy in relieving symptoms related to job stress among nurses. Int. J. Nurs. Pract., Nov 15. doi:10.1111 (2013). [DOI] [PubMed]
- 9.Drew AK, Whyte IM, Bensoussan A, Dawson AH, Zhu X, Myers SP. Chinese herbal medicine toxicology database: monograph on Herba Asari, “xi xin”. J Toxicol Clin Toxicol. 2002;40:169–72. doi: 10.1081/CLT-120004405. [DOI] [PubMed] [Google Scholar]
- 10.Zhu YP. Chinese Materia Medica; Chemistry, Pharmacology and Applications. Florida, USA: CRC Press; 1988. [Google Scholar]
- 11.Huang J, Wang HQ, Zhang C, Li GY, Lin RC, Wang JH. A new tetrahydrofuran-type lignan with anti-inflammatory activity from Asarum heterotropoides Fr. Schmidt var mandshuricum. J Asian Nat Prod Res. 2013;16:387–92. doi: 10.1080/10286020.2013.820713. [DOI] [PubMed] [Google Scholar]
- 12.Xu Y, Cao C, Shang M, Jiang Y, Wang X, Li C, et al. Assessment on anti-nociception and anti-inflammation pharmacodynamics of Asarum heterotropoides var. mandshuricum and Asarum sieboldii. Zhongguo Zhong Yao Za Zhi. 2012;37:625–31. [PubMed] [Google Scholar]
- 13.Hashimoto K, Yanagisawa T, Okui Y, Ikeya Y, Maruno M, Fujita T. Studies on anti-allergic components in the roots of Asiasarum sieboldi. Planta Med. 1994;60:124–7. doi: 10.1055/s-2006-959432. [DOI] [PubMed] [Google Scholar]
- 14.Deng Y, Feng Y, Sun J, Zhou D, Yang L, Lai J. [Study on anti-HPV activity of Asarum heterotropoides] Zhong Yao Cai. 2004;27:665–7. [PubMed] [Google Scholar]
- 15.Koo BS, Park KS, Ha JH, Park JH, Lim JC, Lee DU. Inhibitory effects of the fragrance inhalation of essential oil from Acorus graminens on central nervous system. Biol Pharm Bull. 2003;26:978–82. doi: 10.1248/bpb.26.978. [DOI] [PubMed] [Google Scholar]
- 16.Jeon S, Hur J, Jeong HJ, Koo BS, Pak SC. SuHeXiang Wan essential oil alleviates amyloid beta induced memory impairment through inhibition of tau protein phosphorylation in mice. Am J Chin Med. 2011;39:917–32. doi: 10.1142/S0192415X11009305. [DOI] [PubMed] [Google Scholar]
- 17.Castagne V, Moser P, Roux S, Porsolt RD: Rodent models of depression: forced swim and tail suspension behavioral despair tests in rats and mice. Curr. Protoc. Neurosci.,Chapter 8, Unit 8 10A (2011). [DOI] [PubMed]
- 18.Porsolt RD, Martin P, Lenegre A, Fromage S, Drieu K. Effects of an extract of Ginkgo Biloba (EGB 761) on “learned helplessness” and other models of stress in rodents. Pharmacol Biochem Behav. 1990;36:963–71. doi: 10.1016/0091-3057(90)90107-S. [DOI] [PubMed] [Google Scholar]
- 19.Steru L, Chermat R, Thierry B, Simon P. The tail suspension test: a new method for screening antidepressants in mice. Psychopharmacology (Berl) 1985;85:367–70. doi: 10.1007/BF00428203. [DOI] [PubMed] [Google Scholar]
- 20.Paxinos G, Watson C, Pennisi M, Topple A. Bregma, lambda and the interaural midpoint in stereotaxic surgery with rats of different sex, strain and weight. J Neurosci, methods. 1985;13:139–43. doi: 10.1016/0165-0270(85)90026-3. [DOI] [PubMed] [Google Scholar]
- 21.Doucet MV, Levine H, Dev KK, Harkin A. Small-molecule inhibitors at the PSD-95/nNOS interface have antidepressant-like properties in mice. Neuropsychopharmacology. 2013;38:1575–84. doi: 10.1038/npp.2013.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gibney SM, Fagan EM, Waldron AM, O’Byrne J, Connor TJ, Harkin A. Inhibition of stress-induced hepatic tryptophan 2,3-dioxygenase exhibits antidepressant activity in an animal model of depressive behaviour. Int J Neuropsychopharmacol. 2014;17:1–12. doi: 10.1017/S1461145713000746. [DOI] [PubMed] [Google Scholar]
- 23.Seredenin SB, Voronina TA, Gudasheva TA, Garibova TL, Molodavkin GM, Litvinova SA, et al. Antidepressant effect of dimeric dipeptide GSB-106, an original Low-molecular-weight mimetic of BDNF. Acta Naturae. 2013;5:105–9. [PMC free article] [PubMed] [Google Scholar]
- 24.Chatterjee M, Jaiswal M, Palit G. Comparative evaluation of forced swim test and tail suspension test as models of negative symptom of schizophrenia in rodents. ISRN Psychiatry. 2012;2012:595141. doi: 10.5402/2012/595141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Han P, Han T, Peng W, Wang XR. Antidepressant-like effects of essential oil and asarone, a major essential oil component from the rhizome of Acorus tatarinowii. Pharm Biol. 2013;51:589–94. doi: 10.3109/13880209.2012.751616. [DOI] [PubMed] [Google Scholar]
- 26.Socolsky C, Rates SM, Stein AC, Asakawa Y, Bardon A. Acylphloroglucinols from Elaphoglossum crassipes: antidepressant-like activity of crassipin A. J Nat Prod. 2012;75:1007–17. doi: 10.1021/np200436h. [DOI] [PubMed] [Google Scholar]
- 27.Zhang F, Wang LX, Luo Q, Xiao HB, Liang XM, Cai SQ. [Analysis of volatile constituents of root and rhizome of Asarum heterotropoides Fr var mandshuricum (Maxim) Kitag by gas chromatography–mass spectrometry] Se pu. 2002;20:467–70. [PubMed] [Google Scholar]
- 28.Norte MC, Cosentino RM, Lazarini CA. Effects of methyl-eugenol administration on behavioral models related to depression and anxiety, in rats. Phytomedicine. 2005;12:294–8. doi: 10.1016/j.phymed.2003.12.007. [DOI] [PubMed] [Google Scholar]
- 29.Gu L, Liu YJ, Wang YB, Yi LT. Role for monoaminergic systems in the antidepressant-like effect of ethanol extracts from Hemerocallis citrina. J Ethnopharmacol. 2012;139:780–7. doi: 10.1016/j.jep.2011.11.059. [DOI] [PubMed] [Google Scholar]
- 30.Schechter LE, Ring RH, Beyer CE, Hughes ZA, Khawaja X, Malberg JE, et al. Innovative approaches for the development of antidepressant drugs: current and future strategies. NeuroRx. 2005;2:590–611. doi: 10.1602/neurorx.2.4.590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chen HX, Jin ZL, Zhang LM, Xue R, Xu XD, Zhao N, et al. Antidepressant-like activity of YL-0919: a novel combined selective serotonin reuptake inhibitor and 5-HT1A receptor agonist. PLosOne. 2013;8:e83271. doi: 10.1371/journal.pone.0083271. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 32.Ducottet C, Griebel G, Belzung C. Effects of the selective nonpeptide corticotropin-releasing factor receptor 1 antagonist antalarmin in the chronic mild stress model of depression in mice. Prog Neuropsychopharmacol Biol Psychiatry. 2003;27:625–31. doi: 10.1016/S0278-5846(03)00051-4. [DOI] [PubMed] [Google Scholar]
- 33.Faturi CB, Leite JR, Alves PB, Canton AC, Teixeira-Silva F. Anxiolytic-like effect of sweet orange aroma in Wistar rats. Prog Neuropsychopharmacol Biol Psychiatry. 2010;34:605–9. doi: 10.1016/j.pnpbp.2010.02.020. [DOI] [PubMed] [Google Scholar]
- 34.Li B, Zhao J, Lv J, Tang F, Liu L, Sun Z, et al. Additive antidepressant-like effects of fasting with imipramine via modulation of 5-HT2 receptors in the mice. Prog Neuropsychopharmacol Biol Psychiatry. 2014;48:199–206. doi: 10.1016/j.pnpbp.2013.08.015. [DOI] [PubMed] [Google Scholar]
- 35.Griebel G, Simiand J, Steinberg R, Jung M, Gully D, Roger P, et al. 4-(2-chloro-4-methoxy-5-methylphenyl)-N-[(1S)-2-cyclopropyl-1-(3-fluoro-4-methylp henyl)ethyl]5-methyl-N-(2-propynyl)-1, 3-thiazol-2-amine hydrochloride (SSR125543A), a potent and selective corticotrophin-releasing factor(1) receptor antagonist. II. Characterization in rodent models of stress-related disorders. J Pharmacol Exp Ther. 2002;301:333–45. doi: 10.1124/jpet.301.1.333. [DOI] [PubMed] [Google Scholar]
- 36.Pan Y, Kong L, Xia X, Zhang W, Xia Z, Jiang F. Antidepressant-like effect of icariin and its possible mechanism in mice. Pharmacol Biochem Behav. 2005;82:686–94. doi: 10.1016/j.pbb.2005.11.010. [DOI] [PubMed] [Google Scholar]
- 37.Ku YC, Tsai YJ, Tung CS, Fang TH, Lo SM, Liu YP. Different involvement of ventral and dorsal norepinephrine pathways on norepinephrine reuptake inhibitor-induced locomotion and antidepressant-like effects in rats. Neurosci Lett. 2012;514:179–84. doi: 10.1016/j.neulet.2012.02.088. [DOI] [PubMed] [Google Scholar]
- 38.Kawabata K, Kawai Y, Terao J. Suppressive effect of quercetin on acute stress-induced hypothalamic-pituitary-adrenal axis response in Wistar rats. J Nutr Biochem. 2010;21:374–80. doi: 10.1016/j.jnutbio.2009.01.008. [DOI] [PubMed] [Google Scholar]
- 39.Park H, Shim H, Shim I, Lee T. Neural mechanisms underlying antidepressant-like effects of glycyrrhizae radix in rats. Korean J Oriental physiol Pathol. 2010;24:1053–8. [Google Scholar]
- 40.Bale TL, Vale WW. Increased depression-like behaviors in corticotropin-releasing factor receptor-2-deficient mice: sexually dichotomous responses. J Neurosci. 2003;23:5295–301. doi: 10.1523/JNEUROSCI.23-12-05295.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Maswood S, Barter JE, Watkins LR, Maier SF. Exposure to inescapable but not escapable shock increases extracellular levels of 5-HT in the dorsal raphe nucleus of the rat. Brain Res. 1998;783:115–20. doi: 10.1016/S0006-8993(97)01313-9. [DOI] [PubMed] [Google Scholar]
- 42.Kirby LG, Rice KC, Valentino RJ. Effects of corticotropin-releasing factor on neuronal activity in the serotonergic dorsal raphe nucleus. Neuropsychopharmacology. 2000;22:148–62. doi: 10.1016/S0893-133X(99)00093-7. [DOI] [PubMed] [Google Scholar]
- 43.Ramboz S, Oosting R, Amara DA, Kung HF, Blier P, Mendelsohn M, et al. Serotonin receptor 1A knockout: an animal model of anxiety-related disorder. Proc Natl Acad Sci U S A. 1998;95:14476–81. doi: 10.1073/pnas.95.24.14476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Perona MT, Waters S, Hall FS, Sora I, Lesch KP, Murphy DL, et al. Animal models of depression in dopamine, serotonin, and norepinephrine transporter knockout mice: prominent effects of dopamine transporter deletions. Behav Pharmacol. 2008;19:566–74. doi: 10.1097/FBP.0b013e32830cd80f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Shim ELT. Effects of mixture of lycii radicis cortex and moutan cortex on corticotropin-releasing factor, c-Fos, and tyrosine hydroxylase in forced swimming test. The society for herbal. 2011;26:59–66. [Google Scholar]
- 46.Ebner K, Singewald N. Stress-induced release of substance P in the locus coeruleus modulates cortical noradrenaline release. Naunyn Schmiedebergs Arch Pharmacol. 2007;376:73–82. doi: 10.1007/s00210-007-0185-3. [DOI] [PubMed] [Google Scholar]