

Abstract
Advanced glycation end-products (AGEs) initiate cellular inflammation and contribute to cardiovascular disease in the elderly. AGE can be inhibited by Alagebrium (ALT), an AGE cross-link breaker. Moreover, the beneficial effects of exercise on aging are well recognized. Thus, we investigated the effects of ALT and exercise (Ex) on cardiovascular function in a rat aging model. Compared to young (Y) rats, in sedentary old (O) rats, end-systolic elastance (Ees) decreased (0.9±0.2 vs 1.7±0.4 mm Hg/μL, P<0.05), dP/dtmax was attenuated (6054±685 vs 9540±939 mm Hg/s, P<0.05), ventricular compliance (end-diastolic pressure–volume relationship (EDPVR)) was impaired (1.4±0.2 vs 0.5±0.4 mm Hg/μL, P<0.05) and diastolic relaxation time (tau) was prolonged (21±3 vs 14±2 ms, P<0.05). In old rats, combined ALT+Ex (4 weeks) increased dP/dtmax and Ees (8945±665 vs 6054±685 mm Hg/s, and 1.5±0.2 vs 0.9±0.2 respectively, O with ALT+Ex vs O, P<0.05 for both). Diastolic function (exponential power of EDPVR and tau) was also substantially improved by treatment with Alt+Ex in old rats (0.4±0.1 vs 0.9±0.2 and 16±2 vs 21±3 ms, respectively, O with ALT+EX vs O, P<0.05 for both). Pulse wave velocity (PWV) was increased in old rats (7.0±0.7 vs 3.8± 0.3 ms, O vs Y, P<0.01). Both ALT and Ex alone decreased PWV in old rats but the combination decreased PWV to levels observed in young (4.6±0.5 vs 3.8±0.3 ms, O with ALT+Ex vs Y, NS). These results suggest that prevention of the formation of new AGEs (with exercise) and breakdown of already formed AGEs (with ALT) may represent a therapeutic strategy for age-related ventricular and vascular stiffness.
Keywords: AGE, Ventricular–vascular coupling, Alagebrium, Exercise, Advanced glycation end-products
1. Introduction
Age related cardiovascular disease results in part from the formation of advanced glycation end-products (AGEs). These accrue as a result of the irreversible non-enzymatic glycosylation and modification of proteins including collagen, elastin, and other functional proteins (Zieman and Kass, 2004). Cell surface receptors for AGEs, so called “RAGEs,” have been identified and coupled to pathways that initiate cellular inflammation and tissue damage. Interaction of AGE ligands with RAGE enhances receptor expression initiating a positive feedback loop (for review see Ulrich and Cerami, 2001). AGEs increase tensile stiffness and make these molecules less degradable by metalloproteinases leading to tissue accumulation. Although AGE-formation is increased in hyperglycemic conditions, it also occurs under normoglycemic conditions and is time-dependent, suggesting that it could be important in age-related vascular changes. Thus, AGEs may ultimately contribute to the ventricular and vascular stiffness seen in the aging population (Kass et al., 2001). The recent development of agents, for example Alagebrium, that can break carbon– carbon bonds and impair AGE-formation has important implications for treating age related cardiovascular dysfunction (Kass, 2003). Studies have demonstrated enhanced cardiac compliance, and cardiac output in aged dogs (Asif et al., 2000) and improved arterial compliance (Kass et al., 2001) following treatment with Alagebrium.
Reduced physical activity is well recognized to result in cardiovascular deconditioning (McGuire et al., 2001). Sedentary aging is associated with stiffening of the heart and vasculature, while life-long exercise training prevents these developments (Arbab-Zadeh et al., 2004). However, even 1 year of exercise training starting later in life (after the age of 65) does not seem to be effective at improving either arterial (Shibata and Levine, 2012) or cardiac (Fujimoto et al., 2010) stiffness in human studies. This could be due to the fact that in older individuals AGE products are already present in substantial quantities. Thus, it is possible that one mechanism by which exercise prevents stiffening of the heart and blood vessels is by limiting AGE accumulation within cardiovascular structures. The formation of AGE products is not spontaneously reversible and it is plausible that concurrent pharmacological reversal is necessary to allow full expression of the exercise response.
Ventricular–vascular coupling (VVC), or the interaction between ventricular contraction and systemic arterial vessels, is a central determinant of net cardiovascular performance (Kass, 2002). One of the seminal features of the aging vasculature is an increase in arterial stiffness or decreased arterial compliance (CA). Indices of vascular stiffness (increased pulse pressure, increased PWV) are important risk factors for cardiovascular disease, specifically in the elderly (Mitchell et al., 2010). This has led to a search for agents that increase CA and reduce pressure pulsatility such as novel AGE crosslink breakers (Kass et al., 2001). Although multiple pathways are dysregulated in aging blood vessels (Fitch et al., 2001; Wang et al., 2000), including impaired endothelial vasodilator function and NO signaling, increased oxidative stress, upregulation of matrix metalloproteinases (MMP-2) (Wang and Lakatta, 2002; Wang et al., 2003) and monocyte chemotactic factor and its receptor (on vascular smooth muscle cells) (Spinetti et al., 2004), AGE-accumulation may be critically important in limiting the cardiovascular response to exercise in older adults. As a corollary, AGE cross-link breakdown could enhance the response to exercise in cardiovascular structures. We therefore hypothesized that ALT, a novel AGE crosslink breaker, acts synergistically with exercise and improves the parameters of vascular and ventricular stiffness to a greater degree than either intervention alone. In this study, we determined the effect of ALT and exercise alone, or in combination, on cardiovascular parameters including parameters of vascular stiffness and diastolic and systolic functions.
2. Methods
2.1. Study design
Fifteen young (6 months) and 60 old (22–24 months) male Fisher 344 (F344) rats were obtained from the National Institute on Aging. The rats were housed three animals or less per cage, with controlled temperature and light conditions. The animals were fed and received water ad libitum. All experimental procedures were approved by the Institutional Animal Care and Use Committee of Johns Hopkins University School of Medicine. Fisher rats at 24months develop an increase in cardiac stiffness and a decrease in contractile reserve (Pacher et al., 2004). Old rats were randomized to receive placebo, Alagebrium chloride from Alteon Corp. (ALT-711) at 1 mg/kg/day body weight by IP injection, exercise (40 min/day, 5 days/week), or a combination of ALT and exercise for 4 weeks. The young control rats remained sedentary. Control (Y) and experimental groups each consisted of 7–8 F344 rats. ALT dosage was determined from a review of previous studies, where 1 mg/kg/day was safely administered to rats, monkeys, and dogs to yield significant restorative effects of ALT on cardiovascular and diabetic pathophysiology (Liu et al., 2003; Susic et al., 2004; Vaitkevicius et al., 2001;Wolffenbuttel et al., 1998)
2.2. Exercise regimen
Rats were exercised daily on a rodent treadmill (Columbus Instruments), for a period of 40min per day at about 50% of the maximal exercise tolerance (Hoydal et al., 2007). Rats were acclimatized to the treadmill for 1 week by walking daily for 40 min starting at a speed of 5 m/min with increments of 1 m/min/day. After this period of acclimatization rats were exercised daily for 4 weeks at 12m/min at an incline of 5° for the period of the study.
2.3. Pulse wave velocity measurement
The rats were anesthetized in a closed chamber with isoflurane. Anesthesia was maintained with 1.5% isoflurane (in 100% O2) by mask. Rats were positioned supine on a temperature controlled printed circuit board (THM100, Indus Instruments, Houston, TX) with legs and arms taped to incorporate electrocardiogram electrodes. Body temperature was maintained at 37 °C.
Doppler spectrograms of aortic outflow were acquired with a 2-mm diameter, 10-MHz pulsed Doppler probe (DSPW, Indus Instruments, Houston, TX). Thoracic aortic outflow was acquired at a depth of 4–6 mm with the probe placed to the right of the sternum. Abdominal aortic flow was captured at a depth of 4–5 mm. The nature of these waveforms was verified by comparison to characteristic aortic outflow waveforms. The distance separating the probe locations was also measured. Aortic PWV is calculated as the quotient of the separation distance and the time difference between pulse arrivals, with respect to the R-peak of the ECG. Data analysis of Doppler and ECG signals was done off-line using DSPW software from Indus Instruments.
2.4. Pressure–volume loops
Closed-chest pressure–volume (P–V) relationships were measured using a combined pressure–conductance Millar catheter (SPR- 838, 2.0 Fr) and the Aria combined pressure–conductance system (Millar). Rats were anesthetized with ketamine/xylazine (120mg/kg, 12 mg/kg; ketamine, xylazine, respectively), and a tracheostomy was performed for mechanical ventilation. The conductance catheter was inserted through the right carotid artery and advanced into the left ventricle (LV). A polyethylene catheter was inserted into the femoral artery for measurement of arterial pressure. In addition, a catheter was inserted into the femoral vein for administration of albumin solution. A 2 Fr Fogarty catheter was advanced through the femoral vein into the IVC. Left ventricular P–V relations were measured through preload reduction by transiently inflating the balloon. Hemodynamic measurements were performed as previously described in detail (Barouch et al., 2002; Varghese et al., 2000). Cardiac preload was determined using left ventricular end-diastolic volume (EDV) and pressure (EDP). Myocardial contractility was determined as the peak rate of rise in LV pressure (+dP/dtmax) divided by instantaneous pressure (dP/dtmax-IP). The load-independent end-systolic elastance (Ees) calculated from the linear end-systolic pressure–volume relationship (ESPVR) fitted in the pressure range of 40 to 110 mm Hg. The effective arterial elastance, Ea, was calculated as the ratio of the left ventricular end-systolic pressure and the stroke volume. Ventricular to arterial coupling was assessed as the Ea/Ees ratio. Active diastolic relaxation was measured by peak −dP/dt (−dP/dtmin), and the time constant of ventricular relaxation (tau). The latter is derived from the mono-exponential equation describing the rate of fall of ventricular pressure (regression of dP/dt vs pressure), and is a well validated measure of the rate of diastolic relaxation (Gilbert and Glantz, 1989). Mean arterial pressure (MAP) was calculated from peak systolic pressure (Pmax) and pressure at dP/dtmax (P@dP/dtmax) with the equation MAP=P@dP/dtmax+(Pmax−P@dP/dtmax)/3, which estimates the center of gravity of the analog triangular pressure waveform in each contraction. The total peripheral resistance index (TPRI) is derived from the ratio of MAP and cardiac output index (COI): TPRI=MAP/COI. Ejection fraction (EF) is derived from stroke volume (SV) and EDV as follows: EF=SV/EDV. The end-diastolic P–V relationship (EDPVR), an assessment of diastolic compliance, was determined from the end-diastolic P–V data obtained utilizing IVC occlusion. A nonlinear fit was used to characterize EDPVR, yielding the curve: EDP=C*e(K*EDV) where C and K are real value constants. End-diastolic pressure increases exponentially with respect to rate constant K as age- and treatment-dependent end-diastolic volume increases with higher loads in consecutive contractile cycles. K values depend on the instantaneous rise in pressure (P) over the increase in volume (V), ΔP/ΔV=1/compliance, and could be used to approximate ventricular stiffness and diastolic function. Characterizing the exponential power of the nonlinear fit for EDPVR, these K values are shown in Fig. 2A.
Fig. 2.
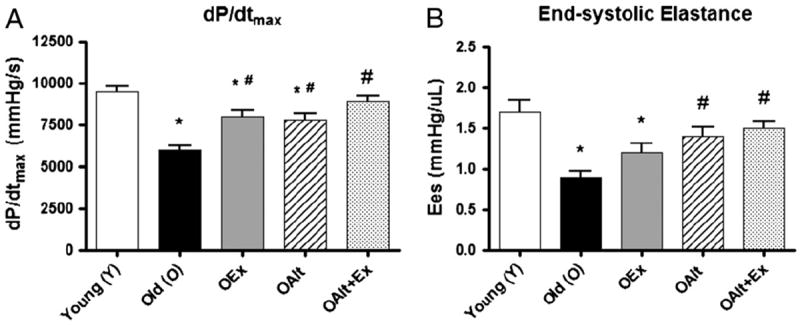
Parameters of myocardial contractility including the load-independent measurement end-systolic elastance (Ees) are restored by exercise and Alagebrium: aging is associated with a large decrease in measures of contractility as seen in representative P–V loop (Fig. 2). Both OALT and OEx, as well as OALT + Ex significantly improve contractility as measured by dP/dtmax. With regard to ESPVR, there is a significant improvement in this parameter in OALT + Ex. (*) P<0.05 vs. Y, (#) P<0.05 vs. O. N-values are the same as those specified in the table legend.
2.5. Statistical analysis
All data are presented asmeans±SD. Experiments were performed in either young, old, old ALT, old exercise or old ALT plus exercise animals, without conducting longitudinal experiments. One way ANOVA followed by a Student–Newman–Keuls post-test was used for group comparisons. A value of P<0.05 was considered significant.
3. Results
Hemodynamic parameters in young (Y) and old (O) rats, and the modulating influence of ALT and Ex, individually and in combination in O rats are outlined in Table 1. A total of two animals died during the study (one old in the ALT treated group and one old in the exercise group).
Table 1.
Hemodynamic parameters in young (Y) and old (O) rats and the modulating effects of ALT and EX individually and in combination in O rats.
| Parameters | Y (n = 8) | O (n = 8) | OAlt (n = 7) | OEx (n = 7) | OAlt+ Ex (n =8) |
|---|---|---|---|---|---|
| HR (bpm) | 340±30 | 280±30* | 290±20* | 300±30* | 320±30 |
| MAP (mm Hg) | 97±9 | 80±7* | 88±5* # | 92 ±3# | 93±9# |
| COI (mL/min/100 g) | 11±2 | 6.7±1.2* | 8.9±1.9# | 8.3±1.7* | 10.9±1.7# |
| SV (μL) | 140±11 | 93±9* | 114±7* # | 109±8* # | 130±6# |
| EF (%) | 68±9 | 33±6* | 44±5* # | 41 ±7* # | 56±6* # |
| ESP (mm Hg) | 110±7 | 90±8* | 97±3* # | 98±7* # | 103±3# |
| dP/dtmax-IP (s−1) | 108±17 | 80±20* | 90±14 | 91±13 | 103±19 |
| −dP/dtmin (mm Hg/s) | 6900±600 | 5000±1000* | 5100±900* | 4800±900* | 5600±1100* |
| Ees (mm Hg/μL) | 1.7±0.4 | 0.9±0.2* | 1.4±0.3# | 1.2±0.3 | 1.6±0.3# |
| Ea (mm Hg/μL) | 1.7±0.4 | 0.8±0.1* | 1.1±0.1 | 1.1±0.3 | 1.5±0.3# |
| Ea/Ees | 1.0 | 0.89 | 0.80 | 0.88 | 0.96 |
| PRSW (mm Hg) | 104±12 | 42±5* | 61±6* # | 57 ±10* # | 82±8* # |
| TPRI (mm Hg*mL/min/100 g) | 8.8 ±1.3 | 12.2±1.9* | 11±2 | 12 ±2* | 9±2# |
| ESV (μL) | 66.1±17.9 | 190.7±20.3* | 145.0±24.4* # | 160.0 ±34.8* # | 102±23.1* # |
| EDV (μDV) | 206±17 | 280±20* | 260±20* | 270 ±30* | 230±20# |
| EDP (mm Hg) | 6.9±1.6 | 11.1±1.4* | 9.0±1.1* # | 8.2±1.0# | 7.2±1.6# |
| Weight (g) | 334±12 | 418±48 | 382±78 | 400 ±80 | 390±55 |
F344 rats: young (Y), 6 months, n=8; old (O), 22–24-months, n=8; old rats undergoing exercise regimen (OEx), n=7; old rats undergoing Alagebrium Alt-711 treatment (OAlt), n=7; old rats undergoing ALT-711 treatment and exercise (OAlt+Ex), n=8. Hemodynamic parameters as Mean±SD. Heart rate (HR); mean arterial pressure (MAP); cardiac output index (COI); stroke volume (SV); ejection fraction (EF); end-diastolic volume (EDV); end-diastolic pressure (EDP); end-systolic pressure (ESP); end systolic volume (ESV); peak rate of systolic pressure at instantaneous pressure (dP/dtmax-IP); peak rate of diastolic pressure relaxation (−dP/dtmin); total peripheral resistance index (TPRI); preload recruitable stroke work (PRSW). Statistical symbols:
P<0.05 vs. Y,
P<0.05 vs. O.
Heart rate (HR), MAP, COI/CI, SV, EF, ESP, dp/dtmax, −IP, −dp/dt and preload recruitable stroke work (PRSW) were all significantly decreased in O compared to Y animals. EDV, EDP and TPRI were significantly increased in O animals compared to Y. While ALT or Ex each caused partial but statistically significant improvement in age-associated hemodynamic perturbations, their combination resulted in a greater and more consistent improvement of these parameters (Table 1, column 2 vs. 5).
Fig. 1A illustrates representative P–V loops in a young and an old rat. The P–V loops are substantially shifted to the right in the old compared to the young rat. Moreover, the peak systolic pressure developed and the slope of the ESPVR are significantly attenuated. Fig. 1B illustrates the representative P–V loops at baseline in young and old rats and following ALT and Ex, individually and in combination in O rats. Again the P–V loops in Y and O rats are significantly different. ALT or Ex partially attenuates the influence of age but their combinations render a P–V relationship in O animals that is almost identical to that observed in Y rats.
Fig. 1.
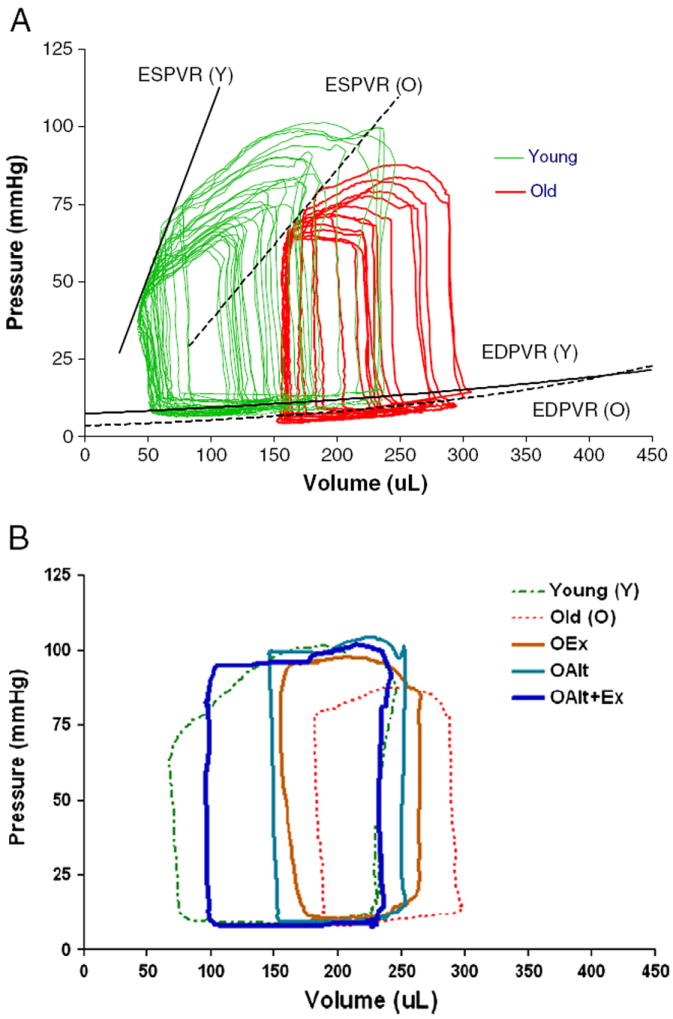
Representative traces of families of P–V loops from O and Y rats demonstrating differences in both ESPVR and EDPVR: (A). P–V loops were obtained using a combined conductance/manometric catheter. EDPVR and ESPVR were determined following IVC transient occlusion with a 2 Fr Fogarty catheter inserted via the femoral vein to the junction of the IVC and right atrium. (B). Representative traces demonstrating the effect of exercise (OEx), ALT treatment (OALT), and combination of ALT and exercise (OALT + Ex) on the P–V loops of O versus Y animals.
3.1. Systolic function
As illustrated in Fig. 2, and outlined in Table 1, there was a significant difference in all indices of systolic function in O compared to Y rats. All parameters of systolic function were restored in rats receiving both exercise and ALT such that the parameters were not statistically significantly different from Y rats. For example, dP/dtmax (Fig. 3A) was reduced by 37% in O vs Y rats consistent with depressed contractile function. Also, SV, a load dependent measure of systolic function was decreased in O vs Y rats but was significantly improved in O rats treated with ALT+Ex. In addition, Ees, the slope of the ESPVR (Fig. 3B), effective arterial elastance (Ea) and preload recruitable stroke work (PRSW), were significantly decreased in O rats. Remarkably, O rats who exercised and received ALT demonstrate a substantial restoration of contractility; animals who received either intervention alone also improved, but not to the same degree as the combined intervention (Table 1). Furthermore, there was a significant improvement of Ees and PRSW in rats receiving combined treatment consistent with an additive/synergistic effect of ALT and exercise on contractile function. Exercise, ALT, and combination therapy all improved EF significantly in old animals. However, these improvements remained lower than the normal EF in young rats. EF is a highly load dependent measure. Regardless of the increase in SV, the increase in EDV in O rats was such that a greater restoration of EF was not observed, even though EDV decreased with exercise and ALT toward the level of young animals. With regard to ventricular–vascular coupling, Ea was significantly decreased in O as compared to Y rats and was not significantly different in the O groups treated with Alt or exercise, but was significantly increased in the combined exercise and Alt group such that it was not different from Y. The Ea/Ees ratio, a metric of the efficiency of coupling between the ventricle and vasculature, was not significantly different across all the groups suggesting that efficient coupling is maintained in all groups.
Fig. 3.
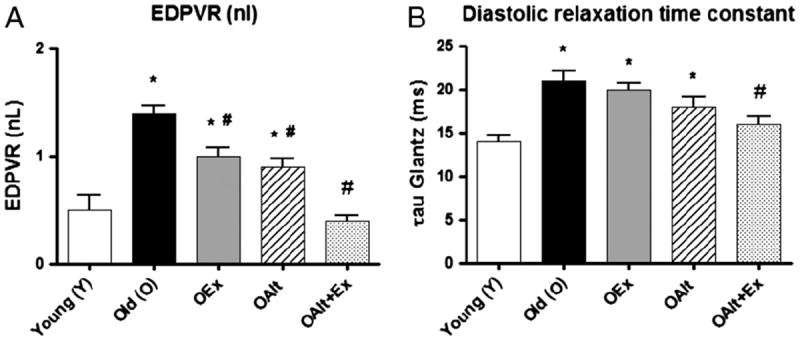
Combination of exercise and Alagebrium improves parameters of diastolic function in old rats: old rats demonstrate an increase in the slope of the non-linear EDPVR (increased diastolic stiffness). In addition, parameters of active ventricular relaxation (tau) are prolonged. OALT+Ex rats demonstrate a significant improvement in measures of diastolic stiffness. Interestingly, ALT alone is also associated with a significant improvement in ventricular relaxation. (*) P<0.05 vs. Y, (#) P<0.05 vs. O. N-values are the same as those specified in the table legend.
3.2. Diastolic function
All indices of diastolic function were significantly impaired in O compared to Y rats including EDV and EDP (Table 1). The P–V loops were shifted to the right in O rats compared to Y. This is accompanied by an increase in the slope of the EDPVR in the old rats compared to the young (Fig. 3A). Indices of ventricular relaxation were also altered. Tau (the time constant of relaxation) (Fig. 3B) and −dP/dTmin (Table 1) were significantly decreased in O vs Y rats. Old rats treated with a combination of ALT and Ex demonstrated significant improvement in indices of diastolic function such that these indices were similar to that of Y control rats. Old animals treated with ALT alone, did not demonstrate a significant improvement in active relaxation (tau) but showed significant improvement in −dP/dtmin (Table 1). Animals that underwent exercise alone had a significant improvement in EDP.
3.3. Vascular stiffness
We used PWV as an in vivo measure of vascular stiffness. As illustrated in Fig. 4, PWV was approximately doubled in O rats compared to Y. Both ALT treatment and exercise alone resulted in an equal and significant decrease in PWV in O rats. Remarkably, the combination of exercise and ALT resulted in a further substantial decrease in PWV such that this parameter was not statistically different to that observed in Y rats. This outcome is consistent with a significant impact of combined exercise and ALT on large artery compliance.
Fig. 4.
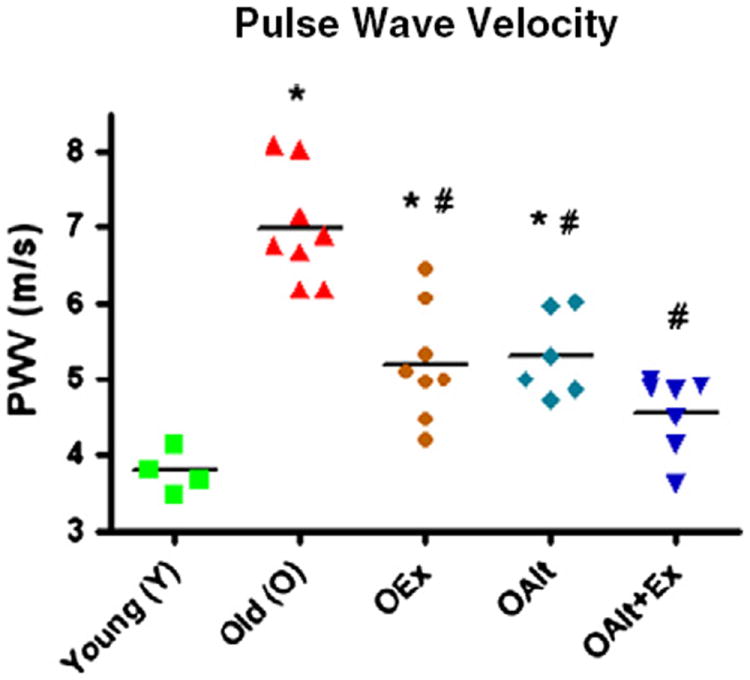
Age is associated with a substantial increase in vascular stiffness as measured by an increase in the pulse wave velocity (PWV). Stiffness is improved with Alagebriumor exercise and by the combination. (*) P<0.01 vs. Y; (#) P<0.01 vs. O; n=4 (Y), n=8 (O), n=8 (OEx), n=6 (OAlt), n=7 (OAlt + Ex).
4. Discussion
We have demonstrated for the first time that the combination of exercise training with the AGE crosslink breaker Alagebrium has a synergistic effect and significantly improves the parameters of both systolic and diastolic functions, and decreases vascular stiffness in a rat model of aging.
Our observations in old rat hearts are consistent with previous studies on aging (Boluyt et al., 2004; Chang et al., 2004; Raya et al., 1997; Rozenberg et al., 2006; Walker et al., 2006) and those of Pacher et al. (2004, 2002) who demonstrated significant impairments in systolic performance, and in diastolic relaxation and stiffness in ~24 month old Fischer 344 rats. Additionally, we have demonstrated that effective arterial elastance (Ea) is decreased in O rats and is restored toward that of Y with exercise and Alt. Furthermore Ea/Es ratio is modestly decreased in O rats and restored to Y value in the O rats treated with a combination of exercise and Alt. This suggests that the combined intervention restores optimal ventricular–arterial coupling. This finding is somewhat consistent with Chang et al. (2004), who have also demonstrated that aging is associated with a decrease in Ea, that Ea is partially restored with the inhibition of AGE. While we found no statistically significant effect of ALT alone on these parameters, we did however observe similar trends. The differences may be accounted for by the difference in AGE crosslink breaker used (Aminoguanidine vs. ALT) or the dose administered.
It is now well accepted that aging in humans is associated with a significant rise in vascular stiffness which causes a significant increase in left ventricular systolic wall stress (for review see Kass, 2002). Effective arterial elastance (Ea) is coupled to an increase in ventricular stiffness (Ees) maintaining myocardial efficiency at near normal levels. This increase in Ea in aging humans is in contrast to the findings in the rat aging model (Chang et al., 2004; Pacher et al., 2004). This discrepancy is explained on the basis of a markedly reduced stroke volume and therefore blood pressure in the O rats, despite an increase in central vascular stiffness. Efficient coupling is however maintained in this rodent model.
Acute changes in Ees are generally interpreted to represent changes in contractility while chronic changes, specifically increases, are indicative of alterations in chamber geometry and structure of the heart. In contrast, dP/dtmax, a contractility specific index, is not altered with aging in humans who in general demonstrate an increase in Ees. In aging humans diastolic dysfunction is the predominant feature, but systolic dysfunction can also develop, albeit perhaps at a later stage, in the disease process.
One of the interesting observations in this study is that ALT alone significantly improved the parameters of active myocardial relaxation as reflected by a decrease in tau. In addition, there was a significant improvement in the parameters of contractility as reflected by an increase in dp/dtmax. These observations suggest that ALT may have effects that modulate the molecular processes involved in active contraction and relaxation. It has recently been demonstrated that glycated proteins stimulate reactive oxygen species production in cardiac myocytes by an NADPH oxidase-dependent mechanism (Zhang et al., 2006). It is well established that not only does NADPH-dependent ROS production mediate myocardial hypertrophy, but it also leads to depressed myocardial contractility and impaired active relaxation. Thus, ALT may improve these parameters by preventing this process. The improvement in active relaxation with ALT may be particularly important for aged humans since even life-long and extensive exercise does not normalize measures of dynamic relaxation (Prasad et al., 2007).
Another interesting observation relates to the decrease inHR seen in O vs Y rats. This depressed HR is restored in ALT+Ex treated rats. Previous studies support a decrease in intrinsic heart rate (following sympathetic and parasympathetic blockade) in vivo in aging rats (Irigoyen et al., 2000) and humans (Jose and Collison, 1970). This has been confirmed in isolated spontaneously beating rat hearts (Friberg et al., 1985). Another possibility is amore profound depth of anesthesia in old rats compared to young rats, as there is a decrease in minimum alveolar concentration (MAC) of volatile anesthetics in old animals compared to young. Prior studies, however, demonstrated similar depressions of heart rate and blood pressure in old rats despite the use of different anesthetic techniques (Capasso et al., 1990; Pacher et al., 2004). Additionally there is evidence supporting the concept that a significant age-related impairment in baroreflex control of HR and sympathetic nerve activity occurs (Lakatta, 1993). These changes are likely to have significant implications for age-related alterations in cardiovascular regulation. It is unclear from this study why combined Ex and ALT therapy might restore HR toward that of the young. One potential explanation is that exercise might improve baroreflex control of HR in old rats. Alternatively, ALT might decrease central vascular stiffness and in that way alter the sensitivity of the carotid baroreceptor “rheostat,” restoring baroreflex sensitivity.
There is increasing evidence that exercise attenuates age-associated diastolic dysfunction in animals (Brenner et al., 2001; Starnes et al., 2003) and humans. For example, sedentary aging results in a large increase in LV diastolic stiffness compared to young but similarly sedentary subjects (Arbab-Zadeh et al., 2004). In contrast, master athletes had normal P–V relations compared to young controls, and similarly normal stress–strain curves demonstrating conclusively that life-long exercise (more than 2 decades in this study) preserves ventricular compliance (Arbab-Zadeh et al., 2004). While the effect of a lifetime of exercise on maintaining ventricular compliance is well established, until recently the effect of exercise training starting later in life remains unknown. Dr. Levine and coworkers have recently demonstrated that 1 year of endurance training had little effect on ventricular compliance in previously sedentary seniors (Fujimoto et al., 2010). On the other hand, there was a significant improvement in ventricular–vascular coupling. Interestingly, this is consistent with our observations in the exercised but not ALT treated rats. EDP and EDPVR were only modestly altered by exercise in contrast to the significant effect of exercise on vascular stiffness.
The large increase in vascular stiffness observed and its secondary effects on the heart (VVC) in the old rats are consistent with what has been previously described by our group and others (Atkinson, 2008; Kim et al., 2009; Santhanam et al., 2010; Soucy et al., 2006). Furthermore, the significant improvement in vascular stiffness, as measured by PWV, in the animals who received either exercise or ALT, and the improvement in the group that received both interventions, are consistent with the proposed biologic effect of each intervention and our original hypothesis: a physiologically meaningful interaction between the two interventions. The increase in vascular stiffness with age results from a number of interacting/underlying mechanisms involving all the components of the vascular wall including the endothelium, structural proteins, vascular smooth muscle and intercellular matrix. The accumulation of AGE in the matrix contributes to vascular stiffness changes with aging. In animal studies, Alagebrium improved vascular and ventricular properties (Asif et al., 2000; Candido et al., 2003; Liu et al., 2003; Vaitkevicius et al., 2001) and in one of the first human trials, ALT decreased pulsatile load as measured by pulse pressure and increased vascular compliance (Kass et al., 2001). Additional studies demonstrated improvement in left ventricular diastolic filling, quality of life, decreased left ventricular mass, left ventricular ejection and reduced aortic stiffness (Little et al., 2005; Liu et al., 2003), while another trial failed to demonstrate significant improvements in cardiac function, or systolic blood pressure with ALT administration without exercise (Hartog et al., 2011). In a large double-blind multicenter trial, ALT was compared to placebo for the treatment of hypertension in patients with and without hypertension (SAPPHIRE and SILVER studies) (Bakris et al., 2004). The primary outcome (drop in systolic blood pressure by 5 mm Hg) was not achieved in this study, however ALT decreased systolic blood pressure by 2–3 mm Hg more than the placebo group. These results are unfortunately difficult to interpret as all groups (both ALT and placebo) had a significant drop in blood pressure of 6–10 mm Hg during the first 2 weeks of the study (Bakris et al., 2004). Currently, there is no clear evidence that ALT alone has lesser or more profound effects in humans than in rodents. Furthermore, there are no human studies that have investigated the combined effect of ALT and exercise on vascular stiffness. It should also be noted that at least in theory, AGEs could de-stabilize the fibrous cap of an atherosclerotic cap pre-disposing to myocardial infarction. However in the more than 1000 subjects studied in clinical trials of Alagebrium to date, there has been no signal of increased myocardial infarction or death. Longer trials of this agent involving a full year of therapy in otherwise healthy seniors are currently ongoing (clinicatrials.gov NCT01014572) and will be necessary to ensure safety of this agent.
Although others have demonstrated a significant effect of ALT in spontaneously hypertensive aged rats (Susic et al., 2004), this study represents the first demonstration of the synergistic effects of ALT in combination with exercise in a purely aging model. Exercise has also been shown to improve vascular characteristics. For example, Tanaka et al. have demonstrated that the predicted age-related increase in central arterial stiffness did not develop in physically active women (Tanaka et al., 1998, 2000). Our data demonstrates a significant improvement in vascular stiffness in old exercised rats compared to age-matched sedentary controls. This observation is consistent with the idea that exercise can improve vascular properties.
What is the underlying mechanism contributing to the exercise and ALT-mediated reduction in vascular stiffness? Vascular stiffness is determined by dynamic (the vascular smooth muscle tone) and structural properties of the vascular wall. NO may regulate vascular stiffness both by acutely altering the tension developed by vascular smooth muscle cells, and by modifying the structure of blood vessels via its effects on vascular matrix protein characteristics (Wilkinson et al., 2004). There is emerging evidence that exercise can improve vascular properties by NO-dependent mechanisms. For example, in rats exercise increases the expression of NOS in the aorta and skeletal muscle arterioles (Spier et al., 2004; Tanabe et al., 2003). In humans, exercise increases NOS phosphorylation and thereby NO signaling (Hambrecht et al., 2003). In addition to promoting vascular remodeling, exercise causes activation of mechano-transduction mechanisms in both endothelial and vascular smooth muscle cells, which result in: 1) stimulation of signaling pathways that up-regulate genes which promote anti-atherogenesis, eNOS expression and phosphorylation, and NO bioavailability, 2) up-regulation of anti-oxidant mechanisms (e.g. superoxide dismutase (SOD)), and 3) potentiation of anti-inflammatory mechanisms (Fukai et al., 2000; Gielen et al., 2005; Hambrecht et al., 2003; Kojda and Hambrecht, 2005). This may further explain the findings by Guo et al. that ALT contributes to oxidative stress reduction by up-regulating activities of SOD and glutathione peroxidase (Guo et al., 2009).
There are a number of potential limitations of this study. We have been unable to measure AGE or RAGE receptors in cardiac tissue using immunohistochemical staining and biochemical assays. This would allow us to confirm that the mechanism by which ALT and Ex are working depends on its crosslink breaking activity. Additionally, we do not have measures of LV mass pre- and post-treatment (or any other echocardiographic data) in order to determine whether ALT cross-link breaking activity alters ventricular structure or wall thickness, and exercise capacity and catecholamine levels were also not measured. Some past studies have questioned the validity of measuring absolute ventricular volume via conductance catheters (Boltwood et al., 1989) (for a detailed review, see Burkhoff, 1990). These studies found that the offset volume is not constant but rather varies with loading conditions and stroke volume appears to be inconsistent if measured by ventriculography vs. a conductance catheter. However, the studies lack adequate validations utilizing sonomicrometry and ventriculography to measure a broad range of left ventricular volumes. At this point conductance catheters still provide accurate measurements during the cardiac cycle against which new methods are compared. Furthermore, conductance catheters offer the advantage of obtaining a continuous signal without the need to invade the chest cavity (Burkhoff, 1990; Kottam et al., 2011). An additional limitation is that PWV was not adjusted for either heart rate or blood pressure, both of which can theoretically change the measured value for PWV without affecting the underlying arterial wall properties. Currently there is no clear consensus in the literature regarding the association of heart rate and pulse wave velocity. While some studies demonstrate an association between PWV and heart rate (Amar et al., 2001; Sa Cunha et al., 1997), others fail to show this effect (Blacher et al., 1999; Nurnberger et al., 2003; Papaioannou et al., 2008). Despite these limitations, our physiologic measurements confirm the salutatory effects of the intervention on both systolic and diastolic functions in the aging rat heart. Lastly there is ongoing debate on the role of exercise intensity on improving the cardiovascular status (Cameron and Dart, 1994; Hoydal et al., 2007; Kemi et al., 2005). Our model of continuous moderate exercise resembles the regime mostly practiced and recommended for adults to improve cardiovascular function (Cameron and Dart, 1994). Recent animal studies however indicate that high intensity aerobic interval training might be more efficient at improving the cardiovascular status (Hoydal et al., 2007; Kemi et al., 2005).
In summary, exercise and ALT in combination improve diastolic stiffness, systolic and diastolic dysfunctions, LV contractility and large artery compliance in a rodent model of aging. This result may have important therapeutic implications for the aging population if it can be confirmed along with its safety profile over a longer period of time in seniors. Further studies extending these results to humans would seem warranted.
Acknowledgments
This work was supported by grants from the NIH, R01AG017479 (to BDL) and R01HL105296 (to DEB), as well as a grant from Alteon Corporation. Alteon had no input into the protocol design, data analysis, data presentation or manuscript preparation.
Disclosures
This work was supported in part by a grant from Alteon Corporation, the makers of Alagebrium, which provided the drug only. Alteon had no input into the protocol design, data analysis, data presentation or manuscript preparation.
References
- Amar J, Ruidavets JB, Chamontin B, Drouet L, Ferrieres J. Arterial stiffness and cardiovascular risk factors in a population-based study. J Hypertens. 2001;19:381–387. doi: 10.1097/00004872-200103000-00005. [DOI] [PubMed] [Google Scholar]
- Arbab-Zadeh A, Dijk E, Prasad A, Fu Q, Torres P, Zhang R, Thomas JD, Palmer D, Levine BD. Effect of aging and physical activity on left ventricular compliance. Circulation. 2004;110:1799–1805. doi: 10.1161/01.CIR.0000142863.71285.74. [DOI] [PubMed] [Google Scholar]
- Asif M, Egan J, Vasan S, Jyothirmayi GN, Masurekar MR, Lopez S, Williams C, Torres RL, Wagle D, Ulrich P, Cerami A, Brines M, Regan TJ. An advanced glycation endproduct cross-link breaker can reverse age-related increases in myocardial stiffness. Proc Natl Acad Sci U S A. 2000;97:2809–2813. doi: 10.1073/pnas.040558497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atkinson J. Age-related medial elastocalcinosis in arteries: mechanisms, animal models, and physiological consequences. J Appl Physiol. 2008;105:1643–1651. doi: 10.1152/japplphysiol.90476.2008. [DOI] [PubMed] [Google Scholar]
- Bakris GL, Bank AJ, Kass DA, Neutel JM, Preston RA, Oparil S. Advanced glycation end-product cross-link breakers. A novel approach to cardiovascular pathologies related to the aging process. Am J Hypertens. 2004;17:23 S–30 S. doi: 10.1016/j.amjhyper.2004.08.022. [DOI] [PubMed] [Google Scholar]
- Barouch LA, Harrison RW, Skaf MW, Rosas GO, Cappola TP, Kobeissi ZA, Hobai IA, Lemmon CA, Burnett AL, O’Rourke B, Rodriguez ER, Huang PL, Lima JA, Berkowitz DE, Hare JM. Nitric oxide regulates the heart by spatial confinement of nitric oxide synthase isoforms. Nature. 2002;416:337–339. doi: 10.1038/416337a. [DOI] [PubMed] [Google Scholar]
- Blacher J, Asmar R, Djane S, London GM, Safar ME. Aortic pulse wave velocity as a marker of cardiovascular risk in hypertensive patients. Hypertension. 1999;33:1111–1117. doi: 10.1161/01.hyp.33.5.1111. [DOI] [PubMed] [Google Scholar]
- Boltwood CM, Jr, Appleyard RF, Glantz SA. Left ventricular volume measurement by conductance catheter in intact dogs. Parallel conductance volume depends on left ventricular size. Circulation. 1989;80:1360–1377. doi: 10.1161/01.cir.80.5.1360. [DOI] [PubMed] [Google Scholar]
- Boluyt MO, Converso K, Hwang HS, Mikkor A, Russell MW. Echocardiographic assessment of age-associated changes in systolic and diastolic function of the female F344 rat heart. J Appl Physiol. 2004;96:822–828. doi: 10.1152/japplphysiol.01026.2003. [DOI] [PubMed] [Google Scholar]
- Brenner DA, Apstein CS, Saupe KW. Exercise training attenuates age-associated diastolic dysfunction in rats. Circulation. 2001;104:221–226. doi: 10.1161/01.cir.104.2.221. [DOI] [PubMed] [Google Scholar]
- Burkhoff D. The conductance method of left ventricular volume estimation. Methodologic limitations put into perspective. Circulation. 1990;81:703–706. doi: 10.1161/01.cir.81.2.703. [DOI] [PubMed] [Google Scholar]
- Cameron JD, Dart AM. Exercise training increases total systemic arterial compliance in humans. Am J Physiol. 1994;266:H693–H701. doi: 10.1152/ajpheart.1994.266.2.H693. [DOI] [PubMed] [Google Scholar]
- Candido R, Forbes JM, Thomas MC, Thallas V, Dean RG, Burns WC, Tikellis C, Ritchie RH, Twigg SM, Cooper ME, Burrell LM. A breaker of advanced glycation end products attenuates diabetes-induced myocardial structural changes. Circ Res. 2003;92:785–792. doi: 10.1161/01.RES.0000065620.39919.20. [DOI] [PubMed] [Google Scholar]
- Capasso JM, Palackal T, Olivetti G, Anversa P. Severe myocardial dysfunction induced by ventricular remodeling in aging rat hearts. Am J Physiol. 1990;259:H1086–H1096. doi: 10.1152/ajpheart.1990.259.4.H1086. [DOI] [PubMed] [Google Scholar]
- Chang KC, Hsu KL, Chou TF, Lo HM, Tseng YZ. Aminoguanidine prevents age-related deterioration in left ventricular–arterial coupling in Fisher 344 rats. Br J Pharmacol. 2004;142:1099–1104. doi: 10.1038/sj.bjp.0705831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fitch RM, Vergona R, Sullivan ME, Wang YX. Nitric oxide synthase inhibition increases aortic stiffness measured by pulse wave velocity in rats. Cardiovasc Res. 2001;51:351–358. doi: 10.1016/s0008-6363(01)00299-1. [DOI] [PubMed] [Google Scholar]
- Friberg P, Nordlander M, Lundin S, Folkow B. Effects of ageing on cardiac performance and coronary flow in spontaneously hypertensive and normotensive rats. Acta Physiol Scand. 1985;125:1–11. doi: 10.1111/j.1748-1716.1985.tb07687.x. [DOI] [PubMed] [Google Scholar]
- Fujimoto N, Prasad A, Hastings JL, Arbab-Zadeh A, Bhella PS, Shibata S, Palmer D, Levine BD. Cardiovascular effects of 1 year of progressive and vigorous exercise training in previously sedentary individuals older than 65 years of age. Circulation. 2010;122:1797–1805. doi: 10.1161/CIRCULATIONAHA.110.973784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fukai T, Siegfried MR, Ushio-Fukai M, Cheng Y, Kojda G, Harrison DG. Regulation of the vascular extracellular superoxide dismutase by nitric oxide and exercise training. J Clin Invest. 2000;105:1631–1639. doi: 10.1172/JCI9551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gielen S, Walther C, Schuler G, Hambrecht R. Anti-inflammatory effects of physical exercise. A new mechanism to explain the benefits of cardiac rehabilitation? J Cardiopulm Rehabil. 2005;25:339–342. doi: 10.1097/00008483-200511000-00005. [DOI] [PubMed] [Google Scholar]
- Gilbert JC, Glantz SA. Determinants of left ventricular filling and of the diastolic pressure–volume relation. Circ Res. 1989;64:827–852. doi: 10.1161/01.res.64.5.827. [DOI] [PubMed] [Google Scholar]
- Guo Y, Lu M, Qian J, Cheng YL. Alagebrium chloride protects the heart against oxidative stress in aging rats. J Gerontol A Biol Sci Med Sci. 2009;64:629–635. doi: 10.1093/gerona/glp023. [DOI] [PubMed] [Google Scholar]
- Hambrecht R, Adams V, Erbs S, Linke A, Krankel N, Shu Y, Baither Y, Gielen S, Thiele H, Gummert JF, Mohr FW, Schuler G. Regular physical activity improves endothelial function in patients with coronary artery disease by increasing phosphorylation of endothelial nitric oxide synthase. Circulation. 2003;107:3152–3158. doi: 10.1161/01.CIR.0000074229.93804.5C. [DOI] [PubMed] [Google Scholar]
- Hartog JW, Willemsen S, van Veldhuisen DJ, Posma JL, van Wijk LM, Hummel YM, Hillege HL, Voors AA. Effects of alagebrium, an advanced glycation endproduct breaker, on exercise tolerance and cardiac function in patients with chronic heart failure. Eur J Heart Fail. 2011;13:899–908. doi: 10.1093/eurjhf/hfr067. [DOI] [PubMed] [Google Scholar]
- Hoydal MA, Wisloff U, Kemi OJ, Ellingsen O. Running speed and maximal oxygen uptake in rats and mice: practical implications for exercise training. Eur J Cardiovasc Prev Rehabil. 2007;14:753–760. doi: 10.1097/HJR.0b013e3281eacef1. [DOI] [PubMed] [Google Scholar]
- Irigoyen MC, Moreira ED, Werner A, Ida F, Pires MD, Cestari IA, Krieger EM. Aging and baroreflex control of RSNA and heart rate in rats. Am J Physiol Regul Integr Comp Physiol. 2000;279:R1865–R1871. doi: 10.1152/ajpregu.2000.279.5.R1865. [DOI] [PubMed] [Google Scholar]
- Jose AD, Collison D. The normal range and determinants of the intrinsic heart rate in man. Cardiovasc Res. 1970;4:160–167. doi: 10.1093/cvr/4.2.160. [DOI] [PubMed] [Google Scholar]
- Kass DA. Age-related changes in ventricular–arterial coupling: pathophysiologic implications. Heart Fail Rev. 2002;7:51–62. doi: 10.1023/a:1013749806227. [DOI] [PubMed] [Google Scholar]
- Kass DA. Getting better without AGE: new insights into the diabetic heart. Circ Res. 2003;92:704–706. doi: 10.1161/01.RES.0000069362.52165.C9. [DOI] [PubMed] [Google Scholar]
- Kass DA, Shapiro EP, Kawaguchi M, Capriotti AR, Scuteri A, deGroof RC, Lakatta EG. Improved arterial compliance by a novel advanced glycation endproduct crosslink breaker. Circulation. 2001;104:1464–1470. doi: 10.1161/hc3801.097806. [DOI] [PubMed] [Google Scholar]
- Kemi OJ, Haram PM, Loennechen JP, Osnes JB, Skomedal T, Wisloff U, Ellingsen O. Moderate vs. high exercise intensity: differential effects on aerobic fitness, cardiomyocyte contractility, and endothelial function. Cardiovasc Res. 2005;67:161–172. doi: 10.1016/j.cardiores.2005.03.010. [DOI] [PubMed] [Google Scholar]
- Kim JH, Bugaj LJ, Oh YJ, Bivalacqua TJ, Ryoo S, Soucy KG, Santhanam L, Webb A, Camara A, Sikka G, Nyhan D, Shoukas AA, Ilies M, Christianson DW, Champion HC, Berkowitz DE. Arginase inhibition restores NOS coupling and reverses endothelial dysfunction and vascular stiffness in old rats. J Appl Physiol. 2009;107:1249–1257. doi: 10.1152/japplphysiol.91393.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kojda G, Hambrecht R. Molecular mechanisms of vascular adaptations to exercise. Physical activity as an effective antioxidant therapy? Cardiovasc Res. 2005;67:187–197. doi: 10.1016/j.cardiores.2005.04.032. [DOI] [PubMed] [Google Scholar]
- Kottam A, Dubois J, McElligott A, Henderson KK. Novel approach to admittance to volume conversion for ventricular volume measurement. Conf Proc IEEE Eng Med Biol Soc. 2011:2514–2517. doi: 10.1109/IEMBS.2011.6090696. [DOI] [PubMed] [Google Scholar]
- Lakatta EG. Cardiovascular regulatory mechanisms in advanced age. Physiol Rev. 1993;73:413–467. doi: 10.1152/physrev.1993.73.2.413. Review. No abstract available. [DOI] [PubMed] [Google Scholar]
- Little WC, Zile MR, Kitzman DW, Hundley WG, O’Brien TX, Degroof RC. The effect of alagebrium chloride (ALT-711), a novel glucose cross-link breaker, in the treatment of elderly patients with diastolic heart failure. J Card Fail. 2005;11:191–195. doi: 10.1016/j.cardfail.2004.09.010. [DOI] [PubMed] [Google Scholar]
- Liu J, Masurekar MR, Vatner DE, Jyothirmayi GN, Regan TJ, Vatner SF, Meggs LG, Malhotra A. Glycation end-product cross-link breaker reduces collagen and improves cardiac function in aging diabetic heart. Am J Physiol Heart Circ Physiol. 2003;285:H2587–H2591. doi: 10.1152/ajpheart.00516.2003. [DOI] [PubMed] [Google Scholar]
- McGuire DK, Levine BD, Williamson JW, Snell PG, Blomqvist CG, Saltin B, Mitchell JH. A 30-year follow-up of the Dallas Bedrest and Training Study: II. Effect of age on cardiovascular adaptation to exercise training. Circulation. 2001;104:1358–1366. [PubMed] [Google Scholar]
- Mitchell GF, Hwang SJ, Vasan RS, Larson MG, Pencina MJ, Hamburg NM, Vita JA, Levy D, Benjamin EJ. Arterial stiffness and cardiovascular events: the Framingham Heart Study. Circulation. 2010;121:505–511. doi: 10.1161/CIRCULATIONAHA.109.886655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nurnberger J, Dammer S, Opazo Saez A, Philipp T, Schafers RF. Diastolic blood pressure is an important determinant of augmentation index and pulse wave velocity in young, healthy males. J Hum Hypertens. 2003;17:153–158. doi: 10.1038/sj.jhh.1001526. [DOI] [PubMed] [Google Scholar]
- Pacher P, Mabley JG, Soriano FG, Liaudet L, Szabo C. Activation of poly(ADP-ribose) polymerase contributes to the endothelial dysfunction associated with hypertension and aging. Int J Mol Med. 2002;9:659–664. [PubMed] [Google Scholar]
- Pacher P, Mabley JG, Liaudet L, Evgenov OV, Marton A, Hasko G, Kollai M, Szabo C. Left ventricular pressure–volume relationship in a rat model of advanced aging-associated heart failure. Am J Physiol Heart Circ Physiol. 2004;287:H2132–H2137. doi: 10.1152/ajpheart.00405.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papaioannou TG, Vlachopoulos CV, Alexopoulos NA, Dima I, Pietri PG, Protogerou AD, Vyssoulis GG, Stefanadis CI. The effect of heart rate on wave reflections may be determined by the level of aortic stiffness: clinical and technical implications. Am J Hypertens. 2008;21:334–340. doi: 10.1038/ajh.2007.52. [DOI] [PubMed] [Google Scholar]
- Prasad A, Popovic ZB, Arbab-Zadeh A, Fu Q, Palmer D, Dijk E, Greenberg NL, Garcia MJ, Thomas JD, Levine BD. The effects of aging and physical activity on Doppler measures of diastolic function. Am J Cardiol. 2007;99:1629–1636. doi: 10.1016/j.amjcard.2007.01.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raya TE, Gaballa M, Anderson P, Goldman S. Left ventricular function and remodeling after myocardial infarction in aging rats. Am J Physiol. 1997;273:H2652–H2658. doi: 10.1152/ajpheart.1997.273.6.H2652. [DOI] [PubMed] [Google Scholar]
- Rozenberg S, Tavernier B, Riou B, Swynghedauw B, Page CL, Boucher F, Leiris J, Besse S. Severe impairment of ventricular compliance accounts for advanced age-associated hemodynamic dysfunction in rats. Exp Gerontol. 2006;41:289–295. doi: 10.1016/j.exger.2005.11.009. [DOI] [PubMed] [Google Scholar]
- Sa Cunha R, Pannier B, Benetos A, Siche JP, London GM, Mallion JM, Safar ME. Association between high heart rate and high arterial rigidity in normotensive and hypertensive subjects. J Hypertens. 1997;15:1423–1430. doi: 10.1097/00004872-199715120-00009. [DOI] [PubMed] [Google Scholar]
- Santhanam L, Tuday EC, Webb AK, Dowzicky P, Kim JH, Oh YJ, Sikka G, Kuo M, Halushka MK, Macgregor AM, Dunn J, Gutbrod S, Yin D, Shoukas A, Nyhan D, Flavahan NA, Belkin AM, Berkowitz DE. Decreased S-nitrosylation of tissue transglutaminase contributes to age-related increases in vascular stiffness. Circ Res. 2010;107:117–125. doi: 10.1161/CIRCRESAHA.109.215228. [DOI] [PubMed] [Google Scholar]
- Shibata S, Levine BD. The effect of exercise training on biologic vascular age in healthy seniors. Am J Physiol Heart Circ Physiol. 2012 Mar;302(6):H1340–H1346. doi: 10.1152/ajpheart.00511.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soucy KG, Ryoo S, Benjo A, Lim HK, Gupta G, Sohi JS, Elser J, Aon MA, Nyhan D, Shoukas AA, Berkowitz DE. Impaired shear stress-induced nitric oxide production through decreased NOS phosphorylation contributes to age-related vascular stiffness. J Appl Physiol. 2006;101:1751–1759. doi: 10.1152/japplphysiol.00138.2006. [DOI] [PubMed] [Google Scholar]
- Spier SA, Delp MD, Meininger CJ, Donato AJ, Ramsey MW, Muller-Delp JM. Effects of ageing and exercise training on endothelium-dependent vasodilatation and structure of rat skeletal muscle arterioles. J Physiol. 2004;556:947–958. doi: 10.1113/jphysiol.2003.060301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spinetti G, Wang M, Monticone R, Zhang J, Zhao D, Lakatta EG. Rat aortic MCP-1 and its receptor CCR2 increase with age and alter vascular smooth muscle cell function. Arterioscler Thromb Vasc Biol. 2004;24:1397–1402. doi: 10.1161/01.ATV.0000134529.65173.08. [DOI] [PubMed] [Google Scholar]
- Starnes JW, Taylor RP, Park Y. Exercise improves postischemic function in aging hearts. Am J Physiol Heart Circ Physiol. 2003;285:H347–H351. doi: 10.1152/ajpheart.00952.2002. [DOI] [PubMed] [Google Scholar]
- Susic D, Varagic J, Ahn J, Frohlich ED. Cardiovascular and renal effects of a collagen cross-link breaker (ALT 711) in adult and aged spontaneously hypertensive rats. Am J Hypertens. 2004;17:328–333. doi: 10.1016/j.amjhyper.2003.12.015. [DOI] [PubMed] [Google Scholar]
- Tanabe T, Maeda S, Miyauchi T, Iemitsu M, Takanashi M, Irukayama-Tomobe Y, Yokota T, Ohmori H, Matsuda M. Exercise training improves ageing-induced decrease in eNOS expression of the aorta. Acta Physiol Scand. 2003;178:3–10. doi: 10.1046/j.1365-201X.2003.01100.x. [DOI] [PubMed] [Google Scholar]
- Tanaka H, DeSouza CA, Seals DR. Absence of age-related increase in central arterial stiffness in physically active women. Arterioscler Thromb Vasc Biol. 1998;18:127–132. doi: 10.1161/01.atv.18.1.127. [DOI] [PubMed] [Google Scholar]
- Tanaka H, Dinenno FA, Monahan KD, Clevenger CM, DeSouza CA, Seals DR. Aging, habitual exercise, and dynamic arterial compliance. Circulation. 2000;102:1270–1275. doi: 10.1161/01.cir.102.11.1270. [DOI] [PubMed] [Google Scholar]
- Ulrich P, Cerami A. Protein glycation, diabetes, and aging. Recent Prog Horm Res. 2001;56:1–21. doi: 10.1210/rp.56.1.1. [DOI] [PubMed] [Google Scholar]
- Vaitkevicius PV, Lane M, Spurgeon H, Ingram DK, Roth GS, Egan JJ, Vasan S, Wagle DR, Ulrich P, Brines M, Wuerth JP, Cerami A, Lakatta EG. A cross-link breaker has sustained effects on arterial and ventricular properties in older rhesus monkeys. Proc Natl Acad Sci U S A. 2001;98:1171–1175. doi: 10.1073/pnas.98.3.1171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varghese P, Harrison RW, Lofthouse RA, Georgakopoulos D, Berkowitz DE, Hare JM. Beta(3)-adrenoceptor deficiency blocks nitric oxide-dependent inhibition of myocardial contractility. J Clin Invest. 2000;106:697–703. doi: 10.1172/JCI9323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walker EM, Jr, Nillas MS, Mangiarua EI, Cansino S, Morrison RG, Perdue RR, Triest WE, Wright GL, Studeny M, Wehner P, Rice KM, Blough ER. Age-associated changes in hearts of male Fischer 344/Brown Norway F1 rats. Ann Clin Lab Sci. 2006;36:427–438. [PubMed] [Google Scholar]
- Wang M, Lakatta EG. Altered regulation of matrix metalloproteinase-2 in aortic remodeling during aging. Hypertension. 2002;39:865–873. doi: 10.1161/01.hyp.0000014506.13322.66. [DOI] [PubMed] [Google Scholar]
- Wang Y-X, Halks-Miller M, Vergona R, Sullivan ME, Fitch R, Mallari C, Martin-McNulty B, Da Cunha V, Freay A, Rubanyi GM, Kauser K. Increased aortic stiffness assessed by pulse wave velocity in apolipoprotein E-deficient mice. Am J Physiol Heart Circ Physiol. 2000;278:H428–H434. doi: 10.1152/ajpheart.2000.278.2.H428. [DOI] [PubMed] [Google Scholar]
- Wang M, Takagi G, Asai K, Resuello RG, Natividad FF, Vatner DE, Vatner SF, Lakatta EG. Aging increases aortic MMP-2 activity and angiotensin II in nonhuman primates. Hypertension. 2003;41:1308–1316. doi: 10.1161/01.HYP.0000073843.56046.45. [DOI] [PubMed] [Google Scholar]
- Wilkinson IB, Franklin SS, Cockcroft JR. Nitric oxide and the regulation of large artery stiffness: from physiology to pharmacology. Hypertension. 2004;44:112–116. doi: 10.1161/01.HYP.0000138068.03893.40. [DOI] [PubMed] [Google Scholar]
- Wolffenbuttel BH, Boulanger CM, Crijns FR, Huijberts MS, Poitevin P, Swennen GN, Vasan S, Egan JJ, Ulrich P, Cerami A, Levy BI. Breakers of advanced glycation end products restore large artery properties in experimental diabetes. Proc Natl Acad Sci U S A. 1998;95:4630–4634. doi: 10.1073/pnas.95.8.4630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang M, Kho AL, Anilkumar N, Chibber R, Pagano PJ, Shah AM, Cave AC. Glycated proteins stimulate reactive oxygen species production in cardiac myocytes: involvement of Nox2 (gp91phox)-containing NADPH oxidase. Circulation. 2006;113:1235–1243. doi: 10.1161/CIRCULATIONAHA.105.581397. [DOI] [PubMed] [Google Scholar]
- Zieman S, Kass D. Advanced glycation end product cross-linking: pathophysiologic role and therapeutic target in cardiovascular disease. Congest Heart Fail. 2004;10:144–149. doi: 10.1111/j.1527-5299.2004.03223.x. quiz 150-1. [DOI] [PubMed] [Google Scholar]