

Abstract
Enteric glia are important components of the enteric nervous system (ENS) and also form an extensive network in the mucosa of the gastrointestinal (GI) tract. Initially regarded as passive support cells, it is now clear that they are actively involved as cellular integrators in the control of motility and epithelial barrier function. Enteric glia form a cellular and molecular bridge between enteric nerves, enteroendocrine cells, immune cells, and epithelial cells, depending on their location. This Review highlights the role of enteric glia in GI motility disorders and in barrier and defense functions of the gut, notably in states of inflammation. It also discusses the involvement of enteric glia in neurological diseases that involve the GI tract.
Introduction
Over the last 25 years there has been an explosion in our understanding of the roles of the diverse cellular elements of the gut. Remarkably, even in the last five years, new cell types have been discovered and completely new functions have been ascribed to cells about which we have long known (for example, see refs. 1, 2). These exciting discoveries have refined our understanding of gut physiology and, to some extent, pathophysiology. Nowhere is this exemplified more vividly than in the enteric nervous system (ENS). The ENS provides local neural control of the gastrointestinal (GI) tract and is required for the coordination of digestive and defensive functions of the gut (3). It consists of nerve cell bodies arranged in two ganglionated plexuses and nerve fibers throughout the external muscle layers, submucosa, and mucosa (3). The ganglia of the ENS form the myenteric and the submucosal plexuses, which are located between the longitudinal and circular muscle layers and in the submucosa, respectively. Neurons of the ENS are supported by a unique peripheral glial cell called enteric glia (4–6). Exciting recent findings have shed new light on the roles of enteric glia in the physiology and pathophysiology of the GI tract.
Enteric glia are increasingly being recognized as playing pivotal roles in health and disease. These cells were once thought to play a passive structural role but now emerge as having significant regulatory functions throughout the GI tract (4, 7, 8). There are two major populations of enteric glia, one in the ENS and the other beneath the epithelium throughout the intestinal mucosa (Figure 1). In the ENS, enteric glia slightly outnumber enteric neurons and are found in both the myenteric and submucosal plexuses (ref. 4 and Figure 2). In this Review I focus on selected new developments in the molecular physiology of enteric glia and relate them to pathophysiological conditions in the gut and to neurological conditions that affect the GI tract. Interested readers are encouraged to consult other recent reviews and articles on enteric glia (4, 7–9).
Figure 1. Schematic representation of the distribution of enteric glia in the GI tract.

Subpopulations of enteric glia are located around all classes of neurons in the myenteric and submucosal plexuses and in the mucosa. In the mucosa, enteric glia lie below the epithelium and connect to the “neuropod” of enteroendocrine cells (EECs) to form a tripartite connection among enteric nerves, enteroendocrine cells, and enteric glia (see text for details). Enteric glia also form a functional bridge between immune cells. Dark blue indicates intrinsic primary afferent neurons; light blue, interneurons; orange, excitatory motor neurons; green, inhibitory motor neurons; purple, secretomotor neurons.
Figure 2. Enteric glia in the myenteric and submucosal plexus.
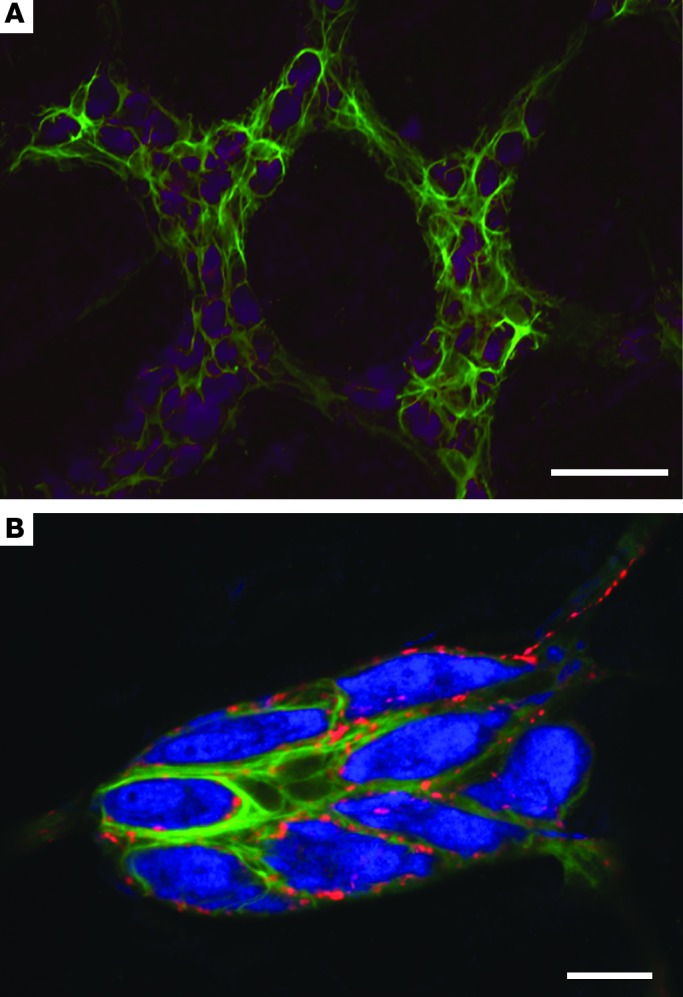
(A) Enteric glia labeled with GFAP (green) surround enteric neurons labeled with Hu C/D (purple) in the myenteric plexus of the mouse colon. Note that the expression of GFAP is not uniform in the myenteric plexus. Scale bar: 50 μm. (B) Enteric glia labeled with GFAP (green) surround enteric neurons labeled with Hu C/D (blue) in the submucosal plexus of the rat colon. Enteric glia and neurons are decorated with punctate nerve terminals expressing calcitonin gene–related peptide (red). Scale bar: 20 μm.
Historical perspective
Glial cells of the ENS were discovered in the late 19th century by Dogiel, but it was not until the 1970s when a detailed ultrastructural analysis of the ENS was conducted by Giorgio Gabella that their identity as a unique cell type was really appreciated (10, 11). Gabella termed these astrocyte-like glial cells “enteric glia” in 1981 (10), though the term had been used previously by Jessen and Mirsky who showed that enteric glia express the glial marker glial fibrillary acidic protein (GFAP) (12). With characteristic precision, Gabella also noted that enteric glia receive specialized contacts from enteric nerves, which he termed “neuroglial junctions” (11). The intimate connections between nerves and glia suggested a functional relationship whose significance remained obscure for many years but which provides the structural basis for many key roles that enteric glia play in the gut. Another important early finding was made by Hanani and colleagues, who showed that enteric glia are dye coupled, forming a functional syncytium (13). Thus, injection of a small-molecular-weight dye into one cell rapidly spreads around the glial network in an enteric ganglion to label 10 to 100 cells over a distance of 300 μm from the site of injection. Below I discuss the significance of the functional connectivity of enteric glia.
Two other early observations are worth highlighting. The first is that in the inflamed ileum of patients with Crohn’s disease enteric glia express MHC class II (14, 15). In controls, MHC class II was weakly expressed or absent, but this expression was markedly upregulated in patients with Crohn’s disease, and Geboes et al. showed that enteric glia were associated with T lymphocytes (14). These data illustrate that enteric glia have immunological functions and form a bridge between the immune system and the neurons of the ENS. This likely has important functional consequences for protecting critical neuronal functions in the face of immunological challenges that frequently plague the digestive system as a result of infection. The second early observation of note is that ablation of enteric glia leads to a rapidly developing fulminant jejunoileitis (16, 17). This massive intestinal inflammation apparently results from breakdown of the intestinal epithelial barrier. This remarkable observation was completely unexpected and illustrated the very significant role of enteric glia in the intestinal mucosa.
This short perspective illustrates the potential role that enteric glia have in physiological and pathophysiological conditions of the GI tract. In the three sections below, I discuss the role of enteric glia in GI motility disorders, in barrier and defense functions of the gut, and in neurological disorders that affect the GI tract.
Role of enteric glia in GI motility disorders
Enteric glia and the control of motility.
Enteric glia were first implicated in the control of GI motility when mice, treated with 6-aminonicotinamide to induce an enteric gliopathy, were found to have diarrhea (18, 19). However, because motility was not directly assessed, it was only recently that this concept gained more traction. Using selective glial disruption techniques, Aubé et al. clearly demonstrated that loss of enteric glia was associated with reduced motility (20). Using a pharmacological approach, Nasser and colleagues also demonstrated reduced motility in animals following glial cell disruption with a selective gliotoxin, fluorocitrate (21). Around the same time as these papers were published, Bassotti and colleagues examined samples of the ileum and colon from patients with slow-transit constipation (22). They showed that there was a widespread and significant loss of enteric glia in most of these specimens, although there was also some reduction in the density of enteric neurons. Taken together, these findings reveal that enteric glia play a role in the physiological control of GI motility. Moreover, they point to enteric glia as cellular elements potentially involved in abnormalities of gut motility (23, 24).
The studies described above raise a number of critical questions, some of which have been addressed in the last few years. In the following sections I examine the mechanisms involved in enteric glial regulation of GI motility.
Enteric glia and neurotransmission in the ENS.
The control of propulsive motility requires coordinated activation of neurons in the ENS. How do glia synchronize their activity with that of enteric neurons? As noted above, Gabella first observed neuroglial junctions in the early 1970s (11). In my laboratory, we developed functional imaging approaches to directly visualize enteric glial activation in response to neuronal stimulation in both mouse and guinea pig myenteric plexus, and demonstrated neuroglial transmission in the ENS (25–27). Building on these findings, Gulbransen’s group showed that the spread of activity in enteric glia, which is observed as an elevation in intracellular calcium, is mediated by connexin-43 hemichannels (28). They utilized genetic approaches in mice to selectively knockout connexin-43 in GFAP-expressing enteric glia. They showed that purinergic signaling to the glial network is substantially blocked in these mice and they confirmed these findings using pharmacological approaches to block connexin-43 hemichannels. Impressively, they then demonstrated that colonic transit was substantially delayed and isometric force generation was reduced in their glial-specific connexin-43 knockout mice. Furthermore, these changes mirrored the effects of aging, in which connexin-43 expression is altered, potentially offering a novel explanation for age-related reductions in colonic motility (28). These new and exciting findings extend work by Broadhead et al., who showed that colonic migrating motor complexes activate enteric glial networks (29). Broadhead and colleagues demonstrated that intracellular calcium waves propagated between closely apposed glia and between glial processes to the glial cell body, where calcium levels were further elevated, suggesting that the glial cell soma acts as an integrator of activity.
These studies raise the question of which neurotransmitters enteric glia respond to. Functional imaging studies, mostly on cultured enteric glial cells from animals, have revealed that enteric glia express receptors for many neurotransmitters (see refs. 4, 30, 31). Boesmans and colleagues recently extended these studies, utilizing enteric glial cultures from human duodenal biopsies, and showed that enteric glia could be activated by all primary fast enteric neurotransmitters, acetylcholine, serotonin, and ATP, although to different degrees (32). This obviously raises the question of whether enteric glia are “innervated” (Figure 2). The early structural work is certainly supportive of this, and further evidence was provided by Vanden Berghe and Klingauf, who demonstrated synaptic release sites on enteric glia (33). However, those studies did not show the transmitter phenotype, and indeed there is still much to do in this regard, but a recent paper from Okamoto et al. elegantly reveals the serotonergic innervation of enteric glia in the myenteric plexus of the colon (34). Importantly, these serotonin neurons have extensive connectivity throughout the myenteric and submucosal plexuses and also innervate interstitial cells of Cajal and blood vessels, suggesting that they are “command neurons” of the colon capable of coordinating motility with secretion and blood flow (34). Previous studies have also shown that enteric glia in the myenteric plexus also receive a cholinergic innervation (35). That enteric glia are innervated along with neuronal and other non-neuronal structures speaks to their importance as part of the integrated regulatory control mechanisms of gut function.
Enteric glia and abnormal GI motility.
The mechanisms that explain how enteric glia are involved in abnormal GI motility have not been extensively studied and remain obscure. Possible mechanisms include immune mediators, inflammatory signaling, and viral infection of enteric glia. As noted above, enteric glia can function as a bridge between the nervous and immune systems, and they may contribute to abnormal motility through immune signaling mechanisms. Such a role has recently been described in an elegant study by Stoffels et al., who examined the mechanisms of postoperative ileus in mice (36). They showed that postoperative ileus was attenuated in mice that lacked the IL-1R or in mice after blocking the IL-1R pharmacologically. They demonstrated that the IL-1R was present on enteric glia and that stimulation of this receptor in cultured enteric glia increased the expression of IL-6 and monocyte chemotactic protein 1 (MCP1), which stimulates the infiltration of immune cells and the local inflammatory conditions associated with postoperative ileus (36). The presence of cytokine receptors on enteric glia is a very interesting feature of this study and should be confirmed with additional experimental approaches. Enteric glia may also contribute to ileus through the production of NO from iNOS (37, 38), though this has yet to be directly demonstrated.
Enteric glia could also be the target of viral infection, as revealed in an examination the myenteric plexus from patients with chronic idiopathic intestinal pseudo-obstruction (39). Here it was shown that about 70% of samples from patients with severe dysmotility had John Cunningham virus–related (JCV-related) T antigen protein expression and 80% had DNA sequences of the JCV T antigen, in contrast to control patients (patients with uncomplicated colon cancer), in whom no protein expression was detected and DNA sequences of the JCV T antigen were found in only 10% of samples. Interestingly, the JCV T antigen was colocalized in GFAP immunoreactive enteric glia and not enteric neurons (39). The functional consequences of this infection and whether or not the infected glia will go on to die remain to be determined. These findings are consistent with previous work showing that the enteric glia are preferentially infected with adenovirus in vitro (40) and suggest that viral infection of glia may lead to dysfunction and contribute to dysmotility.
As noted above, it has been reported that there is a preferential loss of enteric glia in patients with slow-transit constipation, though the underlying mechanisms are completely unknown. This is due in part to a limited understanding of the factors that regulate the maintenance of enteric glia cell numbers in the GI tract. However, one factor that has recently been described is the retinoblastoma 1 tumor suppressor gene (Rb1) (41). Rb1 knockout mice gain weight slowly and suffer from intestinal pseudo-obstruction and severe colonic motility defects. Surprisingly, the most striking alteration in the ENS of these mice is a doubling of the enteric glial cell populations in the myenteric plexus of the ileum and colon, coupled with an enlargement of the size of some NOS-expressing neurons in the distal ileum (41). Other populations of enteric neurons are similar to those in control mice. At this point it is unclear how these changes in enteric glial cell numbers alter motility. However, one can imagine that a lack of coordinated network responses in the glial cells might significantly compromise normal contractility of the bowel.
The factors that lead to loss of enteric glia are also not well understood. However, studies examining the effects of dietary manipulation and aging reveal that enteric glial cell numbers appear to be influenced by both age and diet, suggesting that environmental factors are able to influence glial cell numbers (28, 42–46). Studies of aging reveal a consistent reduction in enteric glial cell density in the myenteric plexus (28, 44). In contrast, the development of obesity caused by feeding a high-fat diet had only minimal effects on glial cell density in the myenteric plexus but did lead to a massive reduction in apparent glial cell density in the duodenal mucosa and the submucosal plexus (45). Interestingly, food restriction in aging rats actually accentuated the loss of enteric glia, although it had been expected that food restriction may reduce the parameters of oxidative stress in the GI tract (46). Finally, another study found that enteric glia of the rat intestine cultured in vitro and exposed to elevated levels of lipids expressed an altered morphology without any change in the cell density (47). An important point to consider is whether changes in glial numbers with age or diet or in the Rb1 knockout mice are reflective of direct effects on glia or are secondary responses in the glia to alterations in the size or number of enteric neurons. Since very little is known about the actual processes that regulate glial number in the ENS, this question cannot be answered at this time. Another point to consider from the discussion above is that an insult to enteric glia may affect the integrity of enteric neurons, implying that gliopathy-mediated neuronal damage might be a cause of abnormal GI motility. Further investigations that directly examine this possibility are warranted.
An important caveat to consider in all of these studies is that the populations of enteric glia are not themselves homogeneous but rather show a remarkable degree of structural, functional, and phenotypic heterogeneity (9). This complicates the issues discussed above, as the roles of enteric glia may well be linked to specific glial subtypes. Importantly, how these different subtypes of enteric glia respond to insults in pathophysiological situations remains to be determined. This may be of considerable relevance given the differential functional responsiveness of these glia.
Role of enteric glia in the mucosa
The significance of the glial network of the mucosa is underscored by the effects of targeted glial ablation, as described previously (16). However, the nature of this glial network in the human gut was not well understood. Recent findings using novel imaging methods reveal the very extensive nature of the distribution of mucosal enteric glia (48). Using optical clearing methods combined with high-resolution confocal microscopy in the human colon, Liu et al. showed that enteric glial processes form a continuous network in the lamina propria from the base of crypts extending up the mucosa to the crypt openings (48). Enteric glia are also in intimate contact with subepithelial myofibroblasts and the epithelial basement membrane. Moreover, the glial network is also in close contact with lymphatic vessels, an association not previously known and whose function has yet to be demonstrated. Recently, Badizadegan et al., identified a novel population of cells with a dendritic morphology in the colonic mucosa that have both neuronal and glia phenotypes (49). These intramucosal neuroglial cells express S-100 and the neuronal marker Tuj1. They were observed in normal human colon and, interestingly, in the mucosa of patients with Hirschsprung disease, challenging the current concepts of the developmental origins of the ENS (49). The function of these newly described cells remains to be determined.
The mucosal enteric glial cell population is positioned to serve as a signaling intermediary between the epithelial cells that line the GI tract and immune and neural elements in the lamina propria of the mucosa (Figure 1). Exactly how this is accomplished and the nature of the signals are now becoming clearer. At an anatomic level, it was recently discovered that enteric glia are not only associated with enteric nerves, but also with a structure called a “neuropod,” a basal extension of enteroendocrine cells (50). Enteroendocrine cells are specialized cells that are scattered throughout the epithelium of the gut wall. These cells release amines and peptides basolaterally, which act as local paracrine regulators or as gut hormones. Recent evidence supports the notion that these cells, which have microvilli that project into the lumen, are capable of “tasting” the luminal environment (51, 52). That enteric glia may be targets of the enteroendocrine transmitters and that enteroendocrine cells could be influenced by glial-derived neurotrophic factor (GDNF) or other signaling molecules opens up new avenues for signaling in the gut. Bohórquez et al. revealed that neuropods were formed more frequently in the presence of nerve growth factor, suggesting this anatomic relationship is plastic in nature and subject to regulation (50). The significance of these findings is not yet clear but imply that factors released from enteroendocrine cells during digestion or by other means regulate glial function, which may in turn regulate barrier function. An interesting potential example in this regard is the peptide glucagon-like peptide 2 (GLP-2), which is released from L cells, a subpopulation of enteroendocrine cells found mostly in the distal ileum and colon (53). GLP-2 has long been known to enhance gut barrier function, and in a recent paper we showed that the enteric glia express GLP-2 receptors and are activated by this peptide (54). While the enhancement of barrier function has not been directly linked to enteric glial function, these findings are consistent with this possibility.
Functionally, many enteric glial-derived factors, including S-nitrosoglutathione, GDNF, and TGF-β, have been shown to play important roles in enhancing barrier function by reducing epithelial permeability (8, 55–57). Enteric glia could be activated to fulfill this homeostatic role by bacterial or other luminal signals or by neural signals. Vagal nerve activation of enteric glia has been linked to enhanced barrier function, and recently nicotinic cholinergic receptor signaling pathways have been shown to downregulate NF-κB in enteric glia, providing a potential intracellular mechanism for this effect (58–60). Enteric glia express TLRs, and these are differentially upregulated in response to pathogenic and non-pathogenic bacteria (61, 62). When the upregulation of TLRs by a bacterial pathogen occurs, it leads to NF-κB activation. An increase in S-100B protein has also been observed and, together with NF-κB activation, leads to enhanced production of NO. The presence of S-100B in enteric glia has previously been shown to increase inducible NO synthase (iNOS), and S-100B–mediated upregulation of iNOS has been demonstrated in ulcerative colitis and also, interestingly, in celiac disease, where there are marked alterations in mucosal integrity (63–65). It seems likely that the role of NO is bactericidal; however, rather than being homeostatic, an excessive production of NO through the upregulation of iNOS in enteric glia could lead to a breakdown in barrier function (66). Of further interest, when enteric glia are activated by stimuli that increase iNOS (lipopolysaccharide and IFN-γ), MHC class II expression is induced, as it is in the ENS from patients with Chagas disease (67). Taken together, these studies demonstrate that enteric glia play a pivotal role in the regulation of gut homeostasis by responding to pathogenic challenges through various defensive mechanisms. They also extend the earlier observations of enhanced expression in Crohn’s disease and raise the possibility that enteric glia upregulate MHC class II in response to specific pathogens as part of the host defense mechanisms to protect the ENS.
The links between inflammation and enteric glial activation were recently tested in a mouse model of intestinal inflammation and using human colonic biopsies (62). In dextran sodium sulphate–induced colitis and human ulcerative colitis, the lipid mediator palmitoylethanolamide (PEA) reduced inflammation through activation of PPARα (62). It also reduced enteric glial S-100B expression, the expression of TLR4 and the production of NO in the animal model and in isolated human and mouse enteric glial cells. In Ppara knockout mice, PEA has no anti-inflammatory effects and does not reduce S-100β or TLR4 expression (or that of other inflammatory markers) (62, 68). While in this study PEA was given exogenously, it is produced naturally in the gut and its levels are regulated by the enzyme fatty acid amide hydrolase, whose expression has been proposed to be important in the control of intestinal inflammation (69). Whether endogenous PEA acting on enteric glia is an important mediator of host defense remains to be determined.
Role of enteric glia in brain disorders
Though it may seem counterintuitive that glial cells residing in the wall of the gut are involved in disorders of the CNS, two recent observations suggest otherwise. The first reveals enteric glia as targets for the misfolded (infective) isoform of the prion protein, which is responsible for transmissible spongiform encephalopathies such as variant Creutzfeldt-Jakob disease. Enteric glia express the normal cellular form the of the prion protein and have been implicated as a reservoir of infective prions, as they serve as a template for replication of the misfolded prion protein (70–72). Following intravenous, intraperitoneal, or intracerebral inoculation with infective prions, misfolded prion is found in enteric glia. Lawson and colleagues demonstrated that intracerebral inoculation with infectious prion protein resulted in the presence of infectious prions in the ileum at the time of clinical disease, about 220 days after inoculation (71). All animals displayed some degree of myenteric neuropathy, with loss of neurofilament M–expressing neurons in the myenteric plexus, though there were no major changes in the submucosal plexus. However, there were markedly altered and morphologically distorted enteric glia in the myenteric plexus, and where this was observed, Hu immunoreactive neurons were shrunken and there was a loss of NOS immunoreactivity. This study and a more recent study by Seelig et al. (73) demonstrate that misfolded prion protein can be transmitted from the brain to the gut, where it can cause pathological changes in enteric glia and neurons. Exactly how this transmission occurs remains to be determined, but likely it is through autonomic neural pathways, including the vagus nerve. The transmission of infective prion from the gut to the brain is well established (74) and involves the ENS and autonomic pathways. Thus enteric glia may be an important reservoir for infective prions that may go from the gut to the brain or vice versa, and they may potentially be a source of prion disease transmission through environmental contamination if the prions are shed into the gut lumen.
The second recent observation is of a role for enteric glia in Parkinson’s disease. Changes in GFAP expression and/or phosphorylation are a feature of neurodegenerative diseases. In Parkinson’s disease, enteric neurons accumulate α-synuclein and exhibit pathological features of Parkinson’s disease (75). Recently, Clairembault et al. showed that enteric glia become reactive by assessing the expression and phosphorylation levels of GFAP in colonic biopsies from patients with Parkinson’s disease (76). Compared with control subjects, those patients had significantly higher GFAP expression and the phosphorylation level of GFAP at serine 13 was significantly lower. These findings provide evidence that reactive gliosis occurs in Parkinson’s disease, and this reinforces the role of the ENS in this important neurodegenerative disease. The inflammatory state of enteric glia in Parkinson’s disease may reduce their functional capacity, contributing to the ENS dysfunction that is responsible for the constipation associated with this condition.
Conclusions and future directions
Enteric glia play an important role in GI homeostasis by acting as cellular integrators in the control of motility and barrier function (Table 1). They may also play a role in the control of intestinal secretion, but less is known in this regard and further studies are warranted. Depending on their location, enteric glia form a cellular and molecular bridge between enteric nerves, enteroendocrine cells, immune cells, and epithelial cells. It seems likely that connectivity among these cells through hemichannels or possibly gap junctions is important in serving these roles, but there is much to be determined in this regard. Novel approaches to study enteric glia, including sophisticated imaging techniques — and particularly studies that explore neuron-glial crosstalk in the ENS — will advance these investigations (31). From a clinical standpoint, enteric glia represent new and probably important targets for drug treatment in disorders of motility and host defense, notably inflammatory conditions of the gut such as inflammatory bowel disease. Their role in neurodegenerative diseases is also worthy of further consideration from both a GI and CNS perspective. The molecular constituents of enteric glia have the potential to become important biomarkers for a range of diseases. Future studies should focus on this possibility, particularly given the relative ease with which these cells can be studied in humans.
Table 1. Functional roles of enteric glia in the GI tract.
Acknowledgments
The author’s laboratory is supported by the Canadian Institutes of Health Research. Thanks to Benjamin Lee, Samantha Trautmann, and Winnie Ho for valuable discussions and help with the illustrations. K.A. Sharkey is the Crohn’s and Colitis Foundation of Canada Chair in Inflammatory Bowel Disease Research at the University of Calgary.
Footnotes
Conflict of interest: The author has declared that no conflict of interest exists.
Reference information:J Clin Invest. 2015;125(3):918–925. doi:10.1172/JCI76303.
References
- 1.Kurahashi M, et al. A functional role for the ‘fibroblast-like cells’ in gastrointestinal smooth muscles. J Physiol. 2011;589(pt 3):697–710. doi: 10.1113/jphysiol.2010.201129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Muller PA, et al. Crosstalk between muscularis macrophages and enteric neurons regulates gastrointestinal motility. Cell. 2014;158(2):300–313. doi: 10.1016/j.cell.2014.04.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Furness JB. The enteric nervous system and neurogastroenterology. Nat Rev Gastroenterol Hepatol. 2012;9(5):286–294. doi: 10.1038/nrgastro.2012.32. [DOI] [PubMed] [Google Scholar]
- 4.Gulbransen BD, Sharkey KA. Novel functional roles for enteric glia in the gastrointestinal tract. Nat Rev Gastroenterol Hepatol. 2012;9(11):625–632. doi: 10.1038/nrgastro.2012.138. [DOI] [PubMed] [Google Scholar]
- 5.Gershon MD, Rothman TP. Enteric glia. Glia. 1991;4(2):195–204. doi: 10.1002/glia.440040211. [DOI] [PubMed] [Google Scholar]
- 6.Ruhl A, Nasser Y, Sharkey KA. Enteric glia. Neurogastroenterol Motil. 2004;16(suppl 1):44–49. doi: 10.1111/j.1743-3150.2004.00474.x. [DOI] [PubMed] [Google Scholar]
- 7.Neunlist M, et al. Enteric glial cells: recent developments and future directions. Gastroenterology. 2014;147(6):1230–1237. doi: 10.1053/j.gastro.2014.09.040. [DOI] [PubMed] [Google Scholar]
- 8.Neunlist M, et al. The digestive neuronal-glial-epithelial unit: a new actor in gut health and disease. Nat Rev Gastroenterol Hepatol. 2013;10(2):90–100. doi: 10.1038/nrgastro.2012.221. [DOI] [PubMed] [Google Scholar]
- 9.Boesmans W, Lasrado R, Vanden Berghe P, Pachnis V. Heterogeneity phenotypic plasticity of glial cells in the mammalian enteric nervous system. Glia. 2014;63(2):229–241. doi: 10.1002/glia.22746. [DOI] [PubMed] [Google Scholar]
- 10.Gabella G. Ultrastructure of the nerve plexuses of the mammalian intestine: the enteric glial cells. Neuroscience. 1981;6(3):425–436. doi: 10.1016/0306-4522(81)90135-4. [DOI] [PubMed] [Google Scholar]
- 11.Gabella G. Fine structure of the myenteric plexus in the guinea-pig ileum. J Anat. 1972;111(pt 1):69–97. [PMC free article] [PubMed] [Google Scholar]
- 12.Jessen KR, Mirsky R. Glial cells in the enteric nervous system contain glial fibrillary acidic protein. Nature. 1980;286(5774):736–737. doi: 10.1038/286736a0. [DOI] [PubMed] [Google Scholar]
- 13.Hanani M, Zamir O, Baluk P. Glial cells in the guinea pig myenteric plexus are dye coupled. Brain Res. 1989;497(2):245–249. doi: 10.1016/0006-8993(89)90269-2. [DOI] [PubMed] [Google Scholar]
- 14.Geboes K, et al. Major histocompatibility class II expression on the small intestinal nervous system in Crohn’s disease. Gastroenterology. 1992;103(2):439–447. doi: 10.1016/0016-5085(92)90832-j. [DOI] [PubMed] [Google Scholar]
- 15.Koretz K, Momburg F, Otto HF, Moller P. Sequential induction of MHC antigens on autochthonous cells of ileum affected by Crohn’s disease. Am J Pathol. 1987;129(3):493–502. [PMC free article] [PubMed] [Google Scholar]
- 16.Bush TG, et al. Fulminant jejuno-ileitis following ablation of enteric glia in adult transgenic mice. Cell. 1998;93(2):189–201. doi: 10.1016/S0092-8674(00)81571-8. [DOI] [PubMed] [Google Scholar]
- 17.Cornet A, et al. Enterocolitis induced by autoimmune targeting of enteric glial cells: a possible mechanism in Crohn’s disease? Proc Natl Acad Sci U S A. 2001;98(23):13306–13311. doi: 10.1073/pnas.231474098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Aikawa H, Suzuki K. Lesions in the skin, intestine, and central nervous system induced by an antimetabolite of niacin. Am J Pathol. 1986;122(2):335–342. [PMC free article] [PubMed] [Google Scholar]
- 19.Aikawa H, Suzuki K. Enteric gliopathy in niacin-deficiency induced by CNS glio-toxin. Brain Res. 1985;334(2):354–356. doi: 10.1016/0006-8993(85)90231-8. [DOI] [PubMed] [Google Scholar]
- 20.Aubé AC, et al. Changes in enteric neurone phenotype and intestinal functions in a transgenic mouse model of enteric glia disruption. Gut. 2006;55(5):630–637. doi: 10.1136/gut.2005.067595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Nasser Y, et al. Role of enteric glia in intestinal physiology: effects of the gliotoxin fluorocitrate on motor and secretory function. Am J Physiol Gastrointest Liver Physiol. 2006;291(5):G912–G927. doi: 10.1152/ajpgi.00067.2006. [DOI] [PubMed] [Google Scholar]
- 22.Bassotti G, et al. Enteric neuropathology of the terminal ileum in patients with intractable slow-transit constipation. Hum Pathol. 2006;37(10):1252–1258. doi: 10.1016/j.humpath.2006.04.027. [DOI] [PubMed] [Google Scholar]
- 23.Bassotti G, Villanacci V, Antonelli E, Morelli A, Salerni B. Enteric glial cells: new players in gastrointestinal motility? Lab Invest. 2007;87(7):628–632. doi: 10.1038/labinvest.3700564. [DOI] [PubMed] [Google Scholar]
- 24.Bassotti G, et al. Enteric glial cells and their role in gastrointestinal motor abnormalities: introducing the neuro-gliopathies. World J Gastroenterol. 2007;13(30):4035–4041. doi: 10.3748/wjg.v13.i30.4035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gulbransen BD, Bains JS, Sharkey KA. Enteric glia are targets of the sympathetic innervation of the myenteric plexus in the guinea pig distal colon. J Neurosci. 2010;30(19):6801–6809. doi: 10.1523/JNEUROSCI.0603-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gulbransen BD, et al. Activation of neuronal P2X7 receptor-pannexin-1 mediates death of enteric neurons during colitis. Nat Med. 2012;18(4):600–604. doi: 10.1038/nm.2679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gulbransen BD, Sharkey KA. Purinergic neuron-to-glia signaling in the enteric nervous system. Gastroenterology. 2009;136(4):1349–1358. doi: 10.1053/j.gastro.2008.12.058. [DOI] [PubMed] [Google Scholar]
- 28.McClain JL, et al. Ca2+ responses in enteric glia are mediated by connexin-43 hemichannels and modulate colonic transit in mice. Gastroenterology. 2014;146(2):497–507. doi: 10.1053/j.gastro.2013.10.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Broadhead MJ, Bayguinov PO, Okamoto T, Heredia DJ, Smith TK. Ca2+ transients in myenteric glial cells during the colonic migrating motor complex in the isolated murine large intestine. J Physiol. 2012;590(pt 2):335–350. doi: 10.1113/jphysiol.2011.219519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Gulbransen BD. Enteric Glia. San Rafael, California, USA: Morgan and Claypool; 2014. [Google Scholar]
- 31.Boesmans W, et al. Imaging neuron-glia interactions in the enteric nervous system. Front Cell Neurosci. 2013;7:183. doi: 10.3389/fncel.2013.00183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Boesmans W, et al. Neurotransmitters involved in fast excitatory neurotransmission directly activate enteric glial cells. Neurogastroenterol Motil. 2013;25(2):e151–e160. doi: 10.1111/nmo.12065. [DOI] [PubMed] [Google Scholar]
- 33.Vanden Berghe P, Klingauf J. Spatial organization and dynamic properties of neurotransmitter release sites in the enteric nervous system. Neuroscience. 2007;145(1):88–99. doi: 10.1016/j.neuroscience.2006.11.048. [DOI] [PubMed] [Google Scholar]
- 34.Okamoto T, et al. Extensive projections of myenteric serotonergic neurons suggest they comprise the central processing unit in the colon. Neurogastroenterol Motil. 2014;26(4):556–570. doi: 10.1111/nmo.12302. [DOI] [PubMed] [Google Scholar]
- 35.Gautron L, et al. Neuronal and nonneuronal cholinergic structures in the mouse gastrointestinal tract and spleen. J Comp Neurol. 2013;521(16):3741–3767. doi: 10.1002/cne.23376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Stoffels B, et al. Postoperative ileus involves interleukin-1 receptor signaling in enteric glia. Gastroenterology. 2014;146(1):176–187e171. doi: 10.1053/j.gastro.2013.09.030. [DOI] [PubMed] [Google Scholar]
- 37.Green CL, Ho W, Sharkey KA, McKay DM. Dextran sodium sulfate-induced colitis reveals nicotinic modulation of ion transport via iNOS-derived NO. Am J Physiol Gastrointest Liver Physiol. 2004;287(3):G706–G714. doi: 10.1152/ajpgi.00076.2004. [DOI] [PubMed] [Google Scholar]
- 38.MacEachern SJ, Patel BA, McKay DM, Sharkey KA. Nitric oxide regulation of colonic epithelial ion transport: a novel role for enteric glia in the myenteric plexus. J Physiol. 2011;589(pt 13):3333–3348. doi: 10.1113/jphysiol.2011.207902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Selgrad M, et al. JC virus infects the enteric glia of patients with chronic idiopathic intestinal pseudo-obstruction. Gut. 2009;58(1):25–32. doi: 10.1136/gut.2008.152512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Abdo H, et al. Enteric glial cells protect neurons from oxidative stress in part via reduced glutathione. FASEB J. 2010;24(4):1082–1094. doi: 10.1096/fj.09-139519. [DOI] [PubMed] [Google Scholar]
- 41.Fu M, et al. Retinoblastoma protein prevents enteric nervous system defects and intestinal pseudo-obstruction. J Clin Invest. 2013;123(12):5152–5164. doi: 10.1172/JCI67653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Stenkamp-Strahm C, Patterson S, Boren J, Gericke M, Balemba O. High-fat diet and age-dependent effects on enteric glial cell populations of mouse small intestine. Auton Neurosci. 2013;177(2):199–210. doi: 10.1016/j.autneu.2013.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.van Haver ER, et al. Postnatal and diet-dependent increases in enteric glial cells and VIP-containing neurones in preterm pigs. Neurogastroenterol Motil. 2008;20(9):1070–1079. doi: 10.1111/j.1365-2982.2008.01160.x. [DOI] [PubMed] [Google Scholar]
- 44.Phillips RJ, Kieffer EJ, Powley TL. Loss of glia and neurons in the myenteric plexus of the aged Fischer 344 rat. Anat Embryol (Berl). 2004;209(1):19–30. doi: 10.1007/s00429-004-0426-x. [DOI] [PubMed] [Google Scholar]
- 45.Baudry C, et al. Diet-induced obesity has neuroprotective effects in murine gastric enteric nervous system: involvement of leptin and glial cell line-derived neurotrophic factor. J Physiol. 2012;590(pt 3):533–544. doi: 10.1113/jphysiol.2011.219717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Schoffen JP, et al. Food restriction enhances oxidative status in aging rats with neuroprotective effects on myenteric neuron populations in the proximal colon. Exp Gerontol. 2014;51:54–64. doi: 10.1016/j.exger.2014.01.001. [DOI] [PubMed] [Google Scholar]
- 47.Voss U, Sand E, Olde B, Ekblad E. Enteric neuropathy can be induced by high fat diet in vivo and palmitic acid exposure in vitro. PLoS One. 2013;8(12):e81413. doi: 10.1371/journal.pone.0081413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Liu YA, et al. 3-D imaging, illustration, and quantitation of enteric glial network in transparent human colon mucosa. Neurogastroenterol Motil. 2013;25(5):e324–e338. doi: 10.1111/nmo.12115. [DOI] [PubMed] [Google Scholar]
- 49.Badizadegan K, et al. Presence of intramucosal neuroglial cells in normal and aganglionic human colon. Am J Physiol Gastrointest Liver Physiol. 2014;307(10):G1002–G1012. doi: 10.1152/ajpgi.00164.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bohórquez DV, et al. An enteroendocrine cell-enteric glia connection revealed by 3D electron microscopy. PLoS One. 2014;9(2):e89881. doi: 10.1371/journal.pone.0089881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Raybould HE. Gut chemosensing: interactions between gut endocrine cells and visceral afferents. Auton Neurosci. 2010;153(1–2):41–46. doi: 10.1016/j.autneu.2009.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Sternini C, Anselmi L, Rozengurt E. Enteroendocrine cells: a site of ‘taste’ in gastrointestinal chemosensing. Curr Opin Endocrinol Diabetes Obes. 2008;15(1):73–78. doi: 10.1097/MED.0b013e3282f43a73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Drucker DJ, Yusta B. Physiology and pharmacology of the enteroendocrine hormone glucagon-like peptide-2. Annu Rev Physiol. 2014;76:561–583. doi: 10.1146/annurev-physiol-021113-170317. [DOI] [PubMed] [Google Scholar]
- 54.de Heuvel E, Wallace L, Sharkey KA, Sigalet DL. Glucagon-like peptide 2 induces vasoactive intestinal polypeptide expression in enteric neurons via phophatidylinositol 3-kinase-γ signaling. Am J Physiol Endocrinol Metab. 2012;303(8):E994–E1005. doi: 10.1152/ajpendo.00291.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Savidge TC, et al. Enteric glia regulate intestinal barrier function and inflammation via release of S-nitrosoglutathione. Gastroenterology. 2007;132(4):1344–1358. doi: 10.1053/j.gastro.2007.01.051. [DOI] [PubMed] [Google Scholar]
- 56.Neunlist M, et al. Enteric glia inhibit intestinal epithelial cell proliferation partly through a TGF-β1-dependent pathway. Am J Physiol Gastrointest Liver Physiol. 2007;292(1):G231–G241. doi: 10.1152/ajpgi.00276.2005. [DOI] [PubMed] [Google Scholar]
- 57.Xiao W, et al. GDNF is involved in the barrier-inducing effect of enteric glial cells on intestinal epithelial cells under acute ischemia reperfusion stimulation. Mol Neurobiol. 2014;50(2):274–289. doi: 10.1007/s12035-014-8730-9. [DOI] [PubMed] [Google Scholar]
- 58.Cheadle GA, Costantini TW, Bansal V, Eliceiri BP, Coimbra R. Cholinergic signaling in the gut: a novel mechanism of barrier protection through activation of enteric glia cells. Surg Infect (Larchmt). 2014;15(4):387–393. doi: 10.1089/sur.2013.103. [DOI] [PubMed] [Google Scholar]
- 59.Cheadle GA, et al. Enteric glia cells attenuate cytomix-induced intestinal epithelial barrier breakdown. PLoS One. 2013;8(7):e69042. doi: 10.1371/journal.pone.0069042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Costantini TW, et al. Targeting α-7 nicotinic acetylcholine receptor in the enteric nervous system: a cholinergic agonist prevents gut barrier failure after severe burn injury. Am J Pathol. 2012;181(2):478–486. doi: 10.1016/j.ajpath.2012.04.005. [DOI] [PubMed] [Google Scholar]
- 61.Turco F, et al. Enteroglial-derived S100B protein integrates bacteria-induced Toll-like receptor signalling in human enteric glial cells. Gut. 2013;63(1):105–115. doi: 10.1136/gutjnl-2012-302090. [DOI] [PubMed] [Google Scholar]
- 62.Esposito G, et al. Palmitoylethanolamide improves colon inflammation through an enteric glia/toll like receptor 4-dependent PPAR-α activation. Gut. 2014;63(8):1300–1312. doi: 10.1136/gutjnl-2013-305005. [DOI] [PubMed] [Google Scholar]
- 63.Esposito G, et al. Enteric glial-derived S100B protein stimulates nitric oxide production in celiac disease. Gastroenterology. 2007;133(3):918–925. doi: 10.1053/j.gastro.2007.06.009. [DOI] [PubMed] [Google Scholar]
- 64.Cirillo C, et al. Proinflammatory stimuli activates human-derived enteroglial cells and induces autocrine nitric oxide production. Neurogastroenterol Motil. 2011;23(9):e372–e382. doi: 10.1111/j.1365-2982.2011.01748.x. [DOI] [PubMed] [Google Scholar]
- 65.Cirillo C, et al. Increased mucosal nitric oxide production in ulcerative colitis is mediated in part by the enteroglial-derived S100B protein. Neurogastroenterol Motil. 2009;21(11):1209–e112. doi: 10.1111/j.1365-2982.2009.01346.x. [DOI] [PubMed] [Google Scholar]
- 66.Xiao WD, et al. The protective effect of enteric glial cells on intestinal epithelial barrier function is enhanced by inhibiting inducible nitric oxide synthase activity under lipopolysaccharide stimulation. Mol Cell Neurosci. 2011;46(2):527–534. doi: 10.1016/j.mcn.2010.12.007. [DOI] [PubMed] [Google Scholar]
- 67.da Silveira AB, et al. Enteroglial cells act as antigen-presenting cells in chagasic megacolon. Hum Pathol. 2011;42(4):522–532. doi: 10.1016/j.humpath.2010.06.016. [DOI] [PubMed] [Google Scholar]
- 68.Esposito G, et al. Palmitoylethanolamide improves colon inflammation through an enteric glia/toll like receptor 4-dependent PPAR-α activation. Gut. 2013;63(8):1300–1312. doi: 10.1136/gutjnl-2013-305005. [DOI] [PubMed] [Google Scholar]
- 69.Storr MA, et al. Targeting endocannabinoid degradation protects against experimental colitis in mice: involvement of CB1 and CB2 receptors. J Mol Med (Berl). 2008;86(8):925–936. doi: 10.1007/s00109-008-0359-6. [DOI] [PubMed] [Google Scholar]
- 70.Albanese V, et al. Evidence for prion protein expression in enteroglial cells of the myenteric plexus of mouse intestine. Auton Neurosci. 2008;140(1–2):17–23. doi: 10.1016/j.autneu.2008.01.008. [DOI] [PubMed] [Google Scholar]
- 71.Lawson VA, et al. The brain to gut pathway: a possible route of prion transmission. Gut. 2010;59(12):1643–1651. doi: 10.1136/gut.2010.222620. [DOI] [PubMed] [Google Scholar]
- 72.Martin GR, et al. Endogenous cellular prion protein regulates contractility of the mouse ileum. Neurogastroenterol Motil. 2012;24(9):e412–e424. doi: 10.1111/j.1365-2982.2012.01970.x. [DOI] [PubMed] [Google Scholar]
- 73.Seelig DM, Mason GL, Telling GC, Hoover EA. Chronic wasting disease prion trafficking via the autonomic nervous system. Am J Pathol. 2011;179(3):1319–1328. doi: 10.1016/j.ajpath.2011.05.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ano Y, Sakudo A, Nakayama H, Onodera T. Uptake and dynamics of infectious prion protein in the intestine. Protein Pept Lett. 2009;16(3):247–255. doi: 10.2174/092986609787601642. [DOI] [PubMed] [Google Scholar]
- 75.Clairembault T, Leclair-Visonneau L, Neunlist M, Derkinderen P. Enteric glial cells: new players in Parkinson’s disease? Mov Disord. doi: 10.1002/mds.25979. [published online ahead of print August 7, 2014]. doi: 10.1002/mds.25979. [DOI] [PubMed] [Google Scholar]
- 76.Clairembault T, et al. Enteric GFAP expression and phosphorylation in Parkinson’s disease. J Neurochem. 2014;130(6):805–815. doi: 10.1111/jnc.12742. [DOI] [PubMed] [Google Scholar]
- 77.Van Landeghem L, et al. Enteric glia promote intestinal mucosal healing via activation of focal adhesion kinase and release of proEGF. Am J Physiol Gastrointest Liver Physiol. 2011;300(6):G976–G987. doi: 10.1152/ajpgi.00427.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Bach-Ngohou K, et al. Enteric glia modulate epithelial cell proliferation and differentiation through 15-deoxy-12,14-prostaglandin J2. J Physiol. 2010;588(pt 14):2533–2544. doi: 10.1113/jphysiol.2010.188409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.von Boyen GB, et al. Nerve growth factor secretion in cultured enteric glia cells is modulated by proinflammatory cytokines. J Neuroendocrinol. 2006;18(11):820–825. doi: 10.1111/j.1365-2826.2006.01478.x. [DOI] [PubMed] [Google Scholar]
- 80.Abdo H, et al. The omega-6 fatty acid derivative 15-deoxy-Δ(1)(2),(1)(4)-prostaglandin J2 is involved in neuroprotection by enteric glial cells against oxidative stress. J Physiol. 2012;590(pt 11):2739–2750. doi: 10.1113/jphysiol.2011.222935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Nagahama M, Semba R, Tsuzuki M, Aoki E. L-arginine immunoreactive enteric glial cells in the enteric nervous system of rat ileum. Biol Signals Recept. 2001;10(5):336–340. doi: 10.1159/000046901. [DOI] [PubMed] [Google Scholar]
- 82.Aoki E, Semba R, Kashiwamata S. Evidence for the presence of L-arginine in the glial components of the peripheral nervous system. Brain Res. 1991;559(1):159–162. doi: 10.1016/0006-8993(91)90300-k. [DOI] [PubMed] [Google Scholar]
- 83.Jessen KR, Mirsky R. Astrocyte-like glia in the peripheral nervous system: an immunohistochemical study of enteric glia. J Neurosci. 1983;3(11):2206–2218. doi: 10.1523/JNEUROSCI.03-11-02206.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zhang W, Segura BJ, Lin TR, Hu Y, Mulholland MW. Intercellular calcium waves in cultured enteric glia from neonatal guinea pig. Glia. 2003;42(3):252–262. doi: 10.1002/glia.10215. [DOI] [PubMed] [Google Scholar]
- 85.Gomes P, et al. ATP-dependent paracrine communication between enteric neurons and glia in a primary cell culture derived from embryonic mice. Neurogastroenterol Motil. 2009;21(8):870–e62. doi: 10.1111/j.1365-2982.2009.01302.x. [DOI] [PubMed] [Google Scholar]
- 86.Joseph NM, et al. Enteric glia are multipotent in culture but primarily form glia in the adult rodent gut. J Clin Invest. 2011;121(9):3398–3411. doi: 10.1172/JCI58186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Laranjeira C, et al. Glial cells in the mouse enteric nervous system can undergo neurogenesis in response to injury. J Clin Invest. 2011;121(9):3412–3424. doi: 10.1172/JCI58200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Hirata I, Austin LL, Blackwell WH, Weber JR, Dobbins WO., 3rd Immunoelectron microscopic localization of HLA-DR antigen in control small intestine and colon and in inflammatory bowel disease. Dig Dis Sci. 1986;31(12):1317–1330. doi: 10.1007/BF01299810. [DOI] [PubMed] [Google Scholar]
- 89.Murakami M, Ohta T, Ito S. Lipopolysaccharides enhance the action of bradykinin in enteric neurons via secretion of interleukin-1beta from enteric glial cells. J Neurosci Res. 2009;87(9):2095–2104. doi: 10.1002/jnr.22036. [DOI] [PubMed] [Google Scholar]
- 90.Murakami M, Ohta T, Otsuguro KI, Ito S. Involvement of prostaglandin E(2) derived from enteric glial cells in the action of bradykinin in cultured rat myenteric neurons. Neuroscience. 2007;145(2):642–653. doi: 10.1016/j.neuroscience.2006.12.052. [DOI] [PubMed] [Google Scholar]
- 91.Ruhl A, Franzke S, Collins SM, Stremmel W. Interleukin-6 expression and regulation in rat enteric glial cells. Am J Physiol Gastrointest Liver Physiol. 2001;280(6):G1163–G1171. doi: 10.1152/ajpgi.2001.280.6.G1163. [DOI] [PubMed] [Google Scholar]