

Abstract
The gut microbiota of many phylogenetically lower termites is dominated by the cellulolytic flagellates of the genus Trichonympha, which are consistently associated with bacterial symbionts. In the case of Endomicrobia, an unusual lineage of endosymbionts of the Elusimicrobia phylum that is also present in other gut flagellates, previous studies have documented strict host specificity, leading to the cospeciation of “Candidatus Endomicrobium trichonymphae” with their respective flagellate hosts. However, it currently remains unclear whether one Trichonympha species is capable of harboring more than one Endomicrobia phylotype. In the present study, we selected single Trichonympha cells from the guts of Zootermopsis nevadensis and Reticulitermes santonensis and characterized their Endomicrobia populations based on internal transcribed spacer (ITS) region sequences. We found that each host cell harbored a homogeneous population of symbionts that were specific to their respective host species, but phylogenetically distinct between each host lineage, corroborating cospeciation being caused by vertical inheritance. The experimental design of the present study also allowed for the identification of an unexpectedly large amount of tag-switching between samples, which indicated that any high-resolution analysis of microbial community structures using the pyrosequencing technique has to be interpreted with great caution.
Keywords: termites, protists, endosymbionts, cospeciation, pyrosequencing
The termite hindgut is colonized by dense assemblages of prokaryotes (bacteria and archaea) and eukaryotic protists that play essential roles in both digestion and host nutrition (4, 20). The gut microbiota of phylogenetically lower termites is dominated by cellulolytic flagellates that are unique to termites and comprise diverse lineages from the phyla Parabasalia and Preaxostyla (order Oxymonadida) (6). In most cases, these flagellates are associated with large numbers of prokaryotic symbionts that colonize both the surface and cytoplasm (19, 38) and sometimes even the nucleus of their hosts (40). There is strong evidence to show that the symbionts complement the nitrogen metabolism of the flagellates and play an important nutritional role in the hindgut microecosystem (3, 6, 19, 20).
The predominant flagellates in the hindgut of several termite lineages are the hypermastigid flagellates of the genus Trichonympha. Originally discovered in Reticulitermes spp. (Rhinotermitidae) (16, 29, 30), members of the genus have also been described in several other termite families (including Termopsidae and Kalotermitidae) and in their sister group, the wood-feeding cockroaches of the genus Cryptocercus (for references, see 51). While some termite species contain only a single species of Trichonympha, others harbor several representatives of this genus.
Termites of the genus Zootermopsis harbor at least four Trichonympha species. Trichonympha campanula (27), Trichonympha sphaerica (26), and Trichonympha collaris (25) have long been recognized based on differences in morphology; the fourth species was discovered using molecular approaches. The first evidence of the existence of this fourth species was the presence of an additional phylotype of small subunit (SSU) rRNA gene sequences in a cell suspension of T. campanula from Zootermopsis nevadensis that was differentiated from the cells of T. campanula by fluorescence in situ hybridization (21). A few years later, this species, Trichonympha postcylindrica, was described based on specimens from the gut of Zootermopsis angusticollis (45).
Although the same four Trichonympha species appear to be present in both Z. nevadensis and Z. angusticollis (21, 25, 45), their relative abundance may differ even among individuals from the same colony (25). Moreover, there is a considerable range of sequence divergence within the populations of each Trichonympha species in Z. angusticollis (45). Current knowledge on the diversity of Trichonympha species in the two Zootermopsis species, including the hitherto unpublished sequence of T. postcylindrica from Z. nevadensis (21), is summarized in Fig. 1.
Fig. 1.

Diversity of 18S rRNA gene sequences of Trichonympha flagellates in Zootermopsis nevadensis (21), Zootermopsis angusticollis (grouped nodes) (45), and different Reticulitermes species (Rlu, Reticulitermes lucifugus; Rsa, Reticulitermes santonensis; Rfl, Reticulitermes flavipes; Rhe, Reticulitermes hesperus) (23). The maximum-likelihood tree was rooted with Trichonympha species of Clusters II and III. Circles indicate node support (○, >80%; ●, >95% bootstrap confidence).
Members of the genus Trichonympha fall into three distinct phylogenetic clusters (9, 23, 37), and all are associated with bacterial symbionts, quite often more than one bacterial species per flagellate host (39, 40, 44). All members of Trichonympha Cluster I harbor large populations of the specific intracellular symbiont, “Candidatus Endomicrobium trichonymphae” (22, 36, 43), which belongs to a distinct lineage of uncultured, insect-associated bacteria (“Endomicrobia”) in the Elusimicrobia phylum (5). The presence of homogeneous bacterial symbiont populations in a single flagellate host cell (18) and an almost perfect congruence of the phylogenies of the bacterial symbionts and their host flagellates (23) suggest that “Ca. Endomicrobium trichonymphae” is propagated exclusively by vertical transmission (cytoplasmic inheritance) in the Trichonympha lineage. In this case, the same number of Endomicrobia species as Trichonympha species is expected in a given termite. This is in apparent contradiction with the composition of a metagenomic library constructed from DNA prepared from the enrichment of “Ca. Endomicrobium trichonymphae” in the gut of Z. nevadensis, which contains a larger number of 16S rRNA phylotypes of Endomicrobia than host species (IMG Project ID: Gi01566) (21). It currently remains unclear whether all individuals of each host species harbor the same symbionts and whether a single Trichonympha cell carries more than one Endomicrobia phylotype.
To clarify this, we isolated single Trichonympha cells from the guts of Z. nevadensis and R. santonensis and characterized their Endomicrobia populations by high-throughput sequencing, using the internal transcribed spacer (ITS) region between their rRNA genes to resolve even low levels of strain variation within both symbionts and hosts.
Materials and Methods
Termites
Zootermopsis nevadensis was collected near the Chilao Flats Campground, Angeles National Forest, California, USA in 2006. Reticulitermes santonensis, which is synonymous with Reticulitermes flavipes (1), was collected near La Gautrelle, Ile d’Oléron, France in 2010. Since then, both species have been maintained in the laboratory on a diet of pine wood and water. Worker termites (pseudergates) were used in all experiments.
Micromanipulation and whole-genome amplification
Termites were dissected and the entire hindgut content was carefully diluted in Solution U (48) in Eppendorf tubes. Aliquots of this suspension (10 μL) were transferred to the wells of a Teflon-coated microscope slide and inspected with an inverted microscope with phase-contrast optics (50-fold magnification). Individual Trichonympha cells were identified by their morphology and captured using a micropipette attached to a micromanipulator, as described previously (47), and transferred to a PCR tube with 50 μL Solution U. The tubes were incubated at 95°C for 10 min to lyse the flagellates, cooled on ice for 2 min, and centrifuged at a slow speed (50 × g) for 1 min to remove cell debris. The supernatant (including the endosymbionts) was used as a template for multiple displacement amplification (MDA) with the REPLI-g UltraFast Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions, except that the reaction time was increased to 4 h.
Library preparation and sequencing
The ITS regions of flagellates and Endomicrobia were amplified by PCR using the MDA products (25-fold diluted) as a template and specific primer pairs for the proximal regions of the flanking rRNA genes (Fig. 2). The combination of the Trichonympha-specific forward primer 18S-Tri-1287f (AAGATTCACGTAGC TGGG; this study) and the universal reverse primer 28S-1r (ATGCTTAAATTCAGCGGGT) (35) yielded PCR products of ~700 bp (30 cycles of amplification: 30 s at 94°C, 30 s at 54°C, and 60 s at 72°C), which were directly sequenced as previously described (47).
Fig. 2.
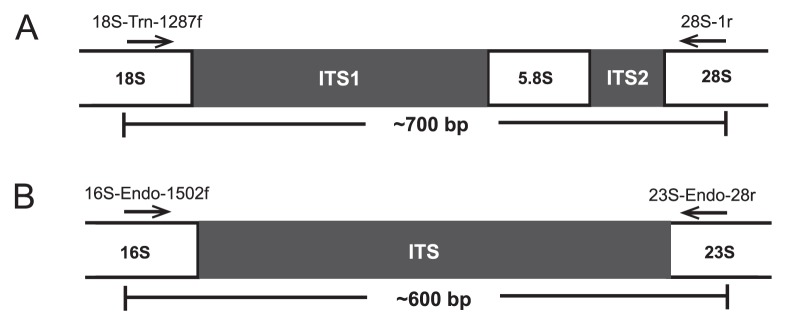
Strategy used for PCR amplification of ITS regions of (A) Trichonympha flagellates and (B) their Endomicrobia symbionts. The primers, their target positions in the proximal regions of the flanking rRNA genes, and the resulting amplicon length are indicated.
The Endomicrobia-specific primers 16S-Endo-1502f (AAGGT AGCCGTACGAGA) and 23S-Endo-28r (ACAGTCTTAGCCAA GGCA) were designed on the basis of all Endomicrobia sequences represented in public databases. Both primers were barcoded as described previously (28). Endomicrobia sequences were also amplified directly from undiluted DNA extracted from the whole-gut homogenates of Z. nevadensis and R. santonensis (35 cycles of amplification: 30 s at 94°C, 30 s at 56°C, and 60 s at 72°C). All samples were commercially sequenced in a single sequencing run (454 GS FLX 64 with Titanium technology; GATC Biotech, Konstanz, Germany).
Bioinformatics
The ITS sequences of the selected flagellates were aligned de novo with MAFFT version 7 (24). After manual curation of the alignment, a neighbor-joining tree was constructed using the ARB software suite (31). The ITS sequences of Endomicrobia were processed as previously described (12). Briefly, pyrotag reads with a minimum length of 250 bp and a maximum expected error of 0.5 were selected and demultiplexed using their barcode sequences (no mismatch allowed). After the removal of barcodes and primer sequences, the sequences were clustered at the 99% similarity level with UPARSE (13). Sequences were dereplicated, and representative phylotypes (most abundant sequence in the respective cluster) were aligned de novo using the MAFFT aligner in the L-INS-I mode with 100 iterations (24). Where necessary, the alignment was manually refined so that all sequences were unambiguously aligned. Maximum-likelihood trees were constructed using RAxML version 8.1.3 (42) with the 16-state GTR-Γ model and 1,000 bootstraps. A heatmap was generated using the R software with the package heatmap.plus (11).
Sequence accession numbers
The ITS region sequences of flagellates have been deposited in GenBank under accession numbers KJ778566–KJ778610. Representative sequences of all Endomicrobia phylotypes obtained from flagellate samples and whole-gut homogenates have been deposited in GenBank under accession numbers KP058245–KP058309.
Results
Isolation and identification of single Trichonympha cells
We successfully amplified the ITS regions of host flagellates (Fig. 2A) and Endomicrobia symbionts (Fig. 2B) from the whole-genome amplification products obtained from the selected Trichonympha cells from Z. nevadensis (34 out of 60 cells) and R. santonensis (9 out of 20 cells). Direct sequencing of the PCR products obtained with flagellate-specific primers yielded clean signals for all species, except for T. campanula, in which multiple bases at several positions of the trace file indicated sequence polymorphism in the ITS region (Supplementary Fig. S1).
A comparative sequence analysis showed that each of the sequences obtained from the morphotypes of T. sphaerica, T. postcylindrica, T. collaris, and T. campanula in Z. nevadensis clustered with their respective relatives from Z. angusticollis (Fig. 3), confirming the morphological assignment of the selected flagellates. The nine sequences from the morphotype of T. agilis formed a sister group of the Trichonympha species from Z. nevadensis.
Fig. 3.

Neighbor-joining tree of the ITS sequences obtained from selected flagellates, illustrating the phylogenetic diversity of the Trichonympha species from Zootermopsis nevadensis (this study) and their relationship to those from Zootermopsis angusticollis (45; sequences are grouped). The tree is based on an alignment of 450 base positions and was rooted with the ITS sequences of Trichonympha agilis from Reticulitermes santonensis. Phase-contrast photomicrographs illustrate the morphology of the respective species (scale bars=50 μm). Circles indicate node support (○, >80%; ●, >95% bootstrap confidence).
Endomicrobia populations in single Trichonympha cells
A pyrosequencing analysis of the PCR products obtained with Endomicrobia-specific primers yielded variable read numbers per flagellate sample (after quality control, see Supplementary Fig. S2). Although the number of Endomicrobia phylotypes (99% sequence identity) obtained from all samples (17 phylotypes from 45 single host cells) was markedly larger than the number of flagellate species investigated (5 species), they all clustered more or less closely according to their respective hosts (Fig. 4A).
Fig. 4.

(A) Maximum-likelihood tree of all Endomicrobia phylotypes (99% sequence similarity) detected in whole-genome amplification (MDA) products of all single flagellate cells. Branches shared by phylotypes from the same host species are color coded; the area of the circles indicates the relative abundance of the major phylotype in each library. Black dots indicate the position of phylotypes that originated from whole-gut samples of Zootermopsis nevadensis or Reticulitermes santonensis. (B) Pie charts indicate the relative abundance of the different Endomicrobia clusters in the whole gut samples. The colors represent the major phylotypes from selected Trichonympha cells; phylotypes from whole-gut samples are shown in grey. Phylotypes in the Pyrsonympha cluster were identified based on the sequences retrieved from selected flagellates from R. santonensis (unpublished results).
While the Endomicrobia obtained from nine different cells of T. agilis comprised only two closely related phylotypes, the symbionts of the other flagellates, particularly T. postcylindrica and T. campanula, were far more diverse (Fig. 4A). Phylogenetically distinct flagellates carried different phylotypes of symbionts, and even the Endomicrobia in host cells with identical ITS sequences sometimes differed slightly in their respective phylotype (e.g., T. sphaerica ZnvTrn10 and ZnvTrn48).
When we analyzed the diversity of Endomicrobia sequences obtained from the whole-gut DNA of Z. nevadensis, we found several additional phylotypes (represented by black branches in Fig. 4A) within the radiation of the Endomicrobia sequences retrieved from selected Trichonympha cells, which indicated that the Trichonympha populations in this termite were not exhaustively sampled. Nevertheless, the vast majority of the Endomicrobia in Z. nevadensis consisted of phylotypes retrieved from T. collaris (41%) and T. sphaerica (33%), while the diverse populations of Endomicrobia associated with T. postcylindrica (13%) and T. campanula (6%) represented only a small part of the community (Fig. 4B). In the case of R. santonensis, the two phylotypes of Endomicrobia from T. agilis accounted for 23% of the phylotypes in the gut homogenate, and the majority of the reads clustered with the ITS sequences of the selected Pyrsonympha flagellates (unpublished results), which form a distinct branch in the phylogeny of Endomicrobia (43).
Pyrosequencing artifacts
Although each of the pyrotag libraries of Endomicrobia from single flagellates consistently contained only one major phylotype (Fig. 5), they always comprised a smaller fraction of reads that were identical with the dominant phylotypes in other libraries. The shared presence of the same Endomicrobia phylotype in different flagellate samples of Z. nevadensis may have been caused by the interspecific transfer of endosymbionts within the gut, whereas the presence of identical phylotypes in Trichonympha species of different termites was unexpected because it can only be explained by multiple recent transfers of endosymbionts between flagellates of different termite species. Therefore, we suspected that the shared phylotypes were artifacts that either resulted from the picking process (e.g., the symbionts stemmed from other, lysed flagellate cells present in the same gut) or were generated during the pyrosequencing process.
Fig. 5.

Heatmap of the relative abundance of different phylotypes of Endomicrobia within pyrotag libraries of all single flagellate cells. The samples were ordered according to the clusters indicated in Fig. 2. The samples that were checked for purity by cloning and Sanger sequencing are in red.
To clarify this, we reanalyzed three of the Endomicrobia samples obtained from individual flagellates (R05, Z13, and Z21), which were selected because they had particularly large proportions of suspicious sequences (see Fig. 5). To exclude the possibility that the minor phylotypes in the corresponding pyrotag libraries stemmed from contamination during the picking process or originated during whole-genome amplification or library preparation (e.g., by contaminated tags), we used aliquots of the samples that had been directly preserved before pyrosequencing (i.e., after the PCR step that introduced the sample-specific tags). The amplicons were ligated into a plasmid vector, and 30 clones of each sample were sequenced. In each case, all sequences in the library were identical to the major phylotype obtained from the respective flagellate, which clearly identified the minor phylotypes in the corresponding pyrotag libraries as artifacts generated during the pyrosequencing process.
This interpretation is supported by the observation that the reads of Endomicrobia phylotype Tsph1, which was present in most T. sphaerica samples and overrepresented in the pyrosequencing run, were recovered with varying abundance from all other flagellate samples (Fig. 5; Supplementary Fig. S2). To test this quantitatively, we compared the number of reads for the major phylotype in each flagellate sample to the number of identical reads recovered from samples of other flagellate species (Fig. 6). The strong linear correlation corroborated the minor phylotypes in each sample not being real, but being artifacts caused by tag switching.
Fig. 6.

Correlation of the number of reads for the major phylotype in each pyrotag library of individual flagellates to the number of identical reads recovered from libraries of other flagellate species. Each dot represents the results obtained for an individual sample.
Discussion
Host specificity of Endomicrobia
In the present study, we showed that each of the Trichonympha cells from both Z. nevadensis and R. santonensis harbored a single phylotype of “Ca. Endomicobium trichonymphae”. The symbiont populations in each host lineage were phylogenetically distinct even if multiple host species were present in the same gut, corroborating the assumption that the apparent cospeciation between the partners in this symbiosis was caused by vertical (cytoplasmic) inheritance of the endosymbionts (23).
Although Ikeda-Ohtsubo and Brune (23) were able to document a congruent topology of the SSU rRNA trees of the symbiotic pairs, the relatively small number of sequences in their clone libraries (10–20 clones) did not allow us to exclude the presence of minor phylotypes of Endomicrobia within each Trichonympha cell. Moreover, since the libraries were obtained from flagellate suspensions containing numerous host cells (100–200 cells per sample), it remained possible that individual flagellates harbored a phylotype entirely different from that of the majority of the cells in the suspension.
Prompted by the finding that a large Endomicrobia clone library of Z. nevadensis (353 clones) contained a markedly larger number of 16S rRNA phylotypes than the Trichonympha species described for this flagellate (21), we reinvestigated the subject. Since the resolution of the 16S rRNA analysis was not sufficient due to the high similarity of the sequences, we used the more variable ITS region, harnessing the advantages of whole-genome amplification and next-generation sequencing technology to achieve a highly resolved analysis of their community structure based on the genomic DNA of the symbiont populations in individual host cells. Beyond the artifacts inherent to the pyrotag method (see below), we concluded that (i) each Trichonympha cell harbored a homogeneous population of Endomicrobia symbionts, (ii) Trichonympha cells of different species always harbored a phylogenetically distinct symbiont population, and (iii) the phylogenetic diversity of Endomicrobia in the two termites investigated in this study was caused by microdiversity in the Trichonympha populations that extended beyond the framework of the currently described species.
The present study could not completely exclude the possibility of an exchange of symbionts between individuals of the same (or closely related) Trichonympha populations present within the same gut. While the Trichonympha species in the wood-feeding cockroach Cryptocercus punctulatus still reproduce sexually during each molt of the host, this trait has been lost in the Trichonympha lineages of termites at an uncertain evolutionary time point (10). However, artificial feeding of Z. angusticollis with the molting hormone 20- hydroxyecdysone still appeared to trigger a sexual cycle in their Trichonympha flagellates (32), and such copulation events may still allow for the mixing of Endomicrobia phylotypes among host cells of the same species on rare occasions. Nevertheless, the strong and ongoing genome reduction demonstrated in “Ca. Endomicrobium trichonymphae” strain Rs-D17 (18) indicates the absence of genetic exchange and a frequent population bottleneck during transmission (50), a well-documented phenomenon in the intracellular symbionts in the bacteriocytes of aphids (33, 34).
Endomicrobia populations in Zootermopsis and Reticulitermes
Assuming that all Endomicrobia species had the same copy number of rRNA genes, the Endomicrobia community in the gut of Z. nevadensis was dominated by phylotypes from T. collaris (43%) and T. sphaerica (31%). These outnumbered the diverse populations of “Ca. Endomicrobium trichonymphae” associated with T. postcylindrica and T. campanula, which accounted for another 19% of the community (Fig. 4B). Considering that the colonization density of Endomicrobia differed among the four Trichonympha phylotypes in Z. nevadensis (21), this is in reasonable agreement with the relative abundance of Trichonympha species in this termite, as reported by Kirby (25) who found that the three species (T. postcylindrica was not recognized as separate from T. campanula) were typically present in comparable numbers. The phylogenetic position of the remaining phylotypes (5% of the reads in the library) recovered only from the gut of Z. nevadensis and not from the selected flagellates (Fig. 4A) suggested that they represented Endomicrobia populations of Trichonympha lineages that were not included among the selected cells, but whose presence was indicated by the microdiversity of the ITS sequences obtained for each morphotype (Fig. 3).
In the case of the T. agilis cells selected from the gut of R. santonensis, the microdiversity of Endomicrobia (Fig. 4A) reflected that of the host flagellates (Fig. 3), which indicated the strict host specificity of the endosymbionts. Kirby (25) reported that, with the exception of Reticulitermes lucifugus, all Reticulitermes species investigated harbored only a single species of Trichonympha, which was in agreement with the number of SSU rRNA genes of Trichonympha species obtained from different Reticulitermes species (Fig. 1). However, the ITS sequences of the selected flagellates revealed the presence of two closely related phylotypes in R. santonensis (Fig. 3), which is consistent with the two phylotypes of “Ca. Endomicrobium trichonymphae” obtained from these samples (Fig. 4).
The reads obtained for the amplified ITS regions of the individual flagellates of T. campanula often contained traces of sequence polymorphism, suggesting that the rRNA gene clusters had multiple and slightly divergent copies. To date, there has been no genome information for any termite gut flagellate; however, the draft genome of the distantly related parabasalid Trichomonas vaginalis contains 254 copies of the 18S rRNA gene (8). ITS regions experience low selective constraint and are considered to evolve rapidly by large insertions and/or deletions (14, 41).
Methodological pitfalls of pyrotag sequencing
In amplicon pyrosequencing, the addition of sequence tags to the amplified target sequences allows for the parallel processing of numerous samples in the same sequencing run (17). The tags are typically added to both ends of the amplicons because this allows for the identification of cases of tag switching (2), which result in impossible tag combinations at each terminus and may affect a large proportion of the reads in pyrotag libraries (7, 49).
However, tag switching can be recognized and removed during quality control only if both termini of the amplicon are present in the same read. If the length of the amplicon exceeds the read length, a common problem with most sequencing platforms, it is not possible to identify mistagged sequences without any prior information. In this case, they either go entirely unnoticed or are purged from the dataset by omitting all reads that do not pass a certain frequency threshold (e.g., 46).
In the present study, the necessities of primer design dictated an amplicon length of approximately 600 bp (the ITS region of Endomicrobia; Fig. 2B), which exceeded even the read length of the 454 Titanium technology (up to 400 bp) (15). However, the low diversity of the Endomicrobia community and the highly improbable presence of identical phylotypes in host flagellates of different termites allowed us to identify reads with suspicious tags. Their frequency amounted to 15% of the reads in the entire sequencing run (Supplementary Fig. S2), which is at the upper end of the proportion of mistagged reads reported in previous studies (49). The complete absence of these reads from the processed samples prior to the sequencing run excludes them resulting from contaminated tags and indicated that they originated during the pyrosequencing step, possibly from tag switching during emulsion PCR, as suggested previously (7). Since the number of mistagged reads in each sample depends on the number of reads carrying a particular tag (Fig. 6; Supplementary Fig. S2), mistagged reads cannot be removed simply by applying a frequency threshold. In cases that are highly sensitive to tag switching, e.g., the identification of core communities shared across multiple samples, such artifacts have to be excluded using an appropriate experimental design. Otherwise, the simultaneous presence of OTUs in different samples of a pyrotag run has to be interpreted with the necessary caution (12).
Supplementary Information
Acknowledgements
This study was funded by the Max Planck Society. Hao Zheng is a doctoral student in the International Max Planck Research School for Environmental, Cellular and Molecular Microbiology (IMPRSMIC), Marburg. Claire L. Thompson was funded by a postdoctoral fellowship of the LOEWE Center for Synthetic Microbiology, Marburg. We thank Jared Leadbetter (Caltech) for providing the termites.
References
- 1.Austin JW, Szalanski AL, Scheffrahn RH, Messenger MT, Dronnet S, Bagnères AG. Genetic evidence for the synonymy of two Reticulitermes species: Reticulitermes flavipes and Reticulitermes santonensis. Ann Entomol Soc Am. 2005;98:395–401. [Google Scholar]
- 2.Binladen J, Gilbert MTP, Bollback JP, Panitz F, Bendixen C, Nielsen R, Willerslev E. The use of coded PCR primers enables high-throughput sequencing of multiple homolog amplification products by 454 parallel sequencing. PLoS One. 2007;2:e197. doi: 10.1371/journal.pone.0000197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Brune A. Microbial symbioses in the digestive tract of lower termites. In: Rosenberg E, Gophna U, editors. Beneficial Microorganisms in Multicellular Life Forms. Springer; Heidelberg: 2011. pp. 3–25. [Google Scholar]
- 4.Brune A, Ohkuma M. Role of the termite gut microbiota in symbiotic digestion. In: Bignell DE, Roisin Y, Lo N, editors. Biology of Termites: A Modern Synthesis. Springer; Dordrecht: 2011. pp. 439–475. [Google Scholar]
- 5.Brune A. Endomicrobia: intracellular symbionts of termite gut flagellates. J Endocytobiosis Cell Res. 2012;23:11–15. [Google Scholar]
- 6.Brune A. Symbiotic digestion of lignocellulose in termite guts. Nat Rev Microbiol. 2014;12:168–180. doi: 10.1038/nrmicro3182. [DOI] [PubMed] [Google Scholar]
- 7.Carlsen T, Aas AB, Lindner D, Vralstad T, Schumacher T, Kauserud H. Don’t make a mista(g)ke: is tag switching an overlooked source of error in amplicon pyrosequencing studies? Fungal Ecol. 2012;5:747–749. [Google Scholar]
- 8.Carlton JM, Hirt RP, Silva JC, et al. Draft genome sequence of the sexually transmitted pathogen Trichomonas vaginalis. Science. 2007;315:207–212. doi: 10.1126/science.1132894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Carpenter KJ, Chow L, Keeling PJ. Morphology, phylogeny, and diversity of Trichonympha (Parabasalia: Hypermastigida) of the wood-feeding cockroach Cryptocercus punctulatus. J Eukaryotic Microbiol. 2009;56:305–313. doi: 10.1111/j.1550-7408.2009.00406.x. [DOI] [PubMed] [Google Scholar]
- 10.Cleveland LR. Hormone-induced sexual cycles of flagellates. I. gametogenesis, fertilization, and meiosis in Trichonympha. J Morphol. 1949;85:197–295. doi: 10.1002/jmor.1050850202. [DOI] [PubMed] [Google Scholar]
- 11.Day A. heatmap.plus—heatmap with more sensible behavior. R package version 1.3. 2012 Available at http://cran.r-project.org/web/packages/heatmap.plus/index.html.
- 12.Dietrich C, Köhler T, Brune A. The cockroach origin of the termite gut microbiota: patterns in bacterial community structure reflect major evolutionary events. Appl Environ Microbiol. 2014;80:2261–2269. doi: 10.1128/AEM.04206-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Edgar RC. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat Methods. 2013;10:996–998. doi: 10.1038/nmeth.2604. [DOI] [PubMed] [Google Scholar]
- 14.García-Martínez J, Acinas SG, Antón AI, Rodriguez-Valera F. Use of the 16S-23S ribosomal genes spacer region in studies of prokaryotic diversity. J Microbiol Methods. 1999;36:55–64. doi: 10.1016/s0167-7012(99)00011-1. [DOI] [PubMed] [Google Scholar]
- 15.Glenn TC. Field guide to next-generation DNA sequencers. Mol Ecol Resour. 2011;11:759–769. doi: 10.1111/j.1755-0998.2011.03024.x. [DOI] [PubMed] [Google Scholar]
- 16.Grassi B, Sandias A. The constitution and development of the society of termites: observations on their habits; with appendices on the parasitic protozoa of Termitidae, and on the Embiidae. Quart J Microsc Sci. 1897;39:245–332. [Google Scholar]
- 17.Hamady M, Walker JJ, Harris JK, Gold NJ, Knight R. Error-correcting barcoded primers for pyrosequencing hundreds of samples in multiplex. Nat Methods. 2008;5:235–237. doi: 10.1038/nmeth.1184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hongoh Y, Sharma VK, Prakash T, et al. Complete genome of the uncultured Termite Group 1 bacteria in a single host protist cell. Proc Natl Acad Sci USA. 2008;105:5555–5560. doi: 10.1073/pnas.0801389105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Hongoh Y, Ohkuma M. Termite gut flagellates and their methanogenic and eubacterial symbionts. In: Hackstein JHP, editor. (Endo)Symbiotic Methanogenic Archaea. Springer; Heidelberg: 2010. pp. 55–79. [Google Scholar]
- 20.Hongoh Y. Toward the functional analysis of uncultivable, symbiotic microorganisms in the termite gut. Cell Mol Life Sci. 2011;68:1311–1325. doi: 10.1007/s00018-011-0648-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ikeda-Ohtsubo W. PhD thesis. Philipps-Universität Marburg; Marburg, Germany: 2007. Endomicrobia in termite guts: symbionts within a symbiont. [Google Scholar]
- 22.Ikeda-Ohtsubo W, Desai M, Stinglt U, Brune A. Phylogenetic diversity of ‘Endomicrobia’ and their specific affiliation with termite gut flagellates. Microbiology. 2007;153:3458–3465. doi: 10.1099/mic.0.2007/009217-0. [DOI] [PubMed] [Google Scholar]
- 23.Ikeda-Ohtsubo W, Brune A. Cospeciation of termite gut flagellates and their bacterial endosymbionts: Trichonympha species and ‘Candidatus Endomicrobium trichonymphae’. Mol Ecol. 2009;18:332–342. doi: 10.1111/j.1365-294X.2008.04029.x. [DOI] [PubMed] [Google Scholar]
- 24.Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30:772–780. doi: 10.1093/molbev/mst010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kirby H. Flagellates of the genus Trichonympha in termites. Univ Calif Publ Zool. 1932;37:349–476. [Google Scholar]
- 26.Kofoid CA, Swezy O. Studies on the parasites of the termites IV. On Leidyopsis sphaerica gen. nov., sp. nov. Univ Calif Publ Zool. 1919;20:99–116. [Google Scholar]
- 27.Kofoid CA, Swezy O. Studies on the parasites of the termites III. On Trichonympha campanula sp. nov. Univ Calif Publ Zool. 1919;20:41–98. [Google Scholar]
- 28.Köhler T, Dietrich C, Scheffrahn RH, Brune A. High-resolution analysis of gut environment and bacterial microbiota reveals functional compartmentation of the gut in wood-feeding higher hermites (Nasutitermes spp.) Appl Environ Microbiol. 2012;78:4691–4701. doi: 10.1128/AEM.00683-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Koidzumi M. Studies on the intestinal protozoa found in the termites of Japan. Parasitology. 1921;13:235–305. [Google Scholar]
- 30.Leidy J. The parasites of the termites. J Acad Nat Sci Philadelphia 2nd Ser. 1881;8:425–477. [Google Scholar]
- 31.Ludwig W, Strunk O, Westram R, et al. ARB: a software environment for sequence data. Nucleic Acids Res. 2004;32:1363–1371. doi: 10.1093/nar/gkh293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Messer AC, Lee MJ. Effect of chemical treatments on methane emission by the hindgut microbiota in the termite Zootermopsis angusticollis. Microb Ecol. 1989;18:275–284. doi: 10.1007/BF02075814. [DOI] [PubMed] [Google Scholar]
- 33.Mira A, Moran NA. Estimating population size and transmission bottlenecks in maternally transmitted endosymbiotic bacteria. Microb Ecol. 2002;44:137–143. doi: 10.1007/s00248-002-0012-9. [DOI] [PubMed] [Google Scholar]
- 34.Moran NA, McCutcheon JP, Nakabachi A. Genomics and evolution of heritable bacterial symbionts. Annu Rev Genet. 2008;42:165–190. doi: 10.1146/annurev.genet.41.110306.130119. [DOI] [PubMed] [Google Scholar]
- 35.Moreira D, von der Heyden S, Bass D, López-García P, Chao E, Cavalier-Smith T. Global eukaryote phylogeny: combined small- and large-subunit ribosomal DNA trees support monophyly of Rhizaria, Retaria and Excavata. Mol Phylogenet Evol. 2007;44:255–266. doi: 10.1016/j.ympev.2006.11.001. [DOI] [PubMed] [Google Scholar]
- 36.Ohkuma M, Sato T, Noda S, Ui S, Kudo T, Hongoh Y. The candidate phylum ‘Termite Group 1’ of bacteria: phylogenetic diversity, distribution, and endosymbiont members of various gut flagellated protists. FEMS Microbiol Ecol. 2007;60:467–476. doi: 10.1111/j.1574-6941.2007.00311.x. [DOI] [PubMed] [Google Scholar]
- 37.Ohkuma M, Noda S, Hongoh Y, Nalepa CA, Inoue T. Inheritance and diversification of symbiotic trichonymphid flagellates from a common ancestor of termites and the cockroach Cryptocercus. Proc R Soc B. 2009;276:239–245. doi: 10.1098/rspb.2008.1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ohkuma M, Brune A. Diversity, structure, and evolution of the termite gut microbial community. In: Bignell DE, Roisin Y, Lo N, editors. Biology of Termites: A Modern Synthesis. Springer; Dordrecht: 2011. pp. 413–438. [Google Scholar]
- 39.Sato T, Hongoh Y, Noda S, Hattori S, Ui S, Ohkuma M. Candidatus Desulfovibrio trichonymphae, a novel intracellular symbiont of the flagellate Trichonympha agilis in termite gut. Environ Microbiol. 2009;11:1007–1015. doi: 10.1111/j.1462-2920.2008.01827.x. [DOI] [PubMed] [Google Scholar]
- 40.Sato T, Kuwahara H, Fujita K, Noda S, Kihara K, Yamada A, Ohkuma M, Hongoh Y. Intranuclear verrucomicrobial symbionts and evidence of lateral gene transfer to the host protist in the termite gut. ISME J. 2014;8:1008–1019. doi: 10.1038/ismej.2013.222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sawada H, Takeuchi T, Matsuda I. Comparative analysis of Pseudomonas syringae pv. actinidiae and pv. phaseolicola based on phaseolotoxin-resistant ornithine carbamoyltransferase gene (argK) and 16S-23S rRNA intergenic spacer sequences. Appl Environ Microbiol. 1997;63:282–288. doi: 10.1128/aem.63.1.282-288.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30:1312–1313. doi: 10.1093/bioinformatics/btu033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Stingl U, Radek R, Yang H, Brune A. “Endomicrobia”: cytoplasmic symbionts of termite gut protozoa form a separate phylum of prokaryotes”. Appl Environ Microbiol. 2005;71:1473–1479. doi: 10.1128/AEM.71.3.1473-1479.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Strassert JFH, Köhler T, Wienemann THG, Ikeda-Ohtsubo W, Faivre N, Franckenberg S, Plarre R, Radek R, Brune A. ‘Candidatus Ancillula trichonymphae’, a novel lineage of endosymbiotic Actinobacteria in termite gut flagellates of the genus Trichonympha. Environ Microbiol. 2012;14:3259–3270. doi: 10.1111/1462-2920.12012. [DOI] [PubMed] [Google Scholar]
- 45.Tai V, James ER, Perlman SJ, Keeling PJ. Single-cell DNA barcoding using sequences from the small subunit rRNA and internal transcribed spacer region identifies new species of Trichonympha and Trichomitopsis from the hindgut of the termite Zootermopsis angusticollis. PLoS One. 2013;8:e58728. doi: 10.1371/journal.pone.0058728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tai V, James ER, Nalepa CA, Scheffrahn RH, Perlman SJ, Keeling PJ. The role of host phylogeny varies in shaping microbial diversity in the hindguts of lower termites. Appl Environ Microbiol. 2015;81:1059–1070. doi: 10.1128/AEM.02945-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Thompson CL, Vier R, Mikaelyan A, Wienemann T, Brune A. ‘Candidatus Arthromitus’ revised: segmented filamentous bacteria in arthropod guts are members of Lachnospiraceae. Environ Microbiol. 2012;14:1454–1465. doi: 10.1111/j.1462-2920.2012.02731.x. [DOI] [PubMed] [Google Scholar]
- 48.Trager W. The cultivation of a cellulose-digesting flagellate, Trichomonas termopsidis, and of certain other termite protozoa. Biol Bull. 1934;66:182–190. [Google Scholar]
- 49.van Orsouw NJ, Hogers RCJ, Janssen A, et al. Complexity reduction of polymorphic sequences (CRoPS™): A novel approach for large-scale polymorphism discovery in complex genomes. PLoS One. 2007;2:e1172. doi: 10.1371/journal.pone.0001172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wernegreen JJ. Genome evolution in bacterial endosymbionts of insects. Nat Rev Genet. 2002;3:850–861. doi: 10.1038/nrg931. [DOI] [PubMed] [Google Scholar]
- 51.Yamin MA. Flagellates of the orders Trichomonadida Kirby, Oxymonadida Grassé, and Hypermastigida Grassi & Foà reported from lower termites (Isoptera families Mastotermitidae, Kalotermitidae, Hodotermitidae, Termopsidae, Rhinotermitidae, and Serritermitidae) and from the wood-feeding roach Cryptocercus (Dictyoptera: Cryptocercidae) Sociobiology. 1979;4:1–119. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.