

Abstract
Although engagement of medial prefrontal cortex (MPFC) underlies self-referencing of information for younger and older adults, the region has not consistently been implicated across age groups for the encoding of self-referenced information. We sought to determine whether making judgments about others as well as the self influenced findings in the previous study. During an fMRI session, younger and older adults encoded adjectives using only a self-reference task. For items later remembered compared to those later forgotten, both age groups robustly recruited medial prefrontal cortex, indicating common neural regions support encoding across younger and older adults when participants make only self-reference judgments. Focal age differences emerged in regions related to emotional processing and cognitive control, though these differences are more limited than in tasks in which judgments also are made about others. We conclude that making judgments about another person differently affects the ways that younger and older adults make judgments about the self, with results of a follow-up behavioral study supporting this interpretation.
1. Introduction
Because older adults often experience deficits in memory, it is important to study techniques that can potentially improve memory function with age. One such technique is self referencing. Previous research has shown that relating information to oneself is a successful encoding strategy (Symons & Johnson, 1997). Adjectives encoded in a self referential manner are retrieved more effectively than those encoded in a semantic, structural, or phonemic manner (Rogers, Kuiper, & Kirker, 1977). The literature suggests that the self is a unique cognitive structure that possesses special mnemonic capabilities (Macrae, Moran, Heatherton, Banfield, & Kelley, 2004). Consistent with this idea, self-referencing has reliably been associated with corresponding activity in the medial prefrontal cortex (mPFC) (Kelley et al., 2002; Macrae et al., 2004; Northoff et al., 2006). Activity in mPFC is elicited in tasks that range from judging whether adjectives describe oneself (e.g., Kelley et al., 2002; Moran, Heatherton, & Kelley, 2009), to self-reflection on physical, mental, or personality attributes (Jenkins & Mitchell, 2011), to self-evaluation of performance (Beer, Lombardo, & Bhanji, 2010), to self-ownership of objects (Turk, van Bussel, Waiter, & Macrae, 2011).
A number of studies show that a self-referencing strategy benefits memory for older adults as much as young adults’ memory, for a commonly-used adjective task (Glisky & Marquine, 2009; Gutchess, Kensinger, Yoon, & Schacter, 2007; Mueller, Wonderlich, & Dugan, 1986), as well as for visual details (Hamami, Serbun, & Gutchess, 2011) and the source of actions (Rosa & Gutchess, 2011). Results also suggest that the strategy operates spontaneously and effortlessly, without depending on attention (Yang, Truong, Fuss, & Bislimovic, 2012). Correspondingly, functional imaging has shown that the mPFC is similarly activated in younger and older adults during self-referencing (Gutchess, Kensinger, & Schacter, 2007). As a result, the activity of the mPFC may remain robust in older adults despite other age related changes to the brain.
The contribution of the mPFC in older adults is less clear when memory is considered. The mPFC has been implicated for younger adults, with activity in the mPFC predicting subsequent memory performance (Macrae et al., 2004). Our previous study attempted to characterize the neural correlates of self-referencing in older adults compared to younger adults during encoding of adjectives. Using a subsequent memory paradigm (Paller & Wagner, 2002), activity was compared at the initial time of encoding for words later remembered compared to words later forgotten (Gutchess, Kensinger, & Schacter, 2010). Results were surprising in that no regions converged across age groups to support the encoding of information into memory. Regions, including superior mPFC, anterior and posterior cingulate, and left inferior prefrontal cortex, exhibiting subsequent memory effects for other person judgments in older adults were associated with forgetting in the self-referential condition for young adults. These findings were unexpected, as previous work implicated mPFC across age groups when making judgments of self vs. others across the age groups (Gutchess, Kensinger, Yoon, et al., 2007) and for subsequent memory in young adults (Macrae et al., 2004). Other neural evidence also suggests that older adults should converge with younger adults with some similar components implicated in self-referencing. Dorsal medial prefrontal cortex was implicated across the age groups for accurate subsequent source memory (Leshikar & Duarte, 2013). With ERP, an earlier emerging old-new component supports self-referential encoding across both age groups (Dulas, Newsome, & Duarte, 2011).
Our previous study that did not find convergence across the age groups employed a typical self-referencing paradigm in which older and younger participants were presented with adjectives at encoding and were asked to make a yes/no response regarding the orienting condition indicated for the trial. The three orienting conditions were the self condition, the other condition, and the case condition. Encoding was followed by a surprise recognition task outside of the scanner. It was predicted that in both younger and older adults, the mPFC would be preferentially activated at encoding during self-referential trials compared to other-referential trials, resulting in a subsequent remembering effect (i.e. greater activity in the mPFC during encoding reflects later memory for the item). However, the results indicated reversals in patterns of activity across age groups rather than failure of one group to differentially engage regions for successful versus unsuccessful encoding. The younger adults exhibited subsequent forgetting effects in all regions of interest (left inferior frontal, left superior mPFC, anterior cingulate, right posterior cingulate) for self-referenced items, and subsequent remembering effects in all regions for other referenced items. The older adults demonstrated a reverse pattern of activity in which they showed remembering effects in all regions of interest for self-referenced items, and forgetting effects in all regions of interest for other-referenced items.
These findings could be the result of fundamental differences between younger and older adults in strategies and attention to incoming information. It is possible that older adults engage elaborative processes that benefit memory for the self while young adults use the same processes to encode information for others (Gutchess et al., 2010). However, this contradicts previous research which has shown that both younger adults and older adults exhibit subsequent memory effects for self referenced information (Glisky & Marquine, 2009; Gutchess, Kensinger, Yoon, et al., 2007) and engage the same region for subsequent source memory (Leshikar & Duarte, 2013).
It is also possible that the task design contributed to the pattern of findings in the Gutchess et al. (2010) paper. Because the task paradigm incorporated three conditions in an intermixed pseudorandom design, it is possible that interference across conditions, or task switching demands, impacted the engagement of mPFC. Participants may have made the wrong judgment, or making judgments about other people could differentially impact how older adults make judgments about the self. The identification of dorsal mPFC and anterior cingulate (ACC) in the prior study is consistent with research revealing that these regions can be engaged when thinking about dissimilar vs. similar others (Mitchell, Macrae, & Banaji, 2006), which could suggest that participants thought about themselves in a relative manner. Another concern with the prior data is that there were relatively few forgotten trials in the young adults; this could have contributed to our failure to find robust subsequent memory effects.
In the current study, we utilized the same subsequent memory paradigm as our prior study, except that we sought to reduce the potential for interference by eliminating the other and the case conditions. By maximizing the number of trials associated with self-referencing we hoped to parse out the brain regions that are active in younger and older adults exclusively during the encoding of self-referenced information. We also wanted to acquire a large number of remembered and forgotten self-referencing trials to better power the comparisons. To achieve this, we employed subsequent recall and recognition tasks (rather than only a subsequent recognition task) in order to increase the number of forgotten items, as should be the case for free recall.
2. Results
Two analyses were conducted on the fMRI data to compare younger and older adults, based on items later free recalled and items later recognized.
2.1 Behavioral results
On the free recall test, young recalled significantly more words (M = 29.18, SD = 11.32) than older adults (M = 20.56, SD = 9.97), t(31) = 2.32, p=.027. On the recognition test, young (M = .59, SD = .18) also significantly outperformed older adults (M = .41, SD = .20) on a measure of corrected recognition (hits minus false alarms), t(21) = 2.68, p=.012.
2.2 Neuroimaging results
2.2.1 Recall analyses
Table 1a&b displays all significant neural coordinates from recall analyses. We first identified regions activated in common across young and older adults. This analysis revealed that a large cluster of activation in medial prefrontal cortex, centered in BA 10 but slightly left of the midline regions commonly identified in the self-referencing literature. See Figure 1a. In terms of age differences, small focal difference emerged, including in orbitofrontal cortex (BA 11), in which young adults exhibited subsequent memory effects whereas older adults tended to show subsequent forgetting effects. Older adults exhibited subsequent memory effects in a small region of middle frontal cortex (BA 46), whereas young adults tended to show subsequent forgetting effects in this region. See Figure 2a & b.
Table 1.
| Table 1A | ||||||
|---|---|---|---|---|---|---|
| A. Common activations across Younger and Older adults for Recall | ||||||
| Region | BA | Activation P eak (x,y,z) | # Voxels | t-value | ||
| L Superior Frontal | 32 | −14 | 48 | 24 | 246 | 3.33 |
| L Anterior Cingulum | 32 | −10 | 42 | 12 | 2.77 | |
| L Superior Medial Frontal | 10 | −10 | 54 | 6 | 2.00 | |
| R Superior Frontal | 9 | 16 | 56 | 40 | 977 | 2.93 |
| L Posterior Cingulum | 23 | −6 | −48 | 32 | 17 | 2.93 |
| R Middle Temporal | 21 | 74 | −16 | −12 | 106 | 2.83 |
| R Cerebellum | N/A | 40 | −60 | −36 | 27 | 2.61 |
| R Middle Temporal Pole | 20 | 58 | 22 | −46 | 69 | 2.49 |
| L Cerebellum | N/A | −4 | −52 | −42 | 30 | 2.46 |
| R Cerebellum | N/A | 6 | −52 | −46 | 2.08 | |
| R Middle Temporal | 21 | 70 | −36 | −6 | 36 | 2.43 |
| L Angular | 39 | −58 | −56 | 30 | 128 | 2.41 |
| L Angular | 39 | −48 | −64 | 34 | 2.13 | |
| L Angular | 39 | −56 | −64 | 30 | 1.96 | |
| L Cerebellum | N/A | −36 | −70 | −32 | 33 | 2.37 |
| L Middle Temporal Pole | 38 | −50 | 18 | −36 | 41 | 2.22 |
| L Middle Temporal Pole | 20 | −44 | 20 | −42 | 2.19 | |
| R Angular | 39 | 50 | −62 | 30 | 27 | 2.16 |
| R Postcentral | 5 | 14 | −50 | 82 | 7 | 2.13 |
| L Middle Temporal | 21 | −44 | −50 | 22 | 8 | 2.06 |
| L Middle Frontal | 9 | −46 | 26 | 44 | 20 | 2.04 |
| L Cerebellum | N/A | −30 | −60 | −36 | 15 | 1.98 |
| R Inferior Frontal | 45 | 54 | 28 | 2 | 14 | 1.91 |
| L Middle Temporal | 21 | −68 | −32 | −2 | 8 | 1.81 |
| Table 1B | ||||||
|---|---|---|---|---|---|---|
| B. Age differences for Recall | ||||||
| Region | BA | Activation P eak (x,y,z) | # Voxels | t-value | ||
| Younger – Older | ||||||
| L Orbitofrontal | 11 | −14 | 54 | −8 | 23 | 4.17 |
| L Inferior Temporal | 21 | −66 | −20 | −18 | 9 | 3.53 |
| Older - Younger | ||||||
| L Lingual | 19 | −24 | −68 | −4 | 292 | 4.33 |
| L Middle Occipital | 19 | −24 | −76 | 12 | 4.20 | |
| L Precuneus | 5 | −10 | −44 | 60 | 30 | 4.20 |
| L Cerebellum | N/A | −48 | −54 | −50 | 44 | 3.90 |
| R Supplementary Motor | 6 | 16 | −14 | 56 | 23 | 3.78 |
| R Superior Temporal | 22 | 62 | −10 | −4 | 9 | 3.74 |
| L Lingual | 18 | −6 | −68 | −2 | 15 | 3.73 |
| L Midcingulate | 23 | −18 | −16 | 46 | 8 | 3.67 |
| L Middle Frontal | 46 | −40 | 38 | 40 | 23 | 3.67 |
| R Supramarginal | 48 | 52 | −22 | 26 | 7 | 3.64 |
| R Inferior Temporal | 37 | 56 | −68 | −2 | 20 | 3.60 |
| R Midcingulate | 23 | 14 | −20 | 38 | 6 | 3.60 |
| R Inferior Occipital | 19 | 48 | −74 | −10 | 30 | 3.55 |
Figure 1.

Commonalities across younger and older adults in subsequent memory (remembered versus forgotten trials). Results show significant activations in the mPFC for both recall (Panel a) and recognition (Panel b).
Figure 2.
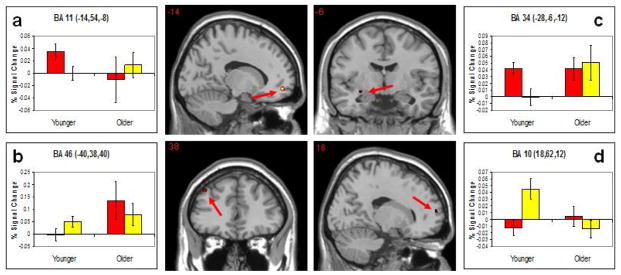
Age differences in subsequent memory for remembered versus forgotten trials. Red bars denote remembered trials and yellow bars denote forgotten trials. Results of the recall analysis show significant activations in the orbitofrontal cortex (Panel a) for younger-older adults and the middle PFC (Panel b) for older-younger adults. Results of the recognition analysis show significant activations in the amygdala (Panel c) for younger-older adults and the superior PFC (Panel d) for older-younger adults.
2.2.2 Recognition analyses
All significant coordinates of neural regions resulting from recognition analyses are presented in Table 2a&b. Common activations across younger and older adults, identified on the basis of recognition data, largely converged with recall analyses. Medial prefrontal cortex, focused more dorsally than the region identified in the recognition analyses, was engaged more for words later correctly recognized compared to those later forgotten for both younger and older adults. See Figure 2b. In terms of age differences, these were again focal, small differences. Young adults engaged the amygdala for items later remembered over forgotten, whereas older adults tended to show a subsequent forgetting effect in this region. Age differences also emerged in superior frontal regions (in BA 10) but these differences reflected a subsequent forgetting effect in young adults whereas the region did not reliable discriminate remembered from forgotten for older adults. See Figure 3c & d.
Table 2.
| Table 2A | ||||||
|---|---|---|---|---|---|---|
| A. Common activations across Younger and Older adults for Recognition | ||||||
| Region | BA | Activation Peak | # Voxels | t-value | ||
| R Superior Medial Prefrontal | 8 | 8 | 38 | 48 | 1182 | 5.14 |
| L Superior Medial Frontal | 8 | −4 | 38 | 46 | 4.15 | |
| L Superior Frontal | 10 | −14 | 58 | 16 | 3.87 | |
| R Cerebellum | N/A | 34 | −58 | −46 | 1283 | 4.59 |
| R Cerebellum | N/A | 40 | −50 | −40 | 2.97 | |
| R Cerebellum | N/A | 14 | −84 | −44 | 2.86 | |
| L Inferior Occipital | 19 | −54 | −76 | −18 | 155 | 4.15 |
| R Cerebellum (Vermis) | N/A | 8 | −54 | −22 | 224 | 3.58 |
| L Cerebellum (Vermis) | N/A | −4 | −54 | −22 | 3.24 | |
| L Angular | 39 | −38 | −56 | 30 | 138 | 3.43 |
| L Middle Occipital | 19 | −34 | −62 | 26 | 2.89 | |
| L Angular | 39 | −36 | −52 | 22 | 2.61 | |
| L Inferior Frontal | 47 | −52 | 40 | −12 | 645 | 3.16 |
| L Inferior Orbitofrontal | 47 | −30 | 26 | −14 | 3.15 | |
| R Inferior Frontal | 47 | 44 | 34 | −16 | 89 | 3.05 |
| R Inferior Orbitofrontal | 47 | 36 | 36 | −14 | 2.75 | |
| L Inferior Frontal | 44 | −38 | 20 | 32 | 49 | 3.01 |
| L Inferior Temporal | 20 | −54 | −20 | −20 | 38 | 2.98 |
| R Middle Frontal | 9 | 44 | 20 | 58 | 9 | 2.94 |
| L Cerebellum | N/A | −32 | −54 | −38 | 37 | 2.89 |
| R Middle Frontal | 10 | 28 | 48 | 8 | 24 | 2.85 |
| L Precuneus | 5 | −12 | −56 | 80 | 33 | 2.85 |
| L Superior Parietal | 5 | −18 | −50 | 80 | 2.60 | |
| L Inferior Frontal | 47 | −28 | 34 | 6 | 13 | 2.76 |
| R Precuneus | 7 | 16 | −78 | 84 | 263 | 2.74 |
| R Precuneus | 7 | 8 | −70 | 84 | 2.26 | |
| L Thalamus | N/A | −12 | −10 | 2 | 95 | 2.55 |
| L Precentral | 48 | −40 | 6 | 48 | 26 | 2.49 |
| L Superior Frontal | 10 | −30 | 64 | 8 | 30 | 2.45 |
| L Orbitofrontal | 11 | −18 | 48 | −6 | 12 | 2.38 |
| L Cerebellum | N/A | −16 | −40 | −40 | 29 | 2.36 |
| L Cerebellum | N/A | −18 | −40 | −32 | 1.84 | |
| L Cerebellum | N/A | −8 | −84 | −38 | 16 | 2.29 |
| R Thalamus | N/A | 20 | −6 | 6 | 36 | 2.25 |
| R Thalamus | N/A | 8 | −6 | 2 | 2.08 | |
| R Superior Parietal | 7 | 30 | −64 | 66 | 108 | 2.11 |
| L Superior Frontal | 9 | −10 | 52 | 50 | 45 | 2.20 |
| R Middle Temporal | 39 | 36 | −60 | 18 | 22 | 2.07 |
| L Inferior Temporal | 20 | −52 | −46 | −18 | 18 | 2.06 |
| R Superior Medial Frontal | 10 | 8 | 62 | 30 | 6 | 2.05 |
| L Cerebellum | N/A | −12 | −72 | −28 | 11 | 1.96 |
| L Middle Frontal | 8 | −34 | 24 | 54 | 5 | 1.92 |
| L Inferior Parietal | 7 | −32 | −58 | 48 | 5 | 1.90 |
| Table 2B | ||||||
|---|---|---|---|---|---|---|
| B. Age differences for Recognition | ||||||
| Region | BA | Activation P eak (x,y,z) | # Voxels | t-value | ||
| Younger – Older | ||||||
| L Precentral | 44 | −62 | 18 | 44 | 35 | 3.97 |
| L Amygdala | 34 | −28 | −6 | −12 | 5 | 3.75 |
| Older - Younger | ||||||
| R Superior Frontal | 10 | 18 | 62 | 12 | 6 | 3.58 |
Figure 3.

Schematic of the task design.
3. Discussion
Across analyses based on recall and recognition data, the results largely converged with prior work implicating mPFC with thinking about the self in younger and older adults (Gutchess, Kensinger, & Schacter, 2007) and with subsequent memory for information related to the self in young adults (Macrae et al., 2004). While the precise locus of the activations differed across the analyses, successful encoding as assessed with recall and recognition measures robustly engaged mPFC across both age groups. These results differ from our prior study of self-referential encoding with age (Gutchess et al., 2010), which failed to identify any regions commonly activated across young and older adults. The current study employed methods largely similar to the prior study, including encoding the same number of words and the use of a recognition paradigm for a subset of the analyses. The primary difference reflected the fact that participants only made judgments about the self-descriptiveness of the words. Judgments were not alternated with judgments of another person in the present study. Thus, we interpret results of this study as reflecting the similarity of the processes engaged across age groups that lead to the successful encoding of information in memory. The robust, convergent results across comparisons of recognition and recall suggest that results are not overall impacted by low numbers of trials in some conditions, as these two memory measures have contrasting profiles: recall analyses have many forgotten trials, whereas recognition analyses have fewer. The divergence of these results from our prior work provides indirect evidence for the idea that a task that includes judgments of both self and other differently impacts the way that young and older adults encode, and perhaps reflect upon, the self.
The age differences revealed by both our recall and recognition analyses are circumscribed, in contrast to the robust pattern of age differences that emerged in our prior study (Gutchess et al., 2010). These differences, however, indicate some intriguing potential differences in the processes that contribute to the successful, or unsuccessful encoding of information into memory. Engaging orbitofrontal cortex for successful recall and the amygdala for successful recognition suggest that emotional processes may contribute to accurate and successful memory formation for younger adults (Dolcos, LaBar, & Cabeza, 2004; Kensinger & Corkin, 2004; Murty, Ritchey, Adcock, & LaBar, 2010), but not for older adults in this task. Whereas this pattern did not emerge in our prior study (Gutchess et al., 2010), we note that the present study used words that were more strongly valenced. The other prefrontal regions implicated by the encoding task suggest that cognitive control regions, such as middle frontal cortex, may support older adults’ successful encoding of words that are later recalled. Engaging cognitive control regions may even impair young adults’ ability to encode the items, as superior frontal cortex is associated with subsequent forgetting in young, but not older, adults. Although speculative, this may reflect the inhibition of external information that is necessary to focus on internal information during social and self-relevant tasks (Buckner, Andrews-Hanna, & Schacter, 2008; Gusnard, Akbudak, Shulman, & Raichle, 2001).
While the present finding of robust mPFC activation during successful encoding poses a striking contrast to our prior results, the lack of a direct comparison poses a challenge to interpreting the processes that differ across the age groups. To further explore the contribution of thinking about another person to thinking about the self, we conducted a follow-up behavioral study to directly investigate the impact on memory across the age groups.
4. Experimental Procedure
4.1 Participants
Seventeen young (ages 18–35; 10 females) and sixteen older (ages 66–83; 10 females) participants volunteered in this study in exchange for monetary compensation1. See Table 3 for participant characteristics. Due to insufficient numbers of trials, one young adult was removed from analyses of recall data (but included in recognition analyses) and two young adults and one older adult were removed from analyses of recognition data (but included in recall analyses) due to insufficient numbers of trials. One additional older participant was not able to participate in the fMRI portion of the study due to discomfort while lying in the head-only scanner. Eligibility for the fMRI portion included right-handedness, English as the primary language, normal or corrected vision, good neurological, psychological and physical health, an absence of medications that affect the central nervous system, and completion of at least 10th grade education. The study was approved by the Partners Institutional Review Board and Brandeis University Institutional Review Boards.
Table 3.
Means and standard deviations for demographics and neuropsychological measures
| Younger | Older | p-value | |
|---|---|---|---|
| Age | 23.41 (4.40) | 76.25 (5.01) | <.001 |
| Years of Educ | 15.53 (1.83) | 16.63 (2.50) | .16 |
| Digit Comparison | 80.41 (14.99) | 55.81 (10.03) | <.001 |
| Shipley Vocabulary | 34.47 (3.57) | 36.63 (2.96) | .07 |
| Mini-mental score | N/A | 28.44 (1.32) |
4.2 Materials and procedure
Informed consent was obtained from each participant before beginning the study. Older participants first completed a pre-test on an earlier date consisting of behavioral and neuropsychological measures (described in section 4.3), whereas young adults completed them at the scanner.
Participants were familiarized with a brief practice session before entering the scanner. Once inside the scanner, approximately 10 minutes of structural MRIs were acquired while the participants were asked to relax. Participants then started the task, viewed 144 different adjectives. Half of the words were positive and half were negative. There were 72 additional lures, or new words, used at recognition outside of the scanner. These 72 lures were rotated in with the other 144 words to create three unique testing conditions.
Participants were asked to judge whether the adjective they saw did or did not describe themselves. The task was presented using E-Prime software (Psychology Software Tools, Pittsburgh, PA), which recorded the participants’ yes or no response from a button box. Participants were instructed to press one of two buttons on the button box, either using their index finger for a “yes” response or using their middle finger for a “no” response. Figure 3 illustrates a schematic of the trial types. Trials in the scanner lasted for a fixed duration of 3 seconds, with 4 seconds for a response, followed by 2–20 jittered seconds of a fixation cross. There were 3 runs of word judgments, each lasting 5 minutes with a short break in between sets. The scanning session lasted approximately 45 minutes. Once outside of the scanner a delay of 30 minutes was administered before beginning a computer recognition task. After the first 15 minutes of the delay, both groups of younger and older participants completed a surprise paper based free recall task. They were asked to, “write down as many words as you remember seeing while in the scanner.” This task was followed by a self paced computer based recognition task (216 trials including 144 old words and 72 new lure words presented in a random order). They responded using yes/no buttons on a keyboard. Afterwards each participant was paid and debriefed.
4.3 Neuropsychological measures
To attain an older population with normal, non-demented cognitive function, we used the Mini-Mental State Examination (MMSE; Folstein, Folstein, & McHugh, 1975) to assess cognitive function. All elderly participants achieved a score of 27 or better (out of 30) on the MMSE, assuring that our sample did not include demented participants. Both groups of young and old participants also completed speed of processing (Hedden et al., 2002) and vocabulary (Shipley, 1986) tasks. Scores can be found in Table 3.
4.4 Image acquisition and data analysis
Data were acquired using a Siemens Avanto 1.5T scanner, using an echo-planar imaging (EPI) sequence (TR=2000ms; TE=40ms) to acquire 26 slices (AC/PC oriented) 5mm thick (20% skip). The stimuli were back-projected onto a white screed located behind the scanner, and the participants viewed images using a mirror attached to the headcoil. High-resolution anatomical images were acquired using a multiplanar rapidly acquired gradient echo (MP-RAGE) sequence.
Preprocessing and data analysis were conducted using SPM2 (Wellcome Department of Cognitive Neurology, London, UK). Functional images were slice-time corrected, realigned to the first image for motion correction, normalized to the Montreal Neurological Institute template, resampled to 2mm cubic voxels, and spatially smoothed using a 6mm full-width half maximum isotropic Gaussian kernel.
In an event-related analysis, encoding events were sorted based on the participant’s subsequent response on the later memory tests. Each event was coordinated with a canonical hemodynamic response function. For the recall task, encoded adjectives were either successfully remembered (i.e. accurately written down) or forgotten (i.e. inaccurately written or not written at all). Similarly for the recognition task, encoded adjectives were either remembered (i.e. a “yes” response) or forgotten (i.e. a “no” response). A separate model was made for the recall and recognition tasks. The two regressors used in each model were Remembered and Forgotten. In order to compare activity associated with successful encoding and failed encoding, we created two contrasts: Remembered – Forgotten and Forgotten – Remembered. Separate contrasts were made for recall and recognition data. Contrasts were smoothed using an 8-mm full-width half maximum isotropic Gaussian kernel, resulting in a total of 10 mm of smoothing across the two stages. Contrasts for young and older adults were compared using a random effects group-analysis for recall and recognition. Two-sample t-tests had a voxel threshold of p<.001 (uncorrected) and an extent threshold of 5 voxels. To assess regions of activity in common for young and older adults, we created a mask of voxels significant at p<.05 for young adults to test for overlapping activations in the older adults for a conjoint probability of p<.0025 using Fisher’s method (Fisher, 1950; Lazar, Luna, Sweeney, & Eddy, 2002), with an extent threshold of 5 voxels. We selected regions of interest and used MarsBar software (Brett, 2002) to extract percent signal change. Extractions were taken from a sphere with a 6mm radius around the peak voxel.
5. Introduction to Experiment 2
To directly investigate the impact of thinking about others on the ways in which one remembers self-relevant information, we directly compared conditions in which younger and older participants only made judgments about the self (comparable to the present fMRI study) to a condition in which younger and older alternated between judgments about the self and other. We included additional conditions that enhanced the relativity with which one made judgments of the self, including making judgments “relative to others” vs. “in general” and comparing the effect across adjectives that were strongly, moderately, or not descriptive of the self. Cross-cultural work has employed relative versus general judgments of the self in order to demonstrate that these decisions can engage distinct processes, which are associated with one’s self construal (e.g., independent vs. interdependent) (Chiao et al., 2009). With age, individuals become more motivated by close personal relationships (Carstensen, Isaacowitz, & Charles, 1999) and may become more oriented to interpersonal, social contexts (Grossmann et al., 2012; Grossmann et al., 2010). Thus, we predicted that older adults’ memory performance would be more affected than young adults’ when they think about the self in a more relative manner. In the present paradigm, that would mean that their memory would be more impacted by the extent to which they thought about others in the task (i.e., by making judgments of another person, and/or thinking about the self relative to others, rather than in an absolute and isolated manner). These effects may occur across levels of self-descriptiveness, or it is possible that they only emerge for words that are neither strongly associated nor dissociated with the self (i.e., words rated at a middle level of self-descriptiveness, rather than strongly descriptive or not descriptive of the self).
6. Results
Recognition accuracy was compared in response to the Self prompts (both general and relative), as these were the conditions completed by all participants. We employed hit rates, as there was only one pool of lure items so that condition-specific false alarm rates did not exist. Hit rates were compared based on the level of self-descriptiveness judged at encoding, by dividing participants’ ratings of how often the word described them into thirds (low, middle, and high).
We conducted a 2x2x2x2 mixed ANOVA on hit rates with Judgment Type (general/relative) and Rating of Self-Descriptiveness (low/middle/high) as within-participant variables and Judgment of Other (self only/self & other) and Age as between-participants variables. Of particular interest based on our interpretation of the results of Experiment 1, there was a significant interaction of Judgment Type x Rating of Self-Descriptiveness x Judgment of Other x Age, F(2, 118) = 3.74, p=.027, η2=.06. See Figure 4. To further understand this interaction, we conducted follow-up ANOVAs separately for each age group. Whereas young adults performed rather consistently across all conditions, with no significant interactions (p=.65 for critical three-way interaction), older adults’ performance differed across conditions, with a significant Judgment Type x Rating of Self-Descriptiveness x Judgment of Others interaction, F(2, 56) = 6.01, p=.004, η2=.18. This interaction was driven by words in the middle rating level for which there was a significant interaction of Judgment Type x Judgment of Others, F(1, 30) = 9.56, p=.004, η2=.24, in a follow-up ANOVA. Whereas older adults in the Self Only condition tended to perform more poorly for words judged in the Relative condition (M = .61, SD = .24) compared to the General condition (M = .72, SD = .20), t(15) = 1.75, p=.10, the opposite was true for older adults in the Self & Other condition. For these participants, words judged in the General condition (M = .54, SD = .26) led to poorer performance than for those judged in the Relative condition (M = .69, SD = .28), t (15) = 2.72, p=.016.
Figure 4.

Behavioral data from Experiment 2. While performance is relatively flat across conditions for younger adults, task context impacts older adults’ performance.
In addition to the primary interaction of interest, additional effects reached significance. There was a significant main effect of age, F(1, 59) = 38.28, p<.001, η2=.39, with younger adults exhibiting higher hit rates than older adults. There was also a main effect of rating, F(2, 118) = 13.90, p<.001, η2=.19, such that hit rates tended to increase with self-descriptiveness, particularly between medium and high levels. There were also significant two-way interactions of Judgment Type x Judgment of Others, F(1, 59) = 5.25, p=.03, η2=.08, Rating x Age, F(2, 118) = 5.28, p=.01, η2=.08, and Judgment Type x Rating, F(2, 118) = 3.24, p=.04, η2=.05. Finally, there was a significant three-way interaction of Judgment Type x Rating x Judgment of Others, F(2, 118) = 6.68, p=.002, η2=.10. Because these effects are qualified by the significant four-way interaction presented above, we do not discuss them further.
7. Discussion
Experiment 2 provides evidence that older adults are more affected than young adults by the social context in which they make judgments, and that this impacts the effectiveness of their memory processes. Young adults’ memory performance is relatively unaffected by the different conditions under which they make judgments about the self. In contrast, older adults performance varies based on the social context of the task in which they make judgments of the self. When the social context of the self judgment is enhanced by alternating between judgments of others and the self, words judged as moderately self-descriptive, relative to others, are better remembered than when one only makes judgments of the self. Therefore, when items are deemed to be somewhat descriptive of the self, judged under the most relative conditions, this enhances memory. This is striking because overall, alternating between judgments of self and other tends to impair older adults’ memory compared to when the task consisted of only self-relevance judgments. This enhancement is particularly pronounced when compared to when older adults endorse items as somewhat descriptive of the self (middle level) in the context of making general judgments of the self. Under these conditions, alternating between self and other judgments impairs memory. While our follow-up analyses focus on comparing the middle level of ratings, it is worth noting that alternating between self and other also appears to disproportionately impair memory for items rated as not self-descriptive by older adults, compared to when they only make self judgments. It may be the case that endorsing an item as somewhat descriptive of the self requires overcoming conflict from thinking about others, at the task and item levels. This additional effortful processing could then result in enhanced memory for older adults. If young adults are less attuned to others and reflect on the self as an independent entity, they should experience less interference across conditions. This would lead to less distinction in the processes engaged across conditions for younger adults, consistent with their relatively stable level of performance.
8. Methods
8.1 Participants
A final sample of 33 young (M age = 20.58, SD = 1.26) and 30 older (M age = 77.06, SD = 5.73) adults participated in the study. An additional 9 participants were excluded from data analysis because they did not have ratings in all levels for all conditions. Younger adults (M = 79.06, SD = 10.73) had higher digit comparison scores than older adults (M age = 57.70, SD = 13.41), t(62) = 7.01, p<.001. Older adults (M = 35.72, SD = 3.38) had higher Shipley vocabulary scores than younger adults (M = 31.22, SD = 2.66), t(62) = 5.92, p<.001.
8.2 Materials and procedure
Participants rated how often a word (e.g., admirable) described them, using a 9-point scale (1=not at all, 5 = sometimes, 9=always). Participants incidentally encoded 144 words. Each word was presented for 5 seconds. Half of the young and half of the older participants made judgments only about the self. For these, half (72) of the words were judged in response to the prompt, “in general I am” and half (72) were decided in response to the prompt, “relative to others I am”. The other half of the participants alternated judgments about the self with judgments about another person (Albert Einstein). For these participants, they judged 36 words in each of the four conditions (self/other x general/relative). Judgment types (general/relative, as well as self/other for participants making both types of judgments) were clustered into mini-blocks consisting of six trials of the same judgment type, with a fixation cross before the next block of trials. Words were counterbalanced across four different lists in order to alternate whether each word was assigned to the general or relative condition, and the self or other judgment.
After a 10 minute delay, participants received the surprise recognition task, which was self-paced. Participants made a keypress to indicate their judgment of old or new for each of the 144 previously studied words as well as 72 new words.
9. General
Discussion Using measures of recall and recognition, we present evidence that medial prefrontal cortex can support successful self-referential encoding for both younger and older adults. In contrast to prior work that employed both self and other person judgments, minimal age differences emerged when participants made only judgments of the self. We suggest that thinking about others induces differences across the age groups in terms of how one reflects upon the self. As we suggested for our prior fMRI study (Gutchess et al., 2010), age groups may differ in the qualitative ways that they approach self-referencing. Older adults may be prone to consider others when thinking about the self, perhaps reflecting on the self in a normative or relative manner, and this can impact the engagement of neural regions that support encoding of self-referential information. While in these studies we did not directly contrast neural regions engaged across age groups when thinking about the self in a general or absolute manner versus a relative manner, the differences in our neural results across fMRI studies and across conditions directly compared in the behavioral study is consistent with this interpretation. Future work may benefit from directly contrasting neural engagement across age groups under different social contexts, as in our work comparing neural engagement under stereotype threat versus resilient conditions (Colton, Leshikar, & Gutchess, 2013). In addition, substantial work is needed with behavioral methods to better characterize the ways in which age groups think about the self in terms of distinctly different qualities, spontaneously and as a result of different task demands and contexts.
Highlights.
Self-referencing engages largely similar neural regions in young and older adults
When participants make only self judgments, findings diverge from prior work
Older adults encode self-relevant information relative to others more than young
Acknowledgments
We gratefully acknowledge experimental assistance from Allie Indeck, Maya Siegel, Christine Hosey, Deborah Chamama, and Brian Dahlben. This research was funded by National Institute on Aging (Grant R21AG032382), and portions of the research were conducted while A.H.G. was a fellow of the American Federation for Aging Research. The Athinoula A. Martinos Center for Biomedical Imaging is supported by the National Center for Research Resources (Grant P41 880 RR14075) and by the MIND Institute.
Footnotes
Note that Colton et al. (2013) includes data from these participants in a distinct set of analyses.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Beer JS, Lombardo MV, Bhanji JP. Roles of medial prefrontal cortex and orbitofrontal cortex in self-evaluation. J Cogn Neurosci. 2010;22(9):2108–2119. doi: 10.1162/jocn.2009.21359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brett M, Anton J-L, Valabregue R, Poline J-B. Region of interest analysis using an SPM toolbox [abstract]. Paper presented at the 8th International Conference on Functional Mapping of the Human Brain; Sendai, Japan. 2002. [Google Scholar]
- Buckner RL, Andrews-Hanna JR, Schacter DL. The brain's default network: anatomy, function, and relevance to disease. Ann N Y Acad Sci. 2008;1124:1–38. doi: 10.1196/annals.1440.011. [DOI] [PubMed] [Google Scholar]
- Carstensen L, Isaacowitz D, Charles S. Taking time seriously: a theory of socioemotional selectivity. American Psychologist. 1999;54(3):165–181. doi: 10.1037//0003-066x.54.3.165. [DOI] [PubMed] [Google Scholar]
- Chiao JY, Harada T, Komeda H, Li Z, Mano Y, Saito D, Iidaka T. Neural basis of individualistic and collectivistic views of self. Human Brain Mapping. 2009;30(9):2813–2820. doi: 10.1002/hbm.20707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colton G, Leshikar ED, Gutchess AH. Age differences in neural response to stereotype threat and resiliency for self-referenced information. Frontiers in Human Neuroscience. 2013;7 doi: 10.3389/fnhum.2013.00537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dolcos F, LaBar KS, Cabeza R. Dissociable effects of arousal and valence on prefrontal activity indexing emotional evaluation and subsequent memory: an event-related fMRI study. Neuroimage. 2004;23(1):64–74. doi: 10.1016/j.neuroimage.2004.05.015. S1053811904002940 [pii] [DOI] [PubMed] [Google Scholar]
- Dulas MR, Newsome RN, Duarte A. The effects of aging on ERP correlates of source memory retrieval for self-referential information. Brain Research. 2011;1377:84–100. doi: 10.1016/j.brainres.2010.12.087. [DOI] [PubMed] [Google Scholar]
- Fisher RA. Statistical methods for research workers. 11. Vol. 5. New York: Hafner; 1950. [Google Scholar]
- Folstein M, Folstein S, McHugh P. Mini-mental state: A practical method for grading the cognitive state of patients for the clinician. Journal of Psychiatric Research. 1975;12:189–198. doi: 10.1016/0022-3956(75)90026-6. [DOI] [PubMed] [Google Scholar]
- Glisky E, Marquine M. Semantic and self-referential processing of positive and negative adjectives in older adults. Memory. 2009;17(2):144–157. doi: 10.1080/09658210802077405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grossmann I, Karasawa M, Izumi S, Na J, Varnum MEW, Kitayama S, Nisbett RE. Aging and wisdom: Culture matters. Psychological Science. 2012;23(10):1059–1066. doi: 10.1177/0956797612446025. [DOI] [PubMed] [Google Scholar]
- Grossmann I, Na JY, Varnum MEW, Park DC, Kitayama S, Nisbett RE. Reasoning about social conflicts improves into old age. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(16):7246–7250. doi: 10.1073/pnas.1001715107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gusnard DA, Akbudak E, Shulman GL, Raichle ME. Medial prefrontal cortex and self-referential mental activity: relation to a default mode of brain function. Proc Natl Acad Sci U S A. 2001;98(7):4259–4264. doi: 10.1073/pnas.071043098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutchess AH, Kensinger EA, Schacter DL. Aging, self-referencing, and medial prefrontal cortex. Soc Neurosci. 2007;2(2):117–133. doi: 10.1080/17470910701399029. 778890605 [pii] [DOI] [PubMed] [Google Scholar]
- Gutchess AH, Kensinger EA, Schacter DL. Functional neuroimaging of self-referential encoding with age. Neuropsychologia. 2010;48:211–219. doi: 10.1016/j.neuropsychologia.2009.09.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutchess AH, Kensinger EA, Yoon C, Schacter DL. Ageing and the self-reference effect in memory. Memory. 2007;15(8):822–837. doi: 10.1080/09658210701701394. 783624081 [pii] [DOI] [PubMed] [Google Scholar]
- Hamami A, Serbun S, Gutchess A. Self-referencing processing and memory specificity with age. Psychology and Aging. 2011;26:636–646. doi: 10.1037/a0022626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedden T, Park D, Nisbett R, Ji L, Jing Q, Jiao S. Cultural variation in verbal versus spatial neuropsychological function across the lifespan. Neuropsychology. 2002;16:65–73. doi: 10.1037//0894-4105.16.1.65. [DOI] [PubMed] [Google Scholar]
- Jenkins AC, Mitchell JP. Medial prefrontal cortex subserves diverse forms of self-reflection. Soc Neurosci. 2011;6(3):211–218. doi: 10.1080/17470919.2010.507948. [DOI] [PubMed] [Google Scholar]
- Kelley WM, Macrae CN, Wyland CL, Caglar S, Inati S, Heatherton TF. Finding the self? An event-related fMRI study. Journal of Cognitive Neuroscience. 2002;14(5):785–794. doi: 10.1162/08989290260138672. [DOI] [PubMed] [Google Scholar]
- Kensinger EA, Corkin S. Two routes to emotional memory: Distinct neural processes for valence and arousal. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(9):3310–3315. doi: 10.1073/pnas.0306408101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lazar NA, Luna B, Sweeney JA, Eddy WF. Combining brains: a survey of methods for statistical pooling of information. Neuroimage. 2002;16(2):538–550. doi: 10.1006/nimg.2002.1107. [DOI] [PubMed] [Google Scholar]
- Leshikar ED, Duarte A. Medial prefrontal cortex supports source memory for self-referenced materials in young and older adults. Cognitive, Affective & Behavioral Neuroscience. 2013 doi: 10.3758/s13415-013-0198-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macrae CN, Moran JM, Heatherton TF, Banfield JF, Kelley WM. Medial prefrontal activity predicts memory for self. Cerebral Cortex. 2004;14(6):647–654. doi: 10.1093/cercor/bhh025. [DOI] [PubMed] [Google Scholar]
- Mitchell JP, Macrae CN, Banaji MR. Dissociable medial prefrontal contributions to judgments of similar and dissimilar others. Neuron. 2006;50(4):655–663. doi: 10.1016/j.neuron.2006.03.040. [DOI] [PubMed] [Google Scholar]
- Moran JM, Heatherton TF, Kelley WM. Modulation of cortical midline structures by implicit and explicit self-relevance evaluation. Soc Neurosci. 2009;4(3):197–211. doi: 10.1080/17470910802250519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller JH, Wonderlich S, Dugan K. Self-referent processing of age-specific material. Psychol Aging. 1986;1(4):293–299. doi: 10.1037/0882-7974.1.4.293. [DOI] [PubMed] [Google Scholar]
- Murty VP, Ritchey M, Adcock RA, LaBar KS. fMRI studies of successful emotional memory encoding: A quantitative meta-analysis. Neuropsychologia. 2010;48(12):3459–3469. doi: 10.1016/j.neuropsychologia.2010.07.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Northoff G, Heinzel A, de Greck M, Bermpohl F, Dobrowolny H, Panksepp J. Self-referential processing in our brain--a meta-analysis of imaging studies of the self. Neuroimage. 2006;31(1):440–457. doi: 10.1016/j.neuroimage.2005.12.002. [DOI] [PubMed] [Google Scholar]
- Paller KA, Wagner AD. Observing the transformation of experience into memory. Trends in Cognitive Sciences. 2002;6(2):93–102. doi: 10.1016/s1364-6613(00)01845-3. [DOI] [PubMed] [Google Scholar]
- Rogers T, Kuiper N, Kirker W. Self-reference and the encoding of personal information. Journal of Personality and Social Psychology. 1977;35(9):677–688. doi: 10.1037//0022-3514.35.9.677. [DOI] [PubMed] [Google Scholar]
- Rosa N, Gutchess A. Source memory for action in young and older adults: Self vs. close or unknown others. Psychology and Aging. 2011;26:625–630. doi: 10.1037/a0022827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shipley W. Shipley Institute of Living Scale. Los Angeles: Western Psychological Services; 1986. [Google Scholar]
- Symons CS, Johnson BT. The self-reference effect in memory: A meta-analysis. Psychological Bulletin. 1997;121(3):371–394. doi: 10.1037/0033-2909.121.3.371. [DOI] [PubMed] [Google Scholar]
- Turk DJ, van Bussel K, Waiter GD, Macrae CN. Mine and me: exploring the neural basis of object ownership. J Cogn Neurosci. 2011;23(11):3657–3668. doi: 10.1162/jocn_a_00042. [DOI] [PubMed] [Google Scholar]
- Yang L, Truong L, Fuss S, Bislimovic S. The effects of aging and divided attention on the self-reference effect in emotional memory: Spontaneous or effortful mnemonic benefits? Memory. 2012;20:596–607. doi: 10.1080/09658211.2012.690040. [DOI] [PubMed] [Google Scholar]