

Abstract
Background
Pseudomonas aeruginosa is an opportunistic pathogen that exploits damaged epithelia to cause infection. Type IV pili (tfp) are polarly located filamentous structures which are the major adhesins for attachment of P. aeruginosa to epithelial cells. The extension and retraction of tfp powers a mode of surface translocation termed twitching motility that is involved in biofilm development and also mediates the active expansion of biofilms across surfaces. Extracellular adenosine triphosphate (eATP) is a key “danger” signalling molecule that is released by damaged epithelial cells to alert the immune system to the potential presence of pathogens. As P. aeruginosa has a propensity for infecting damaged epithelial tissues we have explored the influence of eATP on tfp biogenesis and twitching motility-mediated biofilm expansion by P. aeruginosa.
Results
In this study we have found that eATP inhibits P. aeruginosa twitching motility-mediated expansion of interstitial biofilms at levels that are not inhibitory to growth. We have determined that eATP does not inhibit expression of the tfp major subunit, PilA, but reduces the levels of surface assembled tfp. We have also determined that the active twitching zone of expanding P. aeruginosa interstitial biofilms contain large quantities of eATP which may serve as a signalling molecule to co-ordinate cell movements in the expanding biofilm. The inhibition of twitching motility-mediated interstitial biofilm expansion requires eATP hydrolysis and does not appear to be mediated by the Chp chemosensory system.
Conclusions
Endogenous eATP produced by P. aeruginosa serves as a signalling molecule to co-ordinate complex multicellular behaviours of this pathogen. Given the propensity for P. aeruginosa to infect damaged epithelial tissues, our observations suggest that eATP released by damaged cells may provide a cue to reduce twitching motility of P. aeruginosa in order to establish infection at the site of damage. Furthermore, eATP produced by P. aeruginosa biofilms and by damaged epithelial cells may play a role in P. aeruginosa pathogenesis by inducing inflammatory damage and fibrosis. Our findings have significant implications in the development and pathogenesis of P. aeruginosa biofilm infections.
Electronic supplementary material
The online version of this article (doi:10.1186/s12866-015-0392-x) contains supplementary material, which is available to authorized users.
Keywords: 3’,5’-adenosine triphosphate; ATP; Twitching motility; Type IV pili; Tfp; t4p; eATP
Background
Pseudomonas aeruginosa is a Gram negative bacterium found throughout the environment and is an opportunistic pathogen of a wide variety of eukaryotic hosts [1,2]. P. aeruginosa causes acute and chronic infections in immunocompromised humans and is the major contributor to the morbidity and mortality of individuals with cystic fibrosis [3,4]. P. aeruginosa establishes infections at sites of epithelial tissue damage such as those encountered in patients with severe burns, mechanical ventilation, or corneal damage due to contact lens use [5,6]. P. aeruginosa can establish chronic infections which are associated with the formation of complex, surface-associated communities termed biofilms [7].
Type IV pili (tfp) are the major adhesins of P. aeruginosa that promote attachment to host epithelial cells. These polar, filamentous surface structures also mediate a form of surface translocation termed twitching motility that occurs through the extension, surface binding and retraction of tfp [8]. The active expansion of P. aeruginosa biofilms is a complex, multicellular, collective behaviour that is mediated by twitching motility [9,10]. A number of host derived signals including mucin, serum albumin, oligopeptides, phosphatidylcholine (PC), lactoferrin and low levels of available iron have been shown to stimulate twitching motility-mediated biofilm expansion by P. aeruginosa [11-15]. Interestingly, mucin, serum albumin, lactoferrin, and low iron have also been shown to inhibit the formation of sessile biofilms that form under fully hydrated conditions [12-15]. It has been suggested that the effects of these host-derived compounds may provide a protective advantage to the host by inhibiting the ability of P. aeruginosa to form resistant, sessile biofilms, and thus giving the immune system a better chance of clearing the infection [14].
Another host-derived signalling molecule, extracellular 3’,5’-adenosine triphosphate (eATP) is rapidly released in high levels (5–10 mM) by stressed or injured epithelial cells to alert the host to the presence of invading pathogens, resulting in recruitment of immune system factors to clear the infection [16,17]. As P. aeruginosa displays a propensity for infecting damaged epithelial tissues [5] it is therefore likely to encounter high levels of eATP at the site of damaged epithelium where it initiates infection. In this study we have explored the possibility that eATP may be a host-derived molecule that affects twitching motility in P. aeruginosa and have found that eATP inhibits P. aeruginosa twitching motility-mediated expansion of interstitial biofilms at levels that are not inhibitory to growth.
Results
Extracellular ATP inhibits twitching motility
To determine if eATP influences twitching motility-mediated expansion of interstitial biofilms by P. aeruginosa, we utilised our interstitial twitching motility assay [18] to examine the effect of incorporating eATP into the solidified nutrient media. In this assay P. aeruginosa cells are inoculated at the interface between a glass coverslip and a microscope slide coated in nutrient media that has been solidified with gellan gum. Under these conditions, twitching motility mediates rapid, active biofilm expansion at the interface between the glass coverslip and solidified nutrient media [9,10]. The resultant biofilm is imaged using phase-contrast microscopy and quantitated using image analysis tools. To determine if eATP affects P. aeruginosa PAK twitching motility-mediated expansion of interstitial biofilms, varying concentrations of eATP (0 mM–15 mM) were incorporated into the nutrient media. After 16 h incubation at 37°C the resulting biofilms were imaged (Figure 1A) and the surface area covered by the biofilms calculated (Figure 1B). These analyses revealed that eATP concentrations of 5 mM and above dramatically reduced twitching motility-mediated biofilm expansion to levels equivalent to that of the non-twitching tfp pilin mutant PAKpilA::TcR (Figure 1B) [9,19]. Furthermore, microscopic examination of the interstitial biofilms (Figure 1A) shows that at eATP concentrations of 7.5 mM and above, the resultant biofilms resemble those of the non-twitching PAKpilA::TcR strain as they no longer form the leading-edge rafts and lattice-like trail network which are characteristic micromorphological features of wild-type P. aeruginosa interstitial biofilms in this assay [9,10] (Figure 1A). Interestingly in the presence of 2.5 mM and 5 mM eATP the interstitial biofilms had very large leading edge rafts and had lost the ability to self-organise into the characteristic refined trail network (Figure 1A).
Figure 1.
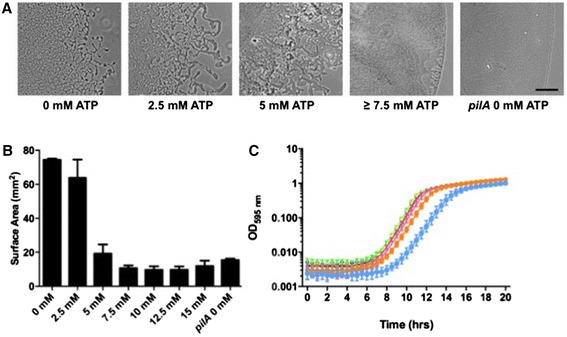
Extracellular ATP inhibits twitching motility-mediated expansion of P. aeruginosa interstitial biofilms. (A) Representative light microscopy images of P. aeruginosa PAK and PAKpilA interstitial biofilms formed at the interface between a microscope slide coated in solidified nutrient media supplemented with a range of eATP concentrations and a coverslip after incubation at 37°C for 16 h. Scale 50 μm. (B) Surface areas of the interstitial biofilms formed by P. aeruginosa PAK and PAKpilA at the interface between a microscope slide coated in solidified nutrient media supplemented with a range of eATP concentrations and a coverslip after incubation at 37°C for 16 h. Mean ± SD from three independent experiments run in duplicate are presented. (C) Planktonic growth of P. aeruginosa PAK at 37°C over 20 h in media supplemented with 0 mM ATP (black), 2.5 mM ATP (green), 5 mM ATP (yellow), 7.5 mM ATP (magenta), 10 mM ATP (pink), 12.5 mM ATP (orange) or 15 mM ATP (blue). Mean ± SD from three independent experiments each consisting of five technical replicates is presented.
We determined if the reduction in interstitial biofilm expansion might be a consequence of reduced growth in the presence of eATP. The planktonic growth of P. aeruginosa PAK in nutrient media supplemented with eATP at concentrations of 0–15 mM was measured by recording OD595nm over a 20 h period (Figure 1C). These assays show that up to 10 mM eATP does not affect planktonic growth, while at 12.5 mM and 15 mM ATP the planktonic growth rate was slightly reduced. These observations indicate that addition of exogenous eATP at concentrations between 2.5 mM and 10 mM specifically inhibits twitching motility-mediated biofilm expansion of P. aeruginosa while planktonic growth remains unaffected.
To further understand the influence of addition of eATP to twitching motility-mediated expansion of interstitial biofilms, we performed high resolution time-lapse imaging of expanding biofilms in the presence of 0 mM, 2.5 mM, 5 mM and 7.5 mM eATP (Additional file 1: Movie 1). In the absence of eATP cells within the trails behind the leading-edge rafts maintain a lattice-like network, which is characteristic of twitching motility-mediated interstitial biofilm expansion [9,10]. However, at eATP concentrations of 2.5 mM and 5 mM cells within the trails are no longer able to maintain this lattice-like network. While most of the cells are still moving in the presence of 2.5 mM and 5 mM eATP, the individual cells have lost the ability to coordinate their movements and instead appear to be moving randomly which inhibits the ability of P. aeruginosa cells to effectively undergo twitching motility-mediated interstitial biofilm expansion. At an eATP concentration of 7.5 mM cells are non-twitching and form a biofilm edge which is similar to that of the non-twitching PAKpilA::TcR strain [9].
Extracellular ATP decreases surface tfp levels
Our observations have indicated that 10 mM eATP causes twitching motility to cease but does not affect planktonic growth. One mechanism via which this may occur is through modulation of tfp production. To investigate this, levels of surface assembled tfp of wild-type PAK cultured on agar containing 0 mM or 10 mM eATP was assessed by ELISA of whole cells and Western analysis of whole cells and sheared tfp using antisera against the major pilin subunit PilA (Figure 2A, B). These analyses indicate that levels of surface assembled tfp are reduced in the presence of 10 mM eATP (Figure 2A, B) whereas the levels of cell-associated pilin are not altered by 10 mM eATP (Figure 2B). These observations suggest that eATP does not affect tfp pilin expression but rather is likely to be reducing tfp assembly or increasing tfp retraction resulting in the observed reduction of surface-assembled tfp levels.
Figure 2.
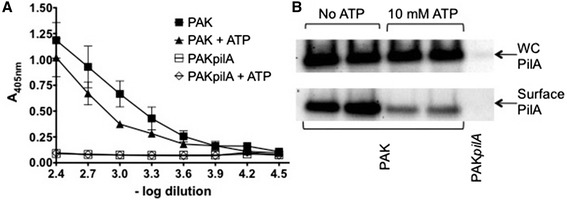
Extracellular ATP decreases levels of surface-assembled tfp. (A) ELISA of surface assembled tfp on whole cells from P. aeruginosa strains PAK and PAKpilA obtained from overnight (20 h) confluent lawns grown at 37°C on LB agar plates containing 0 mM or 10 mM ATP. Levels of surface assembled tfp were detected with anti-PilA anti-serum. Data are presented as the mean ± SD and are representative of results obtained from four independent experiments run in triplicate. (B) Immunoblot of pilin (PilA) detected in whole cell (WC) preparations (top panel) and in sheared surface tfp preparations (bottom panel) of strains PAK (lanes 1 – 4) and PAKpilA (lane 5) obtained from overnight (20 h) confluent lawns grown at 37°C on LB agar plates containing 0 mM or 10 mM ATP. Equal amounts of cells were used in each assay. The results shown in the immunoblots are representative of results obtained in four independent experiments run in duplicate.
The leading edge of actively expanding P. aeruginosa biofilms contains eATP
Our time-lapse microscopy of P. aeruginosa twitching motility-mediated interstitial biofilm expansion in the presence of exogenous eATP indicates that addition of eATP at 2.5–5 mM into the solidified nutrient media appears to inhibit co-ordinated cell movements while an eATP concentration of 7.5 mM causes cell movement to cease (Additional file 1: Movie 1). A possible explanation for this phenomenon is that a gradient of eATP within the active migration zone acts as a directional signal for cells to enable them to differentiate which direction to travel to ensure that overall motility is directed toward virgin territory and away from the mature biofilm. At higher concentrations, eATP may also be acting as a cue to indicate that a cell is no longer located at the leading edge and that movement should cease. Interestingly, Semmler et al. (1999) noted that bacterial cells in the mature regions of the biofilm appear to settle [9], which would be consistent with higher eATP concentrations in this region inducing the cessation of twitching motility.
It has been previously reported that P. aeruginosa secretes eATP in broth culture [20,21]. To explore the possibility that P. aeruginosa produces endogenous eATP in the actively migrating zone at the edge of actively expanding biofilms cultured on solidified nutrient media, we resuspended cells from the actively migrating edge of a colony biofilm and measured the concentration of ATP in the cell-free supernatant. These assays indicate that the active zone of twitching motility at the edge of expanding P. aeruginosa PAK colony biofilms contains about 3 mM eATP. Interestingly, this is significantly higher than eATP concentrations measured in planktonic cultures which were measured in the nm-μm range [20,21]. The higher eATP concentrations that we measured in the active twitching zone of P. aeruginosa colony biofilms may be a consequence of increased secretion and/or reduced diffusion. The ~3 mM concentration of eATP measured in the active twitching zone of the colony biofilm likely reflects the average of the eATP concentrations across the region measured. Our assay was not sufficiently sensitive to determine if there is a gradient of eATP within this zone and if so what was its range. However, given that we have observed that addition of 2.5–10 mM exogenous eATP into the nutrient media appears to inhibit co-ordinated directional movement during twitching motility mediated biofilm expansion and ultimately causes the cells to cease movement, this average value of 3 mM would be consistent with the levels of exogenous eATP confounding an endogenous eATP gradient within the actively migrating zone.
eATP gradients inhibit twitching motility in approaching biofilms
We have observed that if P. aeruginosa is inoculated twice in neighbouring locations on the same media-coated microscope slide that twitching motility at the proximal edges of each adjacent interstitial biofilm is inhibited, resulting in a significantly decreased rate of expansion of the proximal edges compared to the distal edges (Figure 3). As the proximal edges of these biofilms are about 3–8 mm apart, this phenomenon is unlikely to be due to contact-dependent inhibition. Instead these observations suggest that an endogenously produced extracellular signal(s) such as a small molecule is able to diffuse through the media to repress twitching motility-mediated expansion of the neighbouring biofilm. Given our observations that P. aeruginosa colony biofilms contain endogenously produced eATP we hypothesised that these interstitial biofilms may establish a diffused gradient of eATP beyond the colony edge that is sufficient to inhibit twitching motility-mediated expansion of approaching biofilms.
Figure 3.
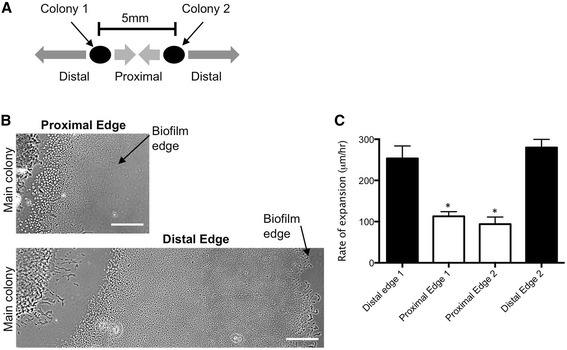
Self-produced extracellular signals inhibit twitching motility-mediated expansion of P. aeruginosa biofilms. P. aeruginosa PAK inoculated at two adjacent locations 5 mm apart on a solidified media-coated microscope slide results in twitching motility-mediated expansion of two neighbouring biofilms at the interstitial space between the media and coverslip. (A) Assay setup: Two P. aeruginosa PAK colonies inoculated on a nutrient media-coated microscope slide 5 mm apart expand to form two neighbouring interstitial biofilms. (B) Representative phase-contrast microscopy images of P. aeruginosa PAK interstitial biofilms formed at the proximal and distal edges of two neighbouring biofilms after incubation at 37°C for 6 h. Scalebar is 200 μm. Images are representative of the twitching motility zones formed in three independent experiments. (C) The rate of expansion via twitching motility away from the main colony at the proximal and distal edges of two neighbouring interstitial biofilms after incubation at 37°C for 6 h. Mean ± SD is presented from three independent experiments (two-tailed Student’s t-test, *p < 0.05).
To explore the possibility that gradients of eATP inhibit twitching motility-mediated expansion of approaching biofilms we established an eATP gradient on media coated slides using ATP-saturated filter discs. Filter discs saturated in H2O were also used to control for the possible influence of increased water content of the medium surrounding the filter discs. P. aeruginosa PAK was then inoculated at a constant distance from the saturated filter discs and a coverslip applied at the edge of the filter disc to establish the interstitial biofilm as per our standard protocol. Time-lapse microscopy was used to examine the effect of the eATP or H2O-saturated discs on twitching motility-mediated interstitial biofilm expansion over time (Additional file 2: Movie 2). This movie demonstrates that while cells continue to flow through the trail network to expand the biofilm towards the H2O-saturated disc, in the presence of an eATP-saturated disc the cells cease twitching motility.
To determine if the observed inhibition by eATP of twitching motility-mediated biofilm expansion by P. aeruginosa strain PAK is likely to be conserved in other P. aeruginosa strains, we examined the influence of eATP gradients on interstitial biofilm expansion by the wild-type P. aeruginosa strains PAO1, PA14 and PA103. These assays showed all strains were similarly inhibited by gradients of eATP (Figure 4A).
Figure 4.
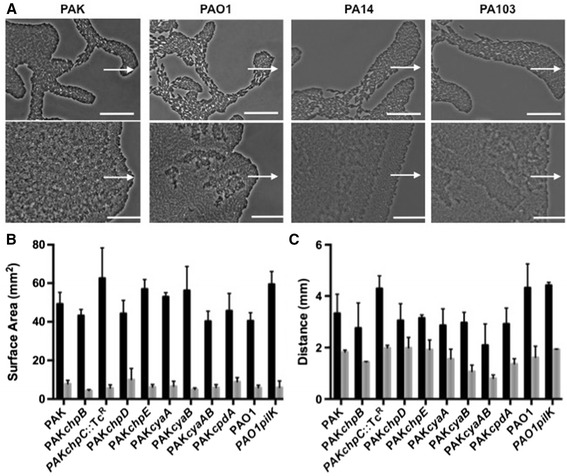
Examination of twitching motility-mediated biofilm expansion in response to eATP by wildtype P. aeruginosa strains and mutants of cyaA , cyaB , cpdA or the Chp chemosensory system. The twitching motility response of P. aeruginosa strains to eATP at the interstitial space between a solidified nutrient media-coated microscope slide and a coverslip after incubation for 15 h at 37°C. (A) Phase-contrast microscopy of interstitial biofilms of wildtype P. aeruginosa strains PAK, PAO1, PA14 and PA103 in the presence of a H2O-saturated disc (upper images) or ATP-saturated disc (lower images). Scalebar is 50 μm and the arrows indicate the direction of expansion towards the disc. Images are representative of three independent experiments. (B) Surface areas of interstitial biofilms formed in the absence of eATP (black bars) or a constant concentration of 7.5 mM eATP (grey bars); (C) the distances expanded towards a H2O-saturated disc (black bars) or ATP-saturated disc (grey bars). The data are represented as the mean ± SD for three independent experiments.
Our observations suggest that P. aeruginosa has the capacity to modulate twitching motility-mediated interstitial biofilm expansion in response to a gradient of eATP. P. aeruginosa twitching motility is regulated by a putative chemosensory system, the Chp system [22-25] which is homologous to the Che chemosensory system which controls swimming motility of Escherichia coli in response to environmental stimuli [26,27]. In P. aeruginosa the core signalling components of this system include a putative histidine kinase encoded by chpA [22]. ChpA is predicted be coupled to a methyl-accepting chemotaxis protein (MCP) receptor, PilJ, by one of two CheW adaptor protein homologues, PilI and ChpC [22]. This complex is predicted to sense a currently unknown environmental signal which results in PilJ undergoing a conformational change, causing ChpA to be autophosphorylated, with the resulting phosphates being transferred to two CheY-like response regulators, PilG and PilH [23,24]. These phosphorylated CheY-homologs are thought to interact with the tfp motor complex to mediate tfp extension and retraction [22,28,29]. Adaptation to the environmental signal(s) is predicted to be mediated through methylation of PilJ by the competing activities of the methyltransferase PilK and the methylestrase ChpB [22,25]. The Chp system gene cluster also encodes the genes chpD and chpE which appear to be part of the operon encoding the Chp system but these are not homologous to components of other chemosensory systems and their function have not as yet been determined [22].
It was not possible to examine mutants of the core Chp system components PilG, PilH, PilI, PilJ and ChpA for defects in eATP responses as these mutants are severely defective in twitching motility [22-24]. However, we explored the possibility that the Chp system may be involved in the control of twitching motility in response to eATP by assaying mutants that retain near wildtype levels of twitching motility and tfp assembly [22,25]. Importantly, as PilK and ChpB are predicted to provide the adaptation mechanism that enables responses to a chemical gradient, we hypothesised that if eATP is indeed sensed by this system it is likely that mutants of pilK and chpB will show defective responses [22,25]. The effects of eATP on the twitching motility of isogenic mutants of the Chp system (chpB, chpC, chpD, chpE and pilK) were examined in our interstitial twitching motility assay in the presence of 7.5 mM eATP incorporated into the media and in our filter disc gradient assay. These assays revealed that all strains showed similar responses to wild-type to both a constant concentration of eATP and an eATP gradient (Figure 4B,C). This suggests that the Chp chemosensory system is not responsible for control of twitching motility-mediated expansion of interstitial biofilms in response to eATP. However, we cannot rule out the possibility that the core components of the Chp system (PilG, PilH, PilI, PilJ and ChpA) are involved.
Twitching motility is modulated by levels of intracellular cAMP (icAMP) [30-32] which are controlled by the adenylate cyclases CyaA and CyaB that catalyse the conversion of ATP to cAMP, and the phosphodiesterase CpdA which degrades cAMP [33,34]. It is possible that eATP is modulating icAMP via these enzymes. We therefore examined the effect of eATP on the twitching motility of isogenic PAK mutants of the icAMP synthesis/degradation system (cyaA, cyaB, cyaAB and cpdA) in our interstitial twitching motility assay in the presence of 7.5 mM eATP incorporated into the media and in our filter disc gradient assay. These assays revealed that all strains showed similar responses to wild-type PAK to both a constant concentration of eATP and an eATP gradient (Figure 4B,C). This suggests that the icAMP synthesis/degradation system is not responsible for control of twitching motility in response to eATP.
Effects of other extracellular nucleotides on twitching motility
To determine whether the observed inhibition of twitching motility was specific for eATP or whether other nucleotides had equivalent effects, gradients of a range of nucleotides were generated using our filter disc gradient assay. Adenosine monophosphate (AMP), adenosine diphosphate (ADP), guanosine triphosphate (GTP), and uridine triphosphate (UTP), did not have any effect on twitching motility even when the interstitial biofilm had approached very close to nucleotide soaked filter disc (Figure 5). The intracellular second messenger signals bis-(3’–5’)-cyclic dimeric guanosine monophosphate (c-di-GMP) and 3’,5’-cyclic adenosine monophosphate (cAMP), which are involved in controlling a transition between motile and sessile modes of life [35] and in tfp assembly and function [30-32], respectively, also had no effect on twitching motility (Figure 5). Interestingly, deoxyadenosine triphosphate (dATP) and cytidine triphosphate (CTP) had similar inhibitory effects as eATP on twitching motility of the approaching interstitial biofilms (Figure 5).
Figure 5.
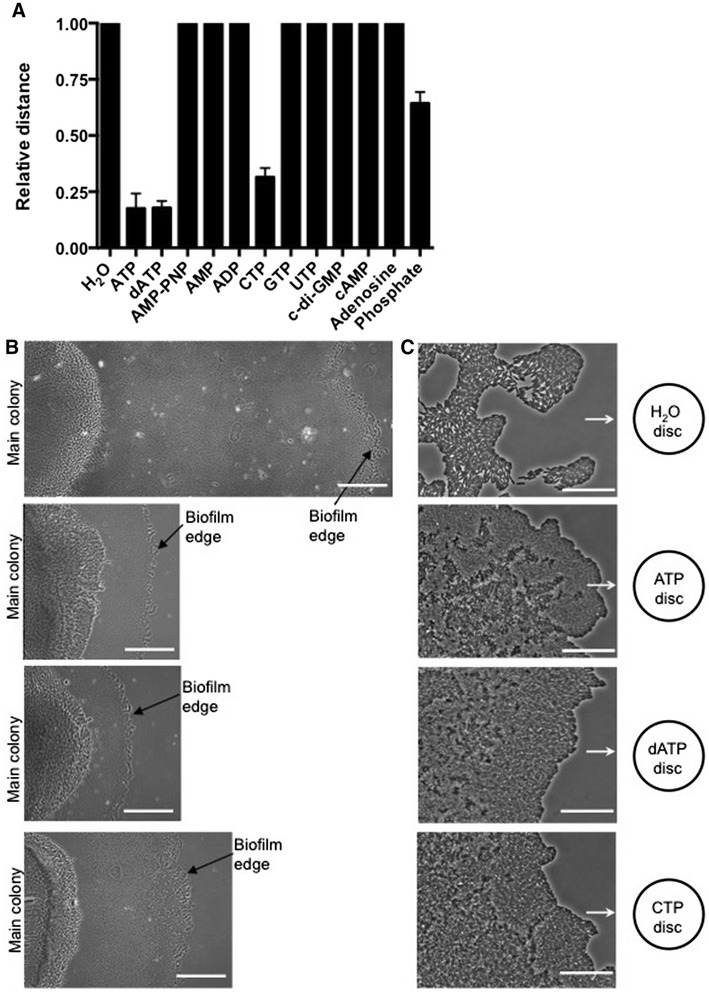
The effect of a range of extracellular nucleotide signals on twitching motility-mediated expansion of P. aeruginosa biofilms. Phase-contrast microscopy of interstitial biofilms of P. aeruginosa PAK in the presence of a range of nucleotides after incubation for 15 h at 37°C. (A) Relative expansion distances of P. aeruginosa PAK interstitial biofilms towards a range of saturated discs after incubation for 15 h at 37°C. A value of 1 indicates that P. aeruginosa expanded up to the edge of the saturated disc. The data are represented as the mean ± SD from four independent experiments. (B) Low magnification images of expansion of the interstitial biofilm away from the main colony towards a H2O, ATP, dATP or CTP-saturated disc (scalebar is 500 μm) and (C) high magnification images of the biofilm edge closest to the disc (scalebar is 50 μm and the arrows indicate the direction of expansion towards the disc). Images are representative of four independent experiments.
To determine if the observed inhibition of twitching motility by eATP required hydrolysis of the molecule, we examined the effect of the non-hydrolysable ATP analogue, adenosine-5’-(β,γ-imido) triphosphate (AMP-PNP) in our filter disc gradient assay. We found that the non-hydrolysable ATP analogue did not affect twitching motility-mediated interstitial biofilm expansion indicating that the observed inhibition of eATP on twitching motility requires ATP hydrolysis (Figure 5). We have determined that adenosine, AMP and ADP do not inhibit twitching motility-mediated interstitial biofilm expansion (Figure 5C). Interestingly, phosphate caused a minor inhibition of twitching motility-mediated interstitial biofilm expansion though not to the extent as was observed for either ATP, dATP or CTP. These observations suggest that the twitching motility response of P. aeruginosa to eATP is likely to involve an enzymatic process involving hydrolysis of ATP as opposed to direct sensing of the ATP molecule, or its hydrolytic products by a signal transduction system.
Discussion
In this study we investigated the effect of eATP on twitching motility by P. aeruginosa. Our observations have revealed that eATP is inhibitory to twitching motility at concentrations that are not inhibitory to planktonic growth. We found that the presence of eATP results in a reduction in surface assembled tfp levels and that inhibition of twitching motility occurs when eATP is either added at a uniform concentration into the solidified nutrient media or when encountered as a gradient from a source external to the biofilm.
Twitching motility mediated biofilm expansion by P. aeruginosa is a complex, multicellular behaviour [9,10] that can lead to the emergence of intricate network patterns of trails that are a characteristic feature of P. aeruginosa interstitial biofilms that form under the conditions of our assay. We have recently determined that extracellular DNA and furrow-mediated stigmergy are involved in this self-organisation of the expanding biofilm [10]. However, it remains unclear how individual bacterial cells are able to determine in which direction to travel to enable overall expansion of the biofilm into new territories. We have shown previously that homoserine lactone quorum sensing is not involved in this process [36]. Our observations in this study indicate that eATP is an important extracellular signalling molecule that co-ordinates twitching motility-mediated biofilm expansion of P. aeruginosa. Time-lapse microscopy suggests that exogenous eATP at concentrations of 2.5–5 mM may be confounding bacteria within the biofilm so that they were unable to maintain directional movement and that higher eATP concentrations signal the cells to cease movement. We also found that the edge of actively expanding P. aeruginosa colony biofilms contain endogenously produced eATP at about 3 mM. Taken together, these observations suggest that a gradient of eATP in the zone of active twitching motility may serve as a signal to direct bacterial traffic toward the virgin territory (low eATP) and away from the older regions of the biofilm (high eATP). High levels of eATP then indicate to the cells that they are no longer near the leading edge and to cease movement.
These observations suggest that a self-produced eATP gradient is sensed by the bacterial cells within the biofilm to direct movement away from regions of high eATP. We examined the possibility that the Chp chemosensory system may be responsible for eATP gradient detection but did not find any evidence that this system is involved. However, as we were unable to examine the core components of this system we cannot completely rule out the possibility. In fact, it is possible that eATP is the central signalling molecule sensed by the Chp chemosensory system to co-ordinate active twitching motility-mediated expansion of the biofilm. Indeed, the observed defects in twitching motility and tfp biogenesis by mutants of these core Chp system components [22-24] are consistent with an inability to detect and respond to eATP.
In this study we determined that ATP hydrolysis appears to be necessary for the inhibition of twitching motility as the non-hydrolysable ATP analogue, AMP-PNP had no effect on P. aeruginosa twitching motility-mediated biofilm expansion. However, we found that none of the possible hydrolytic products of ATP (adenosine, AMP, ADP) are inhibitory and that phosphates are only mildly inhibitory to twitching motility. These observations suggest that eATP or its hydrolytic products are not directly sensed by a signal transduction system to control twitching motility-mediated biofilm expansion. Interestingly, a recent study of eATP release by E. coli and Salmonella sp. in exponential growth phase suggests that this release enhances bacterial survival within stationary phase and that eATP needs to be hydrolysed or degraded at the cell surface [21], which is consistent with our observation that eATP needs to be hydrolysed in order to inhibit P. aeruginosa twitching motility-mediated biofilm expansion. As twitching motility is modulated by icAMP levels and icAMP is generated by the enzymatic conversion of ATP to cAMP we assessed whether the icAMP synthesis/degradation system might be responsible for eATP inhibition of twitching motility. However mutants of this pathway showed no defects in the twitching motility response to eATP indicating that this system is not responsible for eATP hydrolysis. Our observation that CTP and dATP also inhibit P. aeruginosa twitching motility suggests either that the putative enzyme which hydrolyses eATP is also able to use these nucleotides as substrates or that there is more than one hydrolytic enzyme that is able to hydrolyse eATP. As we did not see a similar inhibition of twitching motility in the presence of UTP and GTP, this suggests that the putative enzyme(s) responsible for the hydrolysis of eATP is unable to readily hydrolyse these nucleotides. It is possible that eATP is used by an unidentified kinase to phosphorylate a key component that controls tfp assembly and twitching motility in P. aeruginosa. Interestingly, cAMP and c-di-GMP, two intracellular second messengers which are involved in controlling twitching motility [31,35], did not have any effect on twitching motility when provided exogenously.
P. aeruginosa is an opportunistic pathogen that exploits damaged mucosa to cause infection [5,6]. Interestingly, damage to epithelial cells results in a rapid increase in eATP levels, which results in the recruitment of host immune system factors via the P2X and P2Y receptors [16]. Thus it is likely that P. aeruginosa is exposed to eATP at the site of epithelial damage. As the cytoplasmic concentration of ATP in mammalian cells is of the order of 5–10 mM, injury causing acute plasma membrane damage is expected to result in the release of eATP concentrations in this range at the injury site [37]. Our observations indicate that eATP above 5 mM is sufficient to inhibit twitching motility by P. aeruginosa. This cessation of twitching motility in response to high concentrations of eATP may lead to initiation of infection at the site of epithelial injury. Subsequent cytotoxic damage caused by P. aeruginosa may lead to further release of eATP from the damaged epithelial cells. Interestingly, exposure of mammalian cells to mM concentrations of eATP can lead to necrotic or apoptotic cell death [37]. Our observations indicate that the leading edge of P. aeurginosa biofilms produce eATP in this cytotoxic range which suggests that eATP produced by P. aeruginosa biofilms may be a virulence factor in its own right. Furthermore, chronic P. aeruginosa infection of cystic fibrosis patients is associated with significant inflammatory damage and fibrosis [38] and eATP has recently been associated with pulmonary inflammation and fibrosis via the P2X7 receptor in a murine model of lung fibrosis [39]. Thus it is conceivable that high levels of eATP produced by P. aeruginosa biofilms and through epithelial cell damage contribute to the pathogenesis of chronic P. aeruginosa infection.
Conclusions
This study has shown that eATP produced by P. aeruginosa biofilms functions as an extracellular signalling molecule that co-ordinates cell movements during active biofilm expansion. As P. aeruginosa is an opportunistic pathogen that infects damaged epithelial tissues it is conceivable that the presence of host-derived eATP at sites of epithelial cell damage is exploited by P. aeruginosa to detect potential infection sites. We propose that eATP is an inter-kingdom signalling molecule that contributes to the complex host-pathogen interplay during P. aeruginosa infection.
Methods
Bacterial strains, plasmids and media
The P. aeruginosa strains used in this study are listed in Table 1. P. aeruginosa strains were cultured on Luria-Bertani (LB) [40] broth solidified with agar at 1.5% and grown overnight at 37°C. Planktonic cultures were grown in cation-adjusted Mueller Hinton broth (CAMHB) at 37°C, with shaking at 250 rpm. Twitching motility assays were performed with nutrient media (4 g/L tryptone, 2 g/L, yeast extract, 2 g/L NaCl) solidified with 8 g/L GelGro (ICN). ATP (Sigma Aldrich, St Louis, MO) was added at a final concentration of 1.6–15 mM as indicated. pilK was inactivated by allelic displacement with a tetracycline resistance cassette (TcR) as described previously [22].
Table 1.
List of strains used in this study
| Strain | Relevant characteristics | Source or reference |
|---|---|---|
| PAK | Wild-type P. aeruginosa strain | D. Bradley, Memorial University of Newfoundlands, St John’s, Canada |
| PAO1 | Wild-type P. aeruginosa strain ATCC 15692 | American Type Culture Collection |
| PA14 | Wild-type P. aeruginosa strain | F. Ausubel, Harvard University, Boston, USA |
| PA103 | Wild-type P. aeruginosa strain ATCC 29620 | American Type Culture Collection |
| PAKpilA:TcR | pilA inactivated by allelic displacement with a tetracycline resistance cassette (TcR) | [19] |
| PAKchpB:TcR | chpB inactivated by allelic displacement with a tetracycline resistance cassette (TcR) | [22] |
| PAKchpC:TcR | chpC inactivated by allelic displacement with a tetracycline resistance cassette (TcR) | [22] |
| PAKchpD:TcR | chpD inactivated by allelic displacement with a tetracycline resistance cassette (TcR) | [22] |
| PAKchpE:TcR | chpE inactivated by allelic displacement with a tetracycline resistance cassette (TcR) | [22] |
| PAO1ΔpilK (FA9) | In frame deletion of pilK in wildtype strain PAO1 | [25] |
| PAKΔcyaA | In frame deletion of cyaA | [33] |
| PAKΔcyaB | In frame deletion of cyaB | [33] |
| PAKΔcyaAcyaB | In frame deletions of cyaA and cyaB | [33] |
| PAKΔcpdA | In frame deletion of cpdA | [34] |
Interstitial twitching motility assays
Twitching motility-mediated interstitial biofilm expansion on GelGro-solidified nutrient media was assayed using the solidified nutrient media coated microscope slide assay described previously [18,30]. Interstitial biofilms were examined with phase-contrast microscopy and analysed using FIJI [41].
Filter disc diffusion assays
Filter discs (Whatman 6 mm, GE Healthcare) were soaked with sterile H2O or 100 mM of sodium phosphate or the nucleotides adenosine, adenosine triphosphate (ATP), deoxyadenosine triphosphate (dATP), adenosine monophosphate (AMP), adenosine diphosphate (ADP), cytidine triphosphate (CTP), guanosine triphosphate (GTP), uridine triphosphate (UTP), adenosine-5’-(β,γ-imido)triphosphate (AMP-PNP), 2.9 mM 3’,5’-cyclic diguanylic acid (c-di-GMP) or 30 mM 3’,5’-cyclic adenosine monophosphate (cAMP) and applied to a solidified nutrient media coated microscope slide as follows. 75 μL of the test solution was allowed to fully soak into a filter disc. Each disc was then dried for 2 h and applied to a dried solidified nutrient media coated microscope slide and a gradient allowed to establish for 1 h. The slide was inoculated with the strain of interest 5 mm from the disc, a coverslip applied and incubated at 37°C for the indicated time. In the mutant screening assays each saturated disc was soaked in 30 mM ATP or sterile H2O and used as described above.
Planktonic growth assays
Planktonic growth of P. aeruginosa was followed by recording changes in OD595nm for 20 h. Cells were grown in 96-well microtitre plates, and incubated at 37°C, shaking at 250 rpm. CAMHB or CAMHB supplemented with ATP at indicated concentrations was used in growth assays.
Quantification of eATP in P. aeruginosa biofilms
eATP was detected within P. aeruginosa surface colony biofilms as follows. 30 mL LB 1.5% agar plates were poured and allowed to set overnight at room temperature. The following morning the agar was flipped into a larger petri dish to expose the smooth underside set against the petri dish base which promotes rapid twitching motility-mediated biofilm expansion [9]. 1.5 mL of an overnight P. aeruginosa PAK culture was pelleted by centrifugation (13,000 g, 3 min). The pellet was then gently resuspended and inoculated onto the centre of a flipped 1.5% agar. The large petri-dish lid was applied and incubated in a humid environment for 72 h at 37°C. Cells were harvested from the outer, active twitching edge visualised as a “ground-glass edge”. Harvested cells were resuspended in 1 mL PBS and the eATP detected using a BacTitre-Glo™ assay (Promega Corportation). 500 μL of resuspended cells were pelleted by centrifugation (16, 900 g, 5 min). 110 μl of supernatant was removed and added into triplicate wells of a white 96-well plate (Greiner Bio-one) and 10 μL used to generate four serial 1:11 dilutions, with each well retaining 100 μL final volume. 100 μL of ATP standards ranging from 0–1 μM were also added to the plate in duplicate. 30 μL of room temperature BacTiter-Glo™ Buffer with precombined BacTiter-Glo™ substrate was added to all wells, mixed and incubated at room temperature for 5 min. Luminescence of all wells was read using an integration time of 250 ms.
PilA immunoblotting
Preparation of sheared surface tfp and cell-associated pilin samples was performed as described previously [22] with cells being harvested from plates grown at 37°C on LB agar containing 0 mM or 10 mM ATP for 20 h. Samples were displayed on a 10% Bis-Tris Mini Gel (Life Technologies Corporation) and transferred onto an iBlot® mini gel transfer stack containing a PVDF membrane (Life Technologies Corporation). Membranes were probed with a Western Breeze® chemiluminescent western blot immunodetection kit (Life Technologies Corporation) according to manufacturer’s instructions. A 1:5000 dilution of primary anti-PilA antibody was used.
PilA ELISA
Enzyme-linked immunosorbent assays (ELISAs) to determine the amount of surface tfp of cells grown in the presence and absence of ATP were performed as described previously [22] with cells being harvested from plates grown at 37°C on LB agar containing 0 mM or 10 mM ATP for 20 h.
Acknowledgements
We thank Sarah R. Osvath for technical assistance. L.T. was supported by a UTS Chancellor’s Postdoctoral Fellowship. C.B.W. was supported by an NHMRC Senior Research Fellowship (SRF 571905).
Additional files
Additional file 1: Movie 1.
Exogenous ATP affects twitching motility of P. aeruginosa. Time-lapse phase-contrast microscopy of P. aeruginosa PAK interstitial biofilm expansion on solidified nutrient media-coated microscope slide containing 0 mM, 2.5 mM, 5 mM or 7.5 mM ATP and a coverslip at 37°C. Scalebar is 10 μm. Time (min:s) indicated in top right.
Additional file 2: Movie 2.
Extracellular ATP gradients affect twitching motility of P. aeruginosa. Time-lapse phase-contrast microscopy of P. aeruginosa PAK interstitial biofilm expansion in the presence of an ATP- or H2O-saturated disc after 4 h of incubation at 37°C. Scalebar is 10 μm. Time (min:s) indicated in top right.
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
L.M.N and R.C. performed all experimental work. L.M.N and C.B.W wrote the manuscript. L.M.N., L.T. and C.B.W. conceived and contributed to the design of the study. All authors reviewed the manuscript. All authors read and approved the final manuscript.
Contributor Information
Laura M Nolan, Email: laura.nolan25@gmail.com.
Rosalia Cavaliere, Email: Rosalia.Cavaliere@uts.edu.au.
Lynne Turnbull, Email: Lynne.Turnbull@uts.edu.au.
Cynthia B Whitchurch, Email: Cynthia.Whitchurch@uts.edu.au.
References
- 1.Mahajan-Miklos S, Rahme LG, Ausubel FM. Elucidating the molecular mechanisms of bacterial virulence using non-mammalian hosts. Mol Microbiol. 2000;37(5):981–8. doi: 10.1046/j.1365-2958.2000.02056.x. [DOI] [PubMed] [Google Scholar]
- 2.Rahme LG, Ausubel FM, Cao H, Drenkard E, Goumnerov BC, Lau GW, et al. Plants and animals share functionally common bacterial virulence factors. Proc Natl Acad Sci U S A. 2000;97(16):8815–21. doi: 10.1073/pnas.97.16.8815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lyczak JB, Cannon CL, Pier GB. Establishment of Pseudomonas aeruginosa infection: lessons from a versatile opportunist. Microbes Infect. 2000;2(9):1051–60. doi: 10.1016/S1286-4579(00)01259-4. [DOI] [PubMed] [Google Scholar]
- 4.Tatterson LE, Poschet JF, Firoved A, Skidmore J, Deretic V. CFTR and pseudomonas infections in cystic fibrosis. Front Biosci. 2001;6:D890–7. doi: 10.2741/Tatterso. [DOI] [PubMed] [Google Scholar]
- 5.Ramphal R, Pyle M. Adherence of mucoid and nonmucoid Pseudomonas aeruginosa to acid-injured tracheal epithelium. Infect Immun. 1983;41(1):345–51. doi: 10.1128/iai.41.1.345-351.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kazmierczak BI, Mostov K, Engel JN. Interaction of bacterial pathogens with polarized epithelium. Annu Rev Microbiol. 2001;55:407–35. doi: 10.1146/annurev.micro.55.1.407. [DOI] [PubMed] [Google Scholar]
- 7.Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Sci (New York, NY) 1999;284(5418):1318–22. doi: 10.1126/science.284.5418.1318. [DOI] [PubMed] [Google Scholar]
- 8.Whitchurch CB. Biogenesis and function of type IV pili in pseudomonas species. In: Ramos JL, Levesque RC, editors. Pseudomonas Volume 4 molecular biology of emerging issues. USA: Springer; 2006. pp. 139–88. [Google Scholar]
- 9.Semmler AB, Whitchurch CB, Mattick JS. A re-examination of twitching motility in Pseudomonas aeruginosa. Microbiology. 1999;145:2863–73. doi: 10.1099/00221287-145-10-2863. [DOI] [PubMed] [Google Scholar]
- 10.Gloag ES, Turnbull L, Huang A, Vallotton P, Wang H, Nolan LM, et al. Self-organization of bacterial biofilms is facilitated by extracellular DNA. Proc Natl Acad Sci U S A. 2013;110(28):11541–6. doi: 10.1073/pnas.1218898110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Barker AP, Vasil AI, Filloux A, Ball G, Wilderman PJ, Vasil ML. A novel extracellular phospholipase C of Pseudomonas aeruginosa is required for phospholipid chemotaxis. Mol Microbiol. 2004;53(4):1089–98. doi: 10.1111/j.1365-2958.2004.04189.x. [DOI] [PubMed] [Google Scholar]
- 12.Huang B, Whitchurch CB, Mattick JS. FimX, a multidomain protein connecting environmental signals to twitching motility in pseudomonas aeruginosa. J Bacteriol. 2003;185(24):7068–76. doi: 10.1128/JB.185.24.7068-7076.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hammond A, Dertien J, Colmer-Hamood JA, Griswold JA, Hamood AN. Serum inhibits P aeruginosa biofilm formation on plastic surfaces and intravenous catheters. J Surg Res. 2010;159(2):735–46. doi: 10.1016/j.jss.2008.09.003. [DOI] [PubMed] [Google Scholar]
- 14.Singh PK, Parsek MR, Greenberg EP, Welsh MJ. A component of innate immunity prevents bacterial biofilm development. Nature. 2002;417(6888):552–5. doi: 10.1038/417552a. [DOI] [PubMed] [Google Scholar]
- 15.Patriquin GM, Banin E, Gilmour C, Tuchman R, Greenberg EP, Poole K. Influence of quorum sensing and iron on twitching motility and biofilm formation in pseudomonas aeruginosa. J Bacteriol. 2007;190(2):662–71. doi: 10.1128/JB.01473-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Di Virgilio F. Purinergic signalling in the immune system. A brief update. Purinergic Signal. 2007;3(1–2):1–3. doi: 10.1007/s11302-006-9048-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Burnstock G. Physiology and pathophysiology of purinergic neurotransmission. Physiol Rev. 2007;87(2):659–797. doi: 10.1152/physrev.00043.2006. [DOI] [PubMed] [Google Scholar]
- 18.Turnbull L, Whitchurch CB. Motility assay: twitching motility. Methods Mol Biol (Clifton, NJ) 2014;1149:73–86. doi: 10.1007/978-1-4939-0473-0_9. [DOI] [PubMed] [Google Scholar]
- 19.Watson AA, Mattick JS, Alm RA. Functional expression of heterologous type 4 fimbriae in Pseudomonas aeruginosa. Gene. 1996;175(1–2):143–50. doi: 10.1016/0378-1119(96)00140-0. [DOI] [PubMed] [Google Scholar]
- 20.Hironaka I, Iwase T, Sugimoto S, Okuda KI, Tajima A, Yanaga K, et al. Glucose triggers ATP secretion from bacteria in a growth phase-dependent manner. Appl Environ Microbiol. 2013;79(7):2328–35. doi: 10.1128/AEM.03871-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mempin R, Tran H, Chen C, Gong H, Ho K, Lu S. Release of extracellular ATP by bacteria during growth. BMC Microbiol. 2013;13(1):301. doi: 10.1186/1471-2180-13-301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Whitchurch CB, Leech AJ, Young MD, Kennedy D, Sargent JL, Bertrand JJ, et al. Characterization of a complex chemosensory signal transduction system which controls twitching motility in Pseudomonas aeruginosa. Mol Microbiol. 2004;52(3):873–93. doi: 10.1111/j.1365-2958.2004.04026.x. [DOI] [PubMed] [Google Scholar]
- 23.Darzins A. The pilG gene product, required for Pseudomonas aeruginosa pilus production and twitching motility, is homologous to the enteric, single-domain response regulator CheY. J Bacteriol. 1993;175(18):5934–44. doi: 10.1128/jb.175.18.5934-5944.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Darzins A. Characterization of a Pseudomonas aeruginosa gene cluster involved in pilus biosynthesis and twitching motility: sequence similarity to the chemotaxis proteins of enterics and the gliding bacterium Myxococcus xanthus. Mol Microbiol. 1994;11(1):137–53. doi: 10.1111/j.1365-2958.1994.tb00296.x. [DOI] [PubMed] [Google Scholar]
- 25.Darzins A. The Pseudomonas aeruginosa pilK gene encodes a chemotactic methyltransferase (CheR) homologue that is translationally regulated. Mol Microbiol. 1995;15(4):703–17. doi: 10.1111/j.1365-2958.1995.tb02379.x. [DOI] [PubMed] [Google Scholar]
- 26.Baker MD, Wolanin PM, Stock JB. Signal transduction in bacterial chemotaxis. Bioessays. 2006;28(1):9–22. doi: 10.1002/bies.20343. [DOI] [PubMed] [Google Scholar]
- 27.Wadhams GH, Armitage JP. Making sense of it all: bacterial chemotaxis. Nat Rev Mol Cell Biol. 2004;5(12):1024–37. doi: 10.1038/nrm1524. [DOI] [PubMed] [Google Scholar]
- 28.Guzzo CR, Salinas RK, Andrade MO, Farah CS. PILZ protein structure and interactions with PILB and the FIMX EAL domain: implications for control of type IV pilus biogenesis. J Mol Biol. 2009;393(4):848–66. doi: 10.1016/j.jmb.2009.07.065. [DOI] [PubMed] [Google Scholar]
- 29.Bertrand JJ, West JT, Engel JN. Genetic analysis of the regulation of type IV pilus function by the Chp chemosensory system of Pseudomonas aeruginosa. J Bacteriol. 2010;192(4):994–1010. doi: 10.1128/JB.01390-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Nolan LM, Beatson SA, Croft L, Jones PM, George AM, Mattick JS, et al. Extragenic suppressor mutations that restore twitching motility to fimL mutants of Pseudomonas aeruginosa are associated with elevated intracellular cyclic AMP levels. MicrobiologyOpen. 2012;1(4):490–501. doi: 10.1002/mbo3.49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Inclan YF, Huseby MJ, Engel JN. FimL regulates cAMP synthesis in Pseudomonas aeruginosa. PLoS One. 2011;6(1):e15867. doi: 10.1371/journal.pone.0015867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fulcher NB, Holliday PM, Klem E, Cann MJ, Wolfgang MC. The Pseudomonas aeruginosa Chp chemosensory system regulates intracellular cAMP levels by modulating adenylate cyclase activity. Mol Microbiol. 2010;76(4):889–904. doi: 10.1111/j.1365-2958.2010.07135.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wolfgang MC, Lee VT, Gilmore ME, Lory S. Coordinate regulation of bacterial virulence genes by a novel adenylate cyclase-dependent signaling pathway. Dev Cell. 2003;4(2):253–63. doi: 10.1016/S1534-5807(03)00019-4. [DOI] [PubMed] [Google Scholar]
- 34.Fuchs EL, Brutinel ED, Klem ER, Fehr AR, Yahr TL, Wolfgang MC. In vitro and in vivo characterization of the Pseudomonas aeruginosa Cyclic AMP (cAMP) Phosphodiesterase CpdA, required for cAMP homeostasis and virulence factor regulation. J Bacteriol. 2010;192(11):2779–90. doi: 10.1128/JB.00168-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Simm R, Morr M, Kader A, Nimtz M, Römling U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Mol Microbiol. 2004;53(4):1123–34. doi: 10.1111/j.1365-2958.2004.04206.x. [DOI] [PubMed] [Google Scholar]
- 36.Beatson SA, Whitchurch CB, Semmler ABT, Mattick JS. Quorum sensing is not required for twitching motility in Pseudomonas aeruginosa. J Bacteriol. 2002;184(13):3598–604. doi: 10.1128/JB.184.13.3598-3604.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Di Virgilio F. ATP as a death factor. Biofactors. 1998;8(3–4):301–3. doi: 10.1002/biof.5520080318. [DOI] [PubMed] [Google Scholar]
- 38.Pukhalsky AL, Kapranov NI, Kalashnikova EA, Shmarina GV, Shabalova LA, Kokarovtseva SN, et al. Inflammatory markers in cystic fibrosis patients with lung Pseudomonas aeruginosa infection. Mediators Inflamm. 1999;8(3):159–67. doi: 10.1080/09629359990496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Riteau N, Gasse P, Fauconnier L, Gombault A, Couegnat M, Fick L, et al. Extracellular ATP is a danger signal activating P2X7 receptor in lung inflammation and fibrosis. Am J Respir Crit Care Med. 2010;182(6):774–83. doi: 10.1164/rccm.201003-0359OC. [DOI] [PubMed] [Google Scholar]
- 40.Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. 2. Cold Spring Harbor, N. Y: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- 41.Schindelin J, Arganda-Carreras I, Frise E, Kaynig V, Longair M, Pietzsch T, et al. Fiji: an open-source platform for biological-image analysis. Nat Methods. 2012;9(7):676–82. doi: 10.1038/nmeth.2019. [DOI] [PMC free article] [PubMed] [Google Scholar]