

Abstract
The genera Actinobacillus, Haemophilus, and Pasteurella exhibit extensive polyphyletic branching in phylogenetic trees and do not represent coherent clusters of species. In this study, we have utilized molecular signatures identified through comparative genomic analyses in conjunction with genome based and multilocus sequence based phylogenetic analyses to clarify the phylogenetic and taxonomic boundary of these genera. We have identified large clusters of Actinobacillus, Haemophilus, and Pasteurella species which represent the “sensu stricto” members of these genera. We have identified 3, 7, and 6 conserved signature indels (CSIs), which are specifically shared by sensu stricto members of Actinobacillus, Haemophilus, and Pasteurella, respectively. We have also identified two different sets of CSIs that are unique characteristics of the pathogen containing genera Aggregatibacter and Mannheimia, respectively. It is now possible to demarcate the genera Actinobacillus sensu stricto, Haemophilus sensu stricto, and Pasteurella sensu stricto on the basis of discrete molecular signatures. The other members of the genera Actinobacillus, Haemophilus, and Pasteurella that do not fall within the “sensu stricto” clades and do not contain these molecular signatures should be reclassified as other genera. The CSIs identified here also provide useful diagnostic targets for the identification of current and novel members of the indicated genera.
1. Introduction
The family Pasteurellaceae, the single constituent family of the order Pasteurellales, represents a diverse group of commensal and pathogenic bacteria within the class Gammaproteobacteria. The family currently contains 19 genera, some of which are particularly important human and animal pathogens [1, 2]. The genera Haemophilus contains species responsible for human bacteremia, pneumonia, acute bacterial meningitis, and the sexually transmitted disease chancroid [3–5]; Aggregatibacter species have been implicated in juvenile periodontitis [6]; members of the genera Mannheimia, Pasteurella, and Actinobacillus have been implicated in the causation of shipping fever in cattle, fowl cholera, and pleuropneumonia in pigs, respectively [7–9].
The family Pasteurellaceae was originally proposed as a higher level taxonomic grouping of the related pathogenic genera Actinobacillus, Haemophilus, and Pasteurella [10]. Classification of organisms into these three genera was primarily based on DNA G-C content and a handful of phenotypic traits [11]. The phenotypic traits were later found not to be characteristic of any single genus [12]. Consequently, the genera Actinobacillus, Haemophilus, and Pasteurella each exhibit extensive polyphyly in subsequent 16S rRNA based phylogenies [12, 13]. Additional studies based on individual or concatenated gene sets and DNA-DNA/rRNA-DNA hybridization also support the presence of extensive polyphyly within the genera Actinobacillus, Haemophilus, and Pasteurella [14–20].
Extensive work has been undertaken to amend the classification of the genera Actinobacillus, Haemophilus, and Pasteurella [1, 5, 9, 18]. New genera have been created to house phylogenetically coherent clusters of Actinobacillus, Haemophilus, and Pasteurella. The species [Actinobacillus] actinomycetemcomitans, [Haemophilus] aphrophilus, [Haemophilus] paraphrophilus, and [Haemophilus] segnis have been transferred to the genus Aggregatibacter [21]; the species [Haemophilus] paragallinarum, [Pasteurella] gallinarum, [Pasteurella] avium, and [Pasteurella] volantium have been transferred to the genus Avibacterium [22]; the species [Haemophilus] somnus and [Haemophilus] agni have been transferred to the genus Histophilus [23]; and the species [Pasteurella] haemolytica and [Pasteurella] granulomatis have been transferred to the genus Mannheimia [7]. Additionally, some individual species within the genera Actinobacillus, Haemophilus, and Pasteurella that do not cluster with other members of their genus in phylogenetic trees have been moved or proposed to be moved to novel or neighbouring genera (namely, the transfer of the species [Haemophilus] pleuropneumoniae to the genus Actinobacillus [24], the transfer of the species [Pasteurella] anatis to the genus Gallibacterium [25], the transfer of the species [Pasteurella] trehalosi to the genus Bibersteinia [26], the transfer of the species [Pasteurella] ureae to the genus Actinobacillus [27], and the proposed transfer of the species [Haemophilus] ducreyi to a novel genus [28]). However, despite these changes, the classification of the genera Actinobacillus, Haemophilus, and Pasteurella is still problematic and each genus continues to contain members which exhibit polyphyletic branching [5, 17–20].
Multiple studies have attempted to define a core group of species which cluster around the nomenclatural type species of Actinobacillus, Haemophilus, or Pasteurella as the only true members of these genera (i.e., sensu stricto) [13, 15–17, 29–31], but the taxonomy and phylogeny of these bacteria continue to remain inconclusive [20, 32, 33]. Several methods have been employed for the demarcation of these genera; however, no simple method or criterion is available that can clearly delimit these genera. It has been suggested that genome based studies may provide reliable means of clarifying the evolutionary relationships of these bacteria [33].
Since the availability of the first complete genome sequence of the Haemophilus influenzae [34], a large number of genomes for the members of the family Pasteurellaceae have become available in public databases [35, 36]. The availability of these genomes provides us with an opportunity to complete comprehensive genome scale phylogenetic analyses of the family Pasteurellaceae. These genome sequences have also been utilized to carry out comparative genomic analyses to identify molecular signatures (namely, conserved signature indels (CSIs) in various proteins), commonly shared by all or closely related subsets of species within the family Pasteurellaceae. On the basis of the molecular signatures identified from comparative analyses of Pasteurellaceae genomes in conjunction with core genome based and multilocus sequence based phylogenetic analyses, we have identified sensu stricto clades of Actinobacillus, Haemophilus, and Pasteurella that are supported by 3, 7, and 6 unique molecular signatures, respectively. We also report sets of molecular signatures that are unique characteristics of the pathogen containing genera Aggregatibacter and Mannheimia.
2. Methods
2.1. Multilocus Sequence Analysis
Multilocus sequence analysis was completed for members of the family Pasteurellaceae using widely available nucleotide sequences of the 16S rDNA, infB (translation initiation factor IF-2), recN (DNA repair protein), and rpoB (DNA-directed RNA polymerase subunit beta) genes which have been used, individually or as part of a set, in a number of previous phylogenetic analyses of the family Pasteurellaceae [15–17, 29, 30]. Gene sequences for these four genes were obtained for 52 Pasteurellaceae strains, representing a large majority of the known Pasteurellaceae species, and 2 members of Vibrio cholerae from the NCBI nucleotide database [37]. Species which were missing one of these four genes or which did not have a gene sequence that was at least 50% of the length of the full gene were excluded from the analysis. The four genes were individually aligned using MUSCLE [38] and manually concatenated to create a combined dataset that contained 10 183 nucleotide long alignments. A maximum-likelihood tree based on 100 bootstrap replicates of this alignment was constructed using MEGA 6.0 [39] while employing maximum composite likelihood substitution model.
2.2. Pasteurellaceae Core Genome Phylogenetic Tree
A phylogenetic tree of 76 Pasteurellaceae strains, rooted using 7 members of the family Vibrionaceae, based on the core genome of the family Pasteurellaceae was created for this study. The core set of Pasteurellaceae proteins were identified using the UCLUST algorithm [55] to identify widely distributed protein families with at least 30% sequence identity and 50% sequence length. Proteins families which were present in less than 50% of the input genomes were excluded from further analysis. Potentially paralogous sequences (additional proteins from the same organism in a single protein family) within the remaining protein families were also excluded from further analysis. Each protein family was individually aligned using MAFFT 7 [56]. Aligned amino acid positions which contained gaps in more than 50% of organisms were excluded from further analysis. The remaining amino acid positions were concatenated to create a combined dataset that contained 128 080 amino acid long alignments. An approximately maximum-likelihood tree based on this alignment was constructed using FastTree 2 [57] while employing the Whelan and Goldman substitution model [58].
2.3. Identification of Molecular Signatures (CSIs) for Different Genera of the Family Pasteurellaceae
The detailed outline of the process of identifying CSIs has been recently published [59]. In brief, Blastp searches were performed on all proteins from the genome of Haemophilus influenzae F3047 [47]. Ten to fifteen high scoring homologues that were present in Haemophilus, other Pasteurellaceae, and Gammaproteobacteria species were retrieved, and their multiple sequence alignments were constructed using Clustal X 1.83 [60]. The alignments were visually inspected to identify any conserved inserts or deletions (indels) that are restricted to the particular clades of the family Pasteurellaceae, which are flanked on each side by at least 5-6 identical/conserved residues in the neighbouring 30–40 amino acids. The selected sequences containing the indels and their flanking conserved regions were further evaluated by detailed Blastp searches to determine species distribution and group specificity. The results of these Blast searches were processed using Sig_Create and Seq_Style to construct signature files [59]. Due to space constraints, the sequence alignment files presented here contain sequence information for a limited number of species within the order Pasteurellaceae and a representative selection of outgroup species. However, in each case, all members of the order and outgroups exhibited similar sequence characteristics to the representatives.
3. Results and Discussion
3.1. Phylogenetic Analysis of the Pasteurellaceae
Elucidating an accurate phylogeny of the members of the family Pasteurellaceae has been a long standing challenge in Pasteurellaceae research [10–12, 18, 19]. Early 16S rRNA based studies revealed that the established taxonomy of the family Pasteurellaceae was not consistent with their genetically inferred phylogeny [12, 14]. This has led to a long series of taxonomic revisions within the family Pasteurellaceae, a process which is still taking place today [7, 18, 22, 28]. However, it was subsequently discovered that phylogenetic trees of Pasteurellaceae species based on different genes did not completely agree with each other [15, 16, 31]. In particular, phylogenetic trees based on the 16S rRNA gene, often considered the gold standard in bacterial taxonomy and phylogeny [61, 62], disagreed with highly robust multilocus sequence and concatenated protein sequence based phylogenetic trees [9, 17, 19, 20, 53, 63].
Phylogenetic trees based on concatenated sequences for a large number of unlinked and conserved loci are more reliable and robust than phylogenetic trees based on any single gene or protein [64, 65]. Due to a rapid increase in the availability of genomic sequence data, we are now able to complete genome scale phylogenetic analyses of the family Pasteurellaceae which cover a vast majority of the diversity within the family. In this work we have produced a phylogenetic tree for 74 genome sequenced members of the family Pasteurellaceae based on 128 080 aligned amino acid positions (Figure 1(a)). The branching patterns of the core genome phylogenetic tree produced in this work largely agree with a previous genome based phylogenetic tree produced for a limited number of Pasteurellaceae species [19] and a concatenated protein based phylogenetic tree of the family Pasteurellaceae produced by our lab in a previous study [20]. Additionally, we have also produced a multilocus sequence based phylogenetic tree using the 16S rDNA, infB, recN, and rpoB genes which are commonly used in the phylogenetic analysis of the family Pasteurellaceae (Figure 1(b)) [15–17, 29, 30]. This tree also showed broadly similar branching patterns to past multilocus sequence based phylogenetic trees [17, 18] and to our core genome based phylogenetic tree. Both our core genome based and multilocus sequence based phylogenetic trees provide evidence for a division of the Pasteurellaceae into at least two higher taxonomic groups (families) which are broadly similar to the two clades of Pasteurellales identified in our previous work [20]. A similar division of the family Pasteurellaceae into two or more large groups is seen in many other robust multilocus or concatenated protein based phylogenetic trees [17, 19, 53, 63]; however, this division is not readily apparent in phylogenies based on the 16S rRNA gene [9, 66].
Figure 1.

(a) A maximum-likelihood core genome phylogenetic tree of sequenced members of the family Pasteurellaceae. (b) A maximum-likelihood phylogenetic tree based on concatenated nucleotide sequence alignments of the 16S rDNA, infB, recN, and rpoB genes. Both trees are rooted using members of the Vibrionaceae (not shown). Nodes with >80% bootstrap support are indicated by diamond shaped symbols at the node. Clusters of species representing Actinobacillus sensu stricto, Haemophilus sensu stricto, Pasteurella sensu stricto, Aggregatibacter, and Mannheimia are indicated by brackets. Members of the genera Actinobacillus, Haemophilus, and Pasteurella which do not fall into their respective “sensu stricto” clades are indicated by the presence of square brackets around their generic name (ex. [Pasteurella] pneumotropica).
A majority of the known genera within the family Pasteurellaceae form well-defined and coherent clusters in phylogenetic trees (Figure 1) [9, 17, 19, 20, 66]. The genera Actinobacillus, Haemophilus, and Pasteurella, which were described before the advent of genetic characterization, exhibit polyphyletic branching in all gene and protein based phylogenetic trees, including the core genome based and multilocus sequence based phylogenetic trees created in this work (Figure 1). However, there are large clusters of Actinobacillus, Haemophilus, and Pasteurella species identifiable in the phylogenetic trees which represent the core or “sensu stricto” members of each genera. The clusters of species that represent Actinobacillus sensu stricto, Haemophilus sensu stricto, and Pasteurella sensu stricto are indicated in Figure 1. Members of each genera which fall outside of the sensu stricto clusters, indicated in our phylogenetic trees by the presence of square brackets around their genus name (e.g., [Pasteurella] pneumotropica), are only distantly related to the sensu stricto members of their genus and will require reclassification in order to make their taxonomy and phylogeny concordant.
3.2. The Usefulness of Conserved Signature Indels as Phylogenetic and Taxonomic Markers
Whole genome sequences are a rich resource for the discovery of molecular signatures which are unique to a group of organisms [67–69]. One useful class of shared molecular signatures are conserved signature indels (CSIs), which are insertions/deletions uniquely present in protein sequences from a group of evolutionarily related organisms [59, 70, 71]. The unique, shared presence of multiple CSIs by a group of related species is most parsimoniously explained by the occurrence of the genetic changes that resulted in these CSIs in a common ancestor of the group, followed by vertical transmission of these CSIs to various descendant species [59, 71–73]. Hence, these CSIs represent molecular synapomorphies (markers of common evolutionary decent) which can be used to identify and demarcate specific bacterial groups in molecular terms and for understanding their interrelationships independently of phylogenetic trees [59, 70–72]. CSIs have recently been used to propose important taxonomic changes for a number of bacterial groups (namely, Aquificae, Spirochaetes, Thermotogae, Xanthomonadales, and Borrelia) at different taxonomic ranks [69, 74–77]. In the present work, we have completed comprehensive comparative analysis of Pasteurellaceae genomes (Table 1) in order to identify CSIs that are primarily restricted to the different genera within the family Pasteurellaceae. We have identified 3, 7, and 6 unique molecular signatures which are shared by Actinobacillus sensu stricto, Haemophilus sensu stricto, and Pasteurella sensu stricto, respectively. Information regarding these CSIs and their evolutionary significances is discussed below.
Table 1.
Genome characteristics of the sequenced Pasteurellaceae included in our analyses.
| Organism name | BioProject | Size (Mb) |
Proteins | G-C (%) |
References |
|---|---|---|---|---|---|
| Actinobacillus pleuropneumoniae L20 | CP000569 | 2.27 | 2013 | 41.3 | [40] |
| Actinobacillus pleuropneumoniae serovar 3 str. JL03 | CP000687 | 2.24 | 2036 | 41.2 | [41] |
| Actinobacillus pleuropneumoniae serovar7 str. AP76 | CP001091 | 2.35 | 2142 | 41.2 | STHHb |
| Actinobacillus ureae ATCC 25976a | AEVG0 | 2.30 | 2475 | — | BCMg |
| Actinobacillus minor 202a | ACFT0 | 2.13 | 2050 | 39.3 | McGill University |
| Actinobacillus minor NM305a | ACQL0 | 2.43 | 2411 | 39.3 | McGill University |
| Actinobacillus succinogenes 130Z | CP000746 | 2.32 | 2079 | 44.9 | Joint Genome Institute |
|
| |||||
| Aggregatibacter aphrophilus NJ8700 | CP001607 | 2.31 | 2219 | 42.2 | [19, 42] |
| Aggregatibacter actinomycetemcomitans D11S-1 | CP001733 | 2.20 | 2280 | 44.3 | [43] |
| Aggregatibacter actinomycetemcomitans D7S-1 | CP003496 | 2.31 | 2250 | 44.3 | [44] |
| Aggregatibacter segnis ATCC 33393a | AEPS0 | 1.99 | 1956 | — | BCMg |
|
| |||||
| Gallibacterium anatis UMN179 | CP002667 | 2.69 | 2500 | 39.9 | [45] |
|
| |||||
| Haemophilus aegyptius ATCC 11116a | AFBC0 | 1.92 | 2020 | — | BCMg |
| Haemophilus ducreyi 35000HP | AE017143 | 1.70 | 1717 | 38.2 | Ohio State University |
| Haemophilus haemolyticus M21621a | AFQQ0 | 2.09 | 1894 | — | [46] |
| Haemophilus influenzae 10810 | FQ312006 | 1.98 | 1903 | 38.1 | WTSIh |
| Haemophilus influenzae F3031 | FQ670178 | 1.99 | 1770 | 38.2 | [47] |
| Haemophilus influenzae F3047 | FQ670204 | 2.01 | 1786 | 38.2 | [47] |
| Haemophilus influenzae 22.1-21a | AAZD0 | 1.89 | 2224 | 38.0 | [48] |
| Haemophilus influenzae 3655 | AAZF0 | 1.88 | 1929 | 38.0 | [48] |
| Haemophilus influenzae 6P18H1a | ABWW0 | 1.91 | 1893 | 38.2 | CGS, ASRIe |
| Haemophilus influenzae 7P49H1a | ABWV0 | 1.83 | 1752 | 37.9 | CGS, ASRIe |
| Haemophilus influenzae NT127a | ACSL0 | 1.87 | 1809 | 38.0 | BIGSPc |
| Haemophilus influenzae PittAAa | AAZG0 | 1.88 | 1981 | 38.1 | [48] |
| Haemophilus influenzae PittIIa | AAZI0 | 1.95 | 2028 | 38.0 | [48] |
| Haemophilus influenzae PittHHa | AAZH0 | 1.84 | 1977 | 38.0 | [48] |
| Haemophilus influenzae R3021a | AAZJ0 | 1.88 | 2307 | 37.9 | [48] |
| Haemophilus influenzae RdAWa | ACSM0 | 1.80 | 1718 | 38.0 | BIGSPc |
| Haemophilus influenzae 86-028NP | CP000057 | 1.91 | 1792 | 38.2 | [49] |
| Haemophilus influenzae PittEE | CP000671 | 1.81 | 1613 | 38.0 | [48] |
| Haemophilus influenzae PittGG | CP000672 | 1.89 | 1661 | 38.0 | [48] |
| Haemophilus influenzae Rd KW20 | L42023 | 1.83 | 1657 | 38.2 | [34] |
| Haemophilus influenzae R2846 | CP002276 | 1.82 | 1636 | 38.0 | UW-SBRId |
| Haemophilus influenzae R2866 | CP002277 | 1.93 | 1795 | 38.1 | UW-SBRId |
| Haemophilus parainfluenzae ATCC 33392 a | AEWU0 | 2.11 | 2010 | — | BCMg |
| Haemophilus parainfluenzae T3T1 | FQ312002 | 2.09 | 1975 | 39.6 | WTSIh |
| Haemophilus parasuis 29755a | ABKM0 | 2.22 | 2244 | 39.8 | Iowa State University |
| Haemophilus parasuis SH0165 | CP001321 | 2.27 | 2021 | 40.0 | [50] |
| Haemophilus pittmaniae HK 85a | AFUV0 | 2.18 | 2390 | — | J. Craig Venter Institute |
| Haemophilus sputorum CCUG13788a | AFNK0 | 2.14 | 2073 | — | Aarhus University Hospital |
| Haemophilus parahaemolyticus HK385a | AJSW0 | 1.81 | 1764 | — | J. Craig Venter Institute |
| Haemophilus paraphrohaemolyticus HK411a | AJMU0 | 2.02 | 2025 | — | J. Craig Venter Institute |
| Haemophilus sp. oral taxon 851 str.F0397a | AGRK0 | 1.84 | 1809 | — | GCG-WUf |
|
| |||||
| Histophilus somni 2336 | CP000947 | 2.26 | 1980 | 37.4 | Joint Genome Institute |
| Histophilus somni 129PT | CP000436 | 2.01 | 1798 | 37.2 | [51] |
|
| |||||
| Mannheimia succiniciproducens MBEL55E | AE016827 | 2.31 | 2370 | 42.5 | [52] |
| Mannheimia haemolytica PHL213a | AASA0 | 2.57 | 2695 | 41.1 | [53] |
|
| |||||
| Pasteurella multocida subsp. multocida str. Pm70 | AE004439 | 2.26 | 2012 | 40.4 | [54] |
| Pasteurella dagmatis ATCC 43325a | ACZR0 | 2.25 | 2053 | 37.4 | BCMg |
aThe genomes of these species/strains are currently under scaffolds/contigs status.
bStiftung Tieraerztliche Hochschule Hannover (STHH).
cThe Broad Institute Genome Sequencing Platform (BIGSP).
dUniversity of Washington; Seattle Biomedical Research Institute (UW-SBRI).
eCenter for Genomic Sciences, Allegheny-Singer Research Institute (CGS, ASRI).
fGenome Sequencing Center (GSC) at Washington University (WashU) School of Medicine.
gBaylor College of Medicine (BCM).
hWellcome Trust Sanger Institute (WTSI).
3.3. Molecular Signatures Specific for Actinobacillus sensu stricto
The genus Actinobacillus was originally defined as a group of growth factor independent host-associated rods which shared phenotypic or biochemical similarity with Actinobacillus lignieresii, the type species of the genus [24, 78]. However, the original classification scheme for the genus Actinobacillus led to the inclusion of a highly heterogeneous and polyphyletic grouping of species within the genus [12–14]. An assemblage of Actinobacillus species closely related to Actinobacillus lignieresii has been recognized as Actinobacillus sensu stricto (i.e., the core members of the genus Actinobacillus) in both our phylogenetic analysis (Figure 1) and past phylogenetic analyses [12–14, 17]. Differentiation of Actinobacillus sensu stricto from other Actinobacillus species and the modern criteria for placing novel species within the genus Actinobacillus sensu stricto is heavily reliant on genetic and genomic criteria, namely, DNA-DNA hybridization values, 16S rRNA sequence similarity, and other single gene sequence comparisons [13, 18]. There are currently no known discrete characteristics which are unique to Actinobacillus that define the genus. In this work, we have completed a comprehensive comparative analysis of Pasteurellaceae genomes in order to identify unique, defining molecular signatures for different genera within the family Pasteurellaceae. We have identified 3 CSIs which are unique, defining molecular signatures for the sequenced members of Actinobacillus sensu stricto (namely, Actinobacillus capsulatus, A. pleuropneumoniae, A. suis, and A. ureae). An example of a CSI specific for Actinobacillus sensu stricto is shown in Figure 2. The CSI consists of a 1-amino-acid insertion in a conserved region of a 3′-nucleotidase which is present in all sequenced members of Actinobacillus sensu stricto and absent in all other sequenced Gammaproteobacteria. Sequence information for 2 other CSIs which are also unique characteristics of the Actinobacillus sensu stricto clade is presented in Supplemental Figures 1-2 available online at http://dx.doi.org/10.1155/2015/198560 and their characteristics are briefly summarized in Table 2(A).
Figure 2.

A partial sequence alignment of a 3′-nucleotidase showing a 1-amino-acid insertion identified in all members of Actinobacillus sensu stricto. This insertion was not found in the homologues from any member of the genus Actinobacillus that was not part of the “sensu stricto” clade or any other member of the Gammaproteobacteria. Sequence information for a representative subset of the family Pasteurellaceae and the class Gammaproteobacteria is shown, but unless otherwise indicated, similar CSIs were detected in all members of the indicated group and not detected in any other bacterial species in the top 250 BLAST hits. The dashes (-) in the alignments indicate identity with the residue in the top sequence. GenBank identification (GI) numbers for each sequence are indicated in the second column. Sequence information for other CSIs specific to Actinobacillus sensu stricto are presented in Supplemental Figures 1-2 and their characteristics are summarized in Table 2(A).
Table 2.
Conserved signature indels specific for genera within the family Pasteurellaceae.
| Protein name | Gene name | GenBank identifier | Figure number | Indel size | Indel positiona |
|---|---|---|---|---|---|
| (A) CSIs specific for Actinobacillus sensu stricto | |||||
| 3′-nucleotidase | surE | 126208128 | Figure 2 | 1 aa ins | 367–402 |
| GTP pyrophosphokinase | relA | 126207889 | Sup. Figure 1 | 1 aa ins | 368–412 |
| Anaerobic glycerol-3-phosphate dehydrogenase subunit | glpA | 491834528 | Sup. Figure 2 | 1 aa ins | 359–400 |
|
| |||||
| (B) CSIs specific for Haemophilus sensu stricto | |||||
| Biotin-protein ligase | birA | 144979005 | Figure 3 | 6 aa del | 138–178 |
| Aspartate ammonia-lyase | aspA | 145630289 | Sup. Figure 3 | 1 aa ins | 34–75 |
| NAD(P) transhydrogenase subunit alpha | pntA | 145631394 | Sup. Figure 4 | 1 aa del | 352–378 |
| Fumarate reductase subunit C | frdC | 301169552 | Sup. Figure 5 | 3 aa ins | 31–89 |
| Hypothetical tRNA/rRNA methyltransferase | — | 145636352 | Sup. Figure 6 | 1 aa del | 17–58 |
| Gamma-glutamyl kinase | proB | 145629980 | Sup. Figure 7 | 1 aa ins | 197–253 |
| ACP phosphodiesterase | acpD | 68250119 | Sup. Figure 8 | 2 aa del | 119–159 |
|
| |||||
| (C) CSIs specific for Pasteurella sensu stricto | |||||
| Menaquinone-specific isochorismate synthase | menF | 386834899 | Figure 4 | 4 aa ins | 29–86 |
| tRNA s(4)U8 sulfurtransferase | thiI | 15602400 | Sup. Figure 9 | 2 aa del | 412–446 |
| FKBP-type peptidyl-prolyl cis-trans isomerase | slyD | 378775595 | Sup. Figure 10 | 2 aa del | 151–188 |
| Aspartate-semialdehyde dehydrogenase | asd | 383311492 | Sup. Figure 11 | 1 aa del | 173–245 |
| Lactate permease family transporter | lldP | 492154065 | Sup. Figure 12 | 2 aa ins | 390–427 |
| Cell division protein ftsA | ftsA | 492155843 | Sup. Figure 13 | 1 aa ins | 357–387 |
|
| |||||
| (D) CSIs specific for Aggregatibacter | |||||
| nhaC family sodium:proton antiporter | nhaC | 493769836 | Figure 5(a) | 3 aa ins | 396–437 |
| Outer membrane protein | omp | 261866907 | Sup. Figure 14 | 4 aa del | 25–64 |
| Multidrug transporter murJ | murJ | 365966332 | Sup. Figure 15 | 1 aa del | 190–220 |
| NADH dehydrogenase | nuoE | 387120244 | Sup. Figure 16 | 1 aa ins | 372–412 |
|
| |||||
| (E) CSIs specific for Mannheimia | |||||
| Methyl-galactoside ABC transporter substrate-binding protein | — | 472335016 | Figure 5(b) | 1 aa del | 33–73 |
| UDP-N-acetylmuramoylalanyl-D-glutamate–2,6-diaminopimelate ligase | murE | 472333011 | Sup. Figure 17 | 2 aa del | 418–473 |
| Glutathione-regulated potassium-efflux protein | kefC | 472333189 | Sup. Figure 18 | 1 aa ins | 504–531 |
| Glycerol-3-phosphate acyltransferase | plsB | 472334521 | Sup. Figure 19 | 2 aa del | 214–252 |
3.4. Molecular Signatures Specific for Haemophilus sensu stricto
The classification of novel species into the genus Haemophilus was initially based on phenotypic and biochemical properties, most importantly, the dependence of growth on the presence of factor V and factor X in blood [13, 78, 79]. As with Actinobacillus, the classification of Haemophilus on the basis of phenotypic and biochemical properties has led to the genus containing an extremely heterogeneous group of species [12–14, 32]. Species from the genus Haemophilus have undergone a number of transfers and reclassifications [21–24, 28]. However, the genus remains highly polyphyletic (Figure 1) [17, 19, 28]. The core members of the genus Haemophilus (namely, Haemophilus sensu stricto) consist of Haemophilus influenzae, H. aegyptius, and H. haemolyticus based on 16S rRNA sequence analysis [12–14, 32]. However, phylogenetic analysis based on DNA-DNA hybridization and multilocus sequence analysis suggests that H. parainfluenzae and H. pittmaniae are also members of Haemophilus sensu stricto [30, 80]. Phylogenetic analysis of rpoB, infB, and concatenated gene sets also suggest that [Pasteurella] pneumotropica and related isolates are closely related to Haemophilus sensu stricto [15, 16].
Our comparative analysis of Pasteurellaceae genomes has led to the identification of 7 CSIs that are unique characteristics of Haemophilus sensu stricto which consists of Haemophilus influenzae, H. aegyptius, H. haemolyticus, H. parainfluenzae, H. pittmaniae, and [Pasteurella] pneumotropica (Figure 1). One example of a CSI specific for the members of Haemophilus sensu stricto, shown in Figure 3, consists of a 4-amino-acid deletion in a biotin-protein ligase which is uniquely found in homologs from Haemophilus sensu stricto and absent in all other sequenced Gammaproteobacteria. Sequence information for 6 additional CSIs which are also unique characteristics of Haemophilus sensu stricto is presented in Supplemental Figures 3–8 and their characteristics are briefly summarized in Table 2(B). These CSIs and our phylogenetic trees (Figure 1) suggest that Haemophilus influenzae, H. aegyptius, H. haemolyticus, H. parainfluenzae, H. pittmaniae, and [Pasteurella] pneumotropica share a close evolutionary relationship and should all be considered members of Haemophilus sensu stricto. Additionally, these results also suggest that [Pasteurella] pneumotropica is incorrectly classified as a member of the genus Pasteurella and should be reclassified as “Haemophilus pneumotropica.”
Figure 3.

A partial sequence alignment of 1,4-dihydroxy-2-naphthoate octaprenyltransferase showing a 2-amino-acid insertion identified in all members of Haemophilus sensu stricto. This insertion was not found in the homologues from any member of the genus Haemophilus that was not part of the “sensu stricto” clade or any other member of the Gammaproteobacteria. Sequence information for other CSIs specific to Haemophilus sensu stricto is presented in Supplemental Figures 3–8 and their characteristics are summarized in Table 2(B).
3.5. Molecular Signatures Specific for Pasteurella sensu stricto
The genus Pasteurella is highly heterogeneous and polyphyletic (Figure 1) [13]. Similar to the members of Actinobacillus, bacterial isolates were originally classified as members of the genus Pasteurella based on growth factor independent growth and phenotypic or biochemical similarity to Pasteurella multocida, the type species of the genus [78, 81]. The monophyletic clusters of Pasteurella species that branch with Pasteurella multocida are considered the core members of the genus (namely, Pasteurella sensu stricto) [9, 13, 16, 17]. Our comparative analysis of Pasteurellaceae genomes has led to the identification of 6 CSIs which are unique characteristics for the sequenced members of Pasteurella sensu stricto (namely, Pasteurella multocida and P. dagmatis). An example of a CSI uniquely found in the sequenced members of Pasteurella sensu stricto, consisting of a 4-amino-acid insertion in a conserved region of Menaquinone-specific isochorismate synthase, is shown in Figure 4. This CSI is only found in the sequenced members of Pasteurella sensu stricto and is absent from all other sequenced Gammaproteobacteria. Partial sequence alignments for 5 additional CSIs which are also unique characteristics of Pasteurella sensu stricto are presented in Supplemental Figures 9–13 and their characteristics are briefly summarized in Table 2(C).
Figure 4.

A partial sequence alignment of Menaquinone-specific isochorismate synthase showing a 4-amino-acid insertion identified in all members of Pasteurella sensu stricto. This insertion was not found in the homologues from any member of the genus Pasteurella that was not part of the “sensu stricto” clade or any other member of the Gammaproteobacteria. Sequence information for other CSIs specific to Pasteurella sensu stricto is presented in Supplemental Figures 9–13 and their characteristics are summarized in Table 2(C).
3.6. Molecular Signatures Specific for the Genera Aggregatibacter or Mannheimia
The genus Aggregatibacter was proposed as a novel taxonomic classification for a monophyletic cluster of Actinobacillus and Haemophilus species which branched distinctly from the “sensu stricto” members of their respective clades [21]. Similarly, the genus Mannheimia was proposed as a novel classification for the Pasteurella Haemolytica complex which did not branch with Pasteurella sensu stricto in phylogenetic trees [7]. Currently other than branching in phylogenetic trees or relatedness in DNA-DNA hybridization studies, the members of the genera Aggregatibacter or Mannheimia do not share any single unique or defining biochemical or molecular characteristic that can differentiate them from all other bacteria [5, 82].
In this study we have identified 4 CSIs that are unique molecular characteristics shared by all sequenced species of the genus Aggregatibacter and another 4 CSIs which are uniquely found in all sequenced members of the genus Mannheimia. Examples of CSIs specific to the sequenced members of Aggregatibacter and Mannheimia are shown in Figure 5. A partial sequence alignment of a nhaC family sodium:proton antiporter containing a 3-amino-acid insertion specific for all sequenced species of the genus Aggregatibacter is shown in Figure 5(a) and a partial sequence alignment of a methyl-galactoside ABC transporter substrate-binding protein containing a 1-amino-acid deletion specific for all sequenced species of the genus Mannheimia is shown in Figure 5(b). In each case, the identified CSIs were only found in the sequenced members of the genera Aggregatibacter or Mannheimia and were absent from all other sequenced Gammaproteobacteria. Partial sequence alignments additional CSIs specific for the genera Aggregatibacter or Mannheimia are provided in Supplemental Figures 14–19 and their characteristics are summarized in Tables 2(D)-2(E). These CSIs are the first discrete molecular characteristics which are unique for the genera Aggregatibacter and Mannheimia and support their observed monophyly in phylogenetic trees. Additionally, these CSIs could be useful targets for the development of PCR based diagnostic assays for the genera Aggregatibacter and Mannheimia which amplify the CSI containing DNA segment using the conserved flanking regions of the CSIs [83, 84].
Figure 5.

A partial sequence alignment of (a) a nhaC family sodium:proton antiporter containing a 3-amino-acid insertion specific for all sequenced species of the genus Aggregatibacter (b) a methyl-galactoside ABC transporter substrate-binding protein containing a 1-amino-acid deletion specific for all sequenced species of the genus Mannheimia. In each case, the identified CSIs were only found in the sequenced members of the genera Aggregatibacter or Mannheimia and were absent from all other sequenced Gammaproteobacteria. Sequence information for other CSIs specific to Aggregatibacter or Mannheimia is presented in Supplemental Figures 14–19 and their characteristics are summarized in Tables 2(D) and 2(E).
4. Conclusion
The genera Actinobacillus, Haemophilus, and Pasteurella, within the family Pasteurellaceae, are known to exhibit extensive polyphyletic branching. We have utilized molecular signatures and phylogenetic analyses to clarify the taxonomic boundary of these genera. We have been able to identify large clusters of Actinobacillus, Haemophilus, and Pasteurella species which represent the “sensu stricto” members of these genera. We have identified 3, 7, and 6 unique molecular signatures which are specifically shared by the members of the genera Actinobacillus sensu stricto, Haemophilus sensu stricto, and Pasteurella sensu stricto, respectively. The group specificity of the molecular signatures we have identified in this work is summarized in Figure 6 and their characteristics are briefly summarized in Table 2. Our comparative genomic analyses have not come across any CSIs that were unique characteristics of all sequenced members of the genera Actinobacillus, Haemophilus, or Pasteurella as currently defined, suggesting that the members of these genera that do not fall into the “sensu stricto” clusters should not be considered members of their respective genus.
Figure 6.
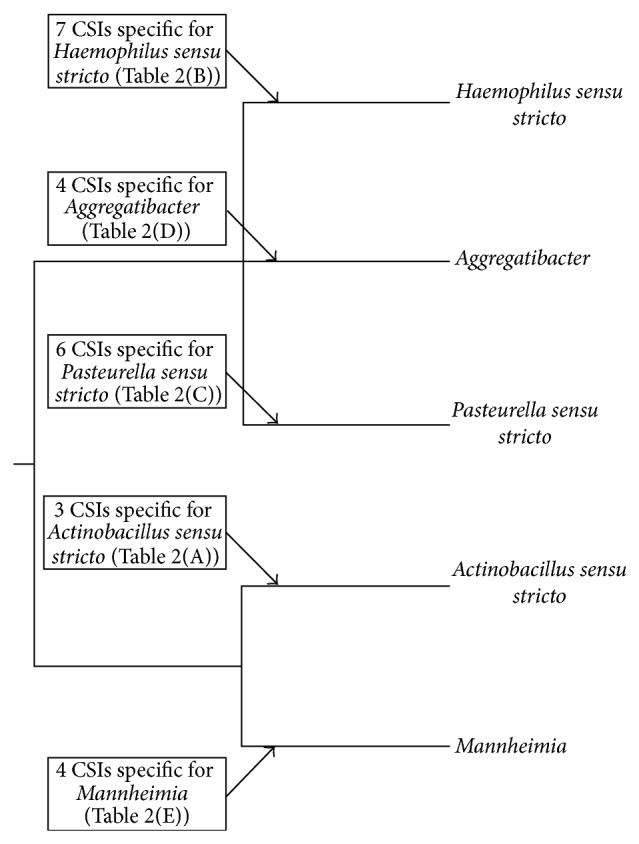
A summary diagram depicting the distribution of identified CSIs for genera within the family Pasteurellaceae.
Examinations of phenotypic and biochemical characteristics do not provide a reliable means of assigning a novel isolate to the genera Actinobacillus, Haemophilus, and Pasteurella [18]. However, based upon the CSIs described in this work, it is now possible to demarcate the genera Actinobacillus sensu stricto, Haemophilus sensu stricto, and Pasteurella sensu stricto on the basis of the presence or absence of unique molecular signatures. It is important to note that the current analysis of CSIs is limited to the currently available genomic sequence data and may show slight variance as additional bacterial genomes are sequenced. However, earlier work on CSIs for other groups of bacteria provides evidence that the identified CSIs have strong predictive value and will likely be found in other members of these groups as more species are sequenced and novel species are isolated [74, 77, 85, 86]. The conserved nature of the sequence regions that contain these CSIs, in conjunction with their strong predictive value, makes CSIs promising targets for the development of highly specific diagnostic assays for Actinobacillus sensu stricto, Haemophilus sensu stricto, Pasteurella sensu stricto, Aggregatibacter, and Mannheimia [83, 84]. Additionally, further analysis of these genus specific CSIs should lead to the discovery of their functional role in their respective organisms and may provide important insights into novel distinguishing features of these groups of organisms.
Supplementary Material
Partial sequence alignments of the CSI containing regions that were unique characteristics of the genera Actinobacillus sensu stricto, Haemophilus sensu stricto, Pasteurella sensu stricto, Aggregatibacter, or Mannheimia.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
Authors' Contribution
Sohail Naushad and Mobolaji Adeolu contributed equally to this work.
References
- 1.Parte A. C. LPSN—list of prokaryotic names with standing in nomenclature. Nucleic Acids Research. 2014;42(1):D613–D616. doi: 10.1093/nar/gkt1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Muehldorfer K., Speck S., Wibbelt G. Proposal of Vespertiliibacter pulmonis gen. nov., sp. nov. and two genomospecies as new members of the family Pasteurellaceae isolated from European bats. International Journal of Systematic and Evolutionary Microbiology. 2014;64(7):2424–2430. doi: 10.1099/ijs.0.062786-0. [DOI] [PubMed] [Google Scholar]
- 3.Spinola S. M., Bauer M. E., Munson R. S., Jr. Immunopathogenesis of Haemophilus ducreyi infection (chancroid) Infection and Immunity. 2002;70(4):1667–1676. doi: 10.1128/iai.70.4.1667-1676.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Christensen H., Bisgaard M. Molecular classification and its impact on diagnostics and understanding the phylogeny and epidemiology of selected members of Pasteurellaceoe of veterinary importance. Berliner und Münchener Tierärztliche Wochenschrift. 2010;123(1-2):20–30. [PubMed] [Google Scholar]
- 5.Nørskov-Lauritsen N. Classification, identification, and clinical significance of Haemophilus and Aggregatibacter species with host specificity for humans. Clinical Microbiology Reviews. 2014;27(2):214–240. doi: 10.1128/cmr.00103-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Henderson B., Ward J. M., Ready D. Aggregatibacter (Actinobacillus) actinomycetemcomitans: a triple A∗ periodontopathogen? Periodontology 2000. 2010;54(1):78–105. doi: 10.1111/j.1600-0757.2009.00331.x. [DOI] [PubMed] [Google Scholar]
- 7.Angen Ø., Mutters R., Caugant D. A., Olsen J. E., Bisgaard M. Taxonomic relationships of the [Pasteurella] haemolytica complex as evaluated by DNA-DNA hybridizations and 16s rRNA sequencing with proposal of Mannheimia haemolytica gen. nov., comb. nov., Mannheimia granulomatis comb. nov., Mannheimia glucosida sp. nov., Mannheimia ruminalis sp. nov. and Mannheimia varigena sp. nov. International Journal of Systematic Bacteriology. 1999;49(1):67–86. doi: 10.1099/00207713-49-1-67. [DOI] [PubMed] [Google Scholar]
- 8.Bossé J. T., Janson H., Sheehan B. J., et al. Actinobacillus pleuropneumoniae: pathobiology and pathogenesis of infection. Microbes and Infection. 2002;4(2):225–235. doi: 10.1016/s1286-4579(01)01534-9. [DOI] [PubMed] [Google Scholar]
- 9.Wilson B. A., Ho M. Pasteurella multocida: from zoonosis to cellular microbiology. Clinical Microbiology Reviews. 2013;26(3):631–655. doi: 10.1128/cmr.00024-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pohl S. Reklassifizierung der gattung Actinobacillus Brumpt 1910, Haemophilus Winslow et al. 1971 und Pasteurella Trevisan 1887 anhand phänotypischer und molekularer daten, insbesondere der DNS-verwandtschaften bei DNS: DNS-hybridisierung in vitro und vorschlag einer neuen familie, Pasteurellaceae [Inaug. Diss.] Marburg, Germany: Philipps-Universität; 1979. [Google Scholar]
- 11.Mannheim W., Pohl S., Holländer R. On the taxonomy of Actinobacillus, Haemophilus, and Pasteurella: DNA base composition, respiratory quinones, and biochemical reactions of representative collection cultures (author's transl) Zentralblatt für Bakteriologie A. 1979;246(4):512–540. [PubMed] [Google Scholar]
- 12.Dewhirst F. E., Paster B. J., Olsen I., Fraser G. J. Phylogeny of 54 representative strains of species in the family Pasteurellaceae as determined by comparison of 16S rRNA sequences. Journal of Bacteriology. 1992;174(6):2002–2013. doi: 10.1128/jb.174.6.2002-2013.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Olsen I., Dewhirst F. E., Paster B. J., Busse H. J. Family I. Pasteurellaceae . In: Brenner D. J., Krieg N. R., Garrity G. M., Staley J. T., editors. Bergey's Manual of Systematic Bacteriology. 2nd. Vol. 2. New York, NY, USA: Springer; 2005. pp. 851–856. [Google Scholar]
- 14.Dewhirst F. E., Paster B. J., Olsen I., Fraser G. J. Phylogeny of the Pasteurellaceae as determined by comparison of 16S ribosomal ribonucleic acid sequences. Zentralblatt für Bakteriologie. 1993;279(1):35–44. doi: 10.1016/s0934-8840(11)80489-5. [DOI] [PubMed] [Google Scholar]
- 15.Christensen H., Kuhnert P., Olsen J. E., Bisgaard M. Comparative phylogenies of the housekeeping genes atpD, infB and rpoB and the 16S rRNA gene within the Pasteurellaceae . International Journal of Systematic and Evolutionary Microbiology. 2004;54(5):1601–1609. doi: 10.1099/ijs.0.03018-0. [DOI] [PubMed] [Google Scholar]
- 16.Korczak B., Christensen H., Emler S., Frey J., Kuhnert P. Phylogeny of the family Pasteurellaceae based on rpoB sequences. International Journal of Systematic and Evolutionary Microbiology. 2004;54(4):1393–1399. doi: 10.1099/ijs.0.03043-0. [DOI] [PubMed] [Google Scholar]
- 17.Kuhnert P., Korczak B. M. Prediction of whole-genome DNA-DNA similarity, determination of G + C content and phylogenetic analysis within the family Pasteurellaceae by multilocus sequence analysis (MLSA) Microbiology. 2006;152(9):2537–2548. doi: 10.1099/mic.0.28991-0. [DOI] [PubMed] [Google Scholar]
- 18.Christensen H., Kuhnert P., Busse H. J., Frederiksen W. C., Bisgaard M. Proposed minimal standards for the description of genera, species and subspecies of the Pasteurellaceae. International Journal of Systematic and Evolutionary Microbiology. 2007;57(1):166–178. doi: 10.1099/ijs.0.64838-0. [DOI] [PubMed] [Google Scholar]
- 19.Bonaventura M. P. D., Lee E. K., DeSalle R., Planet P. J. A whole-genome phylogeny of the family Pasteurellaceae . Molecular Phylogenetics and Evolution. 2010;54(3):950–956. doi: 10.1016/j.ympev.2009.08.010. [DOI] [PubMed] [Google Scholar]
- 20.Naushad H. S., Gupta R. S. Molecular signatures (conserved indels) in protein sequences that are specific for the order Pasteurellales and distinguish two of its main clades. Antonie van Leeuwenhoek. 2012;101(1):105–124. doi: 10.1007/s10482-011-9628-4. [DOI] [PubMed] [Google Scholar]
- 21.Nørskov-Lauritsen N., Kilian M. Reclassification of Actinobacillus actinomycetemcomitans, Haemophilus aphrophilus, Haemophilus paraphrophilus and Haemophilus segnis as Aggregatibacter actinomycetemcomitans gen. nov., comb. nov., Aggregatibacter aphrophilus comb. nov. and Aggregatibacter segnis comb. nov., and emended description of Aggregatibacter aphrophilus to include V factor-dependent and V factor-independent isolates. International Journal of Systematic and Evolutionary Microbiology. 2006;56(9):2135–2146. doi: 10.1099/ijs.0.64207-0. [DOI] [PubMed] [Google Scholar]
- 22.Blackall P. J., Christensen H., Beckenham T., Blackall L. L., Bisgaard M. Reclassification of Pasteurella gallinarum, [Haemophilus] paragallinarum, Pasteurella avium and Pasteurella volantium as Avibacterium gallinarum gen. nov., comb. nov., Avibacterium paragallinarum comb. nov., Avibacterium avium comb. nov. and Avibacterium volantium comb. nov. International Journal of Systematic and Evolutionary Microbiology. 2005;55(1):353–362. doi: 10.1099/ijs.0.63357-0. [DOI] [PubMed] [Google Scholar]
- 23.Angen Ø., Ahrens P., Kuhnert P., Christensen H., Mutters R. Proposal of Histophilus somni gen. nov., sp. nov. for the three species incertae sedis ‘Haemophilus somnus’, ‘Haemophilus agni’ and ‘Histophilus ovis’. International Journal of Systematic and Evolutionary Microbiology. 2003;53(5):1449–1456. doi: 10.1099/ijs.0.02637-0. [DOI] [PubMed] [Google Scholar]
- 24.Pohl S., Bertschinger H. U., Frederiksen W., Mannheim W. Transfer of Haemophilus pleuropneumoniae and the Pasteurella haemolytica-like organism causing porcine necrotic pleuropneumonia to the genus Actinobacillus (Actinobacillus pleuropneumoniae comb. nov.) on the basis of phenotypic and deoxyribonucleic acid relatedness. International Journal of Systematic Bacteriology. 1983;33(3):510–514. doi: 10.1099/00207713-33-3-510. [DOI] [Google Scholar]
- 25.Christensen H., Bisgaard M., Bojesen A. M., Mutters R., Olsen J. E. Genetic relationship among avian isolates classified as Pasteurella haemolytica. 'Actinobacillus salpingitidis' or Pasteurella anatis with proposal of Gallibacterium anatis gen. nov., comb. nov. and description of additional genomospecies with Gallibacterium gen. nov. International Journal of Systematic and Evolutionary Microbiology. 2003;53(1):275–287. doi: 10.1099/ijs.0.02330-0. [DOI] [PubMed] [Google Scholar]
- 26.Blackall P. J., Bojesen A. M., Christensen H., Bisgaard M. Reclassification of [Pasteurella] trehalosi as Bibersteinia trehalosi gen. nov., comb. nov. International Journal of Systematic and Evolutionary Microbiology. 2007;57(4):666–674. doi: 10.1099/ijs.0.64521-0. [DOI] [PubMed] [Google Scholar]
- 27.Mutters R., Pohl S., Mannheim W. Transfer of Pasteurella ureae Jones 1962 to the genus Actinobacillus Brumpt 1910: Actinobacillus ureae comb. nov. International Journal of Systematic Bacteriology. 1986;36(2):343–344. doi: 10.1099/00207713-36-2-343. [DOI] [Google Scholar]
- 28.Christensen H., Kuhnert P. International committee on systematics of prokaryotes. Subcommittee on the taxonomy of Pasteurellaceae. Minutes of the meetings, 25 August 2011, Elsinore, Denmark. International Journal of Systematic and Evolutionary Microbiology. 2012;62(1):257–258. doi: 10.1099/ijs.0.039131-0. [DOI] [PubMed] [Google Scholar]
- 29.Hedegaard J., Okkels H., Bruun B., Kilian M., Mortensen K. K., Nørskov-Lauritsen N. Phylogeny of the genus Haemophilus as determined by comparison of partial infB sequences. Microbiology. 2001;147(9):2599–2609. doi: 10.1099/00221287-147-9-2599. [DOI] [PubMed] [Google Scholar]
- 30.Nørskov-Lauritsen N., Bruun B., Kilian M. Multilocus sequence phylogenetic study of the genus Haemophilus with description of Haemophilus pittmaniae sp. nov. International Journal of Systematic and Evolutionary Microbiology. 2005;55(1):449–456. doi: 10.1099/ijs.0.63325-0. [DOI] [PubMed] [Google Scholar]
- 31.Cattoir V., Lemenand O., Avril J.-L., Gaillot O. The sodA gene as a target for phylogenetic dissection of the genus Haemophilus and accurate identification of human clinical isolates. International Journal of Medical Microbiology. 2006;296(8):531–540. doi: 10.1016/j.ijmm.2006.06.005. [DOI] [PubMed] [Google Scholar]
- 32.Kilian M. Genus III. Haemophilus . In: Brenner D. J., Krieg N. R., Garrity G. M., Staley J. T., editors. Bergey's Manual of Systematic Bacteriology. 2nd. Vol. 2. New York, NY, USA: Springer; 2005. pp. 883–904. [Google Scholar]
- 33.Nørskov-Lauritsen N., Bruun B., Andersen C., Kilian M. Identification of haemolytic Haemophilus species isolated from human clinical specimens and description of Haemophilus sputorum sp. nov. International Journal of Medical Microbiology. 2012;302(2):78–83. doi: 10.1016/j.ijmm.2012.01.001. [DOI] [PubMed] [Google Scholar]
- 34.Fleischmann R. D., Adams M. D., White O., et al. Whole-genome random sequencing and assembly of Haemophilus influenzae Rd. Science. 1995;269(5223):496–512. doi: 10.1126/science.7542800. [DOI] [PubMed] [Google Scholar]
- 35.Wattam A. R., Abraham D., Dalay O., et al. PATRIC, the bacterial bioinformatics database and analysis resource. Nucleic Acids Research. 2014;42(1):D581–D591. doi: 10.1093/nar/gkt1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.NCBI. NCBI Genome Database. 2014. http://www.ncbi.nlm.nih.gov/genome/ [Google Scholar]
- 37.NCBI. NCBI Nucleotide Database, 2014, http://www.ncbi.nlm.nih.gov/nuccore/
- 38.Edgar R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research. 2004;32(5):1792–1797. doi: 10.1093/nar/gkh340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tamura K., Stecher G., Peterson D., Filipski A., Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution. 2013;30(12):2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Foote S. J., Bossé J. T., Bouevitch A. B., Langford P. R., Young N. M., Nash J. H. E. The complete genome sequence of Actinobacillus pleuropneumoniae L20 (serotype 5b) Journal of Bacteriology. 2008;190(4):1495–1496. doi: 10.1128/jb.01845-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Xu Z., Zhou Y., Li L., et al. Genome biology of Actinobacillus pleuropneumoniae JL03, an isolate of serotype 3 prevalent in China. PLoS ONE. 2008;3(1) doi: 10.1371/journal.pone.0001450.e1450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Di Bonaventura M. P., DeSalle R., Pop M., et al. Complete genome sequence of Aggregatibacter (Haemophilus) aphrophilus NJ8700. Journal of Bacteriology. 2009;191(14):4693–4694. doi: 10.1128/jb.00447-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chen C., Kittichotirat W., Si Y., Bumgarner R. Genome sequence of Aggregatibacter actinomycetemcomitans serotype c strain D11S-1. Journal of Bacteriology. 2009;191(23):7378–7379. doi: 10.1128/jb.01203-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Chen C., Kittichotirat W., Chen W., Downey J. S., Si Y., Bumgarner R. Genome sequence of naturally competent Aggregatibacter actinomycetemcomitans serotype a strain D7S-1. Journal of Bacteriology. 2010;192(10):2643–2644. doi: 10.1128/jb.00157-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Johnson T. J., Fernandez-Alarcon C., Bojesen A. M., Nolan L. K., Trampel D. W., Seemann T. Complete genome sequence of Gallibacterium anatis strain UMN179, isolated from a laying hen with peritonitis. Journal of Bacteriology. 2011;193(14):3676–3677. doi: 10.1128/jb.05177-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jordan I. K., Conley A. B., Antonov I. V., et al. Genome sequences for five strains of the emerging pathogen Haemophilus haemolyticus . Journal of Bacteriology. 2011;193(20):5879–5880. doi: 10.1128/jb.05863-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Strouts F. R., Power P., Croucher N. J., et al. Lineage-specific virulence determinants of Haemophilus influenzae biogroup aegyptius . Emerging Infectious Diseases. 2012;18(3):449–457. doi: 10.3201/eid1803.110728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hogg J. S., Hu F. Z., Janto B., et al. Characterization and modeling of the Haemophilus influenzae core and supragenomes based on the complete genomic sequences of Rd and 12 clinical nontypeable strains. Genome Biology. 2007;8(6, article R103) doi: 10.1186/gb-2007-8-6-r103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Harrison A., Dyer D. W., Gillaspy A., et al. Genomic sequence of an otitis media isolate of nontypeable Haemophilus influenzae: comparative study with H. influenzae serotyped, strain KW20. Journal of Bacteriology. 2005;187(13):4627–4636. doi: 10.1128/jb.187.13.4627-4636.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Yue M., Yang F., Yang J., et al. Complete genome sequence of Haemophilus parasuisSH0165. Journal of Bacteriology. 2009;191(4):1359–1360. doi: 10.1128/jb.01682-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Challacombe J. F., Duncan A. J., Brettin T. S., et al. Complete genome sequence of Haemophilus somnus (Histophilus somni) strain 129Pt and comparison to Haemophilus ducreyi 35000HP and Haemophilus influenzae Rd. Journal of Bacteriology. 2007;189(5):1890–1898. doi: 10.1128/jb.01422-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hong S. H., Kim J. S., Lee S. Y., et al. The genome sequence of the capnophilic rumen bacterium Mannheimia succiniciproducens . Nature Biotechnology. 2004;22(10):1275–1281. doi: 10.1038/nbt1010. [DOI] [PubMed] [Google Scholar]
- 53.Gioia J., Qin X., Jiang H., et al. The genome sequence of Mannheimia haemolytica A1: insights into virulence, natural competence, and Pasteurellaceae phylogeny. Journal of Bacteriology. 2006;188(20):7257–7266. doi: 10.1128/jb.00675-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.May B. J., Zhang Q., Li L. L., Paustian M. L., Whittam T. S., Kapur V. Complete genomic sequence of Pasteurella multocida, Pm70. Proceedings of the National Academy of Sciences of the United States of America. 2001;98(6):3460–3465. doi: 10.1073/pnas.051634598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Edgar R. C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics. 2010;26(19):2460–2461. doi: 10.1093/bioinformatics/btq461.btq461 [DOI] [PubMed] [Google Scholar]
- 56.Katoh K., Standley D. M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology and Evolution. 2013;30(4):772–780. doi: 10.1093/molbev/mst010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Price M. N., Dehal P. S., Arkin A. P. FastTree 2—approximately maximum-likelihood trees for large alignments. PLoS ONE. 2010;5(3) doi: 10.1371/journal.pone.0009490.e9490 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Whelan S., Goldman N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Molecular Biology and Evolution. 2001;18(5):691–699. doi: 10.1093/oxfordjournals.molbev.a003851. [DOI] [PubMed] [Google Scholar]
- 59.Gupta R. S. Methods in Microbiology. Vol. 41. Academic Press; 2014. Identification of conserved indels that are useful for classification and evolutionary studies; pp. 153–182. [DOI] [Google Scholar]
- 60.Jeanmougin F., Thompson J. D., Gouy M., Higgins D. G., Gibson T. J. Multiple sequence alignment with Clustal X. Trends in Biochemical Sciences. 1998;23(10):403–405. doi: 10.1016/S0968-0004(98)01285-7. [DOI] [PubMed] [Google Scholar]
- 61.Stackebrandt E., Ebers J. Taxonomic parameters revisited: tarnished gold standards. Microbiology Today. 2006;33(4):p. 152. [Google Scholar]
- 62.Konstantinidis K. T., Stackebrandt E. The Prokaryotes. Springer; 2013. Defining taxonomic ranks; pp. 229–254. [Google Scholar]
- 63.Redfield R. J., Findlay W. A., Bossé J., Kroll J. S., Cameron A. D. S., Nash J. H. E. Evolution of competence and DNA uptake specificity in the Pasteurellaceae . BMC Evolutionary Biology. 2006;6, article 82 doi: 10.1186/1471-2148-6-82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Rokas A., Williams B. I., King N., Carroll S. B. Genome-scale approaches to resolving incongruence in molecular phylogenies. Nature. 2003;425(6960):798–804. doi: 10.1038/nature02053. [DOI] [PubMed] [Google Scholar]
- 65.Wu D., Hugenholtz P., Mavromatis K., et al. A phylogeny-driven genomic encyclopaedia of Bacteria and Archaea. Nature. 2009;462(7276):1056–1060. doi: 10.1038/nature08656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Yilmaz P., Parfrey L. W., Yarza P., et al. The SILVA and 'sll-species living tree project (LTP)' taxonomic frameworks. Nucleic Acids Research. 2013 doi: 10.1093/nar/gkt1209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Gao B., Mohan R., Gupta R. S. Phylogenomics and protein signatures elucidating the evolutionary relationships among the Gammaproteobacteria . International Journal of Systematic and Evolutionary Microbiology. 2009;59(2):234–247. doi: 10.1099/ijs.0.002741-0. [DOI] [PubMed] [Google Scholar]
- 68.Cutiño-Jiménez A. M., Martins-Pinheiro M., Lima W. C., Martín-Tornet A., Morales O. G., Menck C. F. M. Evolutionary placement of Xanthomonadales based on conserved protein signature sequences. Molecular Phylogenetics and Evolution. 2010;54(2):524–534. doi: 10.1016/j.ympev.2009.09.026. [DOI] [PubMed] [Google Scholar]
- 69.Naushad H. S., Gupta R. S. Phylogenomics and molecular signatures for species from the plant pathogen-containing order Xanthomonadales . PLoS ONE. 2013;8(2) doi: 10.1371/journal.pone.0055216.e55216 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Gupta R. S. Applications of conserved indels for understanding microbial phylogeny. In: Oren A., Papke R. T., editors. Molecular Phylogeny of Microorganisms. Norfolk, UK: Caister Academic Press; 2010. pp. 135–150. [Google Scholar]
- 71.Naushad H. S., Lee B., Gupta R. S. Conserved signature indels and signature proteins as novel tools for understanding microbial phylogeny and systematics: Identification of molecular signatures that are specific for the phytopathogenic genera Dickeya, Pectobacterium and Brenneria . International Journal of Systematic and Evolutionary Microbiology. 2014;64(2):366–383. doi: 10.1099/ijs.0.054213-0. [DOI] [PubMed] [Google Scholar]
- 72.Gupta R. S. Protein phylogenies and signature sequences: a reappraisal of evolutionary relationships among archaebacteria, eubacteria, and eukaryotes. Microbiology and Molecular Biology Reviews. 1998;62(4):1435–1491. doi: 10.1128/mmbr.62.4.1435-1491.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Rokas A., Holland P. W. H. Rare genomic changes as a tool for phylogenetics. Trends in Ecology and Evolution. 2000;15(11):454–459. doi: 10.1016/S0169-5347(00)01967-4. [DOI] [PubMed] [Google Scholar]
- 74.Gupta R. S., Lali R. Molecular signatures for the phylum Aquificae and its different clades: Proposal for division of the phylum Aquificae into the emended order Aquificales, containing the families Aquificaceae and Hydrogenothermaceae, and a new order Desulfurobacteriales ord. nov., containing the family Desulfurobacteriaceae . Antonie van Leeuwenhoek. 2013;104(3):349–368. doi: 10.1007/s10482-013-9957-6. [DOI] [PubMed] [Google Scholar]
- 75.Gupta R. S., Mahmood S., Adeolu M. A phylogenomic and molecular signature based approach for characterization of the phylum spirochaetes and its major clades: proposal for a taxonomic revision of the phylum. Frontiers in Microbiology. 2013;4, article 217 doi: 10.3389/fmicb.2013.00217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Adeolu M., Gupta R. S. A phylogenomic and molecular marker based proposal for the division of the genus Borrelia into two genera: the emended genus Borrelia containing only the members of the relapsing fever Borrelia, and the genus Borreliella gen. nov. containing the members of the Lyme disease Borrelia (Borrelia burgdorferi sensu lato complex) Antonie van Leeuwenhoek. 2014;105(6):1049–1072. doi: 10.1007/s10482-014-0164-x. [DOI] [PubMed] [Google Scholar]
- 77.Bhandari V., Gupta R. S. Molecular signatures for the phylum (class) Thermotogae and a proposal for its division into three orders (Thermotogales, Kosmotogales ord. Nov. and Petrotogales ord. Nov.) containing four families (Thermotogaceae, Fervidobacteriaceae fam. Nov., Kosmotogaceae fam. Nov. and Petrotogaceae fam. Nov.) and a new genus Pseudothermotoga gen. Nov. with five new combinations. Antonie van Leeuwenhoek, International Journal of General and Molecular Microbiology. 2014;105(1):143–168. doi: 10.1007/s10482-013-0062-7. [DOI] [PubMed] [Google Scholar]
- 78.Olsen I. Recent approaches to the chemotaxonomy of the Actinobacillus-Haemophilus-Pasteurella group (family Pasteurellaceae) Oral Microbiology and Immunology. 1993;8(6):327–336. doi: 10.1111/j.1399-302x.1993.tb00607.x. [DOI] [PubMed] [Google Scholar]
- 79.Hayashimoto N., Ueno M., Tkakura A., Itoh T. Biochemical characterization and phylogenetic analysis based on 16S rRNA sequences for V-factor dependent members of Pasteurellaceae derived from laboratory rats. Current Microbiology. 2007;54(6):419–423. doi: 10.1007/s00284-006-0531-6. [DOI] [PubMed] [Google Scholar]
- 80.Mutters R., Frederiksen W., Mannheim W. Taxonomy of the group. In: Adlam C., Rutter J. M., editors. Pasturella and Pasteurellosis. London, UK: Academic Press; 1989. pp. 3–34. [Google Scholar]
- 81.Snipes K. P., Biberstein E. L. Pasteurella testudinis sp. nov.: a parasite of desert tortoises (Gopherus agassizi) International Journal of Systematic Bacteriology. 1982;32(2):201–210. doi: 10.1099/00207713-32-2-201. [DOI] [Google Scholar]
- 82.Angen Ø., Ahrens P., Bisgaard M. Phenotypic and genotypic characterization of Mannheimia (Pasteurella) haemolytica-like strains isolated from diseased animals in Denmark. Veterinary Microbiology. 2002;84(1-2):103–114. doi: 10.1016/s0378-1135(01)00439-4. [DOI] [PubMed] [Google Scholar]
- 83.Ahmod N. Z., Gupta R. S., Shah H. N. Identification of a Bacillus anthracis specific indel in the yeaC gene and development of a rapid pyrosequencing assay for distinguishing B. anthracis from the B. cereus group. Journal of Microbiological Methods. 2011;87(3):278–285. doi: 10.1016/j.mimet.2011.08.015. [DOI] [PubMed] [Google Scholar]
- 84.Wong S. Y., Paschos A., Gupta R. S., Schellhorn H. E. Insertion/deletion-based approach for the detection of Escherichia coli O157:H7 in freshwater environments. Environmental Science & Technology. 2014;48(19):11462–11470. doi: 10.1021/es502794h. [DOI] [PubMed] [Google Scholar]
- 85.Gao B., Gupta R. S. Phylogenetic framework and molecular signatures for the main clades of the phylum Actinobacteria . Microbiology and Molecular Biology Reviews. 2012;76(1):66–112. doi: 10.1128/mmbr.05011-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Howard-Azzeh M., Shamseer L., Schellhorn H. E., Gupta R. S. Phylogenetic analysis and molecular signatures defining a monophyletic clade of heterocystous cyanobacteria and identifying its closest relatives. Photosynthesis Research. 2014;122(2):171–185. doi: 10.1007/s11120-014-0020-x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Partial sequence alignments of the CSI containing regions that were unique characteristics of the genera Actinobacillus sensu stricto, Haemophilus sensu stricto, Pasteurella sensu stricto, Aggregatibacter, or Mannheimia.