

Abstract
Even though mouse studies have various advantages, harvesting an adequate number of synovial mesenchymal stem cells (MSCs) is difficult in mice. We investigated whether the total yield of MSCs increased in synovium with inflammation in mice. Infrapatellar fat pads (IFPs) were harvested from 10 knees of 5 mice 3, 7, and 14 days after intraarticular injection of carrageenan. Ten IFPs were also harvested from untreated knees as a control. Seven days after initial plating, the total yield of cells was compared among the 4 groups (n = 4–6). The harvested cells were analyzed for multipotentiality and surface epitopes. Furthermore, knee synovitis was compared among the 4 groups in histology. The number of cells in the 3 and 7 days treated group was significantly higher than the other groups. The harvested cells had characteristics of MSCs. Synovitis in the 3 and 7 days treated groups was significantly severer than the other groups. There seemed to be a relationship between the synovitis score and the total yield of cells derived from IFPs. In mice, it became possible to increase the yield 50-fold by inducing inflammation. This method makes it possible to analyze the molecular mechanisms of cartilage regeneration of synovial MSCs in mice models. © 2014 The Authors. © 2014 The Authors. Journal of Orthopaedic Research published by Wiley Periodicals, Inc. on behalf of the Orthopaedic Research Society. J Orthop Res 33:246–253, 2015.
Keywords: mesenchymal stem cells, synovium, mouse, inflammation, infrapatellar fat pad
Mesenchymal stem cells (MSCs) play important roles in the repair of the human body as well as in normal homeostasis.1 MSCs were first isolated from bone marrow,2 and can be isolated from various mesenchymal tissues including synovium.3–6 Synovium is a thin layer of tissue that lines the joint space and covers a subsynovial tissue, such as infrapatellar fat pad (IFP), which fills the anterior knee compartment. We previously demonstrated that MSCs derived from IFP and those from fibrous synovium on the femur bone had comparable proliferation and chondrogenic potentials and were higher than those of MSCs derived from subcutaneous adipose tissue in humans and rats.7,8 These results indicate that IFP can be the source of synovial MSCs, and that synovial MSCs are an attractive cell source for cartilage and meniscus regeneration.
It would be useful to establish a method to prepare synovial MSCs in mice. Practically, the use of mice is a cost-effective and efficient way to accelerate research. Mice are small, have a short generation time and an accelerated lifespan, keeping the costs, space, and time required to perform research manageable. Mouse systems contribute to the elucidation of the molecular mechanisms of the phenomena for investigators because of the relatively easy gene manipulation in such cells, which is indispensable for the molecular analysis.
Our previous studies described that MSCs in synovial fluid obtained from anterior cruciate ligament (ACL) injury, meniscus injury, and osteoarthritis patients were much more in number than those obtained from healthy volunteers.9–11 These results indicate that many more MSCs exist in knees in unhealthy conditions, such as injury or inflammation. We hypothesized that IFPs in inflammatory conditions have many more MSCs than normal IFPs. Carrageenan, a polysaccharide made from a type of seaweed, is recognized as a cause for arthritis used as a model of acute inflammation.12–14 The aim of this study was to investigate whether the total yield of MSCs derived from IFP increased in inflammatory conditions induced by injection of carrageenan into the knee joints in mice.
MATERIALS AND METHODS
Animals
Animal care and all experiments were conducted in accordance with the guidelines of the animal committee of Tokyo Medical and Dental University. C57BL/6J mice were purchased from Oriental Yeast Co. Ltd (Tokyo, Japan). Mice were housed under a 12-h light–dark cycle and allowed food and water ad libitum.
Synovitis Induced by Carrageenan
Eight-week-old female C57BL/6J mice were used for this study. Mice were anesthetized by the inhalation of 5% isoflurane in oxygen. A small skin incision was created to expose the bilateral knee joints. After that, 31-gauge needles with Hamilton syringes (Hamilton Company, Ren, NV) were inserted through the lateral infrapatellar area toward the intercondylar space of the femur in a deep knee-flexed position. Then, λ-carrageenan (30 μg in 5 μl saline: Sigma–Aldrich, St. Louis, MO) was injected into bilateral knee joints for inducing synovitis.
Isolation and Culture of Mouse Cells From IFPs
Infrapatellar fat pads (IFPs) were harvested from 10 knees of 5 mice 3, 7, and 14 days after intraarticular injection of carrageenan. Ten IFPs were also harvested from untreated knees as a control group. Ten IFPs were minced and digested in a 1 mg/ml collagenase (Wako, Osaka, Japan) for 15 min and passed through a 40-μm filter (Becton Dickinson, Franklin Lakes, NJ).15 Dissociated cells and undigested IFPs were cultured together in a culture dish of 10-cm2 (Nalge Nunc International, Rochester, NY) in Dulbecco's modified Eagle's medium (DMEM: Wako) containing 10% fetal bovine serum (FBS: Invitrogen, Carlsbad, CA), 100 U/ml penicillin and 100 mg/ml streptomycin (Invitrogen) and 5 ng/ml recombinant human FGF-2 (BioVision, Milpitas, CA) at 37°C with 5% humidified CO2. Seven days after initial plating, nonadherent cells, debris and remaining IFPs were washed out with PBS and then the attached cells were harvested with 0.25% trypsin and 1 mM EDTA (Invitrogen), and the number of isolated cells was counted as passage 0 with hemocytometer (Fig. 1).
Figure 1.
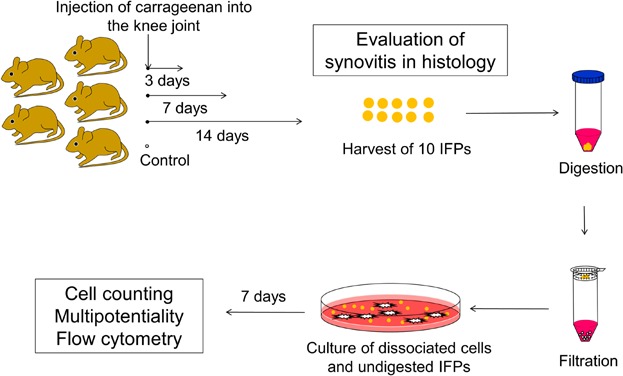
The study scheme. Carrageenan was injected into the knee joints of 5 mice (8-week old). Ten infrapatellar fat pads (IFPs) were harvested 3, 7, and 14 days after the injection. Ten IFPs were also harvested from untreated mice as a control group. Then IFPs were minced, digested, and filtered. Dissociated cells and undigested IFPs were cultured together. Seven days after plating, expanded cells were harvested for analyses. Whole knee joints were also examined for histological evaluation of synovitis 3, 7, and 14 days after the injection and of untreated synovium.
Colony-Forming Assay
One thousand cells in the 3-days treated group were plated and cultured for 7 days in 10-cm2 dishes. The dishes were then stained with 0.5% crystal violet (Wako) in 4% paraformaldehyde for 5 min.
Chondrogenesis
One hundred thousand cells in the 3-days treated group were placed in a 15-ml polypropylene tube (Becton Dickinson) and centrifuged at 450g for 10 min. The pellets were cultured at 37°C with 5% CO2 in 400 μl chondrogenesis medium that contained 1,000 ng/ml BMP-7 (Stryker Biotech, Hopkinton, MA) in high-glucose DMEM (Invitrogen) supplemented with 10 ng/ml transforming growth factor-β3 (R&D Systems, Minneapolis, MN), 100 nM dexamethasone, 50 ng/ml ascorbate-2-phosphate, 40 μg/ml proline, 100 μg/ml pyruvate (Sigma–Aldrich, St. Louis, MO), and 50 mg/ml ITS + Premix (Becton Dickinson). The medium was replaced every 3–4 days for 21 days. For microscopy, the pellets were embedded in paraffin, cut into 5-μm sections, and stained with toluidine blue and type II collagen antibody.16
Adipogenesis
One thousand cells in the 3-days treated group were plated in 10-cm2 dishes and cultured in complete medium for 7 days. The medium was then switched to an adipogenesis medium that consisted of complete medium supplemented with 100 nM dexamethasone, 0.5 mM isobutylmethylxanthine (Sigma–Aldrich), and 50 nM indomethacin (Wako) for an additional 14 days. The adipogenic cultures were fixed in 10% formalin and stained with fresh Oil Red-O (Sigma–Aldrich) solution.17
Calcification
One thousand cells in the 3-days treated group were plated in 10-cm2 dishes and cultured in complete medium for 7 days. The medium was switched to a calcification medium that consisted of complete medium supplemented with 1 nM dexamethasone (Sigma–Aldrich), 20 mM β-glycerol phosphate (Wako), and 50 μg/ml ascorbate-2-phosphate for an additional 14 days. These dishes were fixed in 10% formalin in PBS and stained with 40 mM alizarin red solution (pH 4.1; Sigma–Aldrich).18
Epitope Profile
One million cells were suspended in 500 µl FACS staining buffer (0.2% BSA fraction V and 0.09% Sodium azide in PBS) containing 20 µg/ml antibodies. After incubation for 60 min at 4°C, the cells were washed with PBS and resuspended in 1 ml FACS staining buffer for flow cytometric analysis. Fluorescein isothiocyanate (FITC), phycoerythrin (PE), peridinin chlorophyll protein-Cy5.5 (PerCP-Cy5.5), or Alexa Fluor 647-coupled antibodies against CD11b, CD44, CD45, CD73, CD90, and CD105 (Becton Dickinson) were used. For isotype controls, FITC-, PE-, PerCP-Cy5.5-, or Alexa Fluor 647-coupled nonspecific rat immunoglobulin G (IgG; Becton Dickinson) was substituted for the primary antibody. Cell fluorescence was evaluated by flow cytometry using a FACSVerse instrument (Becton Dickinson). The data were analyzed using FACSuite software (Becton Dickinson).
Histological Analyses of the Knee Joint
The knee joints were dissected, fixed in 4% PFA (paraformaldehyde; Sigma–Aldrich) in the fixed angle (30 degrees), decalcified in 20% EDTA (ethylenediaminetetraacetic acid in PBS, pH 7.4), and embedded in paraffin. Five-micrometer-thick sagittal sections of medial weight-bearing regions were prepared and stained with hematoxylin and eosin (H&E) for the histological evaluation of synovitis. The severity of synovitis was evaluated according to the synovitis scoring system described by Krenn et al.19 Compactly, synovitis was evaluated for enlargement of the synovial cell layer, density of the resident cells and inflammatory infiltrate, in which a full score was 9 and a lower score indicated no synovitis. This synovitis scores were calculated as the total of the synovium attached to the anterior and posterior horn of the meniscus in the knee joint as denoted in the right panel of Figure 5B.
Figure 5.
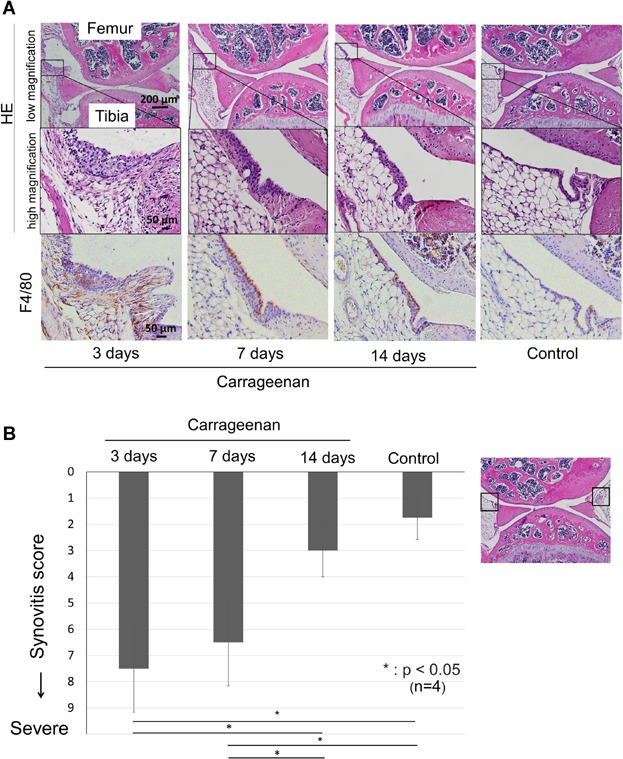
Histological analyses for synovitis induced by carrageenan. (A) Sagittal sections of mice knee joint, stained with H&E and F4/80 antibody for macrophage, 3, 7, and 14 days after injection of carrageenan and as a control. (B) Synovitis score. Severity of synovitis evaluated by scoring system. Average values with standard derivation are shown. For the synovitis score described by Krenn et al.,19 anterior and posterior synovium was evaluated as denoted in the right panel (*p < 0.05 by Tukey–Kramer test).
Immunostaining for Macrophage Infiltration Into Synovial Membrane
Mouse macrophage specific F4/80 staining was performed as described by Blom et al.20 Briefly, sections were deparaffinized, rinsed with PBS, and then incubated in 3% H2O2 in methanol for 30 min at room temperature to deactivate endogenous peroxidase. After blocking with normal rabbit serum (Vector Laboratories, CA) for 30 min, sections were incubated with rat anti-mouse monoclonal F4/80 antibody (1:2000 dilution: AbD Serotec, Raleigh, NC) at 4°C overnight. After extensive washing with PBS, signals were visualized by the Vectastain ABC kit (Vector Laboratories). Therefore, sections were counterstained using hematoxylin.
Statistical Analysis
Statistical analyses were performed using the Tukey–Kramer test and p values less than 0.05 were considered significant for Figures 2B and 5B. Correlation analysis was used for Figure 6.
Figure 2.
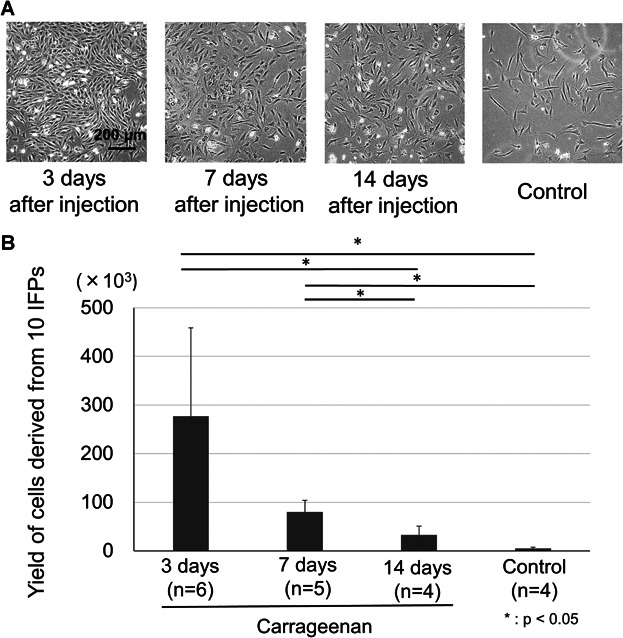
Morphology and the yield of cells derived from IFPs 7 days after the initial plating. (A) Appearance of expanded cells. (B) The yield of cells derived from 10 IFPs. Average values with standard derivation are shown (*p < 0.05 by Tukey–Kramer test).
Figure 6.
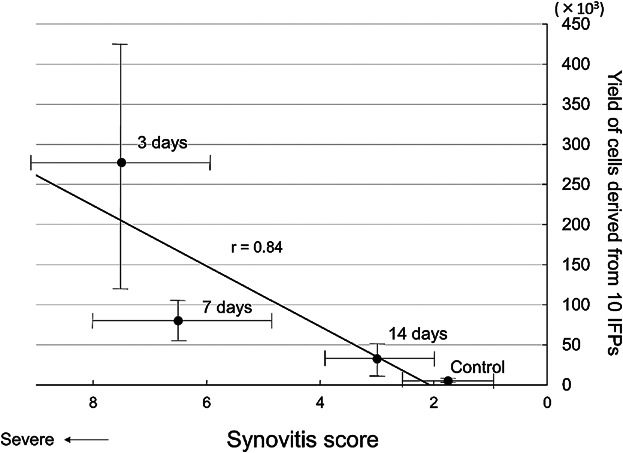
Relationship between the synovitis score and the total yield of cells derived from 10 IFPs. Average values with standard derivation are shown in each value.
RESULTS
Morphology and Yields of Cells Derived from IFPs
The dissociated cells and undigested IFPs were expanded for 7 days. They showed a spindle shape irrespective of carrageenan treated periods (Fig. 2A). The yield of cells in the 3-days treated group was significantly higher than in the control and in the 14-days treated groups (p < 0.05). The yield of cells in the 7-days treated group was less than in the 3-days treated group but still significantly higher than in the control and in the 14-days treated groups (p < 0.05) (Fig. 2B).
Characteristics as MSCs
The cells derived from IFPs treated with carrageenan for 3 days formed colonies (Fig. 3A). They differentiated into chondrocytes (Fig. 3B), adipocytes (Fig. 3C) and were calcified (Fig. 3D) when cultured in the differentiation medium. They expressed CD44, CD73, and CD90 at high rates; CD105 at a moderate rate; and did not express CD11b or CD45 (Fig. 4). The cells derived from IFPs had characteristics of MSCs.
Figure 3.
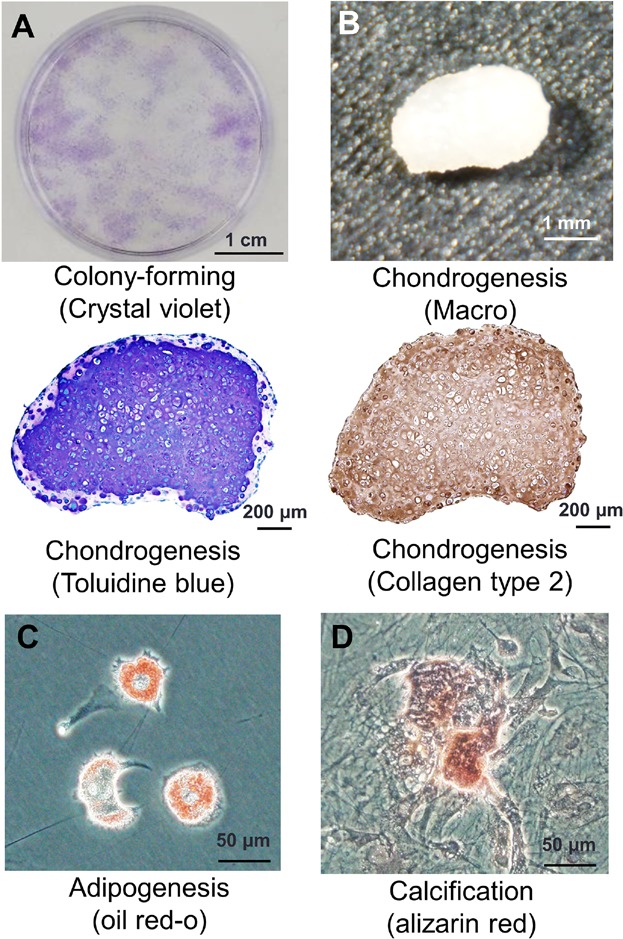
Colony-forming ability and multipotentiality of the cells derived from IFPs. (A) Representative cell colonies stained with crystal violet. (B) Cartilage pellet and its histological sections stained with toluidine blue and type 2 collagen. (C) Adipocytes stained with Oil Red-O. (D) Calcification stained with alizarin red.
Figure 4.
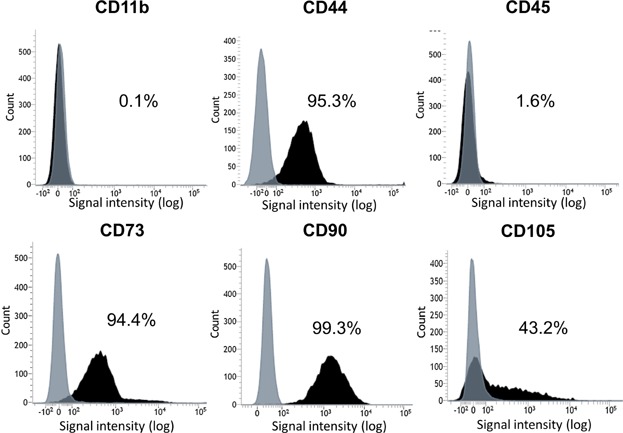
Surface epitopes of the cells expanded from IFPs. Representative histograms are shown as black and isotype controls are shown as gray.
Histological Analyses of Synovitis Induced by Carrageenan
The synovial membrane thickened and its cellularity increased 3 days after intraarticular injection of carrageenan, thickened synovial membrane became dense at 7 days, then its cellularity decreased at 14 days (Fig. 5A). There were numerous F4/80-positive macrophages in the lining cell layer and subsynovial fat layer at 3 days. F4/80-positive macrophages were localized to the lining cell layer at 7 and 14 days. Krenn's synovitis score19 showed that synovitis in the 3 and 7 days treated group were severer than in the other groups (Fig. 5B).
Relationship between Synovitis and Yield of Cells
To analyze the relationship between synovitis and the yield of cells, we plotted the synovitis score at the X-axis and the yield of cells at the Y-axis. There seemed to be a relationship between the synovitis score and the total yield of cells derived from 10 IFPs. The correlation coefficient was 0.84 (Fig. 6).
DISCUSSION
In this study, we demonstrated that the cells derived from mouse IFPs adhered to culture dishes, formed cell colonies, differentiated into chondrocytes and adipocytes, and were calcified. The cells also expressed CD44, CD73, and CD90 at a high rate, CD105 at a moderate rate, and did not express CD11b or CD45. These are consistent with the definition of MSCs by the position statement of the International Society for Cellular Therapy.21
There were some reports in which mouse synovial cells were used for experiments.22–25 However, it is not easy to prepare mouse synovial cells because mouse synovial tissue is very small, and these reports did not mention the detailed methods concerning where to harvest synovium, how many mice were used, how to expand etc. We recently reported the detailed method to prepare mouse synovial MSCs.15 In this current study, we were able to obtain 50 times more mouse synovial MSCs by inducing inflammation.
We induced synovitis by intraarticular injection of carrageenan. Carrageenan is a sulphated mucopolysaccharide extracted from the seaweeds Chondrus spp. and Gigartina spp., commonly known as Irish moss or carrageen moss from red Scottish seaweed.26 These seaweeds were known for its remarkable capacity to stimulate local inflammation dominated by intense macrophage aggregation and by fibroblastic proliferation.27 Carrageenan-induced arthritis is a well-established experimental animal model used to investigate inflammation-mediated articular cartilage degradation in rats, mice and rabbits with a localized synovial inflammatory response, which is indicated by synovial hyperplasia and macrophage accumulation.28,29
In our study, the total yield of MSCs from synovium increased over a 3 days period along with synovitis, in which a great number of macrophages accumulated. Anton et al. demonstrated that MSCs derived from bone marrow were migrated by IL-8, CCL-2 and CCL-5 secreted by macrophage in migration assay.30 These suggest that inflammation increases the yield of mouse synovial MSCs and that some cytokines secreted by macrophages induce migration of MSCs.
We demonstrated that the correlation coefficient was 0.84 between the synovitis score and the total yield of synovial MSCs. Correctly, we should analyze a relationship between synovitis and the yield of MSCs from the same individual mouse. However, it was difficult practically because once IFP was harvested from the knee in order to isolate cells, and the same knee was not unusable for evaluation of synovitis in histology. Therefore, we analyzed with different individual mice. In these mice, mice strains, age, sex and experimental conditions were unified. Generally, the correlation coefficient in the analysis of the same individuals is higher than that of the different individuals. In this study, although different individual mice were used for the synovitis score and for the yield of MSCs, the correlation coefficient for them was relatively high. Consequently, we concluded that total yield of MSCs from synovium is correlated with synovitis.
It was unclear whether the inflammation affects the function of MSCs derived from IFPs. It would be interesting to compare surface epitopes and multipotentiality among the 3, 7, and 14 days treated and control groups. It was effectively impossible to perform these experiments because yields of MSCs at passage 0 in the 7 and 14 days treated and control groups were too low. Previously, we compared MSCs derived from synovial fluid collected just before and 6 days after anterior cruciate ligament operations in humans for multipotentiality and surface epitopes. Though synovial fluid before the operation was transparent and synovial fluid 6 days after the operation was bloody, suggesting this contained a great number of inflammatory cells, there were no differences of properties as MSCs between two populations.9 This supports the claim that inflammation does not affect function of MSCs, but a different result was also reported. Mohanty et al. compared MSCs derived from one marrow of wild mice and IL-1 antagonist knockout mice showing similar phenotypes to rheumatoid arthritis. MSCs derived from mice with rheumatoid arthritis had greater colony-forming, adipocyte differentiation, and osteoblast differentiation capacity than those from normal mice.31
In this study, carrageenan-induced inflammation resulted in a good yield of MSCs. Though there is a possibility that these MSCs had characteristics of tumorigenesis, we do not think so for the following two reasons. Firstly, the morphologies of the cells derived from IFPs appeared to be the same regardless of the presence or absence of inflammation induced by carrageenan. Secondly, there were no obvious differences of proliferation of the MSCs at passage 1 among 4 groups including the control, 3, 7, and 14 days treated groups (data not shown). There seem to be no reports demonstrating that MSCs acquire tumorigenesis at passage 0.
Our purpose was to investigate whether the total yield of MSCs increased in synovium with inflammation in mice. We estimated that the peak of the inflammations induced by intraarticular injection of carrageenan was around the third day after injection according to the previous papers. Bang et al. and Gong et al. demonstrated that the peaks of inflammation of hind-paw or knees were the same from 1 to 4 days after the local injection of carrageenan.32,33 Silván et al. revealed that the peak of inflammation of hind-paw was at 3 days after the local injection of carrageenan.34 Rocha et al. showed that the peaks of inflammation of hind-paw were at 2 and 3 days after the local injection of carrageenan.35 Kim reported that the peaks of inflammation of hind-paw were at 3 and 4 days after the local injection of carrageenan.36 These indicated that the peak of inflammation of IFPs was around 3 days after the intraarticular injection of carrageenan.
Tissue regeneration techniques using MSCs have been considered as new treatments for tissue repair.37 In the field of cartilage regeneration, synovial MSC is a promising tool because of a higher chondrogenic potential compared to MSCs derived from other tissues.8,38 However, the detailed mechanisms in cartilage differentiation of synovial MSCs had been unclear. It is possible to collect plenty of synovial MSCs in mice by our method, which could lead to the elucidation of the molecular mechanism of cartilage differentiation capacity of synovial MSCs because mouse studies have a variety of advantages such as the relative ease of use in gene manipulation. In this study, we used C57BL/6 mice because most of genetically modified mice have a genetic background of C57BL/6, the most widely used strain.
It has been difficult to obtain a sufficient number of synovial MSCs in mice but it was possible to increase the yield more than 50 fold by inducing inflammation. The total yield of MSCs from synovium appeared to be correlated with inflammation in synovium. This method makes it possible for us to analyze the molecular mechanisms of cartilage regeneration of synovial MSCs in mice models.
Acknowledgments
We would like to thank Miyoko Ojima for her help with histology, Hisako Katano for her attractive illustrations and Izumi Nakagawa for managing our laboratory.
REFERENCES
- 1.De Bari C, Dell'accio F. Mesenchymal stem cells in rheumatology: a regenerative approach to joint repair. Clin Sci (Lond) 2007;113:339–348. doi: 10.1042/CS20070126. [DOI] [PubMed] [Google Scholar]
- 2.Pittenger MF, Mackay AM, Beck SC. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143–147. doi: 10.1126/science.284.5411.143. et al. [DOI] [PubMed] [Google Scholar]
- 3.Asakura A, Komaki M, Rudnicki M. Muscle satellite cells are multipotential stem cells that exhibit myogenic, osteogenic, and adipogenic differentiation. Differentiation. 2001;68:245–253. doi: 10.1046/j.1432-0436.2001.680412.x. [DOI] [PubMed] [Google Scholar]
- 4.De Bari C, Dell'Accio F, Tylzanowski P, Luyten FP. Multipotent mesenchymal stem cells from adult human synovial membrane. Arthritis Rheum. 2001;44:1928–1942. doi: 10.1002/1529-0131(200108)44:8<1928::AID-ART331>3.0.CO;2-P. [DOI] [PubMed] [Google Scholar]
- 5.Segawa Y, Muneta T, Makino H. Mesenchymal stem cells derived from synovium, meniscus, anterior cruciate ligament, and articular chondrocytes share similar gene expression profiles. J Orthop Res. 2009;27:435–441. doi: 10.1002/jor.20786. et al. [DOI] [PubMed] [Google Scholar]
- 6.Zuk PA, Zhu M, Mizuno H. Multilineage cells from human adipose tissue: implications for cell-based therapies. Tissue Eng. 2001;7:211–228. doi: 10.1089/107632701300062859. et al. [DOI] [PubMed] [Google Scholar]
- 7.Mochizuki T, Muneta T, Sakaguchi Y. Higher chondrogenic potential of fibrous synovium- and adipose synovium-derived cells compared with subcutaneous fat-derived cells: distinguishing properties of mesenchymal stem cells in humans. Arthritis Rheum. 2006;54:843–853. doi: 10.1002/art.21651. et al. [DOI] [PubMed] [Google Scholar]
- 8.Yoshimura H, Muneta T, Nimura A. Comparison of rat mesenchymal stem cells derived from bone marrow, synovium, periosteum, adipose tissue, and muscle. Cell Tissue Res. 2007;327:449–462. doi: 10.1007/s00441-006-0308-z. et al. [DOI] [PubMed] [Google Scholar]
- 9.Morito T, Muneta T, Hara K. Synovial fluid-derived mesenchymal stem cells increase after intra-articular ligament injury in humans. Rheumatology (Oxford) 2008;47:1137–1143. doi: 10.1093/rheumatology/ken114. et al. [DOI] [PubMed] [Google Scholar]
- 10.Sekiya I, Ojima M, Suzuki S. Human mesenchymal stem cells in synovial fluid increase in the knee with degenerated cartilage and osteoarthritis. J Orthop Res. 2012;30:943–949. doi: 10.1002/jor.22029. et al. [DOI] [PubMed] [Google Scholar]
- 11.Matsukura Y, Muneta T, Tsuji K. Mesenchymal Stem Cells in Synovial Fluid Increase After Meniscus Injury. Clin Orthop Relat Res. 2014;472:1357–1364. doi: 10.1007/s11999-013-3418-4. et al. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hansra P, Moran EL, Fornasier VL, Bogoch ER. Carrageenan-induced arthritis in the rat. Inflammation. 2000;24:141–155. doi: 10.1023/a:1007033610430. [DOI] [PubMed] [Google Scholar]
- 13.Morris CJ. Carrageenan-induced paw edema in the rat and mouse. Methods Mol Biol. 2003;225:115–121. doi: 10.1385/1-59259-374-7:115. [DOI] [PubMed] [Google Scholar]
- 14.Ekundi-Valentim E, Santos KT, Camargo EA. Differing effects of exogenous and endogenous hydrogen sulphide in carrageenan-induced knee joint synovitis in the rat. Br J Pharmacol. 2010;159:1463–1474. doi: 10.1111/j.1476-5381.2010.00640.x. et al. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Futami I, Ishijima M, Kaneko H. Isolation and characterization of multipotential mesenchymal cells from the mouse synovium. PLoS ONE. 2012;7:e45517. doi: 10.1371/journal.pone.0045517. et al. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sekiya I, Colter DC, Prockop DJ. BMP-6 enhances chondrogenesis in a subpopulation of human marrow stromal cells. Biochem Biophys Res Commun. 2001;284:411–418. doi: 10.1006/bbrc.2001.4898. [DOI] [PubMed] [Google Scholar]
- 17.Sekiya I, Larson BL, Vuoristo JT. Adipogenic differentiation of human adult stem cells from bone marrow stroma (MSCs) J Bone Miner Res. 2004;19:256–264. doi: 10.1359/JBMR.0301220. et al. [DOI] [PubMed] [Google Scholar]
- 18.Sakaguchi Y, Sekiya I, Yagishita K. Suspended cells from trabecular bone by collagenase digestion become virtually identical to mesenchymal stem cells obtained from marrow aspirates. Blood. 2004;104:2728–2735. doi: 10.1182/blood-2003-12-4452. et al. [DOI] [PubMed] [Google Scholar]
- 19.Krenn V, Morawietz L, Burmester GR. Synovitis score: discrimination between chronic low-grade and high-grade synovitis. Histopathology. 2006;49:358–364. doi: 10.1111/j.1365-2559.2006.02508.x. et al. [DOI] [PubMed] [Google Scholar]
- 20.Blom AB, van Lent PL, Libregts S. Crucial role of macrophages in matrix metalloproteinase-mediated cartilage destruction during experimental osteoarthritis: involvement of matrix metalloproteinase 3. Arthritis Rheum. 2007;56:147–157. doi: 10.1002/art.22337. et al. [DOI] [PubMed] [Google Scholar]
- 21.Dominici M, Le Blanc K, Mueller I. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–317. doi: 10.1080/14653240600855905. et al. [DOI] [PubMed] [Google Scholar]
- 22.Langdon C, Kerr C, Hassen M. Murine oncostatin M stimulates mouse synovial fibroblasts in vitro and induces inflammation and destruction in mouse joints in vivo. Am J Pathol. 2000;157:1187–1196. doi: 10.1016/S0002-9440(10)64634-2. et al. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dulos J, Verbraak E, Bagchus WM. Severity of murine collagen-induced arthritis correlates with increased CYP7B activity: enhancement of dehydroepiandrosterone metabolism by interleukin-1beta. Arthritis Rheum. 2004;50:3346–3353. doi: 10.1002/art.20509. et al. [DOI] [PubMed] [Google Scholar]
- 24.Rhee DK, Marcelino J, Baker M. The secreted glycoprotein lubricin protects cartilage surfaces and inhibits synovial cell overgrowth. J Clin Invest. 2005;115:622–631. doi: 10.1172/JCI200522263. et al. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Waldburger JM, Palmer G, Seemayer C. Autoimmunity and inflammation are independent of class II transactivator type PIV-dependent class II major histocompatibility complex expression in peripheral tissues during collagen-induced arthritis. Arthritis Rheum. 2011;63:3354–3363. doi: 10.1002/art.30522. et al. [DOI] [PubMed] [Google Scholar]
- 26.Michel G, Nyval-Collen P, Barbeyron T. Bioconversion of red seaweed galactans: a focus on bacterial agarases and carrageenases. Appl Microbiol Biotechnol. 2006;71:23–33. doi: 10.1007/s00253-006-0377-7. et al. [DOI] [PubMed] [Google Scholar]
- 27.Gordon JL, MacIntyre DE, McMillan RM. Stimulation of platelets and macrophages by carrageenin [proceedings] Br J Pharmacol. 1977;61:140P–141P. [PMC free article] [PubMed] [Google Scholar]
- 28.Santer V, Sriratana A, Lowther DA. Carrageenin-induced arthritis: V. A morphologic study of the development of inflammation in acute arthritis. Semin Arthritis Rheum. 1983;13:160–168. doi: 10.1016/0049-0172(83)90002-1. [DOI] [PubMed] [Google Scholar]
- 29.Lowther DA, Gillard GC. Carrageenin-induced arthritis. I. The effect of intraarticular carrageenin on the chemical composition of articular cartilage. Arthritis Rheum. 1976;19:769–776. doi: 10.1002/1529-0131(197607/08)19:4<769::aid-art1780190419>3.0.co;2-v. [DOI] [PubMed] [Google Scholar]
- 30.Anton K, Banerjee D, Glod J. Macrophage-associated mesenchymal stem cells assume an activated, migratory, pro-inflammatory phenotype with increased IL-6 and CXCL10 secretion. PLoS ONE. 2012;7:e35036. doi: 10.1371/journal.pone.0035036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mohanty ST, Kottam L, Gambardella A. Alterations in the self-renewal and differentiation ability of bone marrow mesenchymal stem cells in a mouse model of rheumatoid arthritis. Arthritis Res Ther. 2010;12:R149. doi: 10.1186/ar3098. et al. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bang JS, Oh da H, Choi HM. Anti-inflammatory and antiarthritic effects of piperine in human interleukin 1beta-stimulated fibroblast-like synoviocytes and in rat arthritis models. Arthritis Res Ther. 2009;11:R49. doi: 10.1186/ar2662. et al. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Gong D, Chu W, Jiang L. Effect of Fucoxanthin Alone and in Combination with D-glucosamine Hydrochloride on Carrageenan/kaolin-induced Experimental Arthritis in Rats. Phytother Res. 2014;28:1054–1063. doi: 10.1002/ptr.5093. et al. [DOI] [PubMed] [Google Scholar]
- 34.Silván AM, Abad MJ, Bermejo P, Villar AM. Adjuvant-carrageenan-induced inflammation in mice. Gen Pharmacol. 1997;29:665–669. doi: 10.1016/s0306-3623(96)00561-7. [DOI] [PubMed] [Google Scholar]
- 35.Rocha AC, Fernandes ES, Quintão NL. Relevance of tumour necrosis factor-alpha for the inflammatory and nociceptive responses evoked by carrageenan in the mouse paw. Br J Pharmacol. 2006;148:688–695. doi: 10.1038/sj.bjp.0706775. et al. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kim KS, Kim MH, Yeom M. Arthritic disease is more severe in older rats in a kaolin/carrageenan-induced arthritis model. Rheumatol Int. 2012;32:3875–3879. doi: 10.1007/s00296-011-2286-z. et al. [DOI] [PubMed] [Google Scholar]
- 37.Keating A. Mesenchymal stromal cells: new directions. Cell Stem Cell. 2012;10:709–716. doi: 10.1016/j.stem.2012.05.015. [DOI] [PubMed] [Google Scholar]
- 38.Sakaguchi Y, Sekiya I, Yagishita K, Muneta T. Comparison of human stem cells derived from various mesenchymal tissues: superiority of synovium as a cell source. Arthritis Rheum. 2005;52:2521–2529. doi: 10.1002/art.21212. [DOI] [PubMed] [Google Scholar]