

Abstract
Members of the neuropeptide Y (NPY) family acting via Y2 and/or Y4 receptors have been proposed to participate in the control of ingestive behaviour and energy homeostasis. Since these processes vary between day and night, we explored the circadian patterns of locomotor, exploratory and ingestive behaviour in mice with disrupted genes for Y2 (Y2−/−) or Y4 (Y4−/−) receptors. To this end, the LabMaster system was used and its utility for the analysis of changes in circadian activity and ingestion caused by gene knockout evaluated. Female animals, aged 27 weeks on average, were housed singly in cages fitted with sensors for water and food intake and two infrared frames for recording ambulation and rearing under a 12 h light/dark cycle for 4 days. Relative to WT animals, diurnal locomotion, exploration, drinking and feeding were reduced, whereas nocturnal locomotion was enhanced in Y2−/− mice. In contrast, Y4−/− mice moved more but ate and drank less during the photophase, while they ate more and explored less during the scotophase. Both Y2−/− and Y4−/− mice weighed more than WT mice. These findings attest to a differential role of Y2 and Y4 receptor signalling in the circadian control of behaviours that balance energy intake and energy expenditure. These phenotypic traits can be sensitively and continuously recorded by the LabMaster system.
Keywords: Neuropeptide Y, Y2 receptors, Y4 receptors, circadian pattern, locomotion, exploration, feeding, drinking, energy intake, energy expenditure, energy homeostasis
INTRODUCTION
Neuropeptide Y (NPY) is one of the most widely distributed peptide messengers in the mammalian brain. Its many functional implications include the control of mood, anxiety, stress sensitivity, cognition, seizure activity and energy homeostasis (Eva et al., 2006; Heilig, 2004; Karl and Herzog, 2007; Kask et al., 2002; Lin et al., 2004; Vezzani et al., 1999). Haplotype-driven expression of NPY in humans predicts brain responses to emotional and stress challenges and inversely correlates with trait anxiety (Zhou et al., 2008). The physiological actions of NPY are mediated by several G protein-coupled receptors, five of which (Y1, Y2, Y4, Y5 and y6) have been elucidated at the gene and protein level (Michel et al., 1998; Redrobe et al., 2004a). Apart from NPY, Y receptors are also targeted by peptide YY (PYY) and pancreatic polypeptide. PYY is thought to be an intestinal satiety signal, and the major circulating form of this peptide, PYY(3-36), reduces food intake in rodents and humans primarily via binding to autoinhibitory Y2 receptors in the arcuate nucleus of the hypothalamus (McGowan and Bloom, 2004; Ueno et al., 2008).
The functional implications of Y receptors have been explored by gene knockout approaches and, where available, pharmacological antagonism of Y receptors. Anxiety-and depression-like behaviour is significantly reduced in Y2 receptor knockout (Y2−/−) mice (Redrobe et al., 2003; Tschenett et al., 2003), and a similar anxiolytic and antidepressant phenotype has been observed in Y4 receptor knockout (Y4−/−) mice (Painsipp et al., 2008; Tasan et al., 2009). Body weight (BW) and adiposity are decreased in one strain (Sainsbury et al., 2002a) but increased in another strain of Y2−/−mice (Naveilhan et al., 1999). Food intake is enhanced in either strain of Y2−/− mice (Lin et al., 2004; Naveilhan et al., 1999; Sainsbury et al., 2002a). In contrast, BW, adipose tissue mass and food intake are significantly reduced in Y4−/− mice (Sainsbury et al., 2002b).
Locomotion, exploration, water and food intake are activities subject to circadian regulation. Although Y2 receptor signalling has been found to modulate the circadian clock (Gribkoff et al., 1998; Yannielli and Harrington, 2001), the effect of Y2 and Y4 receptor gene knockouts on the circadian cycle of locomotion, exploration and ingestive behaviour has not yet been examined in a systematic manner. The LabMaster system (TSE Systems, Bad Homburg, Germany) makes it possible to record the circadian patterns of activity and ingestion for prolonged periods of time while the animals remain completely undisturbed. It was the first aim of this study to evaluate the utility and potential of this system in the circadian phenotyping of genetically modified mice. The second aim of the study was to characterize the circadian pattern of locomotion, exploration, drinking and feeding specifically in female Y2−/− and Y4−/− mice, relative to wild-type (WT) mice, in order to analyze how energy intake and energy expenditure are under the regulatory control of Y2 and Y4 receptors.
METHODS
Experimental animals
The study was conducted with female mice of the WT, Y2−/− and Y4−/− genotypes. They all were generated from the same founders on the same mixed C57BL/6:129/SvJ (50 %:50 %) background has been described previously (Sainsbury et al. 2002a, 2002b). The presence or deletion of Y2 and Y4 receptors was verified by polymerase chain reaction (Sainsbury et al., 2002a, 2002b). Homozygous animals were bred at the Institute of Pharmacology of the Medical University of Innsbruck. The breeding scheme included back-crossing of the knockout animals with WT mice approximately every 5th generation. After their transfer to Graz, the animals were allowed to acclimatize in the animal house of the Medical University of Graz for a minimum of 4 weeks. All experiments were carried out at the Institute of Experimental and Clinical Pharmacology of the Medical University of Graz.
Before the experiments, the animals were housed in groups of 2 – 5 per cage, whereas in the experiments they were kept singly in the test cages. In either case, the animals were maintained under controlled temperature (set point 24 °C), controlled relative air humidity (set point 50 %) and a 12 h light/dark cycle (lights on at 07:00 h, lights off at 19:00 h). All experiments were approved by an ethical committee at the Federal Ministry of Science and Research of the Republic of Austria and conducted according to the Directive of the European Communities Council of 24 November 1986 (86/609/EEC). The experiments were designed in such a way that the number of animals used and their suffering was minimized.
LabMaster system
The circadian pattern of locomotion, exploration, drinking and feeding was assessed with the LabMaster system (TSE Systems, Bad Homburg, Germany), which allowed continuous recording of these parameters (Theander-Carrillo et al., 2006) for up to 10 days while the animals remained undisturbed by any investigator. The system consisted of six recording units, each unit comprising a test cage (type III, 42 cm x 26.5 cm x 15 cm, length x width x height), two external infrared frames and a cage lid fitted with two weight transducers. These devices were connected to a personal computer which was used to collect and analyze the data with the LabMaster software. The hardware sampling rate at the infrared frames was 100 Hz, while that at the drinking and feeding sensors was 1 Hz. In contrast, the minimal sampling interval of the LabMaster software was 1 min, which means that the recordings taken by the hardware over 1 min (6,000 and 60, respectively) were summed up at 1 min intervals. In other terms, 720 values of each test parameter were collected over a 12 h interval.
The two weight transducers were employed to quantify ingestive behaviour. To this end, a feeding bin filled with standard rodent chow (altromin 1324 FORTI; Altromin, Lage, Germany) and a drinking bottle filled with tap water were each attached to a transducer on the cage lid, and the animals were allowed to drink and feed ad libitum. The drinking flasks were equipped with a special nipple that prevented the spontaneous leaking of water from the bottle. Water and food intake over time was measured in ml and g, respectively. For data analysis, the amount of water and food ingested over select time intervals was normalized to the BW of animals (ml/g BW, g/g BW).
For recording locomotion and exploration, the two external infra-red frames were positioned in a horizontal manner above one another at a distance of 4.3 cm, with the lower frame being fixed 2.0 cm above the bedding floor. The bottom frame was used to record horizontal locomotion (ambulatory movements) of the mice, while the top frame served to record vertical movements (rearing, exploration). The measures of activity (locomotion, exploration) were derived from the light beam interruptions (counts) of the corresponding infra-red frames. An ambulatory movement was defined as temporally subsequent interruption of any two different light beams in one axis, and the total locomotor activity was calculated by summing up the counts in both the x- and y-axes over select time intervals.
Experimental protocols
In order to enable the mice to adapt to the test room conditions, the group-housed animals were transferred to the test room at least one week before the experiments in the LabMaster system were started on day 0. Two to three days before day 0, the group-housed animals were also habituated to the drinking bottles used in the LabMaster system. On day 0, the mice were weighed and then placed singly in the test cages and maintained there for up to 4 days. The light intensity in the centre of the test cages during the photophase was 230 – 340 lux.
The transfer of the animals to the test cages took place in the morning or early afternoon of day 0. The remaining photophase and the following scotophase of day 0 were allowed for habituation of the mice to the novel environment and, for this reason, were not included in the statistical analysis of the results, although the data of scotophase 0 are shown to illustrate the circadian time course of the test parameters.
Given the multitude of values (720) collected for each test parameter during a 12 h interval, the results were subjected to the following data reduction procedure. First, the data for each test parameter and animal collected during the photophase of days 1 and 2 as well as the data collected during the scotophase of days 1 and 2 were summed up. The respective sum values for the photophase and scotophase on days 1 and 2 were then averaged for each animal. Finally, the mean values of each test parameter during the photophase and scotophase were used for statistical analysis of differences between the genotypes.
Statistics
Statistical analysis was performed on SPSS 15.0 (SPSS Inc., Chicago, IL, USA). Explorative data analysis revealed a violation of normality assumptions for most of the test parameters. Statistical analysis was therefore performed with non-parametric test procedures. Statistical differences among genotypes were determined with Kruskal-Wallis one-way ANOVA for the respective test parameters. Post-hoc analysis of group differences was performed with the Mann-Whitney U test, the P values being adjusted for multiple comparisons with the Bonferroni correction. In view of the exploratory nature of this study, P values ≤ 0.1 (Kirk, 1995) were considered to be statistically significant. All data are presented as means ± SEM, n referring to the number of mice in each group.
In order to test for any effect of the body weight on locomotion and exploration, a two-way analysis of covariance (ANCOVA) for repeated measures was carried out, in which the factors genotype and time (photophase and scotophase) were analyzed along with the covariable body weight. Although being parametric, this test was chosen because the violation of normality assumptions can be explained by the relatively small number of animals in each group and because the F statistics are relatively robust against this kind of violation (Kirk, 1995).
RESULTS
General observations
The experiments were carried out with adult mice as defined by Crawley (2000), i.e., mice aged 6.2 ± 0.2 months (mean ± SEM, n = 74). Their average BW was 23.3 ± 0.2 g (mean ± SEM, n = 74), but there was a significant difference in the BW between the three genotypes under study, both before and after the trials (Figure 1). Thus, Y2−/− and Y4−/− mice weighed significantly more than the respective WT mice at the beginning and end of the experiments. During the test session all animals lost weight to a similar extent, this change being similar in all three genotypes.
Figure 1.
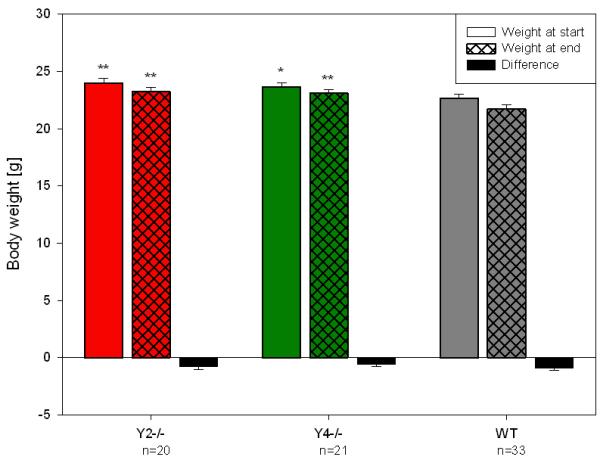
Body weight at the beginning (day 0) and end (day 4) of the experimental trial and weight loss during the experimental trial in WT, Y2−/− and Y4−/− mice. The values represent means ± SEM, n as indicated below the abscissa. * P ≤ 0.1, ** P ≤ 0.05 vs. WT mice (Mann-Whitney U test).
Locomotion and exploration
Circadian pattern
The locomotor (ambulatory) and exploratory (rearing) behaviour as recorded during the scotophase of days 0, 1 and 2 and the photophase of days 1 and 2 showed a characteristic circadian time course (Figure 2A,B). As was expected for nocturnal animals, the activity of the mice was considerably higher during the scotophase than during the photophase. While this circadian pattern of activity was seen in all genotypes, the magnitude of nocturnal activity differed with the genotype (Figures 2 and 3). It is worth noting hat the peak of nocturnal ambulation in all three genotypes tended to decrease over the course of the experiment (Figure 2A). This was also true for nocturnal rearing in Y4−/− mice, whereas in WT and Y2−/− mice nocturnal rearing tended to increase from day 0 to day 1 (Figure 2B). The different behaviour during the scotophase of day 0 was interpreted as a response to the novel environment of the test cages. For this reason, the observations made during the dark phase of day 0 were excluded from further analysis.
Figure 2.

Time course of the circadian (A) locomotor and (B) exploratory activity in WT, Y2−/− and Y4−/− mice. The graphs show the counts of light beam crossings summed up at intervals of 3 h for three consecutive dark phases (shaded areas, experimental days 0, 1 and 2) and two intervening light phases (white areas, experimental days 1 and 2). The values represent means ± SEM, n as indicated in brackets.
Figure 3.

Quantitative estimates of (A) locomotor and (B) exploratory activity during the photo- and scotophase in WT, Y2−/− and Y4−/− mice. The bars represent the counts of light beam crossings summed up and averaged for the photo- and scotophase, respectively, of experimental days 1 and 2. The values represent means ± SEM, n as indicated below the abscissa. * P ≤ 0.1, ** P ≤ 0.05 vs. WT mice, ++ P ≤ 0.05 vs. respective parameters in Y4−/− mice (Mann-Whitney U test).
Time course
Locomotor activity during the scotophase was markedly higher in Y2−/− mice than in WT and Y4−/− mice (Figure 2A). Ambulation peaked during the first quarter of the night in all genotypes, while the subsequent decline of ambulation was much more pronounced in Y2−/− mice than in WT and Y4−/− mice (Figure 2A). During the photophase locomotor activity was very low in all three genotypes (Figure 2B). It is noteworthy, however, that Y2−/− mice were the least active animals during the light phase whereas Y4−/− mice appeared to be most active ones (Figure 2A).
Like locomotion, nocturnal exploration peaked during the first quarter of the night and subsequently declined in both Y2−/− and Y4−/− mice, while in WT mice rearing was maintained at a plateau level during the first half of the scotophase (Figure 2B). The magnitude of nocturnal exploration was lowest in Y4−/− mice, whereas that in Y2−/− mice was similar to that in WT mice. Diurnal rearing activity was very low and indistinguishable between the three genotypes (Figure 2B).
Quantitative differences
Quantitative estimates of locomotor and exploratory activity during the photo- and scotophase in WT, Y2−/− and Y4−/− mice were obtained by summing up the counts of light beam crossings for the whole photophase and scotophase, respectively, and averaging the counts of experimental days 1 and 2.
Relative to WT mice, diurnal ambulation and rearing was significantly reduced in Y2−/− mice, whereas nocturnal locomotion was significantly enhanced and nocturnal rearing remained unchanged (Figure 3A,B). In contrast, Y4−/− mice moved more during the light phase and explored less during the dark phase, compared with WT mice, while their diurnal exploration and nocturnal locomotion stayed unchanged (Figure 3A,B). It also turned out that Y2−/− and Y4−/− mice differed from each other in all parameters of circadian ambulation and rearing (Figure 3A,B).
When the results were subjected to two-way ANCOVA for repeated measures, with the body weight measured at the beginning of the trials as covariable, the residual variance remained unchanged and the genotype-related differences in locomotion and exploration were preserved. It follows that the differences in ambulation and rearing between WT, Y2−/− and Y4−/− mice appeared to be independent of the body weight.
Ingestive behaviour
Time course
As was seen for locomotion and exploration, drinking and feeding followed a characteristic circadian pattern, with the highest activity occurring during the scotophase (Figure 4A,B). Although the circadian time course of water and food intake in the different genotypes under study showed considerable overlap, it is worth noting that the ingestive behaviour of WT and Y2−/− mice peaked roughly in the middle of the dark phase, whereas that of Y4−/− mice reached a maximum already in the first quarter of the night (Figure 4A,B). Of further note is the finding that the ingestive behaviour of WT mice increased progressively during the second half of the photophase, a process that was blunted in Y2−/− and Y4−/− mice (Figure 4A,B).
Figure 4.

Time course of the circadian (A) water and (B) food intake in WT, Y2−/− and Y4−/− mice. The graphs show the water and food consumption summed up at intervals of 3 h for three consecutive dark phases (shaded areas, experimental days 0, 1 and 2) and two intervening light phases (white areas, experimental days 1 and 2). Water consumption is expressed as ml/g BW, and food intake as g/g BW. The values represent means ± SEM, n as indicated in brackets.
Quantitative differences
Quantitative estimates of drinking and feeding during the photo- and scotophase in female WT, Y2−/− and Y4−/− mice were obtained by summing up the consumption rates for the whole photophase and scotophase, respectively, and averaging the counts of experimental days 1 and 2. The consumption rates were expressed relative to the BW determined at the beginning of the experiments.
Y2−/− mice ate and drank significantly less than WT mice during the photophase, whereas nocturnal ingestion tended to be increased, albeit without statistical difference (Figure 5A,B). In addition, the water intake of Y2−/− mice during the photophase was significantly less than that of Y4−/− mice. Compared with WT mice, diurnal drinking and feeding was likewise decreased in Y4−/− mice, whereas nocturnal feeding, but not drinking, was significantly enhanced in these animals (Figure 5A,B).
Figure 5.

Quantitative estimates of (A) water and (B) food intake during the photo- and scotophase in WT, Y2−/− and Y4−/− mice. The bars represent the water and food consumption summed up and averaged for the photo- and scotophase, respectively, of experimental days 1 and 2. Water consumption is expressed as ml/g BW, and food intake as g/g BW. The values represent means ± SEM, n as indicated below the abscissa. ** P ≤ 0.05 vs. WT mice, ++ P ≤ 0.05 vs. respective parameters in Y4−/− mice (Mann-Whitney U test).
Statistical analysis of the cumulative daily food intake failed to reveal any statistical difference between the three genotypes under study (data not shown).
DISCUSSION
General considerations
The overall aim of this study was to test the potential of the LabMaster system in the evaluation of changes in the circadian cycle of activity and ingestion caused by gene knockout. The specific aims were to explore whether the circadian pattern of locomotion, exploration, drinking and feeding differs between WT, Y2−/− and Y4−/− mice. The main results show that knockout of Y2 and Y4 receptors modifies locomotor, exploratory and ingestive behaviour in a differential circadian cycle-related manner. These data have a direct bearing on the proposed implications of the Y2 and Y4 receptor systems in energy homeostasis. By simultaneously recording feeding, drinking, locomotion and exploration it is possible to obtain information on the balance between energy intake and mobility-associated energy expenditure, parameters that are affected by appetite/satiety and have an impact on BW.
Considerable information on the implications of peptides acting via Y2 and/or Y4 receptors in the control of locomotion has been obtained from tests of emotional-affective behaviour (Painsipp et al., 2008; Redrobe et al., 2003; Tasan et al., 2009; Tschenett et al., 2003), many of which rely on the recording of locomotor and exploratory activity during a limited period of observation (Belzung and Griebel, 2001). However, these test paradigms represent stressors that are likely to bias the study results as, for instance, the anorectic effect of intraperitoneally injected PYY(3-36), a preferential Y2 receptor agonist, is inhibited by stress (Halatchev et al., 2004). To avoid these limitations, we recorded ingestive and motor behaviour continuously for several days while the animals were left undisturbed in the same cage.
While the LabMaster system allows to continuously record ingestive and motor activity and to analyze the circadian pattern of these activities, it has the disadvantage that the animals need be socially deprived and kept singly in the test cages. There is evidence for a gender-related difference in the reaction to single housing which appears to be a stressor for female, but not male, mice (Palanza et al., 2001). This issue need be kept in mind when the current results are compared with data in the literature, most of which were obtained in male animals. This instance and the observation that male and female Y2−/− mice differ in their food intake (Sainsbury et al., 2002a) were the reasons why we chose to examine female mice in the current study. In addition, the circadian locomotor behaviour of male Y2−/− and Y4−/− mice has been reported by Tasan et al. (2009). Since an important aspect of our study was to record the activity of the animals undisturbed by any investigator, the estrous cycle was not determined. We consider it unlikely that our data were significantly biased by this potentially confounding factor because the experiments were performed in the strict absence of any male mice and because we know that estrus synchronicity occurs in 53 – 80 % of female mice both within the same and across different cages (Painsipp et al., 2007).
There are several possibilities to explore the implications of the Y2 and Y4 receptor systems in ingestive and motor behaviour. The pharmacological approach involving selective receptor agonists and/or antagonists provides direct information on the acute biological effects of Y2 and Y4 receptor manipulation, but is limited if the long-term involvement of these receptor systems in energy homeostasis and BW is investigated. Apart from the disturbances that the administration of drugs per se may cause, the short-term effects of Y2 or Y4 receptor ligands may not be representative of the long-term implications of the Y2 and Y4 receptor systems in energy balance. For this reason, we chose a genetic approach involving germline Y2−/− and Y4−/− mice. Although the study data attest to distinct roles of the Y2 and Y4 receptor systems in energy homeostasis, it must not be neglected that developmental compensations may mask the full extent of their function. Another limitation of the present study may be the homozygous breeding scheme, although we think it unlikely that this factor introduced a significant source of error, given that the knockout animals were back-crossed with WT mice approximately every 5th generation.
Circadian locomotion and exploration
The circadian cycle of ambulation and rearing was differentially altered in female Y2−/− and Y4−/− mice and, importantly, there was a dissociation of the locomotor and exploratory activities. In addition, there were some distinct variations in the time course of ambulation and rearing. It is important to note that the current data reveal a gender difference, because locomotion of male Y2−/− and Y4−/− mice, averaged over 24 h, was found unchanged, although subtle differences in the time course of nocturnal ambulation between male control and Y4−/− mice were observed (Tasan et al., 2009).
The reduced locomotor and exploratory activity of female Y2−/− mice during the photophase is at variance with the behaviour which these animals display in the open field and on the elevated plus maze. In these tests, locomotion was found to be unchanged or enhanced (Painsipp et al., 2008; Redrobe et al., 2003, 2004b; Tasan et al., 2009; Tschenett et al., 2003), which is likely to represent a novelty-induced reaction. This conclusion is deduced from the finding that ambulation of female Y2−/− mice acutely exposed to the test cages of the LabMaster system on day 0 is much higher than on day 2 when the animals have habituated to the test cage environment (Painsipp et al., 2008). While Y2−/− mice acutely placed in the test cages do not differ from control mice in their locomotion, they move significantly less than control mice on day 2 (Painsipp et al., 2008) as was also found in the current study. Furthermore, the attenuation of diurnal exploration need be seen in the light of reduced attentional functioning and enhanced impulsivity in Y2−/− mice (Greco and Carli, 2006). Together with the finding that female Y2−/− mice moved more during the scotophase in the absence of any change in exploration, the current data indicate that Y2 receptor stimulation by endogenous ligands attenuates the circadian pattern of locomotor and exploratory activity. In addition, our observations emphasize that circadian phase is important to consider in the phenotyping of animals with regard to behaviours that undergo a circadian cycle (Beeler et al., 2006).
Unlike Y2−/− mice, female Y4−/− mice moved more during the light phase in the absence of a change in rearing, yet explored less during the dark phase without any change in nocturnal locomotion. Y4−/− mice of either gender display an immediate increase in locomotion when, during the photophase, they are exposed to the open field or elevated plus maze test paradigms or when they are placed in the test cages of the LabMaster system (Painsipp et al., 2008; Tasan et al., 2009). The current data indicate that this novelty-evoked increase in diurnal ambulation is maintained, although at a significantly lower level (Painsipp et al., 2008), following habituation. Taken all observations together, Y4 receptor signalling appears to be involved in the regulation of diurnal ambulation and nocturnal rearing, which is distinct from the role of Y2 receptor signalling in the circadian cycle of locomotion and exploration.
Ingestive behaviour and body weight
In parallel with reduced locomotion and exploration during the light phase, Y2−/−mice ate and drank significantly less than WT mice during the photophase. In contrast, the enhanced locomotion during the dark phase was associated only with a nominal trend towards increased ingestion, and the cumulative daily ingestion of water and food was not significantly altered. Since the circadian cycle-related changes in ingestive behaviour were associated with a significant increase in BW, it follows that the balance between energy intake and mobility-associated energy expenditure in Y2−/− mice is altered in favour of energy storage. This conclusion is of relevance to the proposed role of Y2 receptor signalling in the regulation of appetite and energy homeostasis (Lin et al., 2004; McGowan and Bloom, 2004). Y2 receptor agonists have been found to reduce food intake, metabolic rate and body weight gain during the light and dark phase (Balasubramaniam et al., 2007; Halatchev et al., 2004; McGowan and Bloom, 2004), but these results are not fully congruent with the outcome of the current and other Y2 receptor knockout studies. Naveilhan et al. (1999) reported that food intake and BW are enhanced in female Y2−/− mice aged 9-10 weeks, while their activity is reduced. In contrast, mice made deficient in Y2 receptors by the same deletion strategy on the same strain background as those used here were found to be underweight and less adipose despite an increase in cumulative daily food intake (Sainsbury et al., 2003, 2002a). The difference between these and the current findings are not fully understood but seem to be in part age- and gender-dependent. Thus, cumulative daily food intake was increased in female Y2−/− mice aged 8 – 12 weeks, while in male Y2−/− mice it was either enhanced at the age of 8 weeks or remained unchanged at the age of 8 - 16 weeks (Sainsbury et al., 2006, 2002a). The female mice used here were on average 27 weeks old, when cumulative daily food intake was no longer different from that of WT mice and BW was increased due to a shift towards energy storage.
Compared with WT mice, diurnal drinking and feeding in Y4−/− mice was decreased along with increased locomotion, whereas nocturnal feeding, but not drinking, was significantly enhanced along with attenuated exploration. Although cumulative daily intake of water and food remained unaltered, the BW of Y4−/− mice was significantly increased. These data suggest that Y4 receptor deletion, like Y2 receptor knockout, shifts the energy balance towards storage rather than expenditure. The effect of Y4 receptor knockout on ingestion and BW may also be age- and gender-dependent, given that particularly in male Y4−/− mice aged 16 weeks or less cumulative daily intake of food was found to be decreased concomitantly with a reduction in BW gain (Sainsbury et al., 2003, 2002b). In another study, however, feeding in male Y4−/− mice aged 16 weeks or less was enhanced in the absence of any BW change (Sainsbury et al., 2006), whereas in the female Y4−/− mice used here, aged 27 weeks on average, BW was enhanced in the absence of a significant change in cumulative daily food intake. The outcome of Y4 receptor knockout studies can only in part be reconciled with the ability of Y4 receptor agonists to inhibit food intake and body weight gain (Balasubramaniam et al., 2007; Halatchev et al., 2004; McGowan and Bloom, 2004). Since the circulating pancreatic polypeptide is elevated in both Y2−/− and Y4−/− mice, it has been hypothesized that the changes in energy homeostasis caused by Y2 or Y4 receptor knockout could in part be brought about by this preferential Y4 receptor agonist (Sainsbury et al., 2003, 2002a, 2002b).
During the test session all animals lost weight, which is likely to reflect that single housing is stressful to female mice (Palanza et al., 2001) and causes weight loss most likely by enhanced metabolic demand (van Leeuwen et al., 1997). The stress of single housing may also interfere with ingestive behaviour, given that nocturnal drinking in colony-housed female Y2−/− and Y4−/− mice is enhanced (Wultsch et al., 2006), whereas the increase in nocturnal drinking in single-housed Y2−/− and Y4−/− mice does not reach statistical significance as observed in the current study.
CONCLUSIONS
The current data reveal that knockout of the Y2 and Y4 genes alters the circadian pattern of locomotor, exploratory, drinking and feeding behaviour in a differential manner. These phenotypic traits can be sensitively and continuously recorded by the LabMaster system which in conjunction with alterations in BW provides information on the balance between energy intake, storage and expenditure. By analyzing the data of our study three major conclusions can be drawn. One, Y2 receptor stimulation by endogenous ligands attenuates the circadian pattern of locomotor and exploratory activity. Two, Y4 receptor signalling participates in the regulation of diurnal ambulation and nocturnal rearing, which is distinct from the role of Y2 receptor signalling. Three, Y2 and Y4 receptor signalling has an impact on circadian ingestion and energy homeostasis.
ACKNOWLEDGEMENTS
This study was supported by the Zukunftsfonds Steiermark (grant 262) and the Austrian Scientific Research Funds (FWF grant L25-B05). The authors thank Professor Andreas Tiran for his support to conduct the study at the Centre for Medical Research of the Medical University of Graz.
Grants:
Zukunftsfonds Steiermark (grant 262)
Austrian Scientific Research Funds (FWF grant L25-B05)
REFERENCES
- Balasubramaniam A, Joshi R, Su C, Friend LA, James JH. Neuropeptide Y (NPY) Y2 receptor-selective agonist inhibits food intake and promotes fat metabolism in mice: combined anorectic effects of Y2 and Y4 receptor-selective agonists. Peptides. 2007;28:235–40. doi: 10.1016/j.peptides.2006.08.041. [DOI] [PubMed] [Google Scholar]
- Beeler JA, Prendergast B, Zhuang X. Low amplitude entrainment of mice and the impact of circadian phase on behavior tests. Physiol Behav. 2006;87:870–80. doi: 10.1016/j.physbeh.2006.01.037. [DOI] [PubMed] [Google Scholar]
- Belzung C, Griebel G. Measuring normal and pathological anxiety-like behaviour in mice: a review. Behav Brain Res. 2001;125:141–9. doi: 10.1016/s0166-4328(01)00291-1. [DOI] [PubMed] [Google Scholar]
- Crawley JN. What’s Wrong With My Mouse?: Behavioral Phenotyping of Transgenic and Knockout Mice. 1st ed Wiley & Sons; 2000. [Google Scholar]
- Eva C, Serra M, Mele P, Panzica G, Oberto A. Physiology and gene regulation of the brain NPY Y1 receptor. Front Neuroendocrinol. 2006;27:308–39. doi: 10.1016/j.yfrne.2006.07.002. [DOI] [PubMed] [Google Scholar]
- Greco B, Carli M. Reduced attention and increased impulsivity in mice lacking NPY Y2 receptors: relation to anxiolytic-like phenotype. Behav Brain Res. 2006;169:325–34. doi: 10.1016/j.bbr.2006.02.002. [DOI] [PubMed] [Google Scholar]
- Gribkoff VK, Pieschl RL, Wisialowski TA, van den Pol AN, Yocca FD. Phase shifting of circadian rhythms and depression of neuronal activity in the rat suprachiasmatic nucleus by neuropeptide Y: mediation by different receptor subtypes. J Neurosci. 1998;18:3014–22. doi: 10.1523/JNEUROSCI.18-08-03014.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halatchev IG, Ellacott KLJ, Fan W, Cone RD. Peptide YY3-36 inhibits food intake in mice through a melanocortin-4 receptor-independent mechanism. Endocrinology. 2004;145:2585–90. doi: 10.1210/en.2003-1754. [DOI] [PubMed] [Google Scholar]
- Heilig M. The NPY system in stress, anxiety and depression. Neuropeptides. 2004;38:213–24. doi: 10.1016/j.npep.2004.05.002. [DOI] [PubMed] [Google Scholar]
- Karl T, Herzog H. Behavioral profiling of NPY in aggression and neuropsychiatric diseases. Peptides. 2007;28:326–33. doi: 10.1016/j.peptides.2006.07.027. [DOI] [PubMed] [Google Scholar]
- Kask A, Harro J, von Hörsten S, Redrobe JP, Dumont Y, Quirion R. The neurocircuitry and receptor subtypes mediating anxiolytic-like effects of neuropeptide Y. Neurosci Biobehav Rev. 2002;26:259–83. doi: 10.1016/s0149-7634(01)00066-5. [DOI] [PubMed] [Google Scholar]
- Kirk RE. Experimental Design: Procedures for Behavioral Sciences. 3rd ed Wadsworth: 1995. [Google Scholar]
- Lin S, Boey D, Herzog H. NPY and Y receptors: lessons from transgenic and knockout models. Neuropeptides. 2004;38:189–200. doi: 10.1016/j.npep.2004.05.005. [DOI] [PubMed] [Google Scholar]
- McGowan BMC, Bloom SR. Peptide YY and appetite control. Curr Opin. 2004 doi: 10.1016/j.coph.2004.06.007. [DOI] [PubMed] [Google Scholar]
- Michel MC, Beck-Sickinger A, Cox H, Doods HN, Herzog H, Larhammar D, Quirion R, Schwartz T, Westfall T. XVI. International Union of Pharmacology recommendations for the nomenclature of neuropeptide Y, peptide YY, and pancreatic polypeptide receptors. Pharmacol Rev. 1998;50:143–50. [PubMed] [Google Scholar]
- Naveilhan P, Hassani H, Canals JM, Ekstrand AJ, Larefalk A, Chhajlani V, Arenas E, Gedda K, Svensson L, Thoren P, Ernfors P. Normal feeding behavior, body weight and leptin response require the neuropeptide Y Y2 receptor. Nat Med. 1999;5:1188–93. doi: 10.1038/13514. [DOI] [PubMed] [Google Scholar]
- Painsipp E, Wultsch T, Edelsbrunner ME, Tasan RO, Singewald N, Herzog H, Holzer P. Reduced anxiety-like and depression-related behavior in neuropeptide Y Y4 receptor knockout mice. Genes Brain Behav. 2008;7:532–42. doi: 10.1111/j.1601-183X.2008.00389.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Painsipp E, Wultsch T, Shahbazian A, Edelsbrunner M, Kreissl MC, Schirbel A, Bock E, Pabst MA, Thoeringer CK, Huber HP, Holzer P. Experimental gastritis in mice enhances anxiety in a gender-related manner. Neuroscience. 2007;150:522–536. doi: 10.1016/j.neuroscience.2007.09.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palanza P, Gioiosa L, Parmigiani S. Social stress in mice: gender differences and effects of estrous cycle and social dominance. Physiol Behav. 2001;73:411–20. doi: 10.1016/s0031-9384(01)00494-2. [DOI] [PubMed] [Google Scholar]
- Redrobe JP, Dumont Y, Herzog H, Quirion R. Neuropeptide Y (NPY) Y2 receptors mediate behaviour in two animal models of anxiety: evidence from Y2 receptor knockout mice. Behav Brain Res. 2003;141:251–5. doi: 10.1016/s0166-4328(02)00374-1. [DOI] [PubMed] [Google Scholar]
- Redrobe JP, Carvajal C, Kask A, Dumont Y, Quirion R. Neuropeptide Y and its receptor subtypes in the central nervous system: emphasis on their role in animal models of psychiatric disorders. In: Michel MC, editor. Handbook of Experimental Pharmacology: Neuropeptide Y and Related Peptides: Bd 162. 1st ed Springer; Berlin: 2004a. pp. 101–36. [Google Scholar]
- Redrobe JP, Dumont Y, Herzog H, Quirion R. Characterization of neuropeptide Y, Y(2) receptor knockout mice in two animal models of learning and memory processing. J Mol Neurosci. 2004b;22:159–66. doi: 10.1385/JMN:22:3:159. [DOI] [PubMed] [Google Scholar]
- Sainsbury A, Baldock PA, Schwarzer C, Ueno N, Enriquez RF, Couzens M, Inui A, Herzog H, Gardiner EM. Synergistic effects of Y2 and Y4 receptors on adiposity and bone mass revealed in double knockout mice. Mol Cell Biol. 2003;23:5225–33. doi: 10.1128/MCB.23.15.5225-5233.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sainsbury A, Bergen HT, Boey D, Bamming D, Cooney GJ, Lin S, Couzens M, Stroth N, Lee NJ, Lindner D, Singewald N, Karl T, Duffy L, Enriquez R, Slack K, Sperk G, Herzog H. Y2Y4 receptor double knockout protects against obesity due to a high-fat diet or Y1 receptor deficiency in mice. Diabetes. 2006;55:19–26. [PubMed] [Google Scholar]
- Sainsbury A, Schwarzer C, Couzens M, Fetissov S, Furtinger S, Jenkins A, Cox HM, Sperk G, Hökfelt T, Herzog H. Important role of hypothalamic Y2 receptors in body weight regulation revealed in conditional knockout mice. Proc Natl Acad Sci U S A. 2002a;99:8938–43. doi: 10.1073/pnas.132043299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sainsbury A, Schwarzer C, Couzens M, Jenkins A, Oakes SR, Ormandy CJ, Herzog H. Y4 receptor knockout rescues fertility in ob/ob mice. Genes Dev. 2002b;16:1077–88. doi: 10.1101/gad.979102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tasan RO, Lin S, Hetzenauer A, Singewald N, Herzog H, Sperk G. Increased novelty-induced motor activity and reduced depression-like behavior in neuropeptide Y (NPY)-Y4 receptor knockout mice. Neuroscience. 2009;158:1717–30. doi: 10.1016/j.neuroscience.2008.11.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Theander-Carrillo C, Wiedmer P, Cettour-Rose P, Nogueiras R, Perez-Tilve D, Pfluger P, Castaneda TR, Muzzin P, Schürmann A, Szanto I, Tschöp MH, Rohner-Jeanrenaud F. Ghrelin action in the brain controls adipocyte metabolism. J Clin Invest. 2006;116:1983–93. doi: 10.1172/JCI25811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tschenett A, Singewald N, Carli M, Balducci C, Salchner P, Vezzani A, Herzog H, Sperk G. Reduced anxiety and improved stress coping ability in mice lacking NPY-Y2 receptors. Eur J Neurosci. 2003;18:143–8. doi: 10.1046/j.1460-9568.2003.02725.x. [DOI] [PubMed] [Google Scholar]
- Ueno H, Yamaguchi H, Mizuta M, Nakazato M. The role of PYY in feeding regulation. Regul Pept. 2008;145:12–6. doi: 10.1016/j.regpep.2007.09.011. [DOI] [PubMed] [Google Scholar]
- van Leeuwen SD, Bonne OB, Avraham Y, Berry EM. Separation as a new animal model for self-induced weight loss. Physiol Behav. 1997;62:77–81. doi: 10.1016/s0031-9384(97)00144-3. [DOI] [PubMed] [Google Scholar]
- Vezzani A, Sperk G, Colmers WF. Neuropeptide Y: emerging evidence for a functional role in seizure modulation. Trends Neurosci. 1999;22:25–30. doi: 10.1016/s0166-2236(98)01284-3. [DOI] [PubMed] [Google Scholar]
- Wultsch T, Painsipp E, Donner S, Sperk G, Herzog H, Peskar BA, Holzer P. Selective increase of dark phase water intake in neuropeptide-Y Y2 and Y4 receptor knockout mice. Behav Brain Res. 2006;168:255–60. doi: 10.1016/j.bbr.2005.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yannielli PC, Harrington ME. Neuropeptide Y in the mammalian circadian system: effects on light-induced circadian responses. Peptides. 2001;22:547–56. doi: 10.1016/s0196-9781(01)00356-4. [DOI] [PubMed] [Google Scholar]
- Zhou Z, Zhu G, Hariri AR, Enoch M, Scott D, Sinha R, Virkkunen M, Mash DC, Lipsky RH, Hu X, Hodgkinson CA, Xu K, Buzas B, Yuan Q, Shen P, Ferrell RE, Manuck SB, Brown SM, Hauger RL, Stohler CS, Zubieta J, Goldman D. Genetic variation in human NPY expression affects stress response and emotion. Nature. 2008;452:997–1001. doi: 10.1038/nature06858. [DOI] [PMC free article] [PubMed] [Google Scholar]