

Abstract
Background
Given that most species that have ever existed on earth are extinct, it stands to reason that the evolutionary history can be better understood with fossil taxa. Bauhinia is a typical genus of pantropical intercontinental disjunction among the Asian, African, and American continents. Geographic distribution patterns are better recognized when fossil records and molecular sequences are combined in the analyses. Here, we describe a new macrofossil species of Bauhinia from the Upper Miocene Xiaolongtan Formation in Wenshan County, Southeast Yunnan, China, and elucidate the biogeographic significance through the analyses of molecules and fossils.
Results
Morphometric analysis demonstrates that the leaf shapes of B. acuminata, B. championii, B. chalcophylla, B. purpurea, and B. podopetala closely resemble the leaf shapes of the new finding fossil. Phylogenetic relationships among the Bauhinia species were reconstructed using maximum parsimony and Bayesian inference, which inferred that species in Bauhinia species are well-resolved into three main groups. Divergence times were estimated by the Bayesian Markov chain Monte Carlo (MCMC) method under a relaxed clock, and inferred that the stem diversification time of Bauhinia was ca. 62.7 Ma. The Asian lineage first diverged at ca. 59.8 Ma, followed by divergence of the Africa lineage starting during the late Eocene, whereas that of the neotropical lineage starting during the middle Miocene.
Conclusions
Hypotheses relying on vicariance or continental history to explain pantropical disjunct distributions are dismissed because they require mostly Palaeogene and older tectonic events. We suggest that Bauhinia originated in the middle Paleocene in Laurasia, probably in Asia, implying a possible Tethys Seaway origin or an “Out of Tropical Asia”, and dispersal of legumes. Its present pantropical disjunction resulted from disruption of the boreotropical flora by climatic cooling after the Paleocene-Eocene Thermal Maximum (PETM). North Atlantic land bridges (NALB) seem the most plausible route for migration of Bauhinia from Asia to America; and additional aspects of the Bauhinia species distribution are explained by migration and long distance dispersal (LDD) from Eurasia to the African and American continents.
Keywords: Bauhinia, Pantropical intercontinental disjunction, Evolution, Biogeography, Paleocene-Eocene thermal maximum, Boreotropical flora, Long distance dispersal
Background
Pantropical intercontinental disjunction is an interesting biogeographic pattern in angiosperms, and is common to several tropical and subtropical genera and families [1],[2]. However, it still remains poorly understood compared with temperate disjunctions due to greater species richness, inaccessibility of study material [3], greater ocean separation, and the vast latitudinal distribution of taxa [2]. Many pantropical taxa are hypothesized to have spanned the Northern Hemisphere during the Paleocene-Eocene Thermal Maximum (PETM), because the warmer climate of the early Palaeogene allowed thermophilic taxa to extend their ranges northward [4]. The exchange of floristic elements among pantropical regions was possible through the North Atlantic land bridges (NALB) and the Bering land bridges (BLB) during the early Eocene [5]-[8]. A “boreotropical” connection across the North Atlantic during the Eocene has long been viewed as a key to understand the disjunction patterns in the Northern Hemisphere, such as the close relationships between Eastern Asian-North American plants [9], which have been extensively studied [10]. Recently, molecular phylogenetic studies combined with molecular clock inferences have allowed a more precise understanding of the process of dispersal, and have hypothesized the divergence times of pantropical distribution patterns for many plant families, such as Melastomataceae [3],[11], Malpighiaceae [8], Annonaceae [12],[13], Myristicaceae [12], Burseraceae [14], Rubiaceae [15], Simaroubaceae [16], and Sapotaceae [1]. Among the above-mentioned studies, the results suggest that modern pantropical disjunction mostly resulted from ancestral boreotropical distribution which was disrupted by late Eocene climatic cooling. That caused the distribution ranges of plant species to shrink to lower and warmer paleolatitudes for the sake of survival, followed by migrations from North to South America [17]-[20].
Explanations for wide geographic ranges of pantropical lineages in Eurasia, America, and Africa, typically invoke three main hypotheses: (1) The vicariance hypothesis developed with the acceptance of plate tectonics theory, which has been used to explain the wide distribution lineages on portions of the ancient Gondwana continent [2],[21]. As a biogeographic mechanism, it has been proposed for the relatively older family, such as Annonaceae [12]. (2) The boreotropical migration hypothesis describes the migration of some tropical lineages between the Old and the New Worlds via the NALB or BLB during the early Tertiary Period, when the climate conditions in the Northern Hemisphere could accommodate tropical vegetations [8],[22]. This hypothesis has been proposed as an explanation for the distribution of several lineages with a classical western Gondwanan disjunction pattern, such as Burseraceae [14]. (3) Long distance dispersal (LDD) has also been proposed as a process that has significantly shaped the formation of modern biotas. This hypothesis is used, especially when the divergence times of lineages are far too young, to be explained by vicariance via tectonic plate movements [8],[21],[23]. Despite the existence of this conceptual framework, there are still relatively few well-resolved biogeographic studies on pantropical clades.
Bauhinia L., is one of the largest genera in subfamily Caesalpinioideae (Leguminosae), comprising approximately 300 morphologically variable species of trees, shrubs, and lianas [24],[25]. It is distributed in tropical to subtropical and warm-temperate Asia, and tropical regions of Africa and the America (Figure 1). Recent studies have been revealed that the Caesalpinioideae are an early offshoot of the Leguminosae [24]-[31]. Recently, Bauhinia leaves were found in the Upper Miocene deposits from Wenshan, Southeast Yunnan, China. We used morphometrics to investigate the leaf shapes of Bauhinia and to compare the leaf shapes of extant and fossil species. Based on this new fossil record, we combined both molecular and fossil data to investigate the biogeographic history of Bauhinia.
Figure 1.
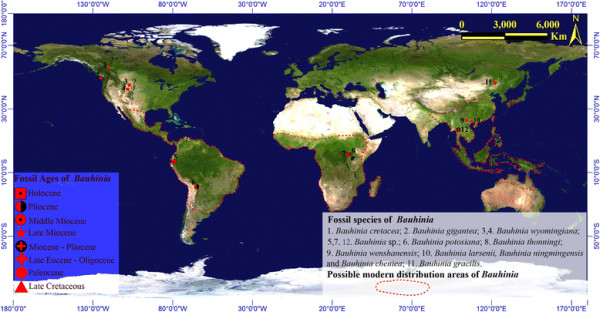
Map showing the locations from whichBauhiniafossils were recovered and the modern distribution ofBauhinia.“?” indicates questionable fossil records. The drawing of map is completed by the authors, and map background is courtesy of National Administration of Surveying, Mapping and Geoinformation.
Several molecular studies have focused on the phylogenetic relationships [25],[26],[32] and diversification of the legumes [33],[34]. Setting the stem node of legumes hypothetically at 60 Ma, the estimated age of the Leguminosae crown node is at 59 Ma [34]. However, the biogeographic history of Bauhinia is still not well resolved. A biogeographic history of Bauhinia should consider well-documented fossils and take into consideration dispersal in different continents. Information from pantropical flora is relatively limited, and divergence times are still unclear without fossil evidence. Well-dated, properly identified fossils and reliable phylogenetic reconstructions are crucial to reveal geographic distribution patterns. Fossils can provide direct evidence on origin time and migration or dispersal pathways to some extent, which are of special interest in biogeographic discussions when combined with molecular analyses.
In this study, we describe a new fossil species of Bauhinia, and use morphometrics to compare it with other species. Then, we combine fossil records and molecular analyses to discuss the timing of lineage diversifications and the biogeographic history of Bauhinia.
Methods
Geological setting
Fossil leaves of Bauhinia were collected in Wenshan County, Southeast Yunnan, China (Figure 2). The fossil-bearing strata belong to the Xiaolongtan Formation [35]. Specimens of Bauhinia were preserved as compressions in marlstone and shaly siltstone from several layers of the same strata (Figure 3). The Xiaolongtan Formation is widely distributed in Central and Southeast Yunnan. Its age is assigned to the Upper Miocene Epoch according to stratigraphic correlation, palynological study [36], mammal fossils [37], and plant macrofossils [38],[39].
Figure 2.
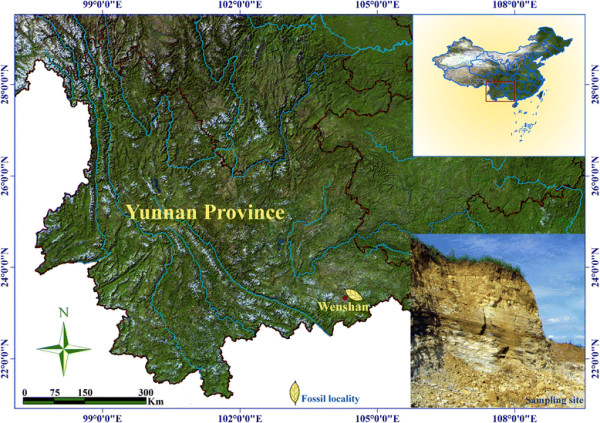
Map showing the location of fossil locality in Wenshan, Southeast Yunnan, China. The fossil site, labeled by a yellow leaf, is approximately 3–4 km southeast of Wenshan County. The drawing of map is completed by the authors, and map background is courtesy of National Administration of Surveying, Mapping and Geoinformation.
Figure 3.
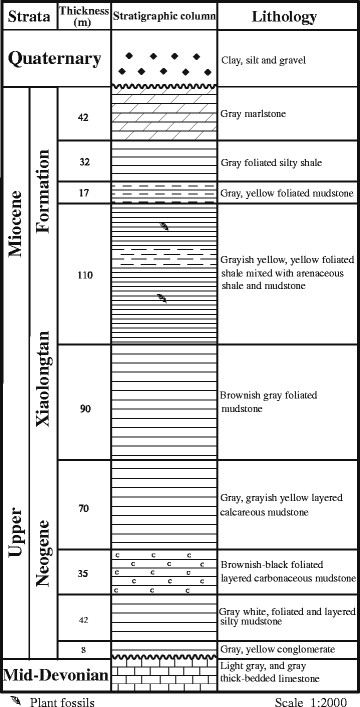
Stratigraphy of Wenshan fossil locality. The fossil leaves of Bauhinia wenshanensis were collected from a grayish-yellow, foliated shale mixed with arenaceous shale and mudstone.
Bauhinia fossil records
Bauhinia fossils have been widely reported from the late Cretaceous period to the Quaternary period [40]-[52], e.g., in Asia, Africa, North America (Table 1; Figure 1), and some wood fossils from India [53]. In Europe the only Bauhinia fossil record is B. olympica from Greece [54]. Early reports of Bauhinia fossils include the Cretaceous Bauhinia cretacea and B. gigantea from Canada and B. wyomingiana from the Cretaceous stratum of the USA [40],[42],[46], however, these Bauhinia records are not credible and their identification is doubtful, because they were not carefully or completely documented, or the samples only consisted of leaf fragments. We did not use B. gracilis as a calibration point of the root node, although it seems the oldest-known fossil records [50]. Because the holotype is a leaf with glandular teeth (Personal opinion, it may belong to Trochodendroides), rather than being an entire-margined leaf of Bauhinia. The original author apparently was confused by some insect damage to the leaf apex, giving a bilobed appearance vaguely similar to undamaged leaves of Bauhinia. But we can set the stem node of legumes hypothetically at 60 Ma to calibrate the phylogeny by following previous molecular research [34]. Extinct Bauhinia with confident identification includes the well-preserved B. larsenii, B. ningmingensis and B. cheniae from the late Eocene or early Oligocene of Guangxi in China, which has clear leaves or connect to a branch and a pod [44],[51]; and our new fossil, which has a clear leaf architecture (Table 1).
Table 1.
Previous published Bauhinia fossils and the Bauhinia fossils utilized in the present study
| Fossil of Bauhinia | Age (Ma) | Locality/Coordinates | Comments | Refs |
|---|---|---|---|---|
| Bauhinia gigantea | 70.6-89.3 | Canada/W125°00′00",N49°45'00" | No photograph, without confidence | [40] |
| Bauhinia wyomingiana | 56.8-60.2 | U. S. A/W108°06'52", N42°16'44" | Leaf fragments, without confidence | [49] |
| Bauhinia wyomingiana | 55.8-60.5 | U. S. A/W107°10'00", N44°55'02" | Leaf fragments, without confidence | [46] |
| Bauhinia cretacea | 93.5-99.6 | Canada/W120°07'57", N56°13'02" | No photograph, without confidence | [41] |
| Bauhinia sp. | 0-1.8 | Congo/E29°49'23", N1°12'48" | Leaf, confidence in identification | [48] |
| Bauhinia thonningi | 1.8-23 | Uganda/E31°00'00", N1°30'00" | Leaf, confidence in identification | [45] |
| Bauhinia potosiana | 1.8-5.3 | Bolivia/E65°00'00", N19°00'00" | Leaf, confidence in identification | [42],[43] |
| Bauhinia sp. | 11.6-15.9 | Ecuador/W79°06'00", S3°23'56" | Leaf, confidence in identification | [47] |
| Bauhinia larsenii | 28.4-37.2 | China/E107°02'14", N22°07'41" | Branch, confidence in identification | [44] |
| Bauhinia gracilis | 65.5-61.1 | China/E130°25'28", N48°53'04" | Leaf fragments, without confidence | [50] |
| Bauhinia sp. | 15.97-11.6 | Thailand/Mae Sot, Changwat Tak | Leaf, confidence in identification | [52] |
| Bauhinia ningmingensis and B. cheniae | 28.4-37.2 | China/E107°02'14", N22°07'41" | Leaf and Branch, confidence in identification | [51] |
| Bauhinia wenshanensis | 5.3-11.6 | China/E104°17'19", N23°20'50" | Leaf, confidence in identification | Present study |
Specimen preparation and geometric morphometric analyses
The gross morphology of new finding fossils was compared with leaves of all the extant species of Bauhinia represented in Chinese National Herbarium, Institute of Botany, CAS (PE); Herbarium, Kunming Institute of Botany, CAS (KUN); and Herbarium of Xishuangbanna Tropical Botanical Garden, CAS (HITBC). We also consulted the relevant literatures about the description of Bauhinia leaves, such as Flora of China[55] and Flora Malesiana[56].
Images of fossil specimens were captured using a Nikon D500 Digital SLR Camera. Several images of extant species were provided by Panasonic DMC-FZ30 in herbaria. The figured specimens, and associated pictures, are deposited in paleoecology group, Xishuangbanna Tropical Botanical Garden, CAS.
Morphometric approaches described the shapes of the specimens quantitatively. We apply geometric morphometrics to investigate foliage-shape in Bauhinia as employed in Menispermaceae endocarps [57]. Shapes were measured for 538 extant specimens (Specimen images are from the Chinese Virtual Herbarium (CVH) at http://www.cvh.org.cn/cms/, and the Muséum national d”Histoire naturelle (P, PC), at http://www.mnhn.fr/le-museum/) (Additional file 1: Table S1), representing 81 species in East-South Asian, American and African continents, and several fossil specimens. The leaf shapes were digitized using tpsDig [58]. The contour of each leaf was divided by four lines along the leaf edge. The first of these was drawn from the pulvinus to the right lobe apex, the second from the right lobe apex to the midvein end point, the third from the midvein end point to the left lobe apex, and the fourth from the left lobe apex to the pulvinus. The pulvinus to the left, right lobe apex was described with 15 equidistant points, and the lobe apex to the midvein point with five equidistant points. All points, except end points, were analyzed as semilandmarks; there were a total of four landmarks and 36 semilandmarks. All landmarks were of type I [59]. Slider vectors were defined as the chord between the previous and next points on the curve using tpsUtil [58]. Consensus shapes were combined and partial warps and partial warp scores were calculated using tpsRelw [60]. Then, we used PAST [61] to perform principal component analysis (PCA) and cluster analysis (CA) on the partial warp scores, to explore the distribution of species in the shape space.
Phylogeny and divergence times
The chloroplast gene tRNA-Leu (trnL) and the trnL-trnF intergenic spacer of 35 Bauhinia species and two outgroups (i.e., Cercis chinensis and C. canadensis) were downloaded from GenBank (Additional file 2: Table S2). The sequences were edited using SeqMan (Lasergene, DNASTAR Inc., Madison, Wisconsin, USA). Multiple sequence alignment was carried out in Clustal X 1.81 [62], and checked visually, refined and adjusted manually.
The combined data set was analyzed using parsimony in PAUP* 4.0b10 [63] with heuristic searching. Starting trees were obtained via stepwise addition, tree bisection reconnection branch swapping, steepest descent, and with the MulTrees and Collapse options in effect, as well as no upper limit for the number of trees held in memory. Support values for all nodes (on a 50% majority rule bootstrap tree) were calculated with the same settings as above for 1,000 replicates; 10 searches with random taxon additions were conducted for each replicate, and the strict consensus tree of all the shortest trees was saved. Bayesian inference relied on MrBayes 3.1.2 [64] and the GTR + I + G model as suggested by Modeltest 3.7 [65]. We used the default of one cold and three heated Markov chain Monte Carlo chains (MCMC), starting from random initial trees, and chains were run for 6,000,000 generations, with sampling every 200th. The default options in MrBayes were used for chain heating and mixing. We discarded a burn-in of the first 2,000,000 generations and used 20,000 trees from the posterior distribution to obtain a majority rule consensus tree.
Divergence times were estimated by BEAST 1.5.4 [66]. The BEAUti interface was used to create an input file for BEAST, in which a general time reversible (GTR) nucleotide substitution model with gamma + invariant sites was applied; an uncorrelated lognormal model was used to describe the relaxed clock. Ten million generations of the MCMC chains were run, with sampling every 1,000th. After discarding the first 1,000 trees as burn in, the samples were summarized in the maximum clade credibility tree using TreeAnnotator 1.4.8 [66], with the posterior probability limit set to 0.5, and summarizing mean node heights. Final trees were evaluated and edited in FigTree 1.3.1 [67]. Statistical support for the clades was determined by assessing Bayesian posterior probabilities. Substitution rates and the 95% highest posterior densities (HPDs) were determined with Tracer in combined runs. Divergence times are given as the mean and the 95% HPDs in millions of years, and the 95% HPDs intervals define the precision of estimation. We used the fossils to calibrate molecular dating, the stem node of legumes hypothetically at 60 Ma in molecular dating as previous molecular research [34]. The new, well-preserved, late Miocene fossil B. wenshanensis is hypothetically dated at ca.11.6 Ma according to the International Stratigraphic Chart 2010 (www.stratigraphy.org), so we used this new fossil to calibrate the crown age for the results of the morphometric analyses and the phylogenetic relationships on the basis of the ITS set (Additional file 3: Table S3).
Results
Systematics
Family: Leguminosae Juss. (or Fabaceae Lindl.)
Subfamily: Caesalpinioideae DC.
Tribe: Cercideae Bronn
Subtribe: Bauhiniinae (Benth.) Walp.
Genus: Bauhinia L.
Species: Bauhinia wenshanensis H.H. Meng et Z.K. Zhou sp. nov.
Twenty-four fossil species of Bauhinia are described as follows. All the voucher specimens were collected from the same locality and stratigraphy, and they are deposited at the same locality.
Holotype: DMS0019, Figure 4A (designated here).
Figure 4.

Bauhinia wenshanensis sp. nov. from the late Miocene of Wenshan, Southeast Yunnan, China. The leaf gross morphology of several species, (Figure 4, A-E). Scale bars = 2 cm.
Isotype: DMS0020, Figure 4B (designated here).
Paratypes: DMS0008, 0021, 0022 (Figure 4C, D, E); DMS0001-0007, DMS0009-0018, DMS0023-0024 (designated here).
Repository: Paleoecology Research Group, Xishuangbanna Tropical Botanical Garden, CAS.
Type locality: Dashidong Town, Wenshan County, Southeast Yunnan, China. (23°20′50″N, 104°17′19″E, alt. 1277 m).
Stratigraphy: Xiaolongtan Formation, Upper Miocene Epoch.
Etymology: The specific epithet wenshanensis is given in reference to the locality Wenshan, where the fossils were collected.
Diagnosis: Adult leaf, petiolate, pulvinate, and simple. Lamina bilobed, apically cordate and symmetrical, apex acuminate, base rounded or ovate. Venation palmate; primary veins 7–9, midvein ending with a free, small projection beyond lamina; secondary venation camptodromous; major secondary veins terminating at the leaf edge.
Description: All leaf fossils are simple, petiolate, and bilobed. The basal portion is slightly ovate, rounded, or cordate. The leaf apex is obtuse, approximately 6–7 cm long and approximately 3–4 cm wide. The two lobes of the lamina share the same pulvinus. The two lobes are attached along the midvein for about 3/4 of the lamina length. Lobes are acutely pointed. The base is rounded to shallowly cordate. The petiole is 2–4 cm long, and thickened at the base. The venation is palmate with 7–9 primary veins in the leaflet. The midvein ends with a free, small apical mucro. Lateral veins are frequently branched. Secondary veins diverge at approximately 45° on the proximal side, fused with other secondary veins or the branches of primary veins to the leaf margin, or arcs between the primary veins. Higher order veins are not visible.
Comparative morphology
PCA and CA among extant Bauhinia and the new fossil B. wenshanensis revealed the leaf shape variability of this genus (Figure 5 and Additional file 4: Figure S1). Species which are visibly or morphologically similar can be distinguished. However, the position of the species within the PCA and CA plots does not reflect the geographic distribution of species in Bauhinia. The results suggest that B. acuminata, B. championii, B. chalcophylla, B. ungulata, B. podopetala, B. madagascariensis, and B. bassacensis are similar to B. wenshanensis (Figure 5). In particular, B. acuminata, B. championii, B. chalcophylla, B. purpurea, and B. podopetala are the closest to B. wenshanensis in leaf shape (Figures 4, 5, and 6). All bear similar strongly bilobed lamina, a bifid leaflet, and a midrib ending in a mucro; especially, the leaf apex is acute. B. acuminata, B. championii, B. purpurea, and B. chalcophylla, are the most similar to the new fossil according to the PCA (Figure 6), and these species are widespread in China today. They are bilobed with a shared pulvinus, and the leaf blade parted to approximately 1/3 to 1/2 of its length.
Figure 5.
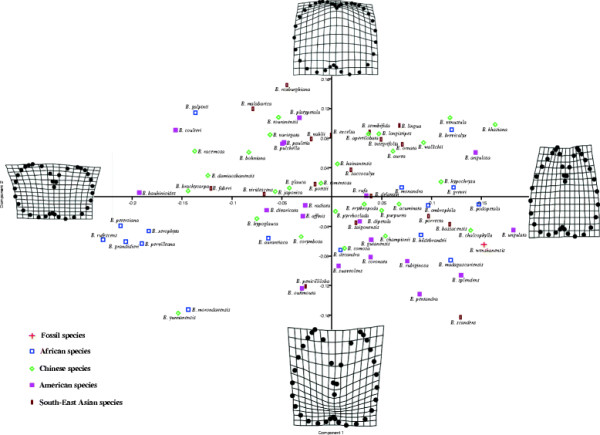
Principal component analysis (PCA) showing the distribution of specimens according to the first and second principal components. Graphs along axes are thin-plate splines of deformation observed along these axes. The biplot shows the contributions of the different descriptors.
Figure 6.

The leaves of extantBauhiniaspecies. (A)B. podopetala. P00149666 (P, PC). (B)B. ungulata. 01685366 (PE). (C) Detail of cleared lamina of B. acuminata. Leaf showing nearly major secondary veins and the intramarginal veins. (D)B. championii. 0169748 (KUN). (E)B. chalcophylla. 0169706 (KUN). (F)B. purpurea. P03100612 (P, PC). (G) Pendent leaves of B. acuminata. Xishuangbanna Tropical Botanical Garden, Mengla, China, 2012. Scale bars = 2 cm.
Phylogenetic relationships and divergence times
The most parsimonious tree and the Bayesian inference tree yielded essentially identical topologies, so only the Bayesian analysis tree is illustrated here (Figure 7). The phylogram shows that Bauhinia species are well-resolved into three main groups. The clades with the high bootstrap values (BP) in the maximum parsimony (MP) analysis also had high posterior probabilities (PP) in the Bayesian analysis. The three main clades recognized in the phylogeny are groups A and B, which include the species in Asia and Africa respectively; and group C, which includes the species in America and B. tomentosa, an Asian species (Figure 7).
Figure 7.
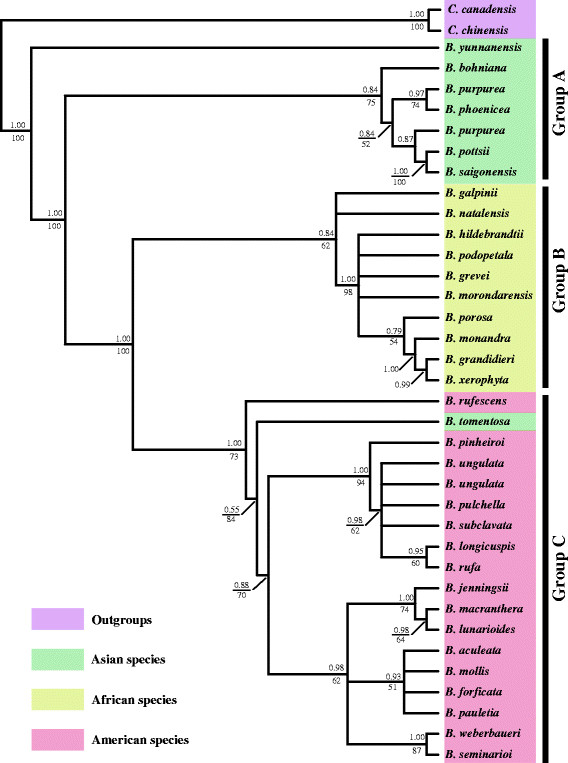
Phylogenetic relationships among extantBauhiniaspecies based on the cpDNA tRNA-Leu (trnL) gene and thetrnL-trnF intergenic spacer data. The numbers above the branches are support values from Bayesian inference/bootstrap resampling.
The Bayesian estimation of divergence times for the three major clades of Bauhinia is presented along with calibration points from fossil records on a chronogram (Figure 8A). The estimated initial divergence time of Bauhinia is ca. 62.7 Ma (Paleocene), and the divergence time of the Asian species B. yunnanensis was ca. 59.8 Ma (Paleocene). The subsequent divergence between the Asian and African-American species groups is estimated at ca. 34.3 Ma (late Eocene), and the diversification times of subclades of the Asian, African, and American species group were ca. 21.8 Ma, ca. 15.3 Ma, and ca. 18.7 Ma, respectively.
Figure 8.
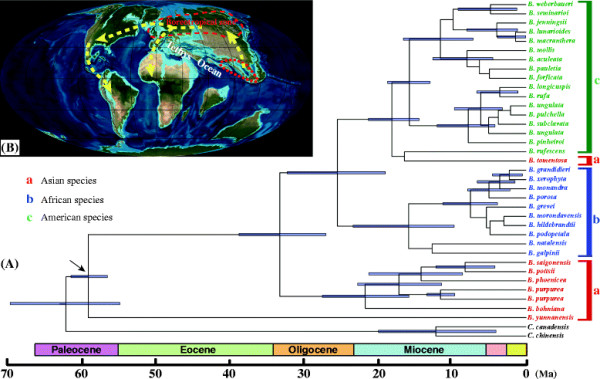
The divergence time and possible dispersal routes. (A) Bayesian divergence time estimates of Bauhinia based on the cpDNA. The blue bars on the nodes indicate 95% posterior credibility intervals. The arrow indicates the stem node of legumes hypothetically at 60 Ma to calibrate the stem age in molecular dating as previous molecular research [34]. (B) The possible dispersal routes of Bauhinia. The paleogeographic map is from Colorado Plateau Geosystems, Inc. Reconstructing the Ancient EARTH.
Discussion
Fossil leaves of Bauhinia
Palmately veined leaves that are divided into two halves, either fused or free, each half with independent nyctinastic movement but borne on a single apical common joint, are unique features of the Leguminosae family [24],[30],[68]. There are 12 genera in the tribe Cercideae with unifoliolate or bifoliolate leaves, and several of the unifoliolate taxa (e.g., Piliostigma, Tylosema, Lysiphyllum, and many Phanera species) with bilobed leaves [69]. However, the secondary veins of these species are entirely different among them. In China, Phanera championii is the homotypic synonym of Bauhinia championii, the present fossil was carefully compared, and these were found to be very similar, with the exception of the secondary veins (Figure 6D). The leaves have moderately developed secondary veins on the midrib and interprimary veins that are easily distinguished from Bauhinia[70], which are incongruent with the secondary veins of the present fossils. Therefore, the present species is thus obviously referred to as Bauhinia.
After comparing the fossil materials with specimens of extant species of the genus, we conclude that the fossil specimens represent a hitherto undescribed taxon of Bauhinia. The morphometric analysis demonstrates the uniqueness of the leaf shape of the fossils. Therefore, a new species is proposed.
Implications of Bauhinia fossil history
We used Bauhinia wenshanensis to calibrate the crown age. B. wenshanensis is morphologically most similar to B. acuminata and B. purpurea according to the PCA and CA that were carried out in the morphometric approach. The phylogenetic analyses also illustrated that B. acuminata and B. purpurea are the closest genetic relatives on the basis of the ITS sequence (Additional file 3: Table S3, Additional file 5: Figure S2), so B. wenshanensis was used to calibrate the crown age of Bauhinia and constrain the age of B. purpurea.
The dating of evolutionary events, such as in phylogeny, biogeography, and evolution studies, has frequently relied on fossils because of their special significance [71]-[75]. The distribution area of the oldest lineage has often been regarded as the original center or diversity center of the taxon [76]. East Asia is one distribution center of some genera of pantropical floral distribution patterns, such as Bauhinia[77]. However, the regions where the oldest known fossils occur might not be the center of origin of the genus [78],[79]. To explore the center of origin, we need to integrate molecular data and fossil taxa in the analyses.
Phylogenetic implications
The previous phylogeny of Bauhinia s.l. used 85 representative species, and indicated that Bauhinia was paraphyletic with the monospecific genus Brenierea clustered within it, but Brenierea is usually described as sister to Bauhinia[25]. Bayesian analysis well resolves and reflects a phylogeny of the extant Bauhinia (Figure 7). Our results revalidated the sister-group relationships and supported three major clades of Bauhinia, groups A, B, and C, representing the Bauhinia species in Asian, African, and the American continents respectively (Figure 7). Group A includs the representative species from Asia, which are all native Asian species from China, Singapore, Thailand and Australia. Group B included the species from Africa (e.g., Mauritius and Madagascar), which were unambiguously supported to be monophyletic. Group C represents species growing in the Neotropics; however, the species B. tomentosa is nested within group C. B. tomentosa is considered to have originated in tropical Asia, and is probably indigenous to India, although widely cultivated as an ornamental plant [55], and B. tomentosa from Hawaii was considered as cultivated species in previous research [25]. Thus, the relationship between B. tomentosa and the other species in group C indicates the close affinity between this Asian species and those of the American continent.
Biogeographic implications of Bauhinia
Based on our calibration point including the hypothetical stem age lacking fossil evidence, plus the crown age based on our fossil, the divergence of Bauhinia from its closest relative lineage in the Leguminosae family occurred at ca. 62.7 Ma (Figure 8A). The biogeographic scenario that Bauhinia originated and initially diversified in Asia is tenable, and consistent with the greatest extant diversity and the highest endemism in Asia (e.g., 23 in 47 species are endemic in China according to Flora of China) [55]. The Asian group is more basal than the American and African groups, and B. yunnanensis is a relatively primitive species, which also suggests that East Asia should be the earliest diversification center.
Here, hypotheses based on vicariance or continental history to explain continental disjunct distributions are dismissed in the biogeography of Bauhinia, because the hypotheses mostly require Palaeogene or older tectonic events [80]. According to our results, Bauhinia originated in Asia and then dispersed in Africa and America. We make the hypothesis that the genus scattered circumboreally across the Northern Hemisphere, although there only fossil record in Europe, Bauhinia olympica (No photograph and just a record; maybe questioned), is from Greece [53], and there are extant plants in North America but without valid Bauhinia fossil records. Then, it entered the North American floristic area through the BLB or the NALB; these land connections and widespread equable climates allowed a relatively homogeneous boreal flora to distribute through a large part of the Northern Hemisphere during the Eocene Epoch [81]-[86].
Oxygen isotope records suggest that very warm climates occurred globally in the late Paleocene to the early Eocene, but cooling proceeded during the mid- to late Eocene with small fluctuations [84]. The drastic cooling events after the PETM made the boreotropical floral elements move to lower latitudes, leading to segregation of the ancestral lineages of modern tropical plants between the North American and Eurasian continents [17],[18],[20],[87]. Bauhinia might have followed the same path and retreated from high to lower latitudes and warmer regions to survive. The absence of extant Bauhinia in Europe does not immediately rule out the NALB as a possible migration pathway. The BLB is situated at higher latitudes than the NALB, which restricted such a tropical or subtropical plant to migrate (e.g., ecological factors, light and temperature). So the NALB might have been the only migration route between Eurasia and America available at any point during the Tertiary for Bauhinia, and should be considered as a migration route for direct migration between southwest Eurasia and North America. The ‘Madrean-Tethyan’ route, wherein a string of volcanic islands at the latitude of the modern Azores allowed migration by ‘island-hopping’ between the two continents between 25 and 38 Ma [22],[88],[89] is also possible. Many tropical taxa have undergone dispersal through the NALB, Malpighiaceae, the “American colonist” scenario had required at least six dispersal events across the Atlantic [8]; Ampelopsis had two independent migrations into Eurasia that are inferred to have occurred in the early Miocene via the NALB [5]. The discovery of more fossil records in Europe would support the NALB route robustly. Here, we suggest that a scenario in which Bauhinia first migrated from Asia to Europe, and then to the America through the NALB, seems the most plausible (Figure 8B).
Additionally, we suggest that the African lineage migrated into Africa via Eurasian continent, particularly from Eurasia to Africa (Figure 8B). The split between the Asian and African-American lineages during the late Eocene to early Oligocene generally coincides with periods of cooling across northern latitudes [90],[91]. This period may also correspond to a time when ancestral acridocarpoids (Malpighiaceae) followed equable climates southward [8]. Additionally, dispersal from Eurasia into Africa seems likely in view of the ancient connections between Africa and Eurasia. Five episodes of major regional change in palaeogeographic and tectonic settings are recognized: late Eocene (37–34 Ma), early late Oligocene (30–27 Ma), latest early to earliest middle Miocene (17–15 Ma), early late Miocene (9–8 Ma), and late early to early middle Pliocene (4–3 Ma) [92]. Most interestingly, this early middle Miocene Africa-Eurasia convergence could explain the Bauhinia migration from Eurasia to Africa and then diversification (diversification time of African subclade is at ca. 15.3 Ma), that inferred from our analyses.
The palaeogene fossil records of Bauhinia are not reliable, and the scenarios that reliable macrofossils of Bauhinia made their debut in the Eocene-Oligocene floras from mid-low latitudes and appeared to lack in the boreotropical floras seem more reasonable; however, we suggest that Bauhinia may have been in the coeval floras at high latitudes (i.e., palaeogene floras in North America and Europe) but not in the right place or time for fossils to be preserved. In addition, the evidence from integrated fossil and molecular evidence supports a tropical Tethys Seaway origin and spread or an “Out of Tropical Asia” dispersal of the Cercideae and the Leguminosae [25],[51]. Under this scenario, we suggest that Bauhinia originated in Asia during the Paleocene and then spread out to other regions, although they were in Asia by the early Oligocene based on direct fossil evidence [44],[51]. Such a scenario is consistent with the hypothesis of the Indomalayan region being a refugium for the boreotropical flora during the global cooling [7],[93]. Lowland tropical forests in this region have experienced relatively climatic and ecological stability since the late Cretaceous in contrast to their American and African counterparts [94], although the uplift of Himalayas and the monsoonal intensification have impacted these regions. Moreover, land connections between Southeast Asia and more northerly regions of the Northern Hemisphere were presented during much of the Tertiary and could have provided migration routes between these areas [95],[96]. The middle Miocene and Holocene fossils in Africa confirm that Bauhinia has been found in Africa since the middle Miocene and is consistent with a migration from Eurasia into Africa. Thus, our data suggest that Bauhinia migrated into Africa from Eurasia and then subsequently into Madagascar.
Nevertheless, the possibility of long distance dispersal (LDD) events from Eurasia to the African and American continents cannot be ruled out. LDD has been viewed as a dominant mechanism for the distribution of many relatively younger tropical plant lineages, such as taxa in Chrysophylloideae [1], Simaroubaceae [16], and Melastomataceae [11]. Some studies have already indicated that LDD events are caused by random incidents [11],[97]-[99], and the dispersal mechanism of the plant itself is sometimes irrelevant for LDD [100]. However, LDD has also been invoked as an explanation for the pantropical disjunction of Bauhinia. We suggest that birds are major agents of Bauhinia plant dispersal, because the birds, with long-distance flight capabilities, are considerably important for leguminous species [101], and the dispersal mechanism of legumes was inferred [102].
Conclusions
We describe a new species of Bauhinia from the late Miocene in Wenshan, Southeast Yunnan, China. Analyses integrating hypothesized age of the stem group, plus fossil and molecular data, suggest that the present pantropical distribution pattern was already established by the middle Paleocene in Laurasia, and possibly originated from Asia. The disjunct distribution of Bauhinia in America, Africa, and Asia is the result of the southward movement of the boreotropical flora in response to the climatic cooling during the late Eocene to the early Oligocene. After originating in Asia, Bauhinia reached the whole of Eurasia and then migrated to the American continent via the NALB, although there are no validly determined Bauhinia fossils in the North American record. And Bauhinia migrated from Eurasia to Africa when the two land masses were connected during the Cenozoic Era. LDD can also explain some of the patterns which were observed. The findings illustrate the power of incorporating fossil records and modern plant distribution areas, together with assumptions about stem group age, to better understand the geographic patterns and the possible dispersal routes of plant species.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
ZKZ conceived and conducted the study. HHM performed the data analyses, evolutionary interpretations, and wrote the manuscript. FMBJ performed data analyses. TS photographed the specimens and revised the manuscript. HHM, FMBJ, TS, YJH, STZ and HJM collected the type specimens. All authors read and approved the final manuscript.
Additional files
Supplementary Material
List of studied specimens. The numbers in the table after a genus name refer to the number of recognized species in the genus.
GenBank accession numbers and their references for sources of cpDNA tRNA-Leu (trnL) gene and trnL-trnF intergenic spacer data of Bauhinia and the outgroups.
GenBank accession numbers and their references for sources of ITS (rRNA internal transcribed spacers) data of Bauhinia and the outgroups.
The cluster analysis (CA) dendrogram of the Bauhinia leaf shape.
The phylogenetic relationships among extant Bauhinia species based on the rDNA ITS. The numbers above the branches are support values from Bayesian inference/bootstrap resampling.
Contributor Information
Hong-Hu Meng, Email: honghumeng@gmail.com.
Frédéric MB Jacques, Email: jacques@xtbg.org.cn.
Tao Su, Email: sutao@xtbg.org.cn.
Yong-Jiang Huang, Email: huangyongjiang@mail.kib.ac.cn.
Shi-Tao Zhang, Email: taogezhang@hotmail.com.
Hong-Jie Ma, Email: 32084917@qq.com.
Zhe-Kun Zhou, Email: zhouzk@xtbg.ac.cn.
Acknowledgements
We would like to thank Dr. Yao-Wu Xing and Dr. He Xu for their assistance in this study. Funding for this research was provided by the National Key Basic Research Project (“973” Project, No. 2012CB821900 to Z.-K. Zhou), and the National Natural Science Foundation of China (No. 41372035 to Z.-K. Zhou, and No. 41272007 to F. MB Jacques).
References
- Bartish I-V, Antonelli A, Richardson J-E, Swenson U. Vicariance or long-distance dispersal: historical biogeography of the pantropical subfamily Chrysophylloideae (Sapotaceae) J Biogeogr. 2011;38:177–190. doi: 10.1111/j.1365-2699.2010.02389.x. [DOI] [Google Scholar]
- Givnish T-J, Renner S-S. Tropical intercontinental disjunctions: Gondwana breakup, immigration from the boreotropics, and transoceanic dispersal. Int J Plant Sci. 2004;165(Supp. 4):1–6. doi: 10.1086/424022. [DOI] [Google Scholar]
- Renner S-S, Clausing G, Meyer K. Historical biogeography of Melastomataceae: the roles of Tertiary migration and long-distance dispersal. Am J Bot. 2001;88:1290–1300. doi: 10.2307/3558340. [DOI] [PubMed] [Google Scholar]
- Milne R-I. Northern Hemisphere plant disjunctions: a window on tertiary land bridges and climate change? Ann Bot. 2006;98:465–472. doi: 10.1093/aob/mcl148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nie Z-L, Sun H, Manchester S-R, Meng Y, Luke Q, Wen J. Evolution of the intercontinental disjunctions in six continents in the Ampelopsis clade of the grape family (Vitaceae) BMC Evol Biol. 2012;12:17. doi: 10.1186/1471-2148-12-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tiffney B-H. Perspectives on the origin of the floristic similarity between eastern Asia and eastern North America. J Arnold Arbor. 1985;66:73–94. [Google Scholar]
- Wolfe J-A. Some aspects of plant geography of the Northern Hemisphere during the Late Cretaceous and Tertiary. Ann Mo Bot Gard. 1975;62:264–279. doi: 10.2307/2395198. [DOI] [Google Scholar]
- Davis C-C, Bell C-D, Mathews S, Donoghue M-J. Laurasian migration explains Gondwanan disjunctions: evidence from Malpighiaceae. Proc Natl Acad Sci U S A. 2002;99:6833–6837. doi: 10.1073/pnas.102175899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wen J. Evolution of eastern Asian and eastern North American disjunct distributions in flowering plants. Annu Rev Ecol Syst. 1999;30:421–455. doi: 10.1146/annurev.ecolsys.30.1.421. [DOI] [Google Scholar]
- Donoghue M-J, Bell C-D, Li J-H. Phylogenetic patterns in Northern Hemisphere plant geography. Int J Plant Sci. 2001;162(Supp. 6):41–52. doi: 10.1086/323278. [DOI] [Google Scholar]
- Renner S-S. Plant dispersal across the tropical Atlantic by wind and sea currents. Int J Plant Sci. 2004;165:23–33. doi: 10.1086/383334. [DOI] [Google Scholar]
- Doyle J-A, Sauquet H, Scharaschkin T, Le Thomas A. Phylogeny, molecular and fossil dating, and biogeographic history of Annonaceae and Myristicaceae (Magnoliales) Int J Plant Sci. 2004;165(Supp. 4):55–67. doi: 10.1086/421068. [DOI] [Google Scholar]
- Erkens RH-J, Maas J-W, Couvreur TL-P. From Africa via Europe to South America: migrational route of a species-rich genus of Neotropical lowland rain forest trees (Guatteria, Annonaceae) J Biogeogr. 2009;36:2338–2352. doi: 10.1111/j.1365-2699.2009.02162.x. [DOI] [Google Scholar]
- Weeks A, Daly D-C, Simpson B-B. The phylogenetic history and biogeography of the frankincense and myrrh family (Burseraceae) based on nuclear and chloroplast sequence data. Mol Phylogenet Evol. 2005;35:85–101. doi: 10.1016/j.ympev.2004.12.021. [DOI] [PubMed] [Google Scholar]
- Razafimandimbison S-G, McDowell T-D, Halford D-A, Bremer B. Origin of the pantropical and nutriceutical Morinda citrifolia L. (Rubiaceae): comments on its distribution range and circumscription. J Biogeogr. 2010;37:520–529. doi: 10.1111/j.1365-2699.2009.02229.x. [DOI] [Google Scholar]
- Clayton J-W, Soltis P-S, Soltis D-E. Recent long-distance dispersal overshadows ancient biogeographical patterns in a pantropical angiosperm family (Simaroubaceae, Sapindales) Syst Biol. 2009;58:395–410. doi: 10.1093/sysbio/syp041. [DOI] [PubMed] [Google Scholar]
- Azuma H, García-Franco J-G, Rico-Gray V, Thien L-B. Molecular phylogeny of the Magnoliaceae: the biogeography of tropical and temperate disjunctions. Am J Bot. 2001;88:2275–2285. doi: 10.2307/3558389. [DOI] [PubMed] [Google Scholar]
- Fritsch P-W. Phylogeny and biogeography of the flowering plant genus Styrax (Styracaceae) based on chloroplast DNA restriction sites and DNA sequences of the internal transcribed spacer region. Mol Phylogenet Evol. 2001;19:387–408. doi: 10.1006/mpev.2001.0933. [DOI] [PubMed] [Google Scholar]
- Van der Hammen T, Cleef A. Trigonobalanus and the tropical amphi-pacific element in the North Andean forest. J Biogeogr. 1983;10:437–440. doi: 10.2307/2844750. [DOI] [Google Scholar]
- Wang Y, Fritsch P-W, Shi S, Almeda F, Cruz B-C, Kelly L-M. Phylogeny and infrageneric classification of Symplocos (Symplocaceae) inferred from DNA sequence data. Am J Bot. 2004;91:1901–1914. doi: 10.3732/ajb.91.11.1901. [DOI] [PubMed] [Google Scholar]
- Queiroz A. The resurrection of oceanic dispersal in historical biogeography. Trends Ecol Evol. 2005;20:68–73. doi: 10.1016/j.tree.2004.11.006. [DOI] [PubMed] [Google Scholar]
- Milne R-I, Abbott R-J. The origin and evolution of Tertiary relict floras. Adv Bot Res. 2002;38:281–314. doi: 10.1016/S0065-2296(02)38033-9. [DOI] [Google Scholar]
- Magallón S, Sanderson M-J. Absolute diversification rates in angiosperm clades. Evolution. 2001;55:1762–1780. doi: 10.1111/j.0014-3820.2001.tb00826.x. [DOI] [PubMed] [Google Scholar]
- Wunderlin R, Larsen K, Larsen S-S. Reorganization of the Cercideae (Fabaceae: Caesalpinioideae) Biol Skrifter. 1987;28:1–40. [Google Scholar]
- Sinou C, Forest F, Lewis G-P, Bruneau A. The genus Bauhinia s.l. (Leguminosae): a phylogeny based on the plastid trnL-trnF region. Botany. 2009;87:947–960. doi: 10.1139/B09-065. [DOI] [Google Scholar]
- Bruneau A, Forest F, Herendeen P-S, Klitgaard B-B, Lewis G-P. Phylogenetic relationships in the Caesalpinioideae (Leguminosae) as inferred from chloroplast trnL intron sequences. Syst Bot. 2001;26:487–514. [Google Scholar]
- Kajita T, Ohashi H, Tateishi Y, Bailey C-D, Doyle J-J. rbcL and legume phylogeny, with particular reference to Phaseoleae, Millettieae, and allies. Syst Bot. 2001;26:515–536. [Google Scholar]
- Käss E, Wink M. Molecular evolution of the leguminosae: Phylogeny of the three subfamilies based on rbcL sequences. Biochem Syst Ecol. 1996;24:365–378. doi: 10.1016/0305-1978(96)00032-4. [DOI] [Google Scholar]
- Pennington R-T, Lavin M, Ireland H, Klitgaard B, Preston J, Hu JM. Phylogenetic relationships of basal papilionoid legumes based upon sequences of the chloroplast trnL intron. Syst Bot. 2001;26:537–556. [Google Scholar]
- Wunderlin R, Larsen K, Larsen S. In: Advances In Legume Systematics, Part 1. Polhill RM, Raven PH, editor. Royal Botanical Gardens, Kew, London; 1981. Cercideae; pp. 107–116. [Google Scholar]
- Zhang D-X. A cladistic analysis of Bauhinia L. (Leguminosae: cercideae) Chin J Bot. 1995;7:55–64. [Google Scholar]
- Hao G, Zhang D-X, Zhang M-Y, Guo L-X, Li S-J. Phylogenetics of Bauhinia subgenus Phanera (Leguminosae: Caesalpinioideae) based on ITS sequences of nuclear ribosomal DNA. Bot Bull Acad Sin. 2003;44:223–228. [Google Scholar]
- Bruneau A, Mercure M, Lewis G-P, Herendeen P-S. Phylogenetic patterns and diversification in the caesalpinioid legumes. Botany. 2008;86:697–718. doi: 10.1139/B08-058. [DOI] [Google Scholar]
- Lavin M, Herendeen P-S, Wojciechowski M-F. Evolutionary rates analysis of Leguminosae implicates a rapid diversification of lineages during the Tertiary. Syst Biol. 2005;54:575–594. doi: 10.1080/10635150590947131. [DOI] [PubMed] [Google Scholar]
- Zhang Y-Z: Lithostratigraphy in Yunnan Province. Wuhan: University of Geosciences Press; 1997.
- Wang W-M. A palynological survey of Neogene strata in Xiaolongtan basin, Yunnan Province of South China. Acta Bot Sin. 1996;38:743–748. [Google Scholar]
- Dong W. Upper Cenozoic stratigraphy and paleoenvironment of Xiaolongtan basin, Kaiyuan, Yunnan Province. Proceedings of the Eighth Annual Meetings of Chinese Society of Vertebrate Paleontology. 2001.
- Zhou Z-K: The Miocene Xiaolongtan fossil flora in Kaiyuan, Yunnan, China. MSc. Thesis. Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences; 1985.
- Zhou Z-K. In: The Evolution of the Late Cretaceous-Cenozoic Floras in China. Tao JR, editor. Science Press, Beijing; 2000. Miocene flora of Yunnan Province; pp. 64–72. [Google Scholar]
- Bell W-A: Flora of the Upper Cretaceous Nanaimo Group of Vancouver Island. British Columbia: E. Cloutier; 1957.
- Bell W-A: Upper Cretaceous Floras of the Dunvegan, Bad Heart, and Milk River Formations of Western Canada. vol. 94th edition. Canada: Dept. of Mines and Technical Surveys; 1963.
- Berry E-W. Fossil plants from Bolivia and their bearing upon the age of uplift of the eastern Andes. Proc US Natl Mus. 1917;54:103–164. doi: 10.5479/si.00963801.54-2229.103. [DOI] [Google Scholar]
- Berry E-W. The fossil flora of Potosi, Bolivia. Johns Hopkins Univ Stud Geol. 1939;13:6–68. [Google Scholar]
- Chen Y-F, Zhang D-X. Bauhinia larsenii, a fossil legume from Guangxi, China. Bot J Linn Soc. 2005;147:437–440. doi: 10.1111/j.1095-8339.2005.00373.x. [DOI] [Google Scholar]
- Eisenmann V. Equidae of the Albertine rift valley, Uganda. Geol Paleobiol Albertine Rift valley, Uganda-Zaire. 1994;2:289–307. [Google Scholar]
- Knowlton F-H. Fossil flora of the Yellowstone National Park. US Geol Surv Mon. 1899;32:651–791. [Google Scholar]
- Kowalski E-A: Middle to Late Miocene environments of Southern Ecuador: temperature, elevation, and fossil plants of the Nabón Basin. PhD thesis. University of Michigan; 2001.
- Van Neer W. The use of fish remains in African archaeozoology. CRA-CNRS Notes et Monographies Techniques. 1983;16:155–167. [Google Scholar]
- Wilf P. Late Paleocene-early Eocene climate changes in southwestern Wyoming: Paleobotanical analysis. Geol Soc Am Bull. 2000;112:292–307. doi: 10.1130/0016-7606(2000)112<292:LPECCI>2.0.CO;2. [DOI] [Google Scholar]
- Tao J-R, Zhou Z-K, Liu Y-S. ᅟ. Science press, Beijing; 2000. The Evolution of the Late Cretaceous-Cenozoic floras in China. [Google Scholar]
- Wang Q, Song Z-Q, Chen Y-F, Shen S, Li Z-Y. Leaves and fruits of Bauhinia (Leguminosae, Caesalpinioideae, Cercideae) from the Oligocene Ningming Formation of Guangxi, South China and their biogeographic implications. BMC Evol Biol. 2014;14:88. doi: 10.1186/1471-2148-14-88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endo S, Fujiyama I. In: Contributions to the Geology And Palaeontology of Southeast Asia 31, Geology And Palaeontology of Southeast Asia. volume 2. Kobayashi T, Toriyama R, editor. University of Tokyo Press, Tokyo; 1966. Some Late Mesozoic and Late Tertiary plants and a fossil insect from Thailand; pp. 191–197. [Google Scholar]
- Awasthi N. In: Advances In Legume Systematics. Part 4. Herendeen PS, Dilcher DL, editor. The fossil records London, Royal Botanical Gardens, Kew; 1992. Indian Fossil Legumes; pp. 225–250. [Google Scholar]
- Unger F. Die fossile Flora von Kumi auf der Insel Euboea. Denkschr Akad Wiss, Wien. 1867;27:1–66. [Google Scholar]
- Chen D-Z, Zhang D-X, Larsen K, Larsen S-S. In: Flora of China. Wu Z-Y, Raven P-H, editor. Missouri Botanical Garden Press, Science Press/St. Louis; 2010. Bauhinia; pp. 6–21. [Google Scholar]
- Hou D, Larsen K, Larsen S-S. Caesalpiniaceae. Flora Malesiana. 1996;12:409–730. [Google Scholar]
- Jacques MB-F, Zhou Z-K. Geometric morphometrics: a powerful tool for the study of shape evolution in Menispermaceae endocarps. Taxon. 2010;59:881–895. [Google Scholar]
- Rohlf F: tpsDig: Digitize landmarks and outlines, version 2.10. State Univ. of New York at Stony Brook: Dept. of Ecology and Evolution; 2006.
- Bookstein F-L: Morphometric tools for landmark data: Geometry and biology. Cambridge: Cambridge University Press; 1997.
- Rohlf F: tpsRelw: Relative warps analysis, version 1.45. State Univ. of New York at Stony Brook: Dept. of Ecology and Evolution; 2007.
- Hammer Ø, Harper D, Ryan P. PAST: Paleontogical statistical software for education and data analysis. Palaeontol Elec. 2001;4:4. [Google Scholar]
- Thompson J-D, Gibson T-J, Plewniak F, Jeanmougin F, Higgins D-G. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucl Acids Res. 1997;25:4876–4882. doi: 10.1093/nar/25.24.4876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swofford D: PAUP 4.0 b10: Phylogenetic analysis using parsimony. Sunderland, MA, USA: Sinauer Associates; 2002.
- Ronquist F, Huelsenbeck J-P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–1574. doi: 10.1093/bioinformatics/btg180. [DOI] [PubMed] [Google Scholar]
- Posada D, Crandall K-A. Modeltest: testing the model of DNA substitution. Bioinformatics. 1998;14:817–818. doi: 10.1093/bioinformatics/14.9.817. [DOI] [PubMed] [Google Scholar]
- Drummond A, Rambaut A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol Biol. 2007;7:214. doi: 10.1186/1471-2148-7-214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rambaut A: FigTree, a graphical viewer of phylogenetic trees. 2007. http://treebioedacuk/software/figtree/.
- Van-Der P. The leaf of Bauhinia. Acta Bot Neerl. 1951;1:287–309. doi: 10.1111/j.1438-8677.1952.tb00016.x. [DOI] [Google Scholar]
- Lewis G: Legumes of the world. Kew: Royal Botanic Gardens; 2005:57–67.
- Zhang D-X. Leaf venation of Cercideae (Leguminosae) J Trop Subtrop Bot. 1994;2:45–57. [Google Scholar]
- Matthew W-D. Climate and evolution. Ann New York Acad Sci. 1915;24:171–318. doi: 10.1111/j.1749-6632.1914.tb55346.x. [DOI] [Google Scholar]
- Ernst C, Barbour R: Turtles of the world. Washington, DC: Smithsonian Institute Press; 1989.
- Seiffert E-R, Simons E-L, Attia Y. Fossil evidence for an ancient divergence of lorises and galagos. Nature. 2003;422:421–424. doi: 10.1038/nature01489. [DOI] [PubMed] [Google Scholar]
- Luo Z-X, Ji Q, Wible J-R, Yuan C-X. An Early Cretaceous tribosphenic mammal and metatherian evolution. Science. 2003;302:1934–1940. doi: 10.1126/science.1090718. [DOI] [PubMed] [Google Scholar]
- Mayr G. Old world fossil record of modern-type hummingbirds. Science. 2004;304:861–864. doi: 10.1126/science.1096856. [DOI] [PubMed] [Google Scholar]
- Wu Z-Y, Zhou Z-K, Sun H, Li D-Z, Peng H. ᅟ. Yunnan Science and Technology Press, Kunming; 2006. The areal-types of seed plants and the origin and differentiation. [Google Scholar]
- Wu Z-Y, Sun H, Zhou Z-K, Li D-Z, Peng H: Floristics of seed plants from China. Beijing: Science press; 2010.
- Gould S-J: Wonderful Life: The Burgess Shale and the nature of history. New York: Norton; 2000.
- Gould S-J: The Burgess Shale and the nature of history. London: Penguin; 1989.
- Schrire B-D, Lavin M, Barker N-P, Forest F. Phylogeny of the tribe Indigofereae (Leguminosae-Papilionoideae): Geographically structured more in succulent-rich and temperate settings than in grass-rich environment. Am J Bot. 2009;96:816–852. doi: 10.3732/ajb.0800185. [DOI] [PubMed] [Google Scholar]
- Chandler ME-J. The Lower Tertiary Floras of Southern England. vol. 4. British Museum (Natural History), London, UK; 1964. A summary and survey of findings in the light of recent botanical observations. [Google Scholar]
- Collinson M, Fowler K, Boulter M. Floristic changes indicate a cooling climate in the Eocene of southern England. Nature. 1981;291:315–317. doi: 10.1038/291315a0. [DOI] [Google Scholar]
- Graham A. ᅟ. Oxford University Press, New York; 1999. Late Cretaceous and Cenozoic history of North American vegetation, north of Mexico. [Google Scholar]
- Miller K-G, Fairbanks R-G, Mountain G-S. Tertiary oxygen isotope synthesis, sea level history, and continental margin erosion. Paleoceanography. 1987;2:1–19. doi: 10.1029/PA002i001p00001. [DOI] [Google Scholar]
- Wolfe J-A. In: Evolution And Diversification Of Land Plants. Iwatsuki K, Raven PH, editor. Springer, Tokyo; 1997. Relations of environmental change to angiosperm evolution during the late Cretaceous and Tertiary; pp. 269–290. [DOI] [Google Scholar]
- Wolfe J-A. A paleobotanical interpretation of Tertiary climates in the Northern Hemisphere: Data from fossil plants make it possible to reconstruct Tertiary climatic changes, which may be correlated with changes in the inclination of the earth's rotational axis. Am Sci. 1978;66:694–703. [Google Scholar]
- Wolfe J-A. Neogene floristic and vegetational history of the Pacific Northwest. Madrono. 1969;20:83–110. [Google Scholar]
- Axelrod D-I. Evolution and biogeography of Madrean-Tethyan sclerophyll vegetation. Ann Mo Boto Gardo. 1975;62:280–334. doi: 10.2307/2395199. [DOI] [Google Scholar]
- Liston A, Rieseberg L-H, Elias T-S. Morphological stasis and molecular divergence in the intercontinental disjunct genus Datisca (Datiscaceae) Aliso. 1989;12:525–542. [Google Scholar]
- Berggren WA, Prothero DR. In: Eocene-Oligocene climatic and biotic evolution. Prothero DR, Berggren WA, editor. Princeton University Press, Princeton, NJ; 1992. Eocene-Oligocene climatic and biotic evolution: An overview; pp. 1–28. [Google Scholar]
- Tiffney B-H, Manchester S-R. The use of geological and paleontological evidence in evaluating plant phylogeographic hypotheses in the Northern Hemisphere Tertiary. Int J Plant Sci. 2001;162(Supp. 6):3–17. doi: 10.1086/323880. [DOI] [Google Scholar]
- Meulenkamp J-E, Sissingh W. Tertiary palaeogeography and tectonostratigraphic evolution of the Northern and Southern Peri-Tethys platforms and the intermediate domains of the African-Eurasian convergent plate boundary zone. Palaeogeogr Palaeoclimatol Palaeoecol. 2003;196:209–228. doi: 10.1016/S0031-0182(03)00319-5. [DOI] [Google Scholar]
- Tiffney B-H. The Eocene North Atlantic land bridge: Its importance in Tertiary and modern phytogeography of the Northern Hemisphere. J Arnold Arbor. 1985;66:243–273. [Google Scholar]
- Ricklefs R-E, Latham R-E, Schluter D. In: Species diversity in ecological communities: historical and geographical perspectives. Ricklefs RE, Schluter D, editor. University of Chicago Press, Chicago; 1993. Global patterns of diversity in mangrove floras; pp. 215–229. [Google Scholar]
- Hall R. Reconstructing cenozoic SE Asia. Geol Soc. 1996;106:153–184. doi: 10.1144/GSL.SP.1996.106.01.11. [DOI] [Google Scholar]
- Hall R. Cenozoic geological and plate tectonic evolution of SE Asia and the SW Pacific: computer-based reconstructions, model and animations. J Asian Earth Sci. 2002;20:353–431. doi: 10.1016/S1367-9120(01)00069-4. [DOI] [Google Scholar]
- Nathan R. Long-distance dispersal of plants. Science. 2006;313:786–788. doi: 10.1126/science.1124975. [DOI] [PubMed] [Google Scholar]
- Nathan R, Katul G-G, Horn H-S, Thomas S-M, Oren R, Avissar R, Pacala S-W, Levin S-A. Mechanisms of long-distance dispersal of seeds by wind. Nature. 2002;418:409–413. doi: 10.1038/nature00844. [DOI] [PubMed] [Google Scholar]
- Wilkinson D-M. Plant colonization: are wind dispersed seeds really dispersed by birds at larger spatial and temporal scales? J Biogeogr. 1997;24:61–65. doi: 10.1111/j.1365-2699.1997.tb00050.x. [DOI] [Google Scholar]
- Higgins S-I, Lavorel S, Revilla E. Estimating plant migration rates under habitat loss and fragmentation. Oikos. 2003;101:354–366. doi: 10.1034/j.1600-0706.2003.12141.x. [DOI] [Google Scholar]
- Snow D-W. Tropical Frugivorous Birds and their foot plants: a world survey. Assoc Tropical Biol Conserv. 1981;13:1–14. [Google Scholar]
- Lavin M, Schrire B-P, Lewis G, Pennington R-T, Delgado-Salinas A, Thulin M, Hughes C-E, Matos A-B, Wojciechowski M-F. Metacommunity process rather than continental tectonic history better explains geographically structured phylogenies in legumes. Phil Trans R Soc Lond B. 2004;359:1509–1522. doi: 10.1098/rstb.2004.1536. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
List of studied specimens. The numbers in the table after a genus name refer to the number of recognized species in the genus.
GenBank accession numbers and their references for sources of cpDNA tRNA-Leu (trnL) gene and trnL-trnF intergenic spacer data of Bauhinia and the outgroups.
GenBank accession numbers and their references for sources of ITS (rRNA internal transcribed spacers) data of Bauhinia and the outgroups.
The cluster analysis (CA) dendrogram of the Bauhinia leaf shape.
The phylogenetic relationships among extant Bauhinia species based on the rDNA ITS. The numbers above the branches are support values from Bayesian inference/bootstrap resampling.