

Abstract
This study aimed to evaluate the ability of a panel of lectins to inhibit the ability of Pasteurella multocida to adhere to and affect the rabbit respiratory epithelium. Nasal septa from rabbit fetuses were cultured with various lectins before the addition of P. multocida. The percentage of bacteria adhering to the epithelium was evaluated semiquantitatively by indirect immunoperoxidase (IIP) staining. The goblet cells (GCs) were counted in semithin sections stained with toluidine blue and served as the main morphological criterion to evaluate the inhibitory effect of the lectins. The lectins PNA, WGA, RCA120, and DBA significantly inhibited the adhesion of P. multocida to the ciliated epithelium (P < 0.05) and prevented the pathogen-induced increase in the number of GCs (P < 0.05) compared with those of positive control tissues. In addition, VVA, SJA, UEA I, DSL, SBA, and ECL significantly inhibited the increase in GCs compared with that of the control tissues. The results suggest that less aggressive therapeutic strategies, such as treatment with lectins, may represent alternative approaches to control bacterial respiratory infections.
1. Introduction
Adhesion of pathogenic microorganisms to epithelial surfaces is an important step in the infection and colonization of a susceptible host [1–4]. Infection by pathogens is generally initiated by the specific recognition of host epithelial surfaces. Receptors present in the mucin layer can act as binding sites during microbial adhesion. Lectin/glycoconjugate interactions are known for their high specificity and play a significant role in the adhesion of bacteria and other microorganisms to the epithelial surfaces of their hosts. In their infection strategy, bacteria often use sugar-binding proteins, such as lectins and the adhesins pili or fimbriae, to recognize and bind to host glycoconjugates [5–7]. In gram-negative bacteria, the lipopolysaccharide (LPS) on the outer membrane has also been reported as an important structure involved in carbohydrate-lectin interactions [8–10].
Bacterial resistance to antibiotics is an ever-increasing concern. An alternative disease intervention strategy is to target important steps in disease pathogenesis rather than targeting the pathogen directly; in this case, we would seek to mitigate disease by limiting pathogen attachment to host cells and thereby reduce colonization using substances that, in contrast to antibacterial agents, do not destroy the pathogens but rather interfere with their first pathogenic step, namely, their attachment to host cells. Accordingly, antiadhesion therapies have been documented for the enteropathogen E. coli K99 in swine and calves; adhesion and infection of this bacterium can be prevented using carbohydrates (CHOs) [11–13]. In the same manner, the specific sialic acid adhesion of Helicobacter pylori to human gastric mucus and erythrocytes was inhibited by high molecular mass constituents derived from cranberries [14].
Pasteurella multocida, which is considered a normal component of the upper respiratory tract flora in a variety of animal species, is a well-known pathogen responsible for a range of diseases and economic losses in bovines, swine, canines, laboratory animals, rabbits, and birds [15–19]. This pathogen has also been associated with respiratory tract infections in humans [20]. Although the pathogenic process is not completely understood, P. multocida can transition from being a normal inhabitant of the host to a pathogen capable of causing disease and death. Strategies to control P. multocida disease include vaccines and antibiotics, which can have limited efficacy [21, 22].
A major capsular component of all P. multocida serogroup A strains is hyaluronic acid [23, 24], the adherent properties of which have been described [25, 26]; additionally, the LPS of P. multocida A serovar 3 strain Pm70 possesses CHO sequences similar to those found on several host epithelial surfaces [27, 28]. These findings suggest that both structures might be susceptible targets for antiadhesive therapy by their corresponding lectins or CHOs.
In a search for alternative strategies to control P. multocida infections with fewer side effects, this ex vivo study explored the possibility of inhibiting P. multocida adhesion to the respiratory epithelium (nasal septum) of fetal rabbits using lectins. The results showed that lectin pretreatment reduced the number of bacteria adhering to the apical surface of the epithelium and blocked the pathogen-induced increase in the number of GCs in respiratory tissues, thus raising the possibility of an alternative control strategy for bacterial infection.
2. Materials and Methods
2.1. P. multocida Strains
P. multocida isolates from Pm147/08 to Pm160/08 were obtained from the turbinates, trachea, or lungs of diseased rabbits with rhinitis and bronchopneumonia [29]. Routine microbiology tests and PCR amplification and sequencing of the hyaD gene in the cap locus, which encodes proteins involved in the synthesis and assembly of the type A capsule (GenBank accession number AF067175) [30, 31], confirmed the identity of P. multocida type A.
P. multocida isolates were passed through mice by intraperitoneal inoculation. Mice were euthanized after the first signs of disease, and the bacterium was recovered from the heart, liver, lung, and trachea and cultured on BHI agar at 37°C for 24 h before use. The bacterial mass was collected and diluted in glucose-enriched essential medium (MEM), achieving a final concentration of 107 CFU/mL via counting and plating.
2.2. Lectins
A total of 18 distinct lectins from three commercial kits (Vector Laboratories) were used (Table 1). These lectins are extracted from various plants and included Con A, DBA, DSL, ECL, GSL I, GSL II, Jacalin, LCA, LEL, PNA, PSA, RCA120, SBA, SJA, STL, UEA I, VVA, and WGA. They were selected based on a range of specificities for multiple sugars [32–37].
Table 1.
Lectins and their characteristics [34, 35, 37]. Man, mannose; Glc, glucose; GalNAc, N-acetylgalactosamine; GlcNAc, N-acetylglucosamine; Gal, galactose; Fuc, fucose; Neu5Ac, N-acetylneuraminic acid.
| Source of lectin | |||
|---|---|---|---|
| Abbreviation | Latin name | Common name | Carbohydrate specificity |
| Con A | Canavalia ensiformis | Jack bean | Man/Glc |
| DBA | Dolichos biflorus | Horse gram | GalNAc(α1-3)/GalNAc |
| DSL | Datura stramonium | Jimson weed or thorn apple | (GlcNAc)2-4 |
| ECL | Erythrina cristagalli | Coral tree | Gal/GalNAc |
| GSL I | Griffonia simplicifolia I | Unknown | Gal/GalNAc |
| GSL II | Griffonia simplicifolia II | Unknown | GlcNAc |
| Jacalin | Artocarpus integrifolia | Jackfruit | Gal/GalNAc |
| LCA | Lens culinaris | Lentil | Man/Glc |
| LEL | Lycopersicon esculentum | Tomato | (GlcNAc)2-4 |
| PNA | Arachis hypogaea | Peanut | Gal β1-3/GalNAc |
| PSA | Pisum sativum | Garden pea | Man/Glc |
| RCA120 | Ricinus communis | Castor oil bean | Galβ1-4/GalNAc |
| SBA | Glycine max | Soybean | Gal/GalNAc |
| SJA | Sophora japonica | Japanese pagoda tree | Gal/GalNAc |
| STL | Solanum tuberosum | Potato | (GlcNAc)2-4 |
| UEA I | Ulex europaeus I | Gorse or furze | Fuc |
| VVA | Vicia villosa | Hair vetch | GalNAc |
| WGA | Triticum vulgaris | Wheat germ | GlcNAc/Neu5Ac |
2.3. Ex Vivo Culture of Rabbit Fetal Nasal Septa
This study was conducted with the approval of the Bioethics Committee of the Faculty of Veterinary Medicine of the National University of Colombia.
Eight pregnant female rabbits at their 26th gestational day were anesthetized with xylazine (5 mg kg−1) and ketamine (35 mg kg−1). The fetuses were delivered by caesarean and immediately euthanized by medullar sectioning; the females were also euthanized with an overdose of anesthetics immediately after the surgery. The skin, mandible, muscle, and palate of the fetuses were removed, and the nasal cavity was cross-sectioned with a sterile blade to obtain three slices, each 0.3 cm thick, from each animal. Sections were washed three times in MEM before tissue culture.
2.4. Inhibitory Effect of Natural Lectins on P. multocida Adherence to the Rabbit Respiratory Epithelium
To evaluate the potential inhibitory effects of lectins on P. multocida adhesion, an experiment was designed to block potential receptors for the bacterium on the apical membrane of epithelial cells. Six cross sections of nasal septa and bacteria were separately evaluated using each lectin (applied at 0.2 μg mL−1); six additional sections cultured without P. multocida or lectin and three sections incubated with each lectin alone were used as negative controls; six additional explants were incubated only with the bacterium and were used as positive controls. Tissue sections were immersed in 10 mL MEM supplemented with lectin in a 5 cm diameter Petri dish and incubated in a humid chamber with 5% CO2 and 95% O2 for 1 h. The samples were then washed three times with MEM to eliminate the nonadherent lectin. Next, 107 CFU of P. multocida was added to the samples, and they were incubated for 2 h. Three explants of each lectin treatment were fixed with 3.7% buffered formalin and three with Trump fixative (40% formalin, 25% glutaraldehyde) for 24 h. Control tissues were fixed in a similar manner [29, 38, 39].
2.5. Tissue Processing
2.5.1. Immunohistochemistry
A polyclonal antiserum raised in an adult female sheep was used. Briefly, as a first dose, 250 μg/mL of P. multocida was injected with complete Freund adjuvant (CFA); 7 days later, a booster dose with incomplete Freund adjuvant was applied. Finally, two doses of the antigen without adjuvant were inoculated one week apart. Animals were bled at 35 days p.i.; serum collected before inoculation served as a negative control. To eliminate cross-reactions, the serum was immunoadsorbed with normal nasal tissues of other fetuses. The serum was diluted 1 : 25 in sterile Tris-buffered saline (pH 7.6); 5 mL of the antiserum was then diluted in 1 mL of macerated sterile tissues previously washed with physiological salt solution. The mixture was centrifuged at 1000 rpm for 1 h at room temperature, and the supernatant was collected and frozen at −20°C until use. The working dilution of the primary antiserum was determined by indirect immunodot; a similar procedure was followed for the second antibody.
The number of bacteria adhered to the ciliated border of the respiratory epithelial cells was assessed by an indirect immunoperoxidase (IIP) technique [40, 41]. The tissues were embedded in paraffin and cut into 3 μm thick sections. The polyclonal antiserum raised in sheep was used as the specific primary antibody, and a commercial antiserum against ovine IgG produced in donkey was used as a secondary antibody (Sigma, Aldrich) [42]. Nasal septa of rabbits affected by the rhinitic and pneumonic forms of the disease were used as positive controls for the IIP technique.
The level of P. multocida adherence to the epithelium was assessed by a semiquantitative procedure as follows: both epithelial surfaces of the nasal septa were considered the full area (100%) to which the bacteria could adhere, and a mean adherence level was determined; no bacteria adhering to the surface were scored as 0%; bacteria attached to >0–30% of the epithelial surface were considered to be of focal adhesion; bacteria adhered to >30–60% of the surface were considered to be multifocal; and bacteria adhered to >60% of the surface were interpreted to be of diffuse adherence.
2.6. Semithin Sections
Tissue sections fixed for 24 h in Trump solution were decalcified in 10% EDTA for seven days, washed with 0.1 mmol phosphate-buffered saline (pH 7.3), postfixed in 1% osmium tetroxide, dehydrated in an ascending alcohol gradient, and finally embedded in Epon 812 (Polysciences). Sections (0.5 μm thick) were cut with a microtome (Microm) and stained with toluidine blue for approximately 30 s.
Tissue sections were evaluated by light microscopy using 100x objective. The number of cells (with cell nuclei used as the counting unit) in eight continuous fields of respiratory epithelium was analyzed in each of 3 replicates. The protective role of lectins was determined by calculating the percentage of GCs relative to those of other respiratory epithelial cells [29, 43]. In addition, increased GC activity was considered when cells showed an enlarged size, increased mucin release, and apical cytoplasm that protruded over their neighbor cells [44].
2.7. Statistical Analysis
The mean proportions of ciliated respiratory epithelium covered by adhered and nonadhered P. multocida were compared among the various lectin treatments and positive control tissues by ANOVA in a completely random model. To determine differences between treatments, a Dunnett test was performed, with P < 0.05 accepted as significant. To analyze the GC activity when the hypothesis was significant (P < 0.05), a Dunnett test was performed to compare each lectin treatment with the positive control [45].
3. Results
3.1. Inhibition of P. multocida Adhesion to the Respiratory Epithelium of Nasal Septa: IIP Staining
Immunostaining of P. multocida on the apical surface of the respiratory epithelium in lectin-treated tissues ranged from localized (>0–30%) (Figure 1(b)) to multifocal (>30–60%) (Figure 1(c)); by contrast, in positive control tissues (tissues exposed only to the bacterium), the immunostaining was similar in all replicates, with a generalized granular appearance covering almost 90% of the epithelial surface (Figure 1(d)).
Figure 1.

IIP technique for determining the percentage of the epithelial surface covered by adherent P. multocida. (a) Normal fetal rabbit respiratory nasal epithelium of IIP, negative control; light microscopy (LM), scale bar = 20 μm. (b) Respiratory epithelium of the nasal septa of rabbit fetus preincubated with PNA plus P. multocida (sparse focal immunostaining (arrows); >0–30% of the epithelial surface); LM, scale bar = 20 μm. (c) Tissue preincubated with VVA and P. multocida showing multifocal immunostaining (>30–60% of the surface covered with bacteria); LM, scale bar = 20 μm. (d) Tissue incubated with P. multocida without lectins (positive control) showing generalized immunostaining (>60% of the epithelial surface covered by bacteria); LM, scale bar = 20 μm.
The lectins LCA (Lens culinaris), PNA (Arachis hypogaea), WGA (Triticum vulgaris), RCA120 (Ricinus communis), and DBA (Dolichos biflorus) significantly (P < 0.05) inhibited P. multocida adherence to the ciliated border of respiratory epithelial cells compared with that of the positive control tissues (Figure 2).
Figure 2.
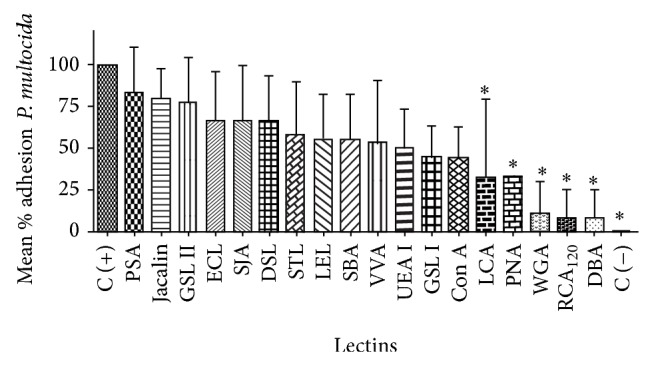
Inhibition of P. multocida adhesion to the ciliated respiratory epithelium of rabbit fetuses exposed to various lectins. Adhesion is expressed as mean percentage of the ciliated border epithelium coated by adhered bacteria. *Significant inhibition (P < 0.05) compared with the positive control tissues. C (−): negative control.
3.2. Semithin Sections
The respiratory epithelia of nasal septa exposed only to P. multocida showed an increase in the number of GCs compared with that of the control specimens (Figures 3(a) and 4). This increased number of GCs (Figure 3(c)) was accompanied by an enhanced production and release of mucus, such that the apical surface of these cells protruded above their neighboring ciliated cells, with their contents liberated into the lumen (Figure 3(c)). No increase in the number or activity of GCs was observed when nasal septa were incubated separately with each lectin.
Figure 3.

Toluidine blue staining of respiratory nasal epithelia of rabbit fetuses exposed to P. multocida. (a) Normal epithelium not exposed to bacteria or lectins; LM, scale bar = 20 μm. (b-c) Nasal septa of positive control tissues exposed only to P. multocida. (b) P. multocida adhering to cilia (arrow); LM, scale bar = 10 μm. (c) GCs showing increased numbers and excretory activity. All of the GCs are protruding above the apical limit of their neighboring cells and liberating their contents (arrows); LM, scale bar = 10 μm. (d) Nasal epithelium exposed to both bacteria and PNA. The lectin inhibited the increase in the number of GCs, though some loss of cilia can still be observed; LM, scale bar = 10 μm.
Figure 4.
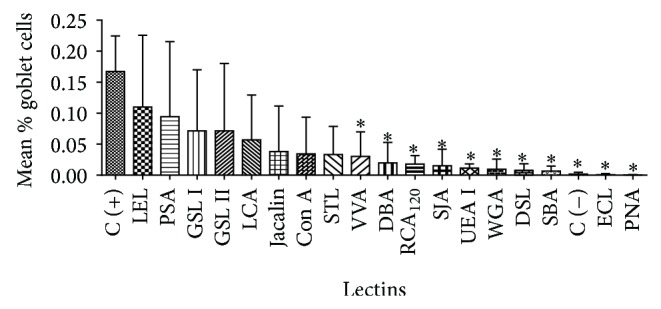
Inhibition of the increase in the percentage of GCs in the respiratory epithelia of nasal septa treated with various lectins. *Significant inhibition (P < 0.05) by lectins compared with positive control tissues. C (−): negative control.
The lectins VVA (Vicia villosa), DBA (Dolichos biflorus), RCA120 (Ricinus communis), SJA (Sophora japonica), UEA I (Ulex europaeus I), WGA (Triticum vulgaris), DSL (Datura stramonium), SBA (Glycine max), ECL (Erythrina cristagalli), and PNA (Arachis hypogaea) also showed significant inhibitions (P < 0.05) of the GC number and activity (Figure 4).
4. Discussion
The current strategies for preventing P. multocida infection in susceptible species, including rabbits, are limited to a few vaccines and antibiotics, but these approaches have shown inconsistent results [46–49]. In this study, we have explored a novel strategy to impede the adherence of this microorganism to the respiratory epithelia of rabbits using lectins while simultaneously investigating which molecules could protect the epithelium without having to kill the bacteria. Implementing an ex vivo experimental protocol, our data demonstrate that certain lectins inhibited the attachment of P. multocida to the respiratory epithelia of fetal rabbit nasal septa and also prevented the pathogen-induced increase in the number of GCs.
In this study, treatment with the lectins PNA (Gal β1–3/GalNAc), WGA (GlcNAc/Neu5Ac), RCA120 (Galβ1–4/GalNAc), DBA (GalNAc (α1–3)/GalNAc), and LCA (Man/Glc) significantly inhibited P. multocida adhesion to rabbit nasal respiratory epithelia and, in the cases of PNA, WGA, RCA120, and DBA also prevented an increase in the number and activity of GCs. These results suggest three possible explanations for the inhibitory activity of these lectins: (1) LCA, PNA, WGA, RCA120, and DBA bind to their specific CHO moieties present on the apical surface of the respiratory epithelium, thereby masking putative CHO receptors for corresponding lectins on the bacterial surface similar to those targeted in this work; (2) these lectins, on their side, recognize CHOs on the bacterial capsule and/or its LPS that serve as ligands for the corresponding lectin receptors on the apical membrane of the respiratory epithelium; and (3) both events occur simultaneously. Mason et al. [50] demonstrated that the PNA, LCA, RCA120, and WGA lectins recognize CHOs on the upper respiratory epithelium of laboratory rodents, partially confirming our results. In addition, Perfumo et al. [51] reported that lectins similar to those tested in this study (DBA, SBA, PNA, and RCA120) bind to receptors on the respiratory epithelial cells of the nasal cavity of healthy swine, as well as those suffering from atrophic rhinitis caused by P. multocida and B. bronchiseptica. Taken together, these findings indicate that CHOs and lectins with corresponding specificities are present on the apical surfaces of the respiratory epithelia of various animal species and most likely also on the surfaces of P. multocida and B. bronchiseptica, serving as receptors and ligands, respectively.
WGA recognizes GlcNAc (β1) and D-glucuronic acid residues, which are constituents of hyaluronan, a major nonsulfated glycosaminoglycan ubiquitous in all connective tissues; hyaluronan has also been described on both the apical and basal surfaces of the airway epithelium [52–55] and in tracheal secretions [56]. Hyaluronic acid is also a major capsular component of all P. multocida serogroup A strains [23, 24]. P. multocida serogroup A has been shown to adhere strongly to HeLa cells, turkey air sac macrophages, and alveolar macrophages; bacterial adhesion was reduced by treatment of the bacteria with hyaluronidase (to reduce the amount of capsule) or the addition of hyaluronic acid [25, 26]. Similarly, a spontaneous acapsular variant of P. multocida serogroup A did not adhere to turkey air sac macrophages [26]. Glorioso et al. [57] studied the adhesion of P. multocida isolated from rabbits to monolayers of HeLa cell cultures and to parakeratotic pharyngeal cells. Their most significant finding was that GlcNAc treatment inhibited bacterial adhesion to both cell lines. These results suggest that lectin-like molecules that serve as receptors for the microorganism are present on both epithelial surfaces. The main adherent role was attributed to P. multocida fimbriae [58]. These findings were complemented by those of Al-Haddawi et al. [59], who demonstrated that P. multocida A3 fimbriae isolated from rabbits recognize GlcNAc on the cilia of the respiratory epithelium; they were also able to competitively block this binding using the same CHO. Conversely, Jacques et al. [60] and Hatfaludi et al. [8] state that, despite the fact that P. multocida possesses various structures able to contribute to its binding of the respiratory epithelial cells of swine and rabbits in vitro, the LPS and type IV fimbria are likely the main mediators of the adhesion, acting as the CHO and lectin ligands, respectively. It is conceivable that, in our studies, WGA simultaneously bound hyaluronan residues on the apical surface of the respiratory epithelium and on the capsule of P. multocida, thereby exerting a double inhibitory effect.
However, the protective effect of WGA in this research could also be attributed to the inhibition of adherence of P. multocida LPS to hyaluronan on the apical membrane of the nasal respiratory epithelial cells. Protective activity against LPS-induced septic shock, acute lung injury, and airway hyperreactivity has been demonstrated by pretreatment with high molecular weight hyaluronan (<5 × 105), indicating that soluble hyaluronan can prevent the adherence of LPS to membrane-bound hyaluronan [61, 62].
The increase in the number and activity of GCs observed in this study could be due to the stimulation of the bacterium itself but is more likely related to the presence of substances liberated by P. multocida, mainly its LPS. Using the same ex vivo model as in this study, a previous study from our group exposed nasal cultures to P. multocida or its LPS separately; in both cases, the main response of the respiratory epithelium was a statistically significant increase in the number and activity of GCs compared with those of controls [29]. Previous studies have also reported hyperplasia and increased activity in the GCs of the respiratory epithelium following exposure to LPS [63–65]. Intranasal instillation of endotoxin in rats induces an inflammatory response characterized by an increase in the quantity or secretion of mucosubstances, as well as the infiltration of inflammatory cells into the epithelium, mainly PMNs [63]. Several mechanisms have been proposed to explain the positive effect of LPS on mucus production. Aside from the activity of various components of the PMNs, the expression of mucin genes, without the need for other secondary mediators, has been induced in cell cultures and nasal explants exposed only to endotoxin [66, 67]. Recent in vivo and in vitro work on the LPS of Pseudomonas aeruginosa reinforced these observations, as the expression of the mucin MUC5AC in mucin-producing cells of the airways was stimulated by treatment with this molecule [68]. Additional experiments by our group, in which the nasal septa of rabbit fetuses were only exposed to P. multocida, showed that the LPS was spontaneously released by the bacterium on the apical surface of the epithelial cells, where it entered the ciliated cells. This event was accompanied by increases in the number and activity of GCs, without any evidence of inflammatory cells [69]. These results reinforce the hypothesis that the LPS of P. multocida was one of the main targets of the inhibitory effect of the lectin WGA observed in this work.
Except for LCA, all lectins that prevented P. multocida adherence to the epithelium (PNA, WGA, RCA120, and DBA) also inhibited the increase in the number of GCs. It is not clear why LCA did not also have an inhibitory effect on these cells.
A different effect was found for VVA, SJA, UEA, DSL, SBA, and ECL, which significantly prevented the elevation in the number of GCs but did not significantly inhibit P. multocida adherence to the respiratory epithelium. Although we do not have a convincing explanation for the lack of inhibitory effect on P. multocida adhesion, part of the inhibitory effect of VVA, SJA, SBA, and ECL on GC number could be attributable to their specificity for CHOs residues containing Gal/GalNAc (and of DSL for GlcNAc), as these are important constituents of the LPS of P. multocida serovar 3, Pm70 [27].
Analysis of the P. multocida 3 Pm70 genome sequence identified two predicted filamentous hemagglutinin genes (fhaB1 and fhaB2) that might have similar adherent activities as that of the filamentous hemagglutinin protein of Bordetella pertussis, FhaB-FhaB1 FhaB2, for the adhesion to host cells [8, 70, 71]. This protein has at least three separate binding activities: a glycosaminoglycan binding site [72–74], an integrin-binding arginine-glycine-aspartate (RGD) sequence [75, 76], and a CHO recognition domain that mediates attachment to ciliated respiratory epithelial cells and macrophages [77–80]. The FhaB1 and FhaB2 proteins of P. multocida could have similar ligand affinity for CHO receptors on the airway epithelia of rabbits.
The results of this study indicate that P. multocida type A binds to the apical surface of rabbit nasal respiratory epithelia through several CHO-containing receptors, likely via bacterial surface structures, such as the capsule, LPS, type IV fimbriae, or lectin-like structures that may specifically bind to those glycosidic receptors, all of which would be potential targets for the inhibitory effect of the lectins observed in this work. We propose that, in addition to the potential use of lectins to inhibit the deleterious effects of P. multocida adhesion and colonization, it could also be more effective to employ the corresponding sugars, that is, D-Man, D-Glc, and GlcNAc, the effects of which have recently been demonstrated [81]. This approach would allow the key receptors on the respiratory epithelial cells to be occupied by the sugars or endocytosed upon sugar binding, reducing their numbers and leaving no binding sites for the microorganisms' sugar molecules [9, 13, 82]. The use of natural substances to inhibit pathogen adhesion to host cells might be an advantageous strategy that does not exert evolutionary pressure to generate more pathogenic forms of the microorganisms [6, 9].
5. Conclusions
A number of lectins were found to impede the attachment of P. multocida to the respiratory epithelia of fetal rabbit nasal septa ex vivo. The inhibition of P. multocida adhesion protected the respiratory cells from the lesions caused by the pathogen, and it is therefore highly probable that the undamaged mucosa would be able to clear the pathogen by innate protective mechanisms, such as mucociliary clearance. Preventing P. multocida colonization and cell damage in the upper respiratory tract may impede the bacteria from reaching deeper regions of the lungs. Our results support future investigations into whether the CHOs identified herein can also inhibit P. multocida adhesion in rabbits in vivo.
Acknowledgments
Special thanks are given to Dr. Noel Verjan from Tolima University, Colombia, and to Dr. Ben Adler from Monash University for their critical review of the paper. The authors are also thankful to Dr. Cristobal Corredor for his invaluable advice on lectins.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Reed W. P., Williams R. C., Jr. Bacterial adherence: first step in pathogenesis of certain infections. Journal of Chronic Diseases. 1978;31(2):67–72. doi: 10.1016/0021-9681(78)90091-7. [DOI] [PubMed] [Google Scholar]
- 2.Neiderman M. S. The pathogenesis of airway colonization: lessons learned from the study of bacterial adherence. European Respiratory Journal. 1994;7(10):1737–1740. doi: 10.1183/09031936.94.07101737. [DOI] [PubMed] [Google Scholar]
- 3.Lu L., Walker W. A. Pathologic and physiologic interactions of bacteria with the gastrointestinal epithelium. The American Journal of Clinical Nutrition. 2001;73(6):1124S–1130S. doi: 10.1093/ajcn/73.6.1124S. [DOI] [PubMed] [Google Scholar]
- 4.Foster T. J. Colonization and infection of the human host by staphylococci: adhesion, survival and immune evasion. Veterinary Dermatology. 2009;20(5-6):456–470. doi: 10.1111/j.1365-3164.2009.00825.x. [DOI] [PubMed] [Google Scholar]
- 5.Imberty A., Varrot A. Microbial recognition of human cell surface glycoconjugates. Current Opinion in Structural Biology. 2008;18(5):567–576. doi: 10.1016/j.sbi.2008.08.001. [DOI] [PubMed] [Google Scholar]
- 6.Hartmann M., Lindhorst T. K. The bacterial lectin FimH, a target for drug discovery—carbohydrate inhibitors of type 1 fimbriae-mediated bacterial adhesion. European Journal of Organic Chemistry. 2011;2011(20-21):3583–3609. doi: 10.1002/ejoc.201100407. [DOI] [Google Scholar]
- 7.Connell H., Agace W., Klemm P., Schembri M., Mårild S., Svanborg C. Type 1 fimbrial expression enhances Escherichia coli virulence for the urinary tract. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(18):9827–9832. doi: 10.1073/pnas.93.18.9827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hatfaludi T., Al-Hasani K., Boyce J. D., Adler B. Outer membrane proteins of Pasteurella multocida . Veterinary Microbiology. 2010;144(1-2):1–17. doi: 10.1016/j.vetmic.2010.01.027. [DOI] [PubMed] [Google Scholar]
- 9.Ofek I., Sharon N. Visions & reflections: a bright future for anti-adhesion therapy of infectious diseases. Cellular and Molecular Life Sciences. 2002;59(10):1666–1667. doi: 10.1007/PL00012494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bravo D., Hoare A., Silipo A., et al. Different sugar residues of the lipopolysaccharide outer core are required for early interactions of Salmonella enterica serovars Typhi and Typhimurium with epithelial cells. Microbial Pathogenesis. 2011;50(2):70–80. doi: 10.1016/j.micpath.2010.11.001. [DOI] [PubMed] [Google Scholar]
- 11.Mouricout M., Petit J. M., Carias J. R., Julien R. Glycoprotein glycans that inhibit adhesion of Escherichia coli mediated by K99 fimbriae: treatment of experimental colibacillosis. Infection and Immunity. 1990;58(1):98–106. doi: 10.1128/iai.58.1.98-106.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mouricout M. Swine and cattle enterotoxigenic Escherichia coli-mediated diarrhea. Development of therapies based on inhibition of bacteria-host interactions. European Journal of Epidemiology. 1991;7(6):588–604. doi: 10.1007/BF00218668. [DOI] [PubMed] [Google Scholar]
- 13.Sharon N. Atomic basis of protein-carbohydrate interactions: an overview. In: Bewley C., editor. Protein-Carbohydrate Interactions in Infectious Diseases. London, UK: Royal Society of Chemistry; 2006. [Google Scholar]
- 14.Burger O., Ofek I., Tabak M., Weiss E. I., Sharon N. A high molecular mass constituent of cranberry juice inhibits helicobacter pylori adhesion to human gastric mucus. FEMS Immunology Medical Microbiology. 2000;29(4):295–301. doi: 10.1111/j.1574-695X.2000.tb01537.x. [DOI] [PubMed] [Google Scholar]
- 15.Harper M., Boyce J. D., Adler B. Pasteurella multocida pathogenesis: 125 years after pasteur. FEMS Microbiology Letters. 2006;265(1):1–10. doi: 10.1111/j.1574-6968.2006.00442.x. [DOI] [PubMed] [Google Scholar]
- 16.García N. Caracterización fenotípica y genética de aislados de Pasteurella multocida obtenidos de ganado porcino [Ph.D. thesis] Madrid, Spain: Universidad Complutense de Madrid; 2010. [Google Scholar]
- 17.Hotchkiss E. J., Hodgson J. C., Schmitt-van de Leemput E., Dagleish M. P., Zadoks R. N. Molecular epidemiology of Pasteurella multocida in dairy and beef calves. Veterinary Microbiology. 2011;151(3-4):329–335. doi: 10.1016/j.vetmic.2011.03.018. [DOI] [PubMed] [Google Scholar]
- 18.Sthitmatee N., Kataoka Y., Sawada T. T. Molecular epidemiology of Japanese avian Pasteurella multocida strains by the single-enzyme amplified fragment length polymorphism and pulsed-field gel electrophoresis. The Journal of Veterinary Medical Science. 2010;72(11):1465–1470. doi: 10.1292/jvms.10-0181. [DOI] [PubMed] [Google Scholar]
- 19.Suelam I. I. A., Samie L. K. Molecular diversity of Pasteurella multocida isolated from different rabbit outbreaks at Zagazig suburbs, Egypt. Global Veterinaria. 2011;6(2):208–212. [Google Scholar]
- 20.Miyoshi S., Hamada H., Miyoshi A., et al. Pasteurella multocida pneumonia: zoonotic transmission confirmed by molecular epidemiological analysis. Geriatrics and Gerontology International. 2012;12(1):159–163. doi: 10.1111/j.1447-0594.2011.00721.x. [DOI] [PubMed] [Google Scholar]
- 21.Kehrenberg C., Thu Tham N. T., Schwarz S. New plasmid-borne antibiotic resistance gene cluster in Pasteurella multocida . Antimicrobial Agents and Chemotherapy. 2003;47(9):2978–2980. doi: 10.1128/aac.47.9.2978-2980.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kumarasamy K. K., Toleman M. A., Walsh T. R., et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. The Lancet Infectious Diseases. 2010;10(9):597–602. doi: 10.1016/s1473-3099(10)70143-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pandit K. K., Smith J. E. Capsular hyaluronic acid in Pasteurella multocida type A and its counterpart in type D. Research in Veterinary Science. 1993;54(1):20–24. doi: 10.1016/0034-5288(93)90005-z. [DOI] [PubMed] [Google Scholar]
- 24.Rosner H., Grimmecke H.-D., Knirel Y. A., Shashkov A. S. Hyaluronic acid and a (1→4]-β-D-xylan, extracellular polysaccharides of Pasteurella multocida (Carter type A) strain 880. Carbohydrate Research. 1992;223:329–333. doi: 10.1016/0008-6215(92)80032-v. [DOI] [PubMed] [Google Scholar]
- 25.Esslinger J., Seleim R. S., Hermann G., Blobel H. Adhesion of Pasteurella multocida to Hela cells and macrophages of different animal species. Revue de Medecine Veterinaire. 1994;145(1):49–53. [Google Scholar]
- 26.Pruimboom I. M., Rimler R. B., Ackermann M. R., Brogden K. A. Capsular hyaluronic acid-mediated adhesion of Pasteurella multocida to turkey air sac macrophages. Avian Diseases. 1996;40(4):887–893. doi: 10.2307/1592313. [DOI] [PubMed] [Google Scholar]
- 27.St. Michael F., Vinogradov E., Li J., Cox A. D. Structural analysis of the lipopolysaccharide from Pasteurella multocida genome strain Pm70 and identification of the putative lipopolysaccharide glycosyltransferases. Glycobiology. 2005;15(4):323–333. doi: 10.1093/glycob/cwi015. [DOI] [PubMed] [Google Scholar]
- 28.Houliston R. S., Bernatchez S., Karwaski M.-F., et al. Complete chemoenzymatic synthesis of the Forssman antigen using novel glycosyltransferases identified in Campylobacter jejuni and Pasteurella multocida . Glycobiology. 2009;19(2):153–159. doi: 10.1093/glycob/cwn117. [DOI] [PubMed] [Google Scholar]
- 29.Esquinas P. Interacción in vitro entre la Pasteurella multocida y/o su lipopolisacárido con epitelio de septo nasal de conejo [M.S. dissertation] Bogotá, Colombia: Universidad Nacional de Colombia; 2007. [Google Scholar]
- 30.Townsend K. M., Boyce J. D., Chung J. Y., Frost A. J., Adler B. Genetic organization of Pasteurella multocida cap loci and development of a multiplex capsular PCR typing system. Journal of Clinical Microbiology. 2001;39(3):924–929. doi: 10.1128/jcm.39.3.924-929.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Steen J. A., Harrison P., Seemann T., et al. Fis is essential for capsule production in Pasteurella multocida and regulates expression of other important virulence factors. PLoS Pathogens. 2010;6(2) doi: 10.1371/journal.ppat.1000750.e1000750 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Barbeito C., Massone A., Quiroga M. Aplicación de Las Técnicas de Lectinhistoquimica. Aplicaciones en Patología Veterinaria. La Plata, Argentina: Universidad Nacional de La Plata; 1990. [Google Scholar]
- 33.Slifkin M., Doyle R. J. Lectins and their application to clinical microbiology. Clinical Microbiology Reviews. 1990;3(3):197–218. doi: 10.1128/cmr.3.3.197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Brooks S. A., Schumacher U., Leathem A. J. C. Lectin Histochemistry. Oxford, UK: Royal Microscopical Society; 1997. Sources of lectins and other reagents for histochemistry. [Google Scholar]
- 35.Rhodes J., Milton J. Lectin-binding specifications. In: Rhodes J., Milton J., editors. Lectins Methods and Protocols. Totowa, NJ, USA: Humana Press; 1998. [Google Scholar]
- 36.Sharon N., Lis H. Plant lectins. In: Sharon N., Lis H., editors. Lectins. Dordrecht, The Netherlands: Springer; 2007. [Google Scholar]
- 37.Turton K., Natesh R., Thiyagarajan N., Chaddock J. A., Acharya K. R. Crystal structures of Erythrina cristagalli lectin with bound N-linked oligosaccharide and lactose. Glycobiology. 2004;14(10):923–929. doi: 10.1093/glycob/cwh114. [DOI] [PubMed] [Google Scholar]
- 38.Stinson M., Wang J. Lectin inhibition of bacterial adhesion to animal cells. Part XII: use of lectins in the investigation in pathogen-host interaction. In: Rhodes J., Milton J., editors. Lectins Methods and Protocols. New York, NY, USA: Humana Press; 1998. [Google Scholar]
- 39.Al-Haj Alia H., Sawadaa T., Hatakeyamab H., Katayamac Y., Ohtsukid N., Itohd O. Invasion of chicken embryo fibroblast cells by avian Pasteurella multocida . Veterinary Microbiology. 2004;104(1-2):55–62. doi: 10.1016/j.vetmic.2004.08.008. [DOI] [PubMed] [Google Scholar]
- 40.Heyderman E. Immunoperoxidase technique in histopathology: applications, methods, and controls. Journal of Clinical Pathology. 1979;32(10):971–978. doi: 10.1136/jcp.32.10.971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ogawa T., Gamoh K., Aoki H., et al. Validation and standardization of virus neutralizing test using indirect immunoperoxidase technique for the quantification of antibodies to rabies virus. Zoonoses and Public Health. 2008;55(6):323–327. doi: 10.1111/j.1863-2378.2008.01128.x. [DOI] [PubMed] [Google Scholar]
- 42.Kenny J. Immunochemical methods in cell and molecular biology: R. J. Mayer and J. H. Walker. pp 325. Academic Press, London. 1987. Biochemical Education. 1989;17(3):p. 164. doi: 10.1016/0307-4412(89)90121-0. [DOI] [Google Scholar]
- 43.Gallego C., Middleton A. M., Martínez N., Romero S., Iregui C. Interaction of Bordetella bronchiseptica and its lipopolysaccharide with in vitro culture of respiratory nasal epithelium. Veterinary Medicine International. 2013;2013:9. doi: 10.1155/2013/347086.347086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Esquinas P., Botero L., Patiño M. D. P., Gallego C., Iregui C. Ultrastructural comparison of the nasal epithelia of healthy and naturally affected rabbits with Pasteurella multocida A. Veterinary Medicine International. 2013;2013:8. doi: 10.1155/2013/321390.321390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Martínez R., Martínez N. Diseño de Experimentos Análisis de Datos Estándar y no Estándar. Bogotá, Colombia: Guadalupe ltda; 1997. [Google Scholar]
- 46.Rimler R. B., Rebers P. A., Rhoades K. R. Fowl cholera: cross-protection induced by Pasteurella multocida separated from infected turkey blood. Avian Diseases. 1979;23(3):730–741. doi: 10.2307/1589749. [DOI] [PubMed] [Google Scholar]
- 47.Suckow M. A., Bowersock T. L., Nielsen K., Chrisp C. E., Frandsen P. L., Janovitz E. B. Protective immunity to Pasteurella multocida heat-labile toxin by intranasal immunization in rabbits. Laboratory Animal Science. 1995;45(5):526–532. [PubMed] [Google Scholar]
- 48.Boyce J., Lo C., Wilkie I., Adler B. Pasteurella in Pathogenesis of Bacterial Infections in Animals. Malden, Mass, USA: Blackwell Publishing; 2010. (C. L. Gyles, J. F. Prescott, G. Songer, C. O. Thoen, Eds.). [Google Scholar]
- 49.Dowling A., Hodgson J. C., Dagleish M. P., Eckersall P. D., Sales J. Pathophysiological and immune cell responses in calves prior to and following lung challenge with formalin-killed Pasteurella multocida biotype A:3 and protection studies involving subsequent homologous live challenge. Veterinary Immunology and Immunopathology. 2004;100(3-4):197–207. doi: 10.1016/j.vetimm.2004.04.008. [DOI] [PubMed] [Google Scholar]
- 50.Mason C. M., Azizi S. Q., dal Nogare A. R. Respiratory epithelial carbohydrate levels of rats with gram-negative bacillary colonization. The Journal of Laboratory and Clinical Medicine. 1992;120(5):740–745. [PubMed] [Google Scholar]
- 51.Perfumo C. J., Mores N., Armocida A. D., Piffer I. A., Massone A. R., Itagaki S.-I. Histochemical and lectinhistochemical studies on nasal mucosa of pigs with or without respiratory diseases. Journal of Veterinary Medical Science. 1998;60(9):1021–1023. doi: 10.1292/jvms.60.1021. [DOI] [PubMed] [Google Scholar]
- 52.Green S. J., Tarone G., Underhill C. B. Distribution of hyaluronate and hyaluronate receptors in the adult lung. Journal of Cell Science. 1988;90(1):145–156. doi: 10.1242/jcs.90.1.145. [DOI] [PubMed] [Google Scholar]
- 53.Casalino-Matsuda S. M., Monzon M. E., Conner G. E., Salathe M., Forteza R. M. Role of hyaluronan and reactive oxygen species in tissue kallikrein-mediated epidermal growth factor receptor activation in human airways. The Journal of Biological Chemistry. 2004;279(20):21606–21616. doi: 10.1074/jbc.m309950200. [DOI] [PubMed] [Google Scholar]
- 54.Pirinen R. T., Tammi R. H., Tammi M. I., et al. Expression of hyaluronan in normal and dysplastic bronchial epithelium and in squamous cell carcinoma of the lung. International Journal of Cancer. 1998;79(3):251–255. doi: 10.1002/(sici)1097-0215(19980619)79:3x003C;251::aid-ijc7x0003e;3.0.co;2-o. [DOI] [PubMed] [Google Scholar]
- 55.Lauer M. E., Erzurum S. C., Mukhopadhyay D., et al. Differentiated murine airway epithelial cells synthesize a leukocyte-adhesive hyaluronan matrix in response to endoplasmic reticulum stress. Journal of Biological Chemistry. 2008;283(38):26283–26296. doi: 10.1074/jbc.M803350200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Monzon M. E., Casalino-Matsuda S. M., Forteza R. M. Identification of glycosaminoglycans in human airway secretions. American Journal of Respiratory Cell and Molecular Biology. 2006;34(2):135–141. doi: 10.1165/rcmb.2005-0256oc. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Glorioso J. C., Jones G. W., Rush H. G. Adhesion of type A Pasteurella multocida to rabbit pharyngeal cells and its possible role in rabbit respiratory tract infections. Infection and Immunity. 1982;35(3):1103–1109. doi: 10.1128/iai.35.3.1103-1109.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ruffolo C. G., Tennent J. M., Michalski W. P., Adler B. Identification,purification, and characterization of the type 4 fimbriae of Pasteurella multocida . Infection and Immunity. 1997;65(1):339–343. doi: 10.1128/iai.65.1.339-343.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Al-Haddawi M. H., Jasni S., Zamri-Saad M., et al. In vitro study of Pasteurella multocida adhesion to trachea, lung and aorta of rabbits. Veterinary Journal. 2000;159(3):274–281. doi: 10.1053/tvjl.1999.0418. [DOI] [PubMed] [Google Scholar]
- 60.Jacques M., Parent N., Foiry B. Adherence of Bordetella bronchiseptica and Pasteurella multocida to porcine nasal and tracheal epithelial cells. Canadian Journal of Veterinary Research. 1988;52(2):283–285. [PMC free article] [PubMed] [Google Scholar]
- 61.Muto J., Yamasaki K., Taylor K. R., Gallo R. L. Engagement of CD44 by hyaluronan suppresses TLR4 signaling and the septic response to LPS. Molecular Immunology. 2009;47(2-3):449–456. doi: 10.1016/j.molimm.2009.08.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Lennon F. E., Singleton P. A. Role of hyaluronan and hyaluronan-binding proteins in lung pathobiology. The American Journal of Physiology—Lung Cellular and Molecular Physiology. 2011;301(2):L137–L147. doi: 10.1152/ajplung.00071.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Harkema J. R., Hotchkiss J. A. In vivo effects of endotoxin on nasal epithelial mucosubstances: ouantitative histochemistry. Experimental Lung Research. 1991;17(4):743–761. doi: 10.3109/01902149109062876. [DOI] [PubMed] [Google Scholar]
- 64.Shimizu T., Takahashi Y., Kawaguchi S., Sakakura Y. Hypertrophic and metaplastic changes of goblet cells in rat nasal epithelium induced by endotoxin. American Journal of Respiratory and Critical Care Medicine. 1996;153(4):1412–1418. doi: 10.1164/ajrccm.153.4.8616574. [DOI] [PubMed] [Google Scholar]
- 65.Tamaoki J., Takeyama K., Yamawaki I., Kondo M., Konno K. Lipopolysaccharide-induced goblet cell hypersecretion in the guinea pig trachea: inhibition by macrolides. The American Journal of Physiology. 1997;272(1):L15–L19. doi: 10.1152/ajplung.1997.272.1.L15. [DOI] [PubMed] [Google Scholar]
- 66.Beckmann N., Tigani B., Sugar R., et al. Noninvasive detection of endotoxin-induced mucus hypersecretion in rat lung by MRI. The American Journal of Physiology—Lung Cellular and Molecular Physiology. 2002;283(1):L22–L30. doi: 10.1152/ajplung.00373.2001. [DOI] [PubMed] [Google Scholar]
- 67.Wagner J. G., van Dyken S. J., Wierenga J. R., Hotchkiss J. A., Harkema J. R. Ozone exposure enhances endotoxin-induced mucous cell metaplasia in rat pulmonary airways. Toxicological Sciences. 2003;74(2):437–446. doi: 10.1093/toxsci/kfg120. [DOI] [PubMed] [Google Scholar]
- 68.Li W., Yan F., Zhou H., et al. P. aeruginosa lipopolysaccharide-induced MUC5AC and CLCA3 expression is partly through Duox1 in vitro and in vivo . PLoS ONE. 2013;8(5) doi: 10.1371/journal.pone.0063945.e63945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Gallego C., Romero S., Esquinas P., Iregui C. Proposed pathogenic roles of the lipopolysaccharide of Pasteurella multocida A during the first steps of the infection in rabbits. Proceedings of the International Pasteurellaceae Conference; 2014; Prato, Italy. [Google Scholar]
- 70.Locht C., Antoine R., Jacob-Dubuisson F. Bordetella pertussis, molecular pathogenesis under multiple aspects. Current Opinion in Microbiology. 2001;4(1):82–89. doi: 10.1016/s1369-5274(00)00169-7. [DOI] [PubMed] [Google Scholar]
- 71.May B. J., Zhang Q., Li L. L., Paustian M. L., Whittam T. S., Kapur V. Complete genomic sequence of Pasteurella multocida, Pm70. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(6):3460–3465. doi: 10.1073/pnas.051634598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Hannah J. H., Menozzi F. D., Renauld G., Locht C., Brennan M. J. Sulfated glycoconjugate receptors for the Bordetella pertussis adhesin filamentous hemagglutinin (FHA) and mapping of the heparin-binding domain on FHA. Infection and Immunity. 1994;62(11):5010–5019. doi: 10.1128/iai.62.11.5010-5019.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Menozzi F. D., Mutombo R., Renauld G., et al. Heparin-inhibitable lectin activity of the filamentous hemagglutinin adhesin of Bordetella pertussis . Infection and Immunity. 1994;62(3):769–778. doi: 10.1128/iai.62.3.769-778.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Mattoo S., Cherry J. D. Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clinical Microbiology Reviews. 2005;18(2):326–382. doi: 10.1128/cmr.18.2.326-382.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Relman D. A., Domenighini M., Toumanen E., Rappuoli R., Falkow S. Filamentous hemagglutinin of Bordetella pertussis: nucleotide sequence and crucial role in adherence. Proceedings of the National Academy of Sciences of the United States of America. 1989;86(8):2637–2641. doi: 10.1073/pnas.86.8.2637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Relman D. A., Tuomanen E. T., Falkow S., Golenbock D. T., Saukkonen K., Wright S. D. Recognition of a bacterial adhesin by an integrin: macrophage CR3 (αMβ2, CD11b/CD18) binds filamentous hemagglutinin of Bordetella pertussis . Cell. 1990;61(7):1375–1382. doi: 10.1016/0092-8674(90)90701-f. [DOI] [PubMed] [Google Scholar]
- 77.Tuomanen E., Towbin H., Rosenfelder G., et al. Receptor analogs and monoclonal antibodies that inhibit adherence of Bordetella pertussis to human ciliated respiratory epithelial cells. The Journal of Experimental Medicine. 1988;168(1):267–277. doi: 10.1084/jem.168.1.267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Saukkonen K., Cabellos C., Burroughs M., Prasad S., Tuomanen E. Integrin-mediated localization of Bordetella pertussis within macrophages: role in pulmonary colonization. Journal of Experimental Medicine. 1991;173(5):1143–1149. doi: 10.1084/jem.173.5.1143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Prasad S. M., Yin Y., Rodzinski E., Tuomanen E. I., Masure H. R. Identification of a carbohydrate recognition domain in filamentous hemagglutinin from Bordetella pertussis . Infection and Immunity. 1993;61(7):2780–2785. doi: 10.1128/iai.61.7.2780-2785.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Cotter P. A., Yuk M. H., Mattoo S., et al. Filamentous hemagglutinin of Bordetella bronchiseptica is required for efficient establishment of tracheal colonization. Infection and Immunity. 1998;66(12):5921–5929. doi: 10.1128/iai.66.12.5921-5929.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Gallego C., Patiño M. P., Martínez N., Iregui C. A. Prevention of Disease and Lesions Caused by P. multocida in Rabbits by Using Carbohydrates. Barcelona, Spain: European Society of Clinical Microbiology and Infectious Diseases; 2014. [Google Scholar]
- 82.Sharon N. Lectins: carbohydrate-specific reagents and biological recognition molecules. The Journal of Biological Chemistry. 2007;282(5):2753–2764. doi: 10.1074/jbc.x600004200. [DOI] [PubMed] [Google Scholar]