

Abstract
Flavobacterium psychrophilum is a Gram-negative bacterium, responsible for the bacterial cold-water disease and the rainbow trout fry syndrome in freshwater salmonid fish. At present, there is only one commercial vaccine in Chile, made with two Chilean F. psychrophilum isolates and another licensed in Europe. The present study analyzed neutrophil migration, as a marker of innate immune activation, in zebrafish (Danio rerio) in response to different F. psychrophilum bath vaccines, which is the first step in evaluating vaccine effectiveness and efficiency in fish. Results indicated that bacterins of the LM-02-Fp isolate were more immunogenic than those from the LM-13-Fp isolate. However, no differences were observed between the same bacteria inactivated by either formaldehyde or heat. Importantly, the same vaccine formulation without an adjuvant only triggered a mild neutrophil migration compared to the complete vaccine. Observations also found that, after a year of storage at 4°C, the activation of the innate immune system by the different vaccines was considerably decreased. Finally, new vaccine formulations prepared with heat and formaldehyde inactivated LM-02-Fp were significantly more efficient than the available commercial vaccine in regard to stimulating the innate immune system.
1. Introduction
Flavobacterium psychrophilum [1], a Gram-negative, filamentous, psychrotrophic bacterium belonging to the phylum Bacteroidetes, is the causative agent of bacterial cold-water disease (BCWD) and rainbow trout fry syndrome (RTFS) in freshwater salmonid fish worldwide [2, 3]. In Chile, this infectious bacterium was first observed in freshwater aquaculture facilities in 1993, and the incidence of F. psychrophilum has dramatically increased since then [4, 5].
Despite the severe impact of this disease, antimicrobial therapies are currently the only method for controlling this condition in farmed fish, and it has been estimated that 55 tons of florfenicol and oxytetracycline each were used in Chilean farms to control outbreaks between 2006 and 2009 [6]. A tentatively licensed commercial vaccine that contains whole-cells inactivated with formaldehyde, Flavomune vaccine (SAG N°2160-BP), was recently developed in Chile. However it is important to note that its efficacy was tested using injection-based challenge models that completely bypassed the protective functions of the fish skin-mucus layer. This is significant due to this layer acting as an important barrier to disease infection in fish.
One key for efficient immunization and a powerful generation of specific antibodies is to ensure efficient antigen presentation by cells from the innate immune system. This event triggers an antigen-specific adaptive immune response that leads to the production of specific antibodies against an invading pathogen [7]. Activated, or noncirculating, leukocytes are involved in the initiation of the adaptive immune response [8]. Recent studies have moreover established various factors that elicit the innate immune response shaping adaptive immunity [8–13]. For instance, splenic neutrophils facilitate the antibody response of marginal zone B cells to microbial antigens [13]. These granulocytes promote B cell survival, as well as the production of IgM, IgG, and IgA antibodies [13, 14].
During the last decade, the zebrafish (Danio rerio) has been positioned as a powerful model for immunity research not only for other fish species, but also for mammals [15–19]. Some outstanding characteristics are its genetic tractability and transparency during the embryonic and larval stages, which facilitate monitoring infection and inflammation processes in vivo [18, 20–24]. Early during zebrafish development, the innate immune system exists in isolation to the adaptive system, which develops later in the larval stages and requires 4–6 weeks before achieving full functionality [25].
In the present work, zebrafish were used to compare the effect of different F. psychrophilum vaccines on the activation of the innate immune system, using neutrophils as specific innate immune system markers. These leukocytes are the first cells to be mobilized in response to injury, and they are the first to infiltrate damaged tissue [26–29]. For all assays, the Tg(mpx:GFP) i114 transgenic zebrafish line was used given that it exclusively expresses the green fluorescent protein (GFP) in neutrophils and allows tracking individual immune cells in live animals [30]. Zebrafish larvae were bath-vaccinated with three strains of bacteria inactivated by heat (VAH) or formaldehyde (VAF) and with or without an adjuvant. Afterwards, neutrophil migration was determined to investigate the effects of vaccination on the activation of the innate immune system.
2. Materials and Methods
2.1. Selection of Bacterial Strains for Vaccine Formulation
Two F. psychrophilum strains (LM-02-Fp and LM-13-Fp) were isolated in 2006 in Chile from the kidneys of clinically infected rainbow trout (Oncorhynchus mykiss) for subsequent vaccine preparation. The LM-02-Fp isolate is a sequence type (ST) 2 strain and antigenically Group 1 (1150), while LM-13-Fp is a ST12 strain and Group 2 (1658) [31, 32]. The identity of each isolate was confirmed as F. psychrophilum by using standard phenotyping procedures [33], including analyses of colony morphology and pigmentation, cell morphology, gliding motility, Gram-staining, cytochrome oxidase and catalase activities, oxidation/fermentation reactions, the presence of cell wall-associated flexirubin-type pigments, and the absorption of Congo red. Each isolate was routinely grown on a tryptone yeast extract salts medium (TYES; 0.4% tryptone, 0.05% yeast extract, 0.02% anhydrous calcium chloride, and 0.05% magnesium sulphate heptahydrate; pH 7.2) in either a liquid or solid state (TYES supplemented with 1% (w/v) agar bacteriological). Bacteria were aerobically incubated at 15°C for 3–5 days. Stock cultures were then frozen and kept at −80°C in Criobilles tubes (AES Laboratory) or in a TYES broth with 10% glycerol.
2.2. Vaccine Preparation
The selected F. psychrophilum isolates were grown in TYES broth for 56 h in order to obtain a final concentration of 3.49 ± 0.51 × 108 colony forming units (CFU) mL−1. Eight vaccines (including F. psychrophilum cells and extracellular products) (Table 1) were prepared using the following two inactivation protocols: (i) LM-02-Fp and LM-13-Fp cultures were inactivated by heat at 60°C for 6 h and then stored at 4°C; and (ii) LM-02-Fp and LM-13-Fp cultures were inactivated through the addition of formalin to a final concentration of 0.7% and then stored at 4°C. The efficiency of each inactivation method was determined by spreading 0.1 mL of culture in 10-fold dilutions (10−1 to 10−8 in duplicate) onto a TYES agar plates. Subsequently, the presence of bacterial colonies was monitored for 5 days. Once the plate counts were negative for F. psychrophilum, 1 mL of the culture was taken directly from each inactivation treatment (VAH or VAF) and seeded onto TYES agar.
Table 1.
Vaccine formulation.
| Strain | Inactivation method | |||
|---|---|---|---|---|
| Heat | Formaldehyde | |||
| Without adjuvant | With adjuvant | Without adjuvant | With adjuvant | |
| LM-02-Fp | VAH 02 | VAH 02 + Adj. | VAF 02 | VAF 02 + Adj. |
| LM-13-Fp | VAH 13 | VAH 13 + Adj. | VAF 13 | VAF 13 + Adj. |
| Control medium | TYES (H) | TYES (H) + Adj. | TYES (F) | TYES (F) + Adj. |
Aliquots from the VAF or VAH vaccines based on LM-02-Fp and LM-13-Fp, respectively, were mixed 1 : 1 with the adjuvant Montanide IMS 1312 VG PR (SEPPIC, France) according to the manufacturer's protocols. In addition to this, the commercial Flavomune vaccine (SAG N°2160.BP), made with two Chilean F. psychrophilum strains (coded 1 and 4), was included for comparative purposes. During the study, the commercial vaccine was kept at 4°C until use.
2.3. Zebrafish Strains and Maintenance
Zebrafish were maintained and raised at Universidad Andrés Bello facilities according to standard protocols [34]. The strain of zebrafish used was Tg(mpx:GFP) i114 [30]. All embryos were collected by natural spawning, staged according to Kimmel and Colleagues [21], and raised in Petri dishes at 28.5°C in an E3 medium (5 mM of NaCl, 0.17 mM of KCl, 0.33 mM of CaCl2, 0.33 mM of MgSO4, and without methylene blue; pH 7.0) as previously described [35]. Embryonic and larval stadium were expressed in hours post-fertilization (hpf).
2.4. Larvae Vaccination and Quantification of Neutrophil Migration
Fifteen Tg(mpx:GFP) i114 larvae were selected at 48 hpf, as previously described [36], and bath-vaccinated by incubation with the different vaccines at a 1 : 10 dilution for 1 min (Table 1). A control medium with or without adjuvant was also included. After the bath-vaccination, larvae were washed three times for 5 min with the E3 medium, and neutrophils were monitored. Quantification of neutrophil migration was performed by analyzing the displacement of GFP positive cells outside of the caudal hematopoietic tissue (CHT) to the region of interest (ROI) (Figure 1) at 2, 4, 6, and 8 hours post-incubation (hpi) under a fluorescent stereoscope as previously described [36].
Figure 1.
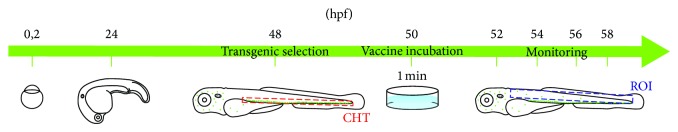
Scheme of vaccine assay. At 48 hpf Tg(mpx:GFP) i114 nonactivated zebrafish larva, with neutrophils located only in the CHT (demarked in red), was selected, bath-incubated, and quantified the number of neutrophils mobilized to the region of interest (ROI, demarked in blue) at 2, 4, 6, and 8 hpi.
2.5. Imaging and Statistical Analyses
Photographs were taken with a QImaging MicroPublisher 5.0 RVT camera using the Olympus SZX16 stereoscope. Images were processed with Photoshop CS5.1, with pictures showing the representative effects of each treatment. Data were analyzed by two-way ANOVA using the Prism 6 software (GraphPad Software).
3. Results
To analyze the innate immune response triggered by the different vaccine formulations, neutrophil behavior was analyzed in live zebrafish larvae (Figure 1). To quantify the number of mobilized neutrophils, a specific region in the larval tail was analyzed (ROI) since neutrophils are normally restricted to the CHT, with few circulating cells. Analysis of quantified neutrophil migration indicated that only VAH and VAF LM-02-Fp vaccines were able to trigger an innate immune response (Figures 2(c), 2(d), 2(k), and 2(l)). The number of neutrophils detected outside the CHT in larvae treated with the VAH or VAF LM-13-Fp vaccine was the same as that with the TYES medium, the negative control (Figures 2(e)–2(h), 2(k), and 2(l)). No statistical differences in the number of mobilized neutrophils were found between same-strain vaccines made from either heat or formaldehyde inactivation (Figures 2(k) and 2(l)). As was expected, when an adjuvant was added to the vaccine formulation all responses considerably increased, even in the TYES medium group (Figures 2(k) and 2(l); Figure S1 in Supplementary Material available online at http://dx.doi.org/10.1155/2015/515187). This result indicates that the adjuvant by itself was able to trigger the innate immune response.
Figure 2.

Effect on innate immune response triggered by freshly prepared vaccines against F. psychrophilum. ((a)–(j)) Representative images of zebrafish tails from larvae at 58 hpf. ((k) and (l)) Graph quantifying the number of neutrophils outside of the CHT (ROI), showing the effect on the innate immune response in larvae bath-vaccinated with (k) heat inactivated bacteria or (l) formaldehyde. Statistical analysis was performed using two-way ANOVA. All embryos ((a)–(j)) were oriented anterior to the left, with the dorsal on top. Data represent the mean ± standard error from three independent experiments with 15 larvae each. * P value < 0.05, *** P < 0.001, and **** P < 0.0001.
In order to determine if vaccine effects changed with time, the same assays as previously described were performed again using one-year-old vaccines. Results indicated that all vaccines had diminished activity (Figure 3). In the case of the VAH and VAF LM-02-Fp vaccines with an adjuvant, a freshly prepared vaccine mobilized an average of 11 neutrophils from the CHT at 8 hpi (Figures 2(k) and 2(l)). This is in contrast to the old vaccines, which moved approximately 8 neutrophils outside of the CHT after 8 hpi (Figure 3). Likewise, the one-year-old VAH and VAF LM-02-Fp vaccines without an adjuvant showed considerably diminished activity. Moreover, the old VAH LM-02-Fp vaccine only started to exert effects, as compared to the control, at 6 hpi, while the VAF LM-02-Fp vaccine showed effects at 8 hpi (Figure 3). These results suggest that a year of storage is too long of a time for maintaining efficacy.
Figure 3.

Effect on innate immune response triggered by one-year-old stored vaccines. ((a) and (b)) Graph quantifying neutrophil migration outside of the CHT (ROI), as triggered by bath-vaccination with formulas prepared from (a) heat or (b) formaldehyde inactivated bacteria. Statistical analysis was performed using two-way ANOVA. Data represent the mean ± standard error from three independent experiments with 15 larvae each. * P value < 0.05, ** P < 0.005, *** P < 0.001, and **** P < 0.0001.
Of significant note, however, is that both vaccines newly formulated with the isolate LM-02-Fp triggered an innate immune response at least twice as powerful as that produced by the commercial Flavomune vaccine (Figure 4).
Figure 4.
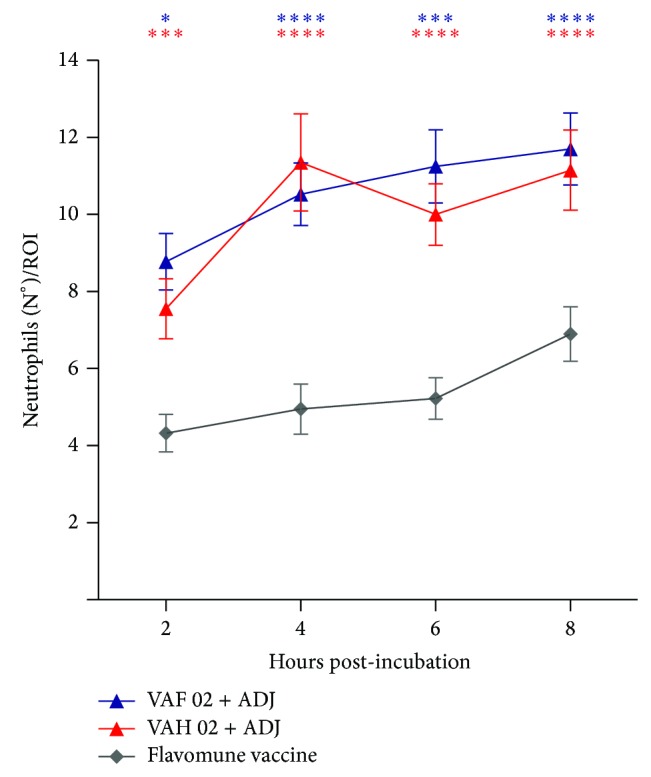
Comparison of the effects on the innate immune response between Flavomune vaccine and newly formulated vaccines. Graph quantifying neutrophil migration to the ROI for larvae bath-vaccinated with VAF-02-Fp + adjutant or VAH-02-Fp + adjutant, thus showing the activation of innate immune response. Statistical analysis was performed with a two-way ANOVA. Data represent the mean ± standard error from three independent experiments with 15 larvae each. * P value < 0.05, *** P < 0.005, and **** P < 0.0001.
4. Discussion
Vaccination is currently one of the most used practices in the prevention of fish diseases. Therefore, it is very important for the aquaculture industry to know the efficiency and effectiveness of the different commercially available vaccines. A preliminary step in selecting the most suitable vaccine against a specific pathogen would be to determine its effect on the innate immune system given that the activation of this system could yield a higher production of specific antibodies. In contrast, antigens unable to activate the innate immune response would probably induce a poor production of antibodies.
In Chile, autochthonous immersion bacterins made from single farm isolates have been used for years as protection against infection by F. psychrophilum [37], but there is still little information on how effective these antipathogen treatments are in terms of relative survival. Moreover, the currently available commercial vaccine against F. psychrophilum is insufficient, although there have been numerous studies towards developing a global commercial vaccine [38].
At the present time, there are no comparative studies in fish regarding the responses triggered by formaldehyde or heat inactivated antigens; only there studies using bath-vaccinations on fish from the same population. In fact, there are no concordant results in previous works performed with different vaccines containing formaldehyde- or heat-inactivated virulent cells of F. psychrophilum. This situation could be explained due to experiments being developed in different fish species (rainbow trout; Coho salmon (Oncorhynchus kisutch) and ayu (Plecoglossus altivelis)). Moreover, the vaccine formula used in previous studies differed in the content of adjuvant [39–45]. Importantly, Madetoja and Colleagues [46] compared the efficacy of formaldehyde- and heat-inactivated antigens in rainbow trout by intraperitoneal injection, demonstrating that both antigens produced high circulating antibody levels and efficient protection against the F. psychrophilum. The present results agree with this study, with LM-02-Fp and LM-13-Fp, regardless of the inactivation protocol, showing similar innate immune responses in zebrafish.
However, the current study used a set of vaccines supplemented with Montanide IMS 1312 VG, an aqueous adjuvant containing liquid particles, and an immunostimulating compound listed as a GRAS substance (generally recognized as safe). This adjuvant improves the presentation of the antigen to the immune cells and keeps it within the fish tissues, thus increasing the length of protection. It is important to highlight that it does not cause off-target effects such as melanosis [47]. In this sense, the present results demonstrate that Montanide IMS 1312 VG can be used in studies on fish vaccination against F. psychrophilum. Moreover, this adjuvant is able to activate an innate immune response. At present, further immunological studies are in progress in rainbow trout in order to establish the adaptive immune responses to specific antigens (unpublished data).
Another factor that should be studied in depth before designing an effective vaccine is the serotyping within the bacterial species. Mata and Colleagues [48] proposed a serotyping system for F. psychrophilum using slide agglutination and compared it to the serological schemes previously proposed by Lorenzen and Olesen [49] and Izumi and Wakabayashi [50]. Three to seven serotypes, mainly associated with the host species, were established [47]. In Chile, Valdebenito and Avendaño-Herrera [51] determined the existence of antigenic heterogeneity within F. psychrophilum with four patterns of serological reactions. According to this serological scheme, LM-02-Fp corresponds to the Group 1, while the strain LM-13-Fp belongs to the minor serological Group 4. Thus, a possible explanation for the differences in the results obtained for LM-02-Fp and LM-13-Fp on the innate immune response in zebrafish may be the existence of different antigenic group. Therefore, differences in serotype should be determined, and cross protection experiments have to be carried out in order to design a vaccine with broad-spectrum effectiveness.
In order to compare effects on the activation of the innate immune system for newly formulated vaccines (VAH and VAF LM-02-Fp) and the commercial Flavomune vaccine, the aforementioned assay was repeated and found that both newly formulated vaccines were largely more immunogenic than Flavomune vaccine. It is important to note that this commercial vaccine contains whole-cells from two Chilean F. psychrophilum strains inactivated by formaldehyde incubation (http://www.veterquimica.cl/contenido/vacuna-flavomune548).
The assay method applied in the present study permitted the analysis of the initial, yet essential, stage of efficient vaccine selection, which is the activation of cellular machinery that will promote antigen presentation and the innate immune response [32]. Results showed that both vaccines with adjuvants (strain LM-02-Fp and LM-13-Fp) exerted similar effects on the innate immune response, but only the LM-02-Fp strain was able to induce neutrophil migration without an adjuvant. From this, it can be speculated that the VAH/VAF LM-02-Fp vaccines could induce the generation of specific antibodies against F. psychrophilum. This isin contrast to theeffects triggered by VAH/VAF LM-13-Fp vaccines, which produced a nonspecific immune response that was mainly due to the adjuvant. These results suggest thatthe VAH and VAF LM-02-Fp vaccines would be a better candidate for vaccine formulation.
Finally, by using zebrafish as a model, another important outcome of this work was providing an in depth investigation on the mechanisms that control adaptive immunity in fish. Presently, fish vaccines are primarily developed according to mammalian knowledge about adaptive immunity, a scenario that might not be the most appropriate given the differences between fish and mammalian immune systems [52–55]. It is currently accepted that fish and particularly zebrafish share similarities with the mammalian immune system [19, 52, 56–58]. For example, both have a comparable set of immune cells [59–63], and the adaptive immune system is active later during embryonic development, after innate immune system development [25]. However, there are also differences as compared with mammalian immune responses, such as with the most abundant immunoglobulin in fish being IgM, whereas in mammals this is IgA [53, 64, 65]. Besides this, several important processes have not yet been described in fish, including antigen presentation and the existence of memory cells. In this regard, the different transgenic zebrafish lines with fluorescently labeled immune cells [30, 60, 61, 66–69] allow this teleost fish to act as a “live indicator” of immune processes, with the whole event of interest being monitored in vivo.
5. Conclusions
The VAH and VAF LM-02-Fp vaccine induced the strongest innate immune response, even when compared with the commercial Flavomune vaccine. However, after a year of storage at 4°C, the capacity of both formulated vaccines to trigger an innate immune response considerably decreased. Finally, the methodology described in this paper, using the zebrafish as fish model, is a suitable approach for analyzing mechanisms that control adaptive immunity in fish, as well pathways of bacterial infection and the innate immune response triggered by different vaccines.
Supplementary Material
Supplementary Figure 1: Effect on the innate immune response triggered by control medium. Plot of quantified neutrophil migration to the ROI. Statistical analysis was performed by two-way ANOVA. Data represent the mean ± standard error from three independent experiments with 15 larvae each. ∗∗∗∗P-value <0.0001.
Acknowledgments
This study was supported by Universidad Andrés Bello Núcleo DI-447-13/N, CONICYT/FONDAP/15110027, and FONDECYT 1120552. The authors thank Catalina Lafourcade for her expert fish care. The Tg(mpx:GFP) transgenic line was kindly provided by Dr. Steve Renshaw.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Bernardet J.-F., Segers P., Vancanneyt M., Berthe F., Kersters K., Vandamme P. Cutting a gordian knot: emended classification and description of the genus Flavobacterium, emended description of the family Flavobacteriaceae, and proposal of Flavobacterium hydatis nom. nov. (basonym, Cytophaga aquatilis Strohl and Tait 1978) International Journal of Systematic Bacteriology. 1996;46(1):128–148. doi: 10.1099/00207713-46-1-128. [DOI] [Google Scholar]
- 2.Nematollahi A., Decostere A., Pasmans F., Haesebrouck F. Flavobacterium psychrophilum infections in salmonid fish. Journal of Fish Diseases. 2003;26(10):563–574. doi: 10.1046/j.1365-2761.2003.00488.x. [DOI] [PubMed] [Google Scholar]
- 3.Barnes M. E., Brown M. L. A review of Flavobacterium psychrophilum biology, clinical signs, and bacterial cold water disease prevention and treatment. The Open Fish Science Journal. 2011;4(1):1–9. [Google Scholar]
- 4.Bustos P. A., Calbuyahue J., Montaña J., Opazo B., Entrala P., Solervisenc R. First isolation of Flexibacter psychrophilus, as causative agent of rainbow trout fry syndrome (RTFS), producing rainbow trout mortality in Chile. Bulletin of the European Association of Fish Pathologists. 1995;15:162–164. [Google Scholar]
- 5.Avendaño-Herrera R., Ilardi P., Fernández J. Significance of Flavobacterium diseases on salmonid farming in Chile. Proceedings of the 2nd Conference on Members of the Genus Flavobacterium; 2009; pp. 21–23. [Google Scholar]
- 6.Henríquez-Núñez H., Evrard O., Kronvall G., Avendaño-Herrera R. Antimicrobial susceptibility and plasmid profiles of Flavobacterium psychrophilum strains isolated in Chile. Aquaculture. 2012;354-355:38–44. doi: 10.1016/j.aquaculture.2012.04.034. [DOI] [Google Scholar]
- 7.Iwasaki A., Medzhitov R. Toll-like receptor control of the adaptive immune responses. Nature Immunology. 2004;5(10):987–995. doi: 10.1038/ni1112. [DOI] [PubMed] [Google Scholar]
- 8.Mantovani A., Cassatella M. A., Costantini C., Jaillon S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nature Reviews Immunology. 2011;11(8):519–531. doi: 10.1038/nri3024. [DOI] [PubMed] [Google Scholar]
- 9.Balázs M., Martin F., Zhou T., Kearney J. F. Blood dendritic cells interact with splenic marginal zone B cells to initiate T-independent immune responses. Immunity. 2002;17(3):341–352. doi: 10.1016/S1074-7613(02)00389-8. [DOI] [PubMed] [Google Scholar]
- 10.Scapini P., Nardelli B., Nadali G., et al. G-CSF-stimulated neutrophils are a prominent source of functional BLyS. Journal of Experimental Medicine. 2003;197(3):297–302. doi: 10.1084/jem.20021343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Schwaller J., Went P., Matthes T., et al. Paracrine promotion of tumor development by the TNF ligand APRIL in Hodgkin's Disease. Leukemia. 2007;21(6):1324–1327. doi: 10.1038/sj.leu.2404627. [DOI] [PubMed] [Google Scholar]
- 12.Puga I., Cols M., Barra C. M., et al. B cell-helper neutrophils stimulate the diversification and production of immunoglobulin in the marginal zone of the spleen. Nature Immunology. 2011;13(2):170–180. doi: 10.1038/ni.2194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Mócsai A. Diverse novel functions of neutrophils in immunity, inflammation, and beyond. The Journal of Experimental Medicine. 2013;210(7):1283–1299. doi: 10.1084/jem.20122220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cerutti A., Cols M., Puga I. Marginal zone B cells: virtues of innate-like antibody-producing lymphocytes. Nature Reviews Immunology. 2013;13(2):118–132. doi: 10.1038/nri3383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Swaim L. E., Connolly L. E., Volkman H. E., Humbert O., Born D. E., Ramakrishnan L. Mycobacterium marinum infection of adult zebrafish causes caseating granulomatous tuberculosis and is moderated by adaptive immunity. Infection and Immunity. 2006;74(11):6108–6117. doi: 10.1128/iai.00887-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Barut B. A., Zon L. I. Realizing the potential of zebrafish as a model for human disease. Physiological Genomics. 2000;2(2):49–51. doi: 10.1152/physiolgenomics.2000.2.2.49. [DOI] [PubMed] [Google Scholar]
- 17.Lieschke G. J., Currie P. D. Animal models of human disease: zebrafish swim into view. Nature Reviews Genetics. 2007;8(5):353–367. doi: 10.1038/nrg2091. [DOI] [PubMed] [Google Scholar]
- 18.Sullivan C., Kim C. H. Zebrafish as a model for infectious disease and immune function. Fish and Shellfish Immunology. 2008;25(4):341–350. doi: 10.1016/j.fsi.2008.05.005. [DOI] [PubMed] [Google Scholar]
- 19.van der Sar A. M., Appelmelk B. J., Vandenbroucke-Grauls C. M. J. E., Bitter W. A star with stripes: zebrafish as an infection model. Trends in Microbiology. 2004;12(10):451–457. doi: 10.1016/j.tim.2004.08.001. [DOI] [PubMed] [Google Scholar]
- 20.Laale H. W. Culture and preliminary observations of follicular isolates from adult zebra fish, Brachydanio rerio . Canadian Journal of Zoology. 1977;55(2):304–309. doi: 10.1139/z77-041. [DOI] [PubMed] [Google Scholar]
- 21.Kimmel C. B., Ballard W. W., Kimmel S. R., Ullmann B., Schilling T. F. Stages of embryonic development of the zebrafish. Developmental Dynamics. 1995;203(3):253–310. doi: 10.1002/aja.1002030302. [DOI] [PubMed] [Google Scholar]
- 22.Traver D., Herbomel P., Patton E. E., et al. The zebrafish as a model organism to study development of the immune system. Advances in Immunology. 2003;81:253–330. [PubMed] [Google Scholar]
- 23.Lieschke G. J., Trede N. S. Fish immunology. Current Biology. 2009;19(16):R678–R682. doi: 10.1016/j.cub.2009.06.068. [DOI] [PubMed] [Google Scholar]
- 24.Allen J. P., Neely M. N. Trolling for the ideal model host: zebrafish take the bait. Future Microbiology. 2010;5(4):563–569. doi: 10.2217/fmb.10.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lam S. H., Chua H. L., Gong Z., Lam T. J., Sin Y. M. Development and maturation of the immune system in zebrafish, Danio rerio: a gene expression profiling, in situ hybridization and immunological study. Developmental and Comparative Immunology. 2004;28(1):9–28. doi: 10.1016/s0145-305x(03)00103-4. [DOI] [PubMed] [Google Scholar]
- 26.Rogers H. W., Tripp C. S., Schreiber R. D., Unanue E. R. Endogenous IL-1 is required for neutrophil recruitment and macrophage activation during murine listeriosis. Journal of Immunology. 1994;153(5):2093–2101. [PubMed] [Google Scholar]
- 27.Pedrosa J., Saunders B. M., Appelberg R., Orme I. M., Silva M. T., Cooper A. M. Neutrophils play a protective nonphagocytic role in systemic Mycobacterium tuberculosis infection of mice. Infection and Immunity. 2000;68(2):577–583. doi: 10.1128/iai.68.2.577-583.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Appelberg R. Neutrophils and intracellular pathogens: beyond phagocytosis and killing. Trends in Microbiology. 2007;15(2):87–92. doi: 10.1016/j.tim.2006.11.009. [DOI] [PubMed] [Google Scholar]
- 29.Saitoh T., Komano J., Saitoh Y., et al. Neutrophil extracellular traps mediate a host defense response to human immunodeficiency virus-1. Cell Host & Microbe. 2012;12(1):109–116. doi: 10.1016/j.chom.2012.05.015. [DOI] [PubMed] [Google Scholar]
- 30.Renshaw S. A., Loynes C. A., Trushell D. M. I., Elworthy S., Ingham P. W., Whyte M. K. B. Atransgenic zebrafish model of neutrophilic inflammation. Blood. 2006;108(13):3976–3978. doi: 10.1182/blood-2006-05-024075. [DOI] [PubMed] [Google Scholar]
- 31.Avendaño-Herrera R., Houel A., Irgang R., et al. Introduction, expansion and coexistence of epidemic Flavobacterium psychrophilum lineages in Chilean fish farms. Veterinary Microbiology. 2014;170(3-4):298–306. doi: 10.1016/j.vetmic.2014.02.009. [DOI] [PubMed] [Google Scholar]
- 32.Nathan C. Neutrophils and immunity: challenges and opportunities. Nature Reviews Immunology. 2006;6(3):173–182. doi: 10.1038/nri1785. [DOI] [PubMed] [Google Scholar]
- 33.Bernardet J.-F., Nakagawa Y., Holmes B., et al. Proposed minimal standards for describing new taxa of the family Flavobacteriaceae and emended description of the family. International Journal of Systematic and Evolutionary Microbiology. 2002;52(3):1049–1070. doi: 10.1099/ijs.0.02136-0. [DOI] [PubMed] [Google Scholar]
- 34.Westerfield M. The Zebrafish Book: A Guide for the Laboratory Use of the Zebrafish (Daniorerio) 5th. Eugene, Ori, USA: University of Oregon Press; 1994. [Google Scholar]
- 35.Haffter P., Granato M., Brand M., et al. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio . Development. 1996;123:1–36. doi: 10.1242/dev.123.1.1. [DOI] [PubMed] [Google Scholar]
- 36.Barros-Becker F., Romero J., Pulgar A., Feijóo C. G. Persistent oxytetracycline exposure induces an inflammatory process that improves regenerative capacity in zebrafish larvae. PLoS ONE. 2012;7(5) doi: 10.1371/journal.pone.0036827.e36827 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bravo S., Midtlyng P. J. The use of fish vaccines in the Chilean salmon industry 1999–2003. Aquaculture. 2007;270(1–4):36–42. doi: 10.1016/j.aquaculture.2007.06.017. [DOI] [Google Scholar]
- 38.Gómez E., Méndez J., Cascales D., Guijarro J. A. Flavobacterium psychrophilum vaccine development: a difficult task. Microbial Biotechnology. 2014;7(5):414–423. doi: 10.1111/1751-7915.12099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Holt R. A. Cytophaga psychrophila, the causative agent of bacterial cold-water disease in salmonid fish [Ph.D. thesis] Corvallis, Ore, USA: Oregon State University; 1987. [Google Scholar]
- 40.Obach A., Baudin Laurencin F. Vaccination of rainbow trout Oncorhynchus mykiss against the visceral form of coldwater disease. Diseases of Aquatic Organisms. 1992;12:13–15. doi: 10.3354/dao012013. [DOI] [Google Scholar]
- 41.Lorenzen E. Studies on Flexibacter psychrophilus in Relation to Rainbow Trout Fry Syndrome (RTFS) [Ph.D. thesis] Copenhagen, Denmark: National Veterinary Laboratory, Aarhus & Royal Veterinary and Agricultural University; 1994. [Google Scholar]
- 42.Rahman M. H., Ototake M., Iida Y., Yokomizo Y., Nakanishi T. Efficacy of oil-adjuvanted vaccine for coldwater disease in ayu Plecoglossus altivelis . Fish Pathology. 2000;35(4):199–203. doi: 10.3147/jsfp.35.199. [DOI] [Google Scholar]
- 43.Rahman M. H., Kuroda A., Dijkstra J. M., Kiryu I., Nakanishi T., Ototake M. The outer membrane fraction of Flavobacterium psychrophilum induces protective immunity in rainbow trout and ayu. Fish and Shellfish Immunology. 2002;12(2):169–179. doi: 10.1006/fsim.2001.0362. [DOI] [PubMed] [Google Scholar]
- 44.Rahman M. H., Ototake M., Nakanishi T. Water-soluble adjuvants enhance the protective effect of Flavobacterium psychrophilum vaccines in ayu Plecoglossus altivelis . Fish Pathology. 2003;38(4):171–176. doi: 10.3147/jsfp.38.171. [DOI] [Google Scholar]
- 45.LaFrentz B. R., LaPatra S. E., Jones G. R., Congleton J. L., Sun B., Cain K. D. Characterization of serum and mucosal antibody responses and relative per cent survival in rainbow trout, Oncorhynchus mykiss (Walbaum), following immunization and challenge with Flavobacterium psychrophilum . Journal of Fish Diseases. 2002;25(12):703–713. doi: 10.1046/j.1365-2761.2002.00424.x. [DOI] [Google Scholar]
- 46.Madetoja J., Lönnström L.-G., Björkblom C., et al. Efficacy of injection vaccines against Flavobacterium psychrophilum in rainbow trout, Oncorhynchus mykiss (Walbaum) Journal of Fish Diseases. 2006;29(1):9–20. doi: 10.1111/j.1365-2761.2005.00676.x. [DOI] [PubMed] [Google Scholar]
- 47.Håstein T., Gudding R., Evensen Ø. Bacterial vaccines for fish: an update of the current situation world-wide. Developments in Biologicals (Basel) 2005;121:55–74. [PubMed] [Google Scholar]
- 48.Mata M., Skarmeta A., Santos Y. A proposed serotyping system for Flavobacterium psychrophilum . Letters in Applied Microbiology. 2002;35(2):166–170. doi: 10.1046/j.1472-765x.2002.01157.x. [DOI] [PubMed] [Google Scholar]
- 49.Lorenzen E., Olesen N. J. Characterization of isolates of Flavobacterium psychrophilum associated with coldwater disease or rainbow trout fry syndrome II: serological studies. Diseases of Aquatic Organisms. 1997;31(3):209–220. doi: 10.3354/dao031209. [DOI] [Google Scholar]
- 50.Izumi S., Wakabayashi H. Further study on serotyping of Flavobacterium psychrophilum . Fish Pathology. 1999;34(2):89–90. doi: 10.3147/jsfp.34.89. [DOI] [Google Scholar]
- 51.Valdebenito S., Avendaño-Herrera R. Phenotypic, serological and genetic characterization of Flavobacterium psychrophilum strains isolated from salmonids in Chile. Journal of Fish Diseases. 2009;32(4):321–333. doi: 10.1111/j.1365-2761.2008.00996.x. [DOI] [PubMed] [Google Scholar]
- 52.Meeker N. D., Trede N. S. Immunology and zebrafish: spawning new models of human disease. Developmental and Comparative Immunology. 2008;32(7):745–757. doi: 10.1016/j.dci.2007.11.011. [DOI] [PubMed] [Google Scholar]
- 53.Uribe C., Folch H., Enriquez R., Moran G. Innate and adaptive immunity in teleost fish: a review. Veterinarni Medicina. 2011;56(10):486–503. [Google Scholar]
- 54.Magnadóttir B., Jónsdóttir H., Helgason S., Björnsson B., Solem S. T., Pilström L. Immune parameters of immunised cod (Gadus morhua L.) Fish & Shellfish Immunology. 2001;11(1):75–89. doi: 10.1006/fsim.2000.0296. [DOI] [PubMed] [Google Scholar]
- 55.Magnadottir B., Gudmundsdottir S., Gudmundsdottir B. K., Helgason S. Natural antibodies of cod (Gadus morhua L.): specificity, activity and affinity. Comparative Biochemistry and Physiology—B Biochemistry and Molecular Biology. 2009;154(3):309–316. doi: 10.1016/j.cbpb.2009.07.005. [DOI] [PubMed] [Google Scholar]
- 56.Trede N. S., Langenau D. M., Traver D., Look A. T., Zon L. I. The use of zebrafish to understand immunity. Immunity. 2004;20(4):367–379. doi: 10.1016/s1074-7613(04)00084-6. [DOI] [PubMed] [Google Scholar]
- 57.Lin B., Cao Z., Lin Y., et al. Acute phase response in zebrafish upon Aeromonas salmonicida and Staphylococcus aureus infection: striking similarities and obvious differences with mammals. Molecular Immunology. 2007;44(4):295–301. doi: 10.1016/j.molimm.2006.03.001. [DOI] [PubMed] [Google Scholar]
- 58.Yoder J. A., Nielsen M. E., Amemiya C. T., Litman G. W. Zebrafish as an immunological model system. Microbes and Infection. 2002;4(14):1469–1478. doi: 10.1016/S1286-4579(02)00029-1. [DOI] [PubMed] [Google Scholar]
- 59.Lieschke G. J., Oates A. C., Crowhurst M. O., Ward A. C., Layton J. E. Morphologic and functional characterization of granulocytes and macrophages in embryonic and adult zebrafish. Blood. 2001;98(10):3087–3096. doi: 10.1182/blood.V98.10.3087. [DOI] [PubMed] [Google Scholar]
- 60.Wittamer V., Bertrand J. Y., Gutschow P. W., Traver D. Characterization of the mononuclear phagocyte system in zebrafish. Blood. 2011;117(26):7126–7135. doi: 10.1182/blood-2010-11-321448. [DOI] [PubMed] [Google Scholar]
- 61.Page D. M., Wittamer V., Bertrand J. Y., et al. An evolutionarily conserved program of B-cell development and activation in zebrafish. Blood. 2013;122(8):e1–e11. doi: 10.1182/blood-2012-12-471029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Lewis K. L., del Cid N., Traver D. Perspectives on antigen presenting cells in zebrafish. Developmental and Comparative Immunology. 2014;46(1):63–73. doi: 10.1016/j.dci.2014.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Zhang S., Cui P. Complement system in zebrafish. Developmental and Comparative Immunology. 2014;46(1):3–10. doi: 10.1016/j.dci.2014.01.010. [DOI] [PubMed] [Google Scholar]
- 64.Acton R. T., Weinheimer P. F., Hall S. J., Niedermeier W., Shelton E., Bennett J. C. Tetrameric immune macroglobulins in three orders of bony fishes. Proceedings of the National Academy of Sciences of the United States of America. 1971;68(1):107–111. doi: 10.1073/pnas.68.1.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Wilson M. R., Warr G. W. Fish immunoglobulins and the genes that encode them. Annual Review of Fish Diseases. 1992;2:201–221. doi: 10.1016/0959-8030(92)90064-5. [DOI] [Google Scholar]
- 66.Bertrand J. Y., Kim A. D., Teng S., Traver D. CD41+ cmyb+ precursors colonize the zebrafish pronephros by a novel migration route to initiate adult hematopoiesis. Development. 2008;135(10):1853–1862. doi: 10.1242/dev.015297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Langenau D. M., Ferrando A. A., Traver D., et al. In vivo tracking of T cell development, ablation, and engraftment in transgenic zebrafish. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(19):7369–7374. doi: 10.1073/pnas.0402248101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ellett F., Pase L., Hayman J. W., Andrianopoulos A., Lieschke G. J. Mpeg1 promoter transgenes direct macrophage-lineage expression in zebrafish. Blood. 2011;117(4):e49–e56. doi: 10.1182/blood-2010-10-314120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Hall C., Flores M., Storm T., Crosier K., Crosier P. The zebrafish lysozyme C promoter drives myeloid-specific expression in transgenic fish. BMC Developmental Biology. 2007;7, article 42 doi: 10.1186/1471-213x-7-42. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figure 1: Effect on the innate immune response triggered by control medium. Plot of quantified neutrophil migration to the ROI. Statistical analysis was performed by two-way ANOVA. Data represent the mean ± standard error from three independent experiments with 15 larvae each. ∗∗∗∗P-value <0.0001.