

SUMMARY
Histone lysine methylation is a critical regulator of chromatin-templated processes such as gene transcription and DNA repair, and is dynamically controlled by enzymes that write and erase this post-translational modification (PTM). While histone methylation has been well studied, the functions of non-histone lysine methylation and its regulatory enzymes, particularly outside the nucleus, are poorly defined. In this issue of Cancer Discovery, Whetstine and colleagues shed light on a new role for the lysine demethylase KDM4A as a regulator of protein translation and identify a single nucleotide polymorphism (SNP) in the KDM4A gene as a candidate biomarker for mTOR inhibitor therapy.
While it has been more than a half century since the discovery of lysine methylation, insight into the function(s) of this post-translational modification (PTM) were fueled by seminal discoveries that linked site-specific methylation on histones to gene transcription (1). Since then, much focus has been placed on the role of histone lysine methylation as a regulator of chromatin structure and function in human health and disease (2), including the discovery of at least 50 predicted lysine methyltransferase enzymes (KMTs) (3). Until recently, lysine methylation was considered an irreversible PTM. It is now appreciated that two classes of enzymes, consisting of more than 30 predicted members, function as lysine demethylases (KDMs) (4). Among them is KDM4A/JMJD2a, a member of the α-ketoglutarate and Fe(II)-dependent dioxygenases known as JMJC demethylases. KDM4A has three known substrate lysines, all on histones (5, 6), and has identified functions as a regulator of gene expression, DNA damage signaling, DNA replication, and site-specific copy number regulation (7). Moreover, KDM4A itself is copy gained and lost in various cancers and protein expression correlates positively with proliferation, metastasis and poor prognosis in cancers of the bladder and lung.
In this issue of Cancer Discovery, Whetstine and colleagues publish two manuscripts defining an unexpected and exciting role for KDM4A outside the nucleus as a regulator of protein translation (8, 9). Noting that KDM4A protein levels were detectable in cytoplasmic biochemical fractionations from retinal pigment epithelium cells, Whetstine and colleagues identified by mass spectrometry, co-immunoprecipitation, and polysome fractionation the association of KDM4A with members of the translation initiation machinery. They further showed that KDM4A depletion by shRNA changed the distribution of translation factors in polysome fractions, suggesting that proper complex assembly/disassembly is dependent on KDM4A. KDM4A depletion was then shown to result in decreased global protein synthesis, an effect also seen with rapamycin – the pioneering inhibitor of the mTOR signaling pathway. mTOR regulates protein synthesis in part by phosphorylating and releasing the negative regulator of translation 4E-BP1 from eIE4E, an initiation factor that binds the 7-methyl-guanosine cap on mature mRNAs, and by phosphorylating and activating S6 kinase, which has numerous downstream translation regulatory targets (Figure 1). Notably, protein synthesis inhibition and sensitivity to several mTOR inhibitors was potentiated by KDM4A depletion and small molecule inhibition in HEK 293T cells, suggesting a rationale combination therapeutic approach.
Figure 1. Compartmentalized cellular functions of KDM4A.
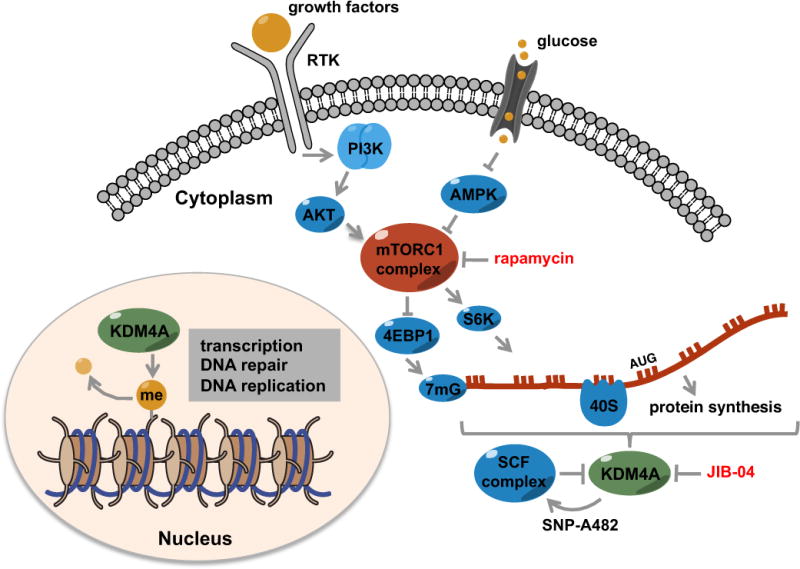
Cartoon depicting the known nuclear and newly uncovered cytoplasmic roles of KDM4A. Shown are receptor tyrosine kinase (RTK) and glucose signaling axes that elicit control on mTORC1, a key regulator of protein synthesis. Whetstine and colleagues show that KDM4A regulates protein synthesis in the cytoplasm through its association with translation initiation factors. They further show that an identified SNP-482 in KDM4A promotes its turnover by targeting it to the SCF ubiquitin ligase complex. Loss of KDM4A or inhibition of catalytic activity with a small molecule (JIB-04) enhances therapeutic effects of rapamycin and other mTORC1 inhibitors.
Whetstine and colleagues also showed with a drug screen (87 preclinical and clinical compounds) against 86 genotyped lung cancer cell lines that cells homozygous for a coding single nucleotide polymorphism (SNP) in KDM4A (SNP-A482) were hypersensitive to mTOR inhibitors. Biochemical analysis of SNP-A482 revealed decreased stability in KDM4A, which was attributed to polyubiquitination by the SCF ubiquitin ligase complex that resulted in increased protein turnover. These data suggest SNP-A482 may be a biomarker for mTOR sensitivity, and indeed, Whetstine and colleagues showed that SNP-A482 is associated with poor prognosis in non-small cell lung cancer patients. Collectively, these data redefine the role of KDM4A and highlight a new function for this demethylase outside the nucleus as a regulator of protein translation. By demonstrating that KDM4A inhibition can be achieved by a small molecule, and with the realization that a coding SNP potentiates the therapeutic effects of mTOR inhibitors, these findings reveal a new biomarker and possible future therapeutic approaches in cancers.
Moving forward, a number of exciting questions remain to be answered. First, what are the translation-associated sites of lysine methylation that KDM4A demethylates, and how do they impact translation? Indeed, the presence of methyl-lysine on the translation machinery, including the ribosome and elongation factors, has been known for several decades, and recent mass spectrometry-based proteomics analyses have revealed a number of newly discovered lysine methylation sites on translation components and beyond (10). However, how lysine methylation impacts translation itself is poorly understood. It will be exciting to resolve which ribosomal subunits are methylated, and how these methylation events (both their establishment and removal) contributes to the proper timing and promotion of translation. It may be that removal of lysine methylation on the ribosome removes an inhibitory effector protein that regulates the ribosome – most likely a factor connected to the mTOR pathway. Alternatively, a site of lysine methylation could be directly impacting translation itself, and removal of this methylation event may increase the rate of translation by improving some aspect of ribosome function. Finally, it may be that KDM4A, while associated with ribosomes, has another target that itself influences translation.
Another unanswered question is what enzyme mediates the methylation that KDM4A removes? Significant to point out, there are many sites of lysine methylation that have been identified outside the nucleus (10), in addition to a wide number of lysine methyltransferases that could mediate them (3). This also raises the question: do other chromatin writers and erasers also contribute to ribosome regulation? The work by Whetstine and colleagues underscores the need to identify enzymes regulating these PTMs. Careful analysis of the subcellular localization of lysine methyltransferases and demethylases will provide fundamental insights needed to begin addressing this important question.
How is KDM4A recruited to polysomes? KDM4A is targeted to chromatin by its tandem Tudor domain, a specialized protein fold that recognizes trimethyl-lysine in a sequence-specific manner. It is attractive to speculate that, like histones, KDM4A uses its tandem Tudor domain to regulate its translation complex association by engaging sites of lysine methylation. It is also interesting to note that Whetstine and colleagues show that the catalytic dead form of KDM4A constitutively associates with translation components in polysome fractionations. This suggests a negative feedback model of complex association such that KDM4A activity may release the demethylase from its binding partners in the translation complex. It will also be of interest to determine mechanisms controlling the subcellular localization of KDM4A and design mutants or fusions of KDM4A that restrict this demethylase to the cytoplasm or nucleus, particularly since it is now unclear whether the therapeutic benefit seen from small molecule inhibitors of KDM4A like JIB-04 is a result of inhibiting gene regulatory functions of KDM4A, its effects on translation, or most likely both.
Is cytoplasmic KDM4A regulated by stress and/or nutrient signaling? The relationship between KDM4A and signals integrating on the mTOR pathway will be an important area of future study, particularly if we are to consider targeting KDM4A in combination with inhibitors of these deregulated signaling axes in cancers. It will be exciting to determine both how cytoplasmic KDM4A responds to growth factors and nutrients like glucose (Figure 1) and how pharmacological interventions at nodal points along these signaling axes regulate KDM4A function outside the nucleus.
Is the therapeutic effect of KDM4A inhibition mediated through the inhibition of its nuclear role(s) and/or its cytoplasmic association with the ribosome? These studies underscore the necessity for careful biochemical analysis of chromatin regulatory factors and their mutations, particularly since many epigenetic factors are now being considered as next-generation targets for cancer therapy. Key to the future success of epigenetic drug discovery programs, and for the rationale design of therapeutic combinations, will be comprehensive biochemical characterization of the protein-protein interaction networks and function(s) these epigenetic regulators have, combined with well-designed measures to monitor target inhibition in cells.
Whetstine and colleagues nicely reinforce the concept that non-histone lysine methylation has important biological and clinical ramifications. Along these lines, it will be important to determine the extent to which other cytoplasmic proteins are targeted for methylation/demethylation events by previously presumed nuclear enzymes. Studies defining the “lysine methylome” have unveiled over 1400 methylated proteins to date (10) – almost half of which are annotated by Uniprot to be in both the nucleus and cytoplasm. Similarly, about half of the presumed histone lysine methyltransferases (3) and demethylases (4) are annotated in both the nucleus and the cytoplasm. We predict the observations made by Whetstine and colleagues with KDM4A are likely to be just the tip of the iceberg, and that many future examples of cytoplasmic regulation by histone-modifying enzymes will be made in the future.
Acknowledgments
The authors acknowledge financial support from the National Institutes of Health (NIH) to S.B.R. (K99 CA181343) and B.D.S. (R01 GM110058). B.D.S. also acknowledges financial support from the W.M. Keck Foundation and the National Science Foundation. B.M.D. acknowledges support from the NIH (R01 GM100919), the Carolina Partnership and the University Cancer Research Fund, UNC Chapel Hill. We apologize to authors whose works we could not cite due to space limitations.
Footnotes
CONFLICT OF INTEREST
The authors disclose no potential conflicts of interest.
References
- 1.Lee DY, Teyssier C, Strahl BD, Stallcup MR. Role of protein methylation in regulation of transcription. Endocrine reviews. 2005;26:147–70. doi: 10.1210/er.2004-0008. [DOI] [PubMed] [Google Scholar]
- 2.Greer EL, Shi Y. Histone methylation: a dynamic mark in health, disease and inheritance. Nature reviews Genetics. 2012;13:343–57. doi: 10.1038/nrg3173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Petrossian TC, Clarke SG. Uncovering the human methyltransferasome. Molecular & cellular proteomics: MCP. 2011;10:M110 000976. doi: 10.1074/mcp.M110.000976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kooistra SM, Helin K. Molecular mechanisms and potential functions of histone demethylases. Nature reviews Molecular cell biology. 2012;13:297–311. doi: 10.1038/nrm3327. [DOI] [PubMed] [Google Scholar]
- 5.Klose RJ, Yamane K, Bae Y, Zhang D, Erdjument-Bromage H, Tempst P, et al. The transcriptional repressor JHDM3A demethylates trimethyl histone H3 lysine 9 and lysine 36. Nature. 2006;442:312–6. doi: 10.1038/nature04853. [DOI] [PubMed] [Google Scholar]
- 6.Whetstine JR, Nottke A, Lan F, Huarte M, Smolikov S, Chen Z, et al. Reversal of histone lysine trimethylation by the JMJD2 family of histone demethylases. Cell. 2006;125:467–81. doi: 10.1016/j.cell.2006.03.028. [DOI] [PubMed] [Google Scholar]
- 7.Black JC, Manning AL, Van Rechem C, Kim J, Ladd B, Cho J, et al. KDM4A lysine demethylase induces site-specific copy gain and rereplication of regions amplified in tumors. Cell. 2013;154:541–55. doi: 10.1016/j.cell.2013.06.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Van Rechem C, Black JC, Boukhali M, Aryee MJ, Graslund S, Haas W, et al. Lysine Demethylase KDM4A Associates with Translation Machinery and Regulates Protein Synthesis. Cancer discovery. 2015 doi: 10.1158/2159-8290.CD-14-1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Van Rechem C, Black JC, Greninger P, Zhao Y, Donado C, Burrowes PD, et al. A Coding Single Nucleotide Polymorphism in Lysine Demethylase KDM4A Associates with Increased Sensitivity to mTOR Inhibitors. Cancer discovery. 2015 doi: 10.1158/2159-8290.CD-14-1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Moore KE, Gozani O. An unexpected journey: Lysine methylation across the proteome. Biochimica et biophysica acta. 2014;1839:1395–403. doi: 10.1016/j.bbagrm.2014.02.008. [DOI] [PMC free article] [PubMed] [Google Scholar]