

Abstract
The Drosophila transcription factor Cabut/dTIEG (Cbt) is a growth regulator, whose expression is modulated by different stimuli. Here, we determine Cbt association with chromatin and identify Yorkie (Yki), the transcriptional co-activator of the Hippo (Hpo) pathway as its partner. Cbt and Yki co-localize on common gene promoters, and the expression of target genes varies according to changes in Cbt levels. Down-regulation of Cbt suppresses the overgrowth phenotypes caused by mutations in expanded (ex) and yki overexpression, whereas its up-regulation promotes cell proliferation. Our results imply that Cbt is a novel partner of Yki that is required as a transcriptional co-activator in growth control.
Keywords: Cabut, dTIEG, GAF, growth, Yorkie
Introduction
Gene expression is regulated through the integrated action of, among others, many cis-regulatory elements, including core promoters and enhancers located at greater distances from transcription start sites (TSS) (reviewed in 1). The combinatorial binding of transcription factors (TF) to these elements can lead to diverse types of transcriptional output, and an understanding of this mechanism is crucial, for example, in the context of development. In fact, the final size and shape of an organism require a complex genetic network of signaling molecules, the final outcome of which must be finely regulated in space and time to ensure a proper response.
The transcription factor Cabut/dTIEG (Cbt) is the fly ortholog of TGF-β-inducible early genes 1 and 2 (TIEG1 and TIEG2) in mammals, which belong to the evolutionary conserved TIEG family 2. TIEGs are zinc finger proteins of the Krüppel-like factor (KLF) family that can function as either activators or repressors depending on the cellular context, the promoter to which they bind or the interacting partners 3. TIEG proteins participate in a wide variety of cellular processes, from development to cancer, and regulate genes that control proliferation, apoptosis, regeneration or differentiation 4, 5.
Drosophila cbt was identified and characterized from an overexpression screen of EP lines conducted to determine genes involved in establishing epithelial planar cell polarity 6, 7. This TF is ubiquitously expressed in the wing disk, and its expression increases in response to metabolic, hormonal and stress signals. Cbt levels rise upon inhibition of TOR signaling 8, 9, and it is among the most highly Mlx-regulated genes 10. Among its functions, it is known that Cbt is required during dorsal closure downstream of JNK signaling 7, that it is a modulator of different signaling pathways involved in wing patterning and proliferation 11 and that it promotes ectopic cell cycling when overexpressed 12. Moreover, Cbt is a crucial downstream mediator gene of the JNK signaling required during wing disk regeneration 5. In spite of this, little is known about its downstream target genes or its precise mechanism of action. Here, we report a novel function for Cbt as a partner of Yki (Yorkie). Yki is the key effector of growth control and the downstream element of the highly conserved Hpo (Hippo) signaling pathway. The Hpo pathway limits organ size by phosphorylating and inhibiting Yki, a key regulator of proliferation and apoptosis. Yki can also act as an oncogene, since it has potent growth-promoting activity (reviewed in 13). Our results show a role for Cbt as a transcriptional activator with the capacity to modulate Yki growth response.
Results and Discussion
Widespread occupancy of Cbt on chromatin
To characterize Cbt target genes, we performed chromatin immunoprecipitation and high-throughput sequencing (ChIP-Seq) from third instar larval wing imaginal disks 14, 15. Analysis of Cbt-bound regions (Supplementary Table S1) in the entire genome revealed that approximately 70% of its peaks were located in proximal promoters or introns (Fig1A and B and Supplementary Fig S1A), consistent with its role as a transcriptional regulator. Thus, we identified 2,060 putative target genes in the wing disk (Supplementary Table S2). Gene Ontology (GO) 16 analysis indicated that this subset of genes was enriched in transcriptional activity, cell migration, mitotic cell cycle and signaling pathways known to play a role in imaginal disk development (Supplementary Fig S1B). As expected, among Cbt targets we found previously described genes such as salm (spalt major) or cbt itself 11, 17, but also several unidentified target genes such as wg (wingless) or vg (vestigial) (Fig1B).
Figure 1.

- A Genome distribution of Cbt peaks (left). Coverage of each class of gene region in the entire genome (right). Cbt peaks are preferentially located in promoter regions (from −1,000 to 100 bp from the TSS).
- B UCSC genome browser screenshot of the Cbt ChIP-Seq profile across regions of known (cbt and salm) and unknown targets (wg and vg). The height of the profile represents the number of reads, and peaks are depicted as black boxes.
- C Ranking of Cbt target genes according to their expression from RNA-Seq in wing imaginal disk (left). Heatmap (right) showing the presence (black) and absence (white) of H3K4me3 and H3K27me3 histone modifications on these genes (data from15).
- D Cbt ChIP analysis of GFP-negative cells (GFP−, black) and GFP-positive cells (GFP+, green) from sorted nub-Gal4; UAS-GFP wing disks. Control samples (mock) of GFP− (gray) and GFP+ (light green). Real-time PCR results were normalized against the mock sample (negative) and are depicted as fold enrichment. Error bars represent the SEM. T-test (*)P ≤ 0.05, n + 3.
- E Comparison of mRNA expression levels of identified Cbt target genes quantified by real-time PCR in wt (wild-type) (black) and ap > cbt (gray) wing disks. Error bars represent SEM. T-test (*)P ≤ 0.05, n + 4.
- F-I (F and H) Wg and Vg (red) staining in wt disks (control). (G and I) Ectopic cbt expression in the medial region of the wing disk using ptc-GAL4 (green) induces up-regulation of Wg (G-G', red) and Vg (I-I', red) in the wing pouch. Scale bar + 50 µm.
Cbt association around the TSS may be an indication of its function as a primary regulatory element, but does not provide any information about its role as an activator or a repressor. To elucidate this question, we examined published data on chromatin modifications 15 as well as recently obtained RNA-Seq data from the wing disk (Pérez-Lluch, in preparation) and ranked Cbt targets according to their expression level (Fig1C). Although at different levels, target genes are mostly expressed in the wing disk. This positive correlation with gene expression was also detected in the extensive overlap between Cbt occupancy and trimethylated histone 3 lysine 4 (H3K4me3). In contrast, only 13% of Cbt target genes correlated with the repressive chromatin mark H3K27me3 (Supplementary Fig S1C–G). Although 200 Cbt targets seemed to present both modifications, these may be coupled to the differential expression pattern of several genes in the wing disk. To clarify whether Cbt binds to active or inactive genes, we next examined Cbt occupancy of genes known to be differentially expressed in a subpopulation of cells within the wing disk tissue. The gene nub (nubbin) is expressed in the wing primordium 18. We induced GFP expression in the wing pouch using a nub-GAL4 driver and performed ChIP assays followed by quantitative PCR (qPCR) in sorted cells (Fig1D and Supplementary Fig S1E). In the vicinity of the TSS of genes expressed in the wing pouch, such as rn (rotund) and nub, we only found Cbt in GFP-positive cells. Cbt was also present in the promoter of cycA (cyclin A), both in GFP-positive and GFP-negative cells, in accordance with its expression throughout the entire wing disk (wing pouch and notum). These observations indicate that Cbt might act as a positive activator of transcription in this tissue. To further confirm this, we examined the expression of selected targets after cbt overexpression. Induction of cbt in the dorsal domain of the wing using an ap-GAL4 (apterous) driver led to a clear increase in the expression levels of target genes, as detected by qPCR (Fig1E). We also ectopically expressed cbt in the ptc (patched) domain of the wing disk using the ptc-GAL4 driver and examined the pattern of Wg (normally restricted to cells adjacent to the D/V boundary in the wing blade and to two rings in the hinge region) and Vg (expressed throughout the wing blade) by immunostaining. After cbt induction, we observed spread staining of Wg in the boundary and ring regions (Fig1F–G'). Likewise, analysis of Vg revealed increased protein levels in the region where cbt was upregulated (1H–I'). No ectopic expression of Wg or Vg was detected in regions far from where they are normally expressed, suggesting that cbt alone is not sufficient to ectopically activate transcription of these genes but can modulate or cooperate with factors that promote their basal expression. Taken together, our results suggest that Cbt functions as a transcriptional activator in the wing disk. Nevertheless, we cannot discard its contribution to repression in some contexts or through binding to different partners, as previous experiments have demonstrated the relevance of the Sin3A interaction domain for Cbt's repressive role 17.
Cbt associates with GAF and Yki proteins
TIEG factors contain three conserved C-terminal zinc finger motifs that seem to bind to GC-rich sequences in vertebrates 19, 20, 21, 22. To characterize the set of motifs enriched within Cbt binding sites, we used different pattern discovery methods (see Materials and Methods, Supplementary Fig S2 and Supplementary Table S3). Among others, we detected GC sequences and the Sp1 motif, as expected for a TIEG family member, but in addition, one of the most enriched motifs comprised GAGA-binding sequences (Fig2A). We did not find enrichment of the proposed consensus TIEG motif 5′GGTGTG3′ 23, which suggests that Cbt binds to degenerated or alternative motifs or may function through its interaction with other TFs. A recent study identified a novel Mad-like motif in promoters of Cbt-regulated genes 12. However, this new motif does not coincide with previously reported Cbt binding data 17, 24.
Figure 2.
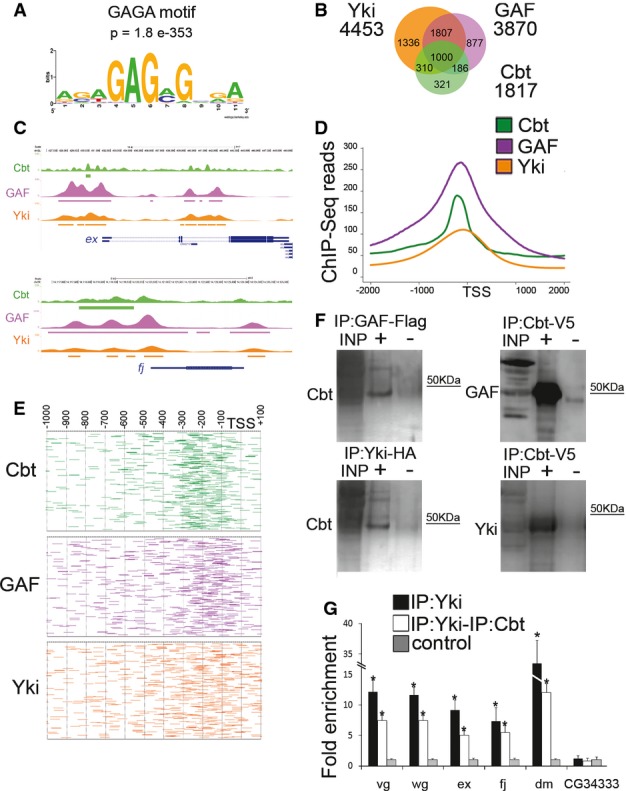
- A Motif and significance score for GAF DNA-binding protein at Cbt-bound targets. De novo analysis with MEME-ChIP identified GAF motif (GAGA) as one of the most representative motifs enriched within Cbt binding regions.
- B Venn diagram showing the overlap between Cbt (green), GAF (purple) and Yki (orange) in the promoters of their target genes.
- C UCSC Genome Browser overview of Cbt (green), GAF (purple) and Yki (orange) ChIP-Seqs in ex and fj regions. Peaks are represented as boxes in different colors: Cbt (green), GAF (purple) and Yki (orange).
- D Distribution of Cbt (green), GAF (purple) and Yki (orange) ChIP-Seq reads over the TSS of common target genes.
- E Distribution of Cbt (green), GAF (purple) and Yki (orange) location in the promoters of their target genes.
- F Western blots showing proteins immunoprecipitated from S2 cells transfected with Yki-HA, GAF-Flag or Cbt-V5 cells and detected by anti-Cbt, anti-GAF and anti-Yki antibodies. Input (INP), immunoprecipitated samples (+) and negative control (−).
- G ChIP-reChIP of Yki-HA and Cbt from S2 cells tested by real-time PCR. The order of antibodies is Yki-HA ChIP and Cbt. Yki-HA ChIP (black), ChIP-reChIP (white). CG34333 promoter region was used as a negative bound region, and results were normalized against input and the mock sample (negative control, gray) and are depicted as fold enrichment. Error bars represent fold enrichment error. T-test (*)P ≤ 0.05, n + 3.
Source data are available online for this figure.
Association of Cbt genome occupancy with regions bound by other TFs expressed in wing disk using reported data 25 showed strong correlations between Cbt, Yki and the general factor GAF (GAGA factor) (Fig2B). More than 70% of Cbt targets were also Yki targets, and of these, around 76% were occupied by the three of them (Fig2B). Fig2C shows the profiles of these factors in known Yki targets such as ex (expanded) and fj (four-jointed). Mapping the targets on common gene promoters revealed that most of Cbt and GAF were located mainly at the promoter region close to the TSS (Fig2E). Although this distribution was more scattered in the case of Yki, the majority of targets were observed in the same region. Projection of the mean reads over the TSS of the full set of genes confirmed this observation (Fig2D). Analysis of Cbt and GAF in polytene chromosomes confirmed their co-localization in particular bands (Supplementary Fig S3A and B). The interaction of Cbt with Yki and GAF proteins was confirmed by co-immunoprecipitation in S2 cells. Western blotting revealed that Cbt specifically co-precipitate with Yki and GAF (Fig2F and Supplementary Fig S3C). To finally verify co-binding of Cbt and Yki in specific targets, we used the ChIP-reChIP technique and found that both factors bind in close vicinity on the same regions (Fig2G), which confirms they physically interact. Altogether, our data suggest that Cbt, GAF and Yki act together to regulate gene transcription. Whether this interaction is direct or mediated by other proteins remains to be elucidated.
Close inspection of members of the Hpo pathway showed that several upstream and downstream components were Cbt targets (Fig3A). To further investigate the requirement of Cbt in controlling Yki target genes expression, we examined the levels of fj, DIAP1 (Drosophila inhibitor of apoptosis) and ex under conditions of elevated or reduced expression of cbt. fj, DIAP1 and ex reporters exhibited higher levels in the cbt overexpression domains (Fig3B–G'), whereas loss-of-function clones showed reduced DIAP1 expression (Fig3H) and fj levels decreased in the presence of cbt RNAi (Supplementary Fig S3E). As expected, and consistent with the occupancy profile, no changes were observed in the expression of bantam, a component of the Yki pathway not bound by Cbt (Supplementary Fig S3G).
Figure 3.
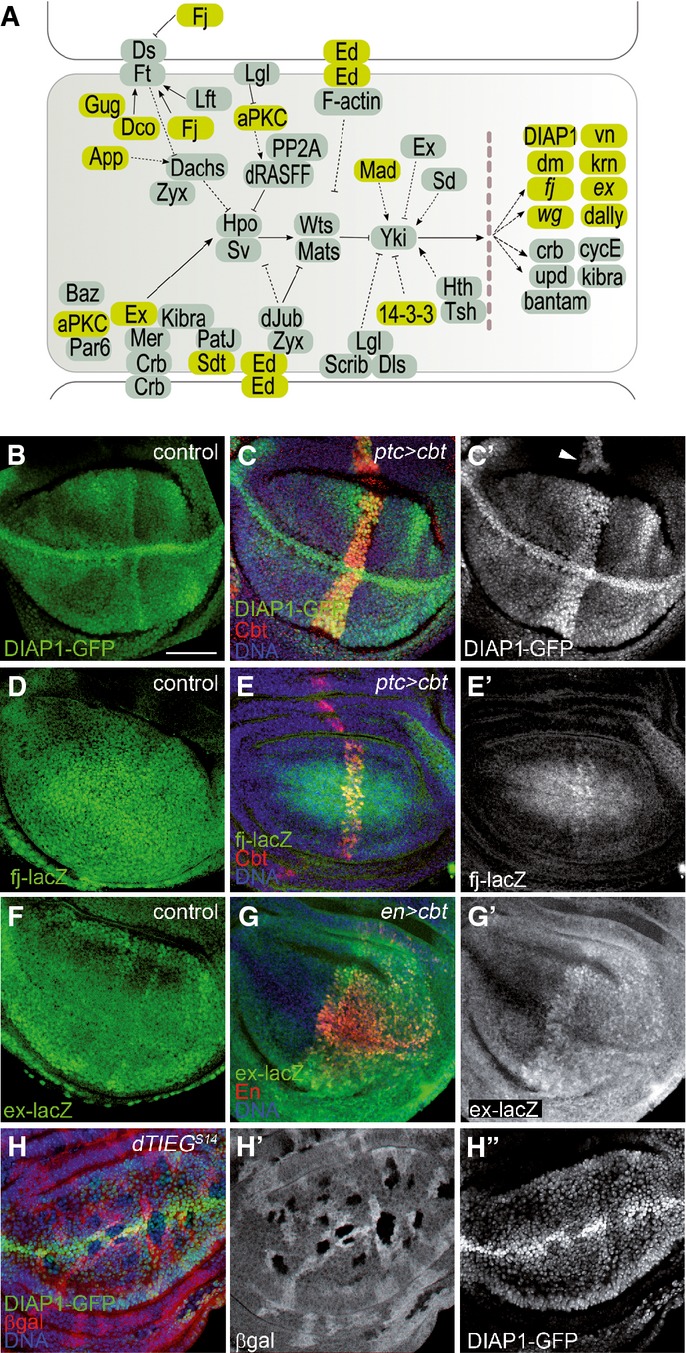
- A Representation of the Hpo pathway and Yki target genes (according to the Kegg pathway 36). Genes identified as Cbt targets that belong to the pathway are highlighted in green.
- B-G Expression pattern of DIAP1, fj and ex reporters after overexpressing cbt (red) in wing disks. Scale bar + 50 µm. (B–C') DIAP1-GFP expression in wt (control, B) and ptc>cbt disks (C–C'). (D–E') fj-lacZ expression (green) in wt (control, D) and ptc > cbt disks (E–E'). (F–G') ex-lacZ expression in wt (control, F) and en > cbt disks (G–G').
- H Analysis of DIAP1-GFP (green) in cbt mutant clones (dTIEGS14, black).
Many studies have emphasized the complexity of Yki and its mammalian homologs YAP and TAZ regulation, including multiple combinations with associate proteins in distinct target genes (reviewed in 13). Besides DNA-binding partners such Sd (Scalloped) and Hth (Homothorax) in Drosophila 26, 27, Yki can cooperate with other factors directly on target promoters, such as the cell cycle-related gene dE2F1 28. Remarkably, a recent report shows that Cbt and dE2F1 regulate an overlapping set of cell cycle genes 12. In the Dpp pathway, Mad (Mothers against decapentaplegic) and Yki interact to form a transcription complex to activate their common targets 29. This association is conserved through evolution, as YAP and TAZ interact with Smad proteins to potentiate transcriptional activity 30. Recent studies have also identified Mask (Multiple ankyrin repeats single KH domain) as a novel cofactor for Yki/YAP, required to induce target gene expression 31, 32. Our results highlight the role of Cbt as a new Yki partner involved in the activation of some Yki target gene expression. This function of Cbt may occur in part through association with GAF as well as chromatin remodeler complexes 25.
Cbt modulates Yki activity
Since overexpression of cbt results in an increase in proliferation as well as wing size 11 (Supplementary Fig S4), we hypothesized that Cbt's role in size control could be mediated through its association with Yki. To address this question, we depleted cbt levels and analyzed the effect on the growth of ex mutant clones and in clones overexpressing yki in wing and eye-antenna imaginal disks. The Yki target gene ex acts as an upstream positive modulator of the Hpo pathway, and in accordance with its role as a tumor suppressor, its loss-of-function mutation results in large clones (Figs4B and 5B) 33. Expression of cbt RNAi in this mutant background caused a clear reduction in the clone size (Figs4D, 5D and Supplementary Fig S5). In the same direction, the overgrowth known to occur by overexpression of a yki-activated form (Figs4H and 5H) 34 is prevented in a mutant cbt background (Figs4I and 5I) as well as expressing cbt RNAi (Fig4S and Supplementary Fig S5). Moreover, impaired growth caused by yki depletion could not be rescued increasing cbt levels (Figs4K–O and 5K–O) and overexpression of yki and cbt triggered massive growth in imaginal tissues (Supplementary Fig S5). Finally, depletion of cbt in adult organs (wings and eyes) also reduced Yki-mediated overgrowth (Figs4P–T and 5P–T), indicating a general function for Cbt in the regulation of Hippo pathway-mediated tissue growth.
Figure 4.

- A-E Wing disks containing GFP-marked MARCM clones of the exe1 mutant allele (B), cbt RNAi (C) and exe1 mutant clones with cbt RNAi (D). Scale bar + 50 µm. (E) Quantification of clone area of control (black), ex mutant (gray), cbt RNAi (white) and ex e1 clones with cbt RNAi (green). Error bars represent SEM. T-test (*)P ≤ 0.003, n + 10.
- F-J Wing disks containing GFP-marked MARCM clones of the dTIEGS14 mutant allele (G), YkiS168A overexpression (H) and dTIEGS14 mutant clones with ykiS168A overexpression (I). (J) Quantification of clone area of control (black), cbt mutant (gray), yki overexpression (white) and cbt mutant with yki overexpression (green). Error bars represent SEM. T-test (*)P ≤ 0.0003, n + 10.
- K-O Wing disks containing GFP-marked MARCM clones of the ykiB5 mutant allele (L), cbt overexpression (M) and ykiB5 mutant clones with cbt/dTIEG overexpression (N). (L) Quantification of clone area of control (black), yki mutant (gray), cbt overexpression (white) and yki mutant with cbt overexpression (green). Error bars represent SEM. T-test (*)P ≤ 0.004, n + 10.
- P-T Wings expressing cbt RNAi (R), ykiS168A (Q) and ykiS168A with cbt RNAi (S) under spaltE/Pv promoter for 24 h. Scale bar + 0.5 mm. (T) Quantification of wing area of control (black) cbt RNAi (gray), ykiS168A (white) and cbt RNAi with yki (green). Error bars represent SD. T-test (*)P ≤ 0.00001, n + 50.
Figure 5.

- A-E Eye disks containing GFP-marked MARCM clones of the exe1 mutant allele (B), cbt RNAi (C) and exe1 mutant clones with cbt RNAi (D). Scale bar + 50 µm. (E) Quantification of clone area of control (black), ex mutant (gray), cbt RNAi (white) and ex mutant with cbt RNAi (green). Error bars represent SEM. T-test (*) P ≤ 0.05, n + 10.
- F-J Eye disks containing GFP-marked MARCM clones of the dTIEGS14 mutant allele (G), YkiS168A overexpression (H) and dTIEGS14 mutant clones with ykiS168A overexpression (I). (J) Quantification of clone area of control (black), cbt mutant (gray), yki overexpression (white) and cbt mutant with yki overexpression (green). Error bars represent SEM. T-test (*) P ≤ 0.03, n + 10.
- K-O Eye disks containing GFP-marked MARCM clones of the ykiB5 mutant allele (L), cbt overexpression (M) and ykiB5 mutant clones with cbt overexpression (N). (L) Quantification of clone area of control (black), yki mutant (gray), cbt overexpression (white) and yki mutant with cbt overexpression (green). Error bars represent SEM. T-test (*) P ≤ 0.0001, n + 10
- P-T Eyes containing clones expressing cbt RNAi (R), ykiS168A (Q) and ykiS168A with cbt RNAi (S). Scale bar + 400 µm. (T) Quantification of the percentage occupied for eye clone area from the whole eye, of control (black) cbt RNAi (gray), ykiS168A (white) and cbt RNAi with yki (green). Error bars represent SEM. T-test (*) P ≤ 0.02, n + 10.
In addition to its role during development, it has been shown that Cbt expression is highly regulated by stress and metabolic conditions 9, 10. Cbt has also been identified as a JNK-inducible gene during dorsal closure 7, and we have shown that JNK and tissue damage trigger cbt transient overexpression to promote wing disk regeneration, indicating that its levels must be finely controlled during regenerative growth 5. Moreover, cbt heterozygous mutant disks fail to proliferate and do not regenerate 5, and it is known that during regeneration, the JNK pathway triggers Yki translocation to the nucleus to promote the proliferative response 35. Altogether, our data support a model for Cbt acting as a modulator of Yki activity in the transcriptional regulatory mechanisms that control tissue growth.
Materials and Methods
ChIP-Seq
Cbt ChIP-Seq from wing imaginal disks was performed using a specific antibody against Cbt 14. One thousand four-hundred disks from Canton S third instar larva were pooled and used as a source of chromatin as described 15. IP buffer and 2 μl of Cbt/dTIEG antibody were used for immunoprecipitation. Immunoprecipitated and input samples were processed and sequenced following Solexa/Illumina protocols at the Ultrasequencing Unit of the CRG (Barcelona, Spain). 8 ng of each sample was used, and fragments between 300 and 350 bp were size-selected before sequencing. ChIP-Seq profiles and target regions were deposited in NCBI GEO under the accession number GSE40958.
Cell sorting and real-time PCR
Five hundred dissected disks were dissociated after incubation in trypsin solution for 1 h. Cells were collected in Schneider medium with DAPI and sorted by a cytometer. Recovered cells were then processed for chromatin immunoprecipitation as previously described 15. IP buffer and 2 μl of Cbt antibody were used. Real-time PCRs were normalized against the mock (negative) sample and depicted as fold enrichment.
Acknowledgments
We thank N. Tapon for stocks, plasmids and insightful suggestions, J. Bernués for plasmids and GAF antibody, KD. Irvine for Yki antibody, S. Pérez-Lluch for ChIP-Seq and cell sorting support and M. Milán and I. Rodríguez for stocks. We also thank the Confocal Unit of the CCiTUB (Universitat de Barcelona) and M. Bosch for help with image analysis. M.R.R. was supported by a FPI fellowship, and this project was funded by Grants BFU2009-09781, CSD2007-00008 and BFU2012-36888, Ministerio de Economía y Competitividad, Spain.
Author contributions
MR-R, FS and MC designed the project and analyzed results; MR-R performed the experiments; EB performed computational analysis; NP provided materials, reagents and insightful discussions; and MR-R and MC wrote the manuscript.
Conflict of interest
The authors declare that they have no conflict of interest.
Supporting Information
Supplementary Information
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Source Data For Supplementary Figure S3C
Review Process File
Source Data For Figure 2F
References
- Lenhard B, Sandelin A, Carninci P. Metazoan promoters: emerging characteristics and insights into transcriptional regulation. Nat Rev Genet. 2012;13:233–245. doi: 10.1038/nrg3163. [DOI] [PubMed] [Google Scholar]
- Munoz-Descalzo S, Belacortu Y, Paricio N. Identification and analysis of cabut orthologs in invertebrates and vertebrates. Dev Genes Evol. 2007;217:289–298. doi: 10.1007/s00427-007-0144-5. [DOI] [PubMed] [Google Scholar]
- Kim J, Shin S, Subramaniam M, Bruinsma E, Kim TD, Hawse JR, Spelsberg TC, Janknecht R. Histone demethylase JARID1B/KDM5B is a corepressor of TIEG1/KLF10. Biochem Biophys Res Commun. 2010;401:412–416. doi: 10.1016/j.bbrc.2010.09.068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramaniam M, Hawse JR, Johnsen SA, Spelsberg TC. Role of TIEG1 in biological processes and disease states. J Cell Biochem. 2007;102:539–548. doi: 10.1002/jcb.21492. [DOI] [PubMed] [Google Scholar]
- Blanco E, Ruiz-Romero M, Beltran S, Bosch M, Punset A, Serras F, Corominas M. Gene expression following induction of regeneration in Drosophila wing imaginal discs. Expression profile of regenerating wing discs. BMC Dev Biol. 2010;10:94. doi: 10.1186/1471-213X-10-94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rorth P, Szabo K, Bailey A, Laverty T, Rehm J, Rubin GM, Weigmann K, Milan M, Benes V, Ansorge W, et al. Systematic gain-of-function genetics in Drosophila. Development. 1998;125:1049–1057. doi: 10.1242/dev.125.6.1049. [DOI] [PubMed] [Google Scholar]
- Munoz-Descalzo S, Terol J, Paricio N. Cabut, a C2H2 zinc finger transcription factor, is required during Drosophila dorsal closure downstream of JNK signaling. Dev Biol. 2005;287:168–179. doi: 10.1016/j.ydbio.2005.08.048. [DOI] [PubMed] [Google Scholar]
- Guertin DA, Guntur KV, Bell GW, Thoreen CC, Sabatini DM. Functional genomics identifies TOR-regulated genes that control growth and division. Curr Biol. 2006;16:958–970. doi: 10.1016/j.cub.2006.03.084. [DOI] [PubMed] [Google Scholar]
- Bulow MH, Aebersold R, Pankratz MJ, Junger MA. The Drosophila FoxA ortholog Fork head regulates growth and gene expression downstream of Target of rapamycin. PLoS ONE. 2010;5:e15171. doi: 10.1371/journal.pone.0015171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Havula E, Teesalu M, Hyötyläinen T, Seppälä H, Hasygar K, Auvinen P, Orešič M, Sandmann T, Hietakangas V. Mondo/ChREBP-Mlx-regulated transcriptional network is essential for dietary sugar tolerance in Drosophila. PLoS Genet. 2013;9:e1003438. doi: 10.1371/journal.pgen.1003438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez I. Drosophila TIEG is a modulator of different signalling pathways involved in wing patterning and cell proliferation. PLoS ONE. 2011;6:e18418. doi: 10.1371/journal.pone.0018418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song M, Zhang Y, Katzaroff AJ, Edgar BA, Buttitta L. Hunting complex differential gene interaction patterns across molecular contexts. Nucleic Acids Res. 2014;42:e57. doi: 10.1093/nar/gku086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh H, Irvine KD. Yorkie: the final destination of Hippo signaling. Trends Cell Biol. 2010;20:410–417. doi: 10.1016/j.tcb.2010.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belacortu Y, Weiss R, Kadener S, Paricio N. Expression of Drosophila Cabut during early embryogenesis, dorsal closure and nervous system development. Gene Expr Patterns. 2011;11:190–201. doi: 10.1016/j.gep.2010.11.004. [DOI] [PubMed] [Google Scholar]
- Perez-Lluch S, Blanco E, Carbonell A, Raha D, Snyder M, Serras F, Corominas M. Genome-wide chromatin occupancy analysis reveals a role for ASH2 in transcriptional pausing. Nucleic Acids Res. 2011;39:4628–4639. doi: 10.1093/nar/gkq1322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet. 2000;25:25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belacortu Y, Weiss R, Kadener S, Paricio N. Transcriptional activity and nuclear localization of Cabut, the Drosophila ortholog of vertebrate TGF-beta-inducible early-response gene (TIEG) proteins. PLoS ONE. 2012;7:e32004. doi: 10.1371/journal.pone.0032004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng M, Diaz-Benjumea FJ, Vincent JP, Wu J, Cohen SM. Specification of the wing by localized expression of wingless protein. Nature. 1996;381:316–318. doi: 10.1038/381316a0. [DOI] [PubMed] [Google Scholar]
- Cook T, Gebelein B, Mesa K, Mladek A, Urrutia R. Molecular cloning and characterization of TIEG2 reveals a new subfamily of transforming growth factor-beta-inducible Sp1-like zinc finger-encoding genes involved in the regulation of cell growth. J Biol Chem. 1998;273:25929–25936. doi: 10.1074/jbc.273.40.25929. [DOI] [PubMed] [Google Scholar]
- Kaczynski J, Cook T, Urrutia R. Sp1- and Krüppel-like transcription factors. Genome Biol. 2003;4:206. doi: 10.1186/gb-2003-4-2-206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spittau B, Krieglstein K. Klf10 and Klf11 as mediators of TGF-beta superfamily signaling. Cell Tissue Res. 2012;347:65–72. doi: 10.1007/s00441-011-1186-6. [DOI] [PubMed] [Google Scholar]
- Subramaniam M, Harris SA, Oursler MJ, Rasmussen K, Riggs BL, Spelsberg TC. Identification of a novel TGF-beta-regulated gene encoding a putative zinc finger protein in human osteoblasts. Nucleic Acids Res. 1995;23:4907–4912. doi: 10.1093/nar/23.23.4907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chrisman HR, Tindall DJ. Identification and characterization of a consensus DNA binding element for the zinc finger transcription factor TIEG/EGRalpha. DNA Cell Biol. 2003;22:187–199. doi: 10.1089/104454903321655819. [DOI] [PubMed] [Google Scholar]
- Brown JL, Grau DJ, DeVido SK, Kassis JA. An Sp1/KLF binding site is important for the activity of a Polycomb group response element from the Drosophila engrailed gene. Nucleic Acids Res. 2005;33:5181–5189. doi: 10.1093/nar/gki827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh H, Slattery M, Ma L, Crofts A, White KP, Mann RS, Irvine KD. Genome-wide association of Yorkie with chromatin and chromatin-remodeling complexes. Cell Rep. 2013;3:309–318. doi: 10.1016/j.celrep.2013.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goulev Y, Fauny JD, Gonzalez-Marti B, Flagiello D, Silber J, Zider A. SCALLOPED Interacts with YORKIE, the Nuclear Effector of the Hippo Tumor-Suppressor Pathway in Drosophila. Curr Biol. 2008;18:435–441. doi: 10.1016/j.cub.2008.02.034. [DOI] [PubMed] [Google Scholar]
- Peng HW, Slattery M, Mann RS. Transcription factor choice in the Hippo signaling pathway: homothorax and yorkie regulation of the microRNA bantam in the progenitor domain of the Drosophila eye imaginal disc. Genes Dev. 2009;23:2307–2319. doi: 10.1101/gad.1820009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nicolay BN, Bayarmagnai B, Islam ABMMK, Lopez-Bigas N, Frolov MV. Cooperation between dE2F1 and Yki/Sd defines a distinct transcriptional program necessary to bypass cell cycle exit. Genes Dev. 2011;25:323–335. doi: 10.1101/gad.1999211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh H, Irvine KD. Cooperative Regulation of Growth by Yorkie and Mad through bantam. Dev Cell. 2011;20:109–122. doi: 10.1016/j.devcel.2010.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrigno O, Lallemand F, Verrecchia F, L'Hoste S, Camonis J, Atfi A, Mauviel A. Yes-associated protein (YAP65) interacts with Smad7 and potentiates its inhibitory activity against TGF-beta/Smad signaling. Oncogene. 2002;21:4879–4884. doi: 10.1038/sj.onc.1205623. [DOI] [PubMed] [Google Scholar]
- Sansores-Garcia L, Atkins M, Moya IM, Shahmoradgoli M, Tao C, Mills GB, Halder G. Mask is required for the activity of the hippo pathway effector Yki/YAP. Curr Biol. 2013;23:229–235. doi: 10.1016/j.cub.2012.12.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sidor CM, Brain R, Thompson BJ. Mask proteins are cofactors of Yorkie/YAP in the hippo pathway. Curr Biol. 2013;23:223–228. doi: 10.1016/j.cub.2012.11.061. [DOI] [PubMed] [Google Scholar]
- Hamaratoglu F, Willecke M, Kango-Singh M, Nolo R, Hyun E, Tao C, Jafar-Nejad H, Halder G. The tumour-suppressor genes NF2/Merlin and Expanded act through Hippo signalling to regulate cell proliferation and apoptosis. Nat Cell Biol. 2006;8:27–36. doi: 10.1038/ncb1339. [DOI] [PubMed] [Google Scholar]
- Oh H, Irvine KD. In vivo regulation of Yorkie phosphorylation and localization. Development. 2008;135:1081–1088. doi: 10.1242/dev.015255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun G, Irvine KD. Regulation of Hippo signaling by Jun kinase signaling during compensatory cell proliferation and regeneration, and in neoplastic tumors. Dev Biol. 2011;350:139–151. doi: 10.1016/j.ydbio.2010.11.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanehisa M, Goto S. KEGG: kyoto encyclopaedia of genes and genomes. Nucleic Acids Res. 2000;28:27–30. doi: 10.1093/nar/28.1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Information
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Source Data For Supplementary Figure S3C
Review Process File
Source Data For Figure 2F